


ISSN: 0973-7510
E-ISSN: 2581-690X
Enterobacteriaceae pathogens such as Escherichia coli, Salmonella sp., Shigella sp., Proteus sp., and Klebsiella pneumoniae cause a wide range of gastrointestinal and other mucosal infections. These bacteria acquire antibiotic resistance very quickly and evolve into multi-drug resistant strains thereby making the treatment very difficult. The outer membrane proteins (OMPs) in Enterobacteriaceae are potential vaccine candidates owing for their high immunogenicity and amino acid conservation. The OmpA is one such protein which need to be investigated for the development of a potential subunit vaccine against multiple infections casued by the pathogens of Enterobacteriaceae. To investigate this, we expressed and purified the highly conserved OmpA of S. typhimurium and studied the antibody mediated cross reactivity with the other Enterobacteriaceae pathogens. This was validated through dot ELISA performed with the hyperimmune sera raised against rOmpA of S. typhimurium. We further analyzed the sequence of OmpA protein and clearly understood that the B-cell epitopes in the protein are highly conserved are responsible for cross reactivity among the Enterobacteriaceae pathogens. This work led to findings that provide strong evidence for the application of OmpA in broad-spectrum subunit vaccine against enteric infections.
Enterobacteriaceae, OMPs, OmpA, Antibodies, B-cell Epitopes, Cross Reactivity
Enterobacteriaceae pathogens namely E. coli, K. pneumoniae, and members of the genus Salmonella, Shigella, Proteus cause serious gastrointestinal infections, bacteremia, and other mucosal infections.1-5 The E. coli, Salmonella and Shigella are also major foodborne pathogens transmitted by fecal-oral route. Primarily, these pathogens cause foodborne outbreaks globally by contaminating food and drinking water.1-5 The E. coli is an opportunistic pathogen and has evolved into several serotypes namely enteropathogenic E. coli (EPEC), enteroaggregative E. coli (EAEC), enteroinvasive E. coli (EIEC), enterohemorrhagic E. coli (EHEC), enterotoxigenic E. coli (ETEC) and diffuse adherent E. coli (DAEC).6 These pathogenic forms of E. coli cause serious gastrointestinal infections, sepsis, urinary tract infections (UTIs), meningitis in children and adults with poor immunity.7 Shigella is phylogenetically a very close relative of E. coli in Enterobacteriaceae family, and it is associated with severe gastrointestinal infections even in low CFU count.8,9 Shigella includes four species i.e., S. boydii, S. dysenteriae, S. flexneri and S. sonnei. These bacteria cause an acute intestinal infection called shigellosis.10
Salmonella is another prominent pathogenic member of Enterobacteriaceae that causes diarrhea, typhoid, paratyphoid, salmonellosis, and gastroenteritis etc.11 Members of the genus Proteus like P. mirabilis and P. vulgaris cause UTIs and asymptomatic bacteriuria.12,13 Apart from these pathogens, K. pneumoniae is another important pathogen of Enterobacteriaceae which is also an opportunistic pathogen that is responsible for nosocomial respiratory and urinary tract infections.14-16 Also, K. pneumoniae is the second most frequent bacteria to cause UTIs and bacteremia.14-16
Treatment for these infections has remained a major concern because of their ability to evolve as multi-drug resistant strains over time.17 In this scenario, vaccination and passive immune therapy are the promising alternatives to control widespread infections by these enteric pathogens.
For the development of any immuno-prophylactic agent, selection of the right virulence factor as an antigen is very essential. Previous reports from our lab and others suggest that outer membrane proteins (OMPs) of Enterobacteriaceae as strong immunogenic components in vaccine and passive immune therapies.18-21 The OMPs comprise of β-barrel structures that are essential in maintaining membrane integrity and selective permeability in gram negative bacteria including Enterobacteriaceae.22-24 These OMPs are highly immunogenic and are important virulent factors during infection. Furthermore, studies have shown that the OMPs in Enterobacteriaceae are highly conserved with greater amino acid sequence homology which will vouch for cross-protection against wide range of Enterobacteriaceae infections.18-21 The OmpA is such a strong immunogen which elicited strong immune response against Salmonella and E. coli in mouse models.25,26 However, there is lack of information on the nature of epitope conservation and antibody cross reactivity against this antigen among the pathogenic Enterobacteriaceae members. Therefore, our study will really help the current vaccinologists in developing a broad-spectrum epitope-based vaccine against Enterobacteriaceae pathogens simultaneously.
In the present study, we expressed the highly conserved OmpA protein of S. typhimurium and carried out several experiments to understand the cross reactivity by polyclonal antibodies against OmpA of the standard strains used in the study. The antibodies generated against the OmpA of S. typhimurium has shown cross reactiivty against the other Enterobacteriaceae pathogens such as E. coli, Salmonella, Shigella, Proteus and K. pneumoniae. Sequence and immuninformatics analysis of OmpA has revealed the presence of several conserved B-cell epitopes that are responsible for the cross reactivity by polyclonal antibodies.
Bacterial strains, plasmids, and growth conditions
All the standard cultures used in this study were enlisted in Table 1. The bacteria were cultured overnight in Luria Bertani (LB) broth at 37°C. Dehydrated growth media and antibiotics were from HiMedia labs, Mumbai, India. Isopropy-β-D-thiogalactopyranoside (IPTG) was purchased from Thermo Scientific, India. The S. typhimurium OmpA was cloned into pET22b vector between EcoRI and HindIII restriction sites using GeneRunner software and outsourced from Eurofins, Bengaluru, India. The DH5a and BL21 (DE3) pLys strains of E. coli were used as cloning and expression hosts to express rOmpA of S. typhimurium. These strains were purchased from Invitrogen, USA. Anti-6X histidine mouse monoclonal antibodies and the HRP-conjugated anti-mouse rabbit secondary antibodies were from Sigma Aldrich, India.
Transformation, expression, and purification
The recombinant OmpA gene from S. typhimurium ATCC 14028 was cloned in pET22b vector and sequenced at Eurofins Analytical Services, Bengaluru, India. The recombinant plasmid was transformed into chemically competent E. coli DH5a host cells by heat shock at 42°C. The transformants were screened by colony PCR using T7 promoter and terminator primers with appropriate controls as reported elsewhere.27,28 One of positive colonies was sub cultured and the plasmid DNA was isolated from the transformed cells for further transformation into E. coli BL21 (DE3) pLys expression host cells. The overnight culture was inoculated into 200 mL of LB broth (tryptone=2g; yeast extract=1g; NaCl=2g; pH=7.0) containing 50 µg/mL of ampicillin and 35 µg/mL of chloramphenicol and grown at 37°C overnight. After reaching desired O.D600 value of ~0.6, the rOmpA gene was induced with 1mM of isopropyl β-D-galactopyranoside (IPTG) and cultured for another 5 hrs for the efficient protein expression. After this, the cells were harvested, subjected to osmotic shock and the periplasmic extracts were purified using imidazole and Ni-NTA metal affinity chromatography. The eluted protein extracts were pooled and subjected to buffer exchange in 1X sterile PBS (NaCl=137 mM; KCl=2.7 mM; Na2HPO4=4.3 mM; KH2PO4=1.47 mM; pH=7.4). The purified rOmpA protein was lyophilized, diluted with 1 X sterile PBS, and quantified by Lowry’s assay with BSA standards.29
Immunization
Five-week-old female BALB/c mice were caged in polypropylene cages with rice hulls as bedding material. The mice were provided with food pellets, mineral water from Kent water purifier and maintained in 12-hr light and dark cycles. Fifty micrograms of purified rOmpA were emulsified with Freund’s complete adjuvant and administered sub-cutaneous during the first immunization. Similarly, 50 μg of rOmpA antigen emulsified with Freund’s incomplete adjuvant was administered as booster dose during the second and third immunizations on 14th and 28th day post first immunization. Forty-five days post first immunization, the mice were sacrificed, and blood was harvested by cardiac puncture for serum isolation. After this, Antibody cross reactivity was studied against the Enterobacteriaceae strains through dot ELISA.
Dot ELISA
The standard Enterobacteriaceae cultures enlisted in Table 1 were cultured overnight and immediately used for determining the polyclonal antibody cross reactivity using dot ELISA. Briefly, cells were harvested from the 500 µL of overnight culture and resuspended in carbonate-bicarbonate buffer (pH 9.2). Then 10 µL of the cell suspension was coated on to nitrocellulose strips and airdried followed by blocking in 5% milk solution for 1 hour at room temperature. The strips were washed with PBST solution 4-5 times and incubated in anti-OmpA polyclonal antibody serum. The unbound antibodies were washed, and strips were further incubated with anti-mouse HRP conjugate secondary antibody, washed, and developed with diaminobenzidine (DAB) tetrahydrochloride solution and 0.003% H2O2.
Table (1):
List of Enterobacteriaceae strains used in the study
Organism/strain |
Strain |
|---|---|
Salmonella typhimurium |
ATCC 14028 |
Salmonella paratyphi |
ATCC 9150 |
Shigella flexneri |
ATCC 9199 |
Shigella boydii |
ATCC 9207 |
Klebsiella pneumoniae |
ATCC 10031 |
Proteus vulgaris |
ATCC 13315 |
Escherichia coli |
ATCC 10536 |
Salmonella paratyphi |
MTCC 735 |
Salmonella typhi |
Gwalior isolate |
Salmonella enteritidis |
Gwalior isolate |
Sequence retrieval and alignment
The complete amino acid sequence of OmpA protein of S. typhimurium LT12 strain bearing an accession ID of P02936 was retrieved from Uniprot resource (https://www.uniprot.org/uniprotkb/P02936/entry). Later this sequence was used to mine the complete amino acid sequences of OmpA from other Enterobacteriaceae such as E. coli, Shigella sp., K. pneumoniae and Proteus sp. (Table 2). Multiple sequence alignment (MSA) was performed for these sequences using CLUSTAL Omega tool (https://www.ebi.ac.uk/Tools/msa/clustalo/).
B-cell epitopes
Initially B-cell epitopes were predicted in the core peptide region (21-350 aa) of OmpA of S. typhimurium. Then amino acid sequence conservation in the predicted B-cell epitopes of S. typhimurium OmpA was identified in sequence alignment file containing OmpA sequences of other Enterobacteriaceae pathogens. For the prediction of B-cell epitopes in OmpA of S. typhimurium, we used BepiPred 2.0 (https://services.healthtech.dtu.dk/services/BepiPred-2.0/) tool30 with default parameters.
Generation and purification of rOmpA
The IPTG induction of E. coli BL21 (DE3) pLys cells carrying rOmpA-pET22b plasmid resulted in expression of a ~ 38-kDa protein which was further purified from the periplasmic extracts through Ni-NTA chromatography (Figure 1a). Then after the pooled eluents were buffer exchanged and purified under native conditions to retain the protein structure for its efficient biological activity in vivo. The protein concentration was found to be ~ 1mg/mL which was determined by Lowry’s method. The rOmpA protein of S. typhimurium also contains 6X His tag at the C-terminal end of the protein. Using the mouse monoclonal antibodies raised against the 6X-His tag, the rOmpA protein was detected through western blot (Figure 1b). Reactivity of the 6X-His antibodies was observed at the vicinity of 35-kDa band in western blot.
Cross reactivity by polyclonal antibodies
Upon confirming rOmpA through Western blot followed by the successful immunization of rOmpA protein, the mice were sacrificed once the antibody titer reached 1:64000 after the third booster dose. The high antibody titer indicates strong immunogenic potential of OmpA protein. The harvested serum was used for dot ELISA experiments to determine cross reactivity of anti-OmpA polyclonal antibodies against other Enterobacteriaceae pathogens. Dot ELISA results showed the cross reactivity of anti-OmpA antibodies against other Enterobacteriaceae strains such as E. coli, Shigella sp., Proteus sp., K. pneumoniae and other Salmonella serovars (Figure 1c). As we expected through our in-silico analysis, high degree of amino acid sequence conservation led to the presence of several conserved B-cell epitopes (Figure 2 and 3) which can mediate the cross reactivity by anti-S. typhimurium OmpA polyclonal antibodies towards overnight cultures of standard Enterobacteriaceae cultures we used in this study.
Amino acid sequence conservation in OmpA
We performed MSA analysis for all the retrieved amino acid sequences of OmpA from the Enterobacteriaceae pathogens that are included in the study. The percent identity matrix of OmpA protein among different pathogens was represented in Table 2. The OmpA protein is highly conserved in the monophyletic group of Enterobacteriaceae such as E. coli, Shigella, Salmonella and K. pneumoniae sp. (Table 2). Moreover, considerable amount of amino acid sequence similarity is seen in P. mirabilis and P. vulgaris too which are somewhat distinct from E. coli, Shigella, Salmonella and K. pneumoniae (Table 2). Overall, almost ~ 66% of the amino acid residues in OmpA protein are highly conserved vowing to the existence of conserved B-cell epitopes to which the antibodies can cross react.
Table (2):
Percentage similarity index showing the sequence homology of OmpA among the Enterobacteriaceae pathogens
P. mirabilis |
P. vulgaris |
K. pneumoniae |
S. paratyphi B |
S. paratyphi A |
Styphimurium |
S. enteritidis |
S. typhi |
S. paratyphi C |
S. dysenteriae |
S. boydii |
E. coli |
S. flexneri |
S. sonnei |
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
P. mirabilis |
100.00 |
92.60 |
70.86 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.86 |
71.47 |
71.17 |
P. vulgaris |
92.60 |
100.00 |
70.86 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.25 |
70.86 |
71.47 |
71.17 |
K. pneumoniae |
70.86 |
70.86 |
100.00 |
88.04 |
88.04 |
87.73 |
87.69 |
88.04 |
88.04 |
86.81 |
86.20 |
85.89 |
86.77 |
86.50 |
S. paratyphi B |
70.25 |
70.25 |
88.04 |
100.00 |
99.09 |
99.09 |
99.09 |
99.39 |
99.39 |
94.85 |
95.15 |
93.64 |
94.53 |
95.15 |
S. paratyphi A |
70.25 |
70.25 |
88.04 |
99.09 |
100.00 |
99.39 |
99.39 |
99.09 |
99.09 |
94.85 |
95.15 |
93.64 |
94.53 |
95.15 |
Styphimurium |
70.25 |
70.25 |
87.73 |
99.09 |
99.39 |
100.00 |
100.00 |
99.70 |
99.70 |
95.15 |
95.15 |
93.64 |
94.53 |
95.15 |
S. enteritidis |
70.25 |
70.25 |
87.69 |
99.09 |
99.39 |
100.00 |
100.00 |
99.70 |
99.70 |
95.14 |
95.14 |
93.62 |
94.53 |
95.14 |
S. typhi |
70.25 |
70.25 |
88.04 |
99.39 |
99.09 |
99.70 |
99.70 |
100.00 |
100.00 |
95.15 |
95.15 |
93.64 |
94.53 |
95.15 |
S. paratyphi C |
70.25 |
70.25 |
88.04 |
99.39 |
99.09 |
99.70 |
99.70 |
100.00 |
100.00 |
95.15 |
95.15 |
93.64 |
94.53 |
95.15 |
S. dysenteriae |
70.25 |
70.25 |
86.81 |
94.85 |
94.85 |
95.15 |
95.14 |
95.15 |
95.15 |
100.00 |
99.09 |
97.27 |
98.18 |
98.18 |
S. boydii |
70.25 |
70.25 |
86.20 |
95.15 |
95.15 |
95.15 |
95.14 |
95.15 |
95.15 |
99.09 |
100.00 |
97.58 |
98.48 |
98.48 |
E. coli |
70.86 |
85.89 |
93.64 |
93.64 |
93.64 |
93.62 |
93.64 |
93.64 |
97.27 |
97.58 |
100.00 |
99.09 |
98.48 |
|
S. flexneri |
71.47 |
71.47 |
86.77 |
94.53 |
94.53 |
94.53 |
94.53 |
94.53 |
94.53 |
98.18 |
98.48 |
99.09 |
100.00 |
99.39 |
S. sonnei |
71.17 |
71.17 |
86.50 |
95.15 |
95.15 |
95.15 |
95.14 |
95.15 |
95.15 |
98.18 |
98.48 |
98.48 |
99.39 |
100.00 |
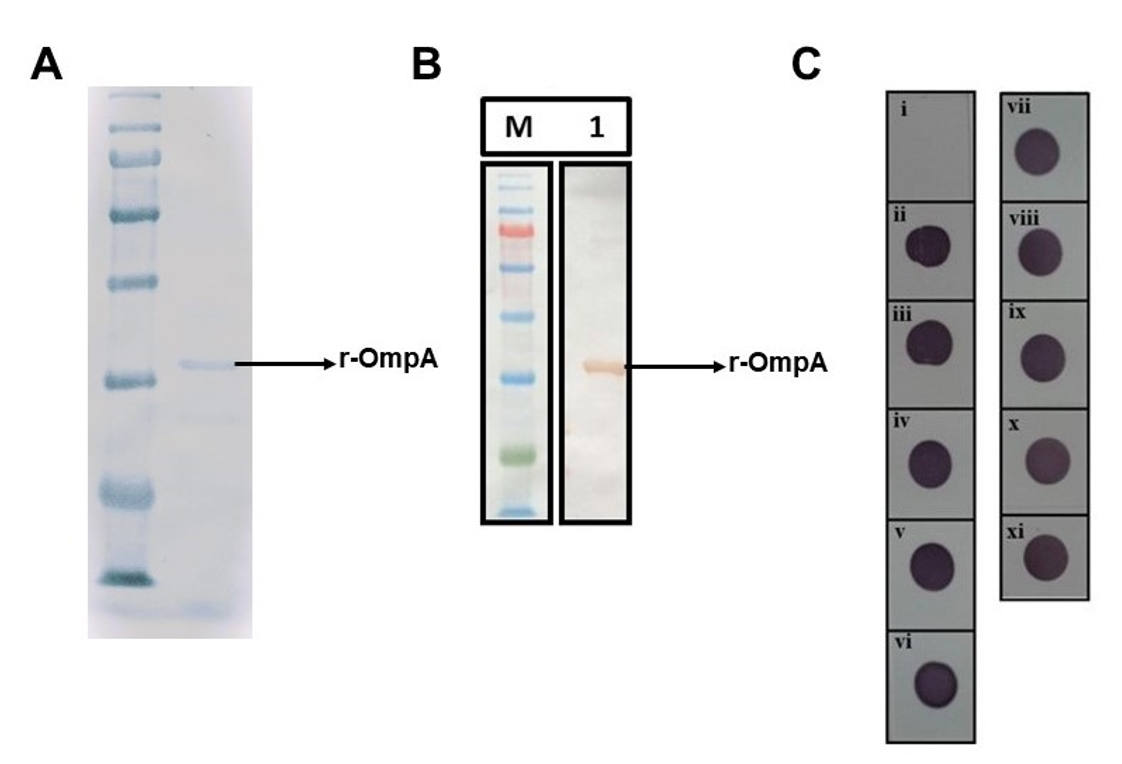
Figure 1. Purification of r-OmpA protein and polyclonal antibody mediated cross reactivity towards the Enterobacteriaceae strains. A. SDS-PAGE image showing the purified r-OmpA. B. Western blot image showing the reactivity of anti-6X histidine antibodies to the purified r-OmpA at ~ 38-kDa. C. Cross reactivity of polyclonal antibodies in the hyperimmune sera was determined through dot ELISA. The polyclonal antibodies in the serum were not reactive to i. negative control, but reactive to ii. Salmonella typhimurium ATCC 14028, iii. Salmonella paratyphi ATCC 9150, iv. Shigella flexneri ATCC 9199, v. Shigella boydii ATCC 9207, vi. Klebsiella pneumoniae ATCC 10031, vii. Proteus vulgaris ATCC 13315, viii. Escherichia coli ATCC 10536, ix. Salmonella paratyphi MTCC 735, x. Salmonella typhi Gwalior isolate and xi. Salmonella enteritidis Gwalior isolate
B-cell epitope conservation in the OmpA
Using BepiPred 2.0 tool, we predicted a total of 7 B-cell epitopes in the OmpA of S. typhimurium (Figure 1). These are QYHDTGFIHNDGPTHENQLGA (38-58 aa), DWLGRMPYKGDNINGAYK (77-94 aa), DTKSNVPGGPSTKDHDTGV (126-144 aa), WTNNIGDANTIGTRPDNGLLSV GVSYRFGQQEAAPVVAPAPAPAPEVQTKHF (168-219 aa), LFNFNKSTLKPEGQQ (226-240 aa), RIGSDAYNQGLSEK (267-280 aa) and GESNPVTGNTCDNVKPRAALIDCL (305-328 aa) respectively (Figure 2). Later, the predicted B-cell epitopes were searched for their amino acid sequence conservation in the alignment of 14 sequences of Enterobacteriaceae pathogens we used for analysis (Figure 3). Among all the 7 predicted epitopes, DWLGRMPYKGDNINGAYK (77-94 aa) and GESNPVTGNTCDNVKPRAALIDCL (305-328 aa) are highly conserved with 72.22% and 75% of the amino acids covering all the 14 pathogens (Figure 3 and 4). Whereas three epitopes WTNNIGDANTIGTRPDNGLLSVGVSYRFG QQEAAPVVAPAPAPAPEVQTKHF (168-219 aa), LFNFNKSTLKPEGQQ (226-240 aa) and RIGSDAYNQGLSEK (267-280 aa) are the next highly conserved ones with at least 60% of amino acid residues covering all the 14 sequences (Figure 3 and 4). Only two epitopes QYHDTGFIHNDGPTHENQLGA (38-58 aa) and DTKSNVPGGPSTKDHDTGV (126-144 aa) are with least amino acid conservation (38.09% and 26.31%) in the OmpA of all the 14 sequences (Figure 3 and 4).
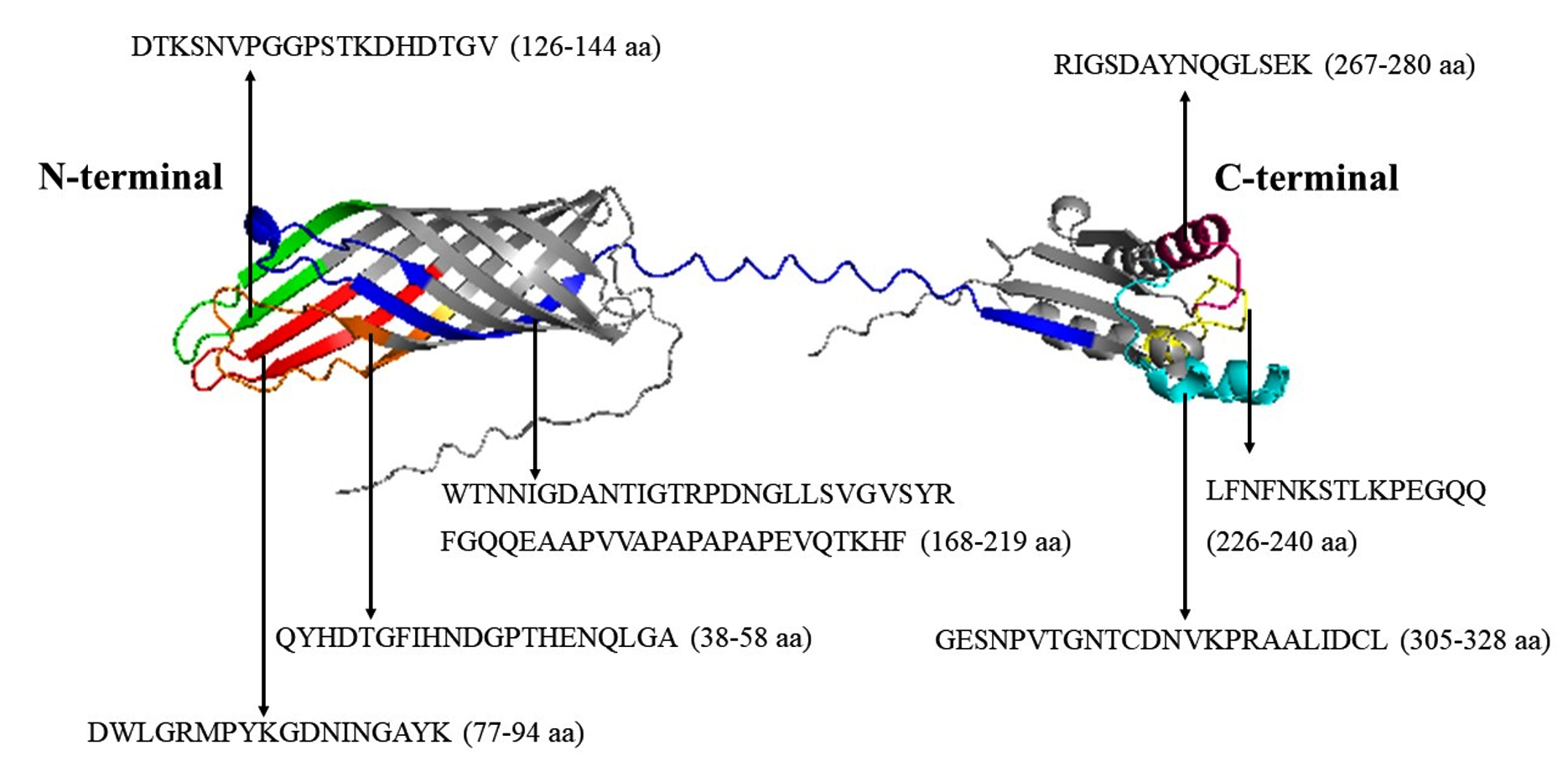
Figure 2. The B-cell epitopes predicted in OmpA of S. typhimurium. The B-cell epitopes were highlighted in different colors (orange, red, green, blue, yellow, pink, and cyan, respectively) three-dimensional tertiary structure of OmpA of S. typhimurium LT2 strain

Figure 3. Multiple sequence alignment showing amino acid sequence conservation of B-cell epitopes in the OmpA among Enterobacteriaceae pathogens. The B-cell epitopes in the OmpA antigen of S. typhimurium are highly conserved in the other Enterobacteriaceae pathogens such as E. coli, Shigella, K. pneumoniae and Proteus sp. This indicates the likelihood of polyclonal antibody mediated cross reactivity among Enterobacteriaceae pathogens.
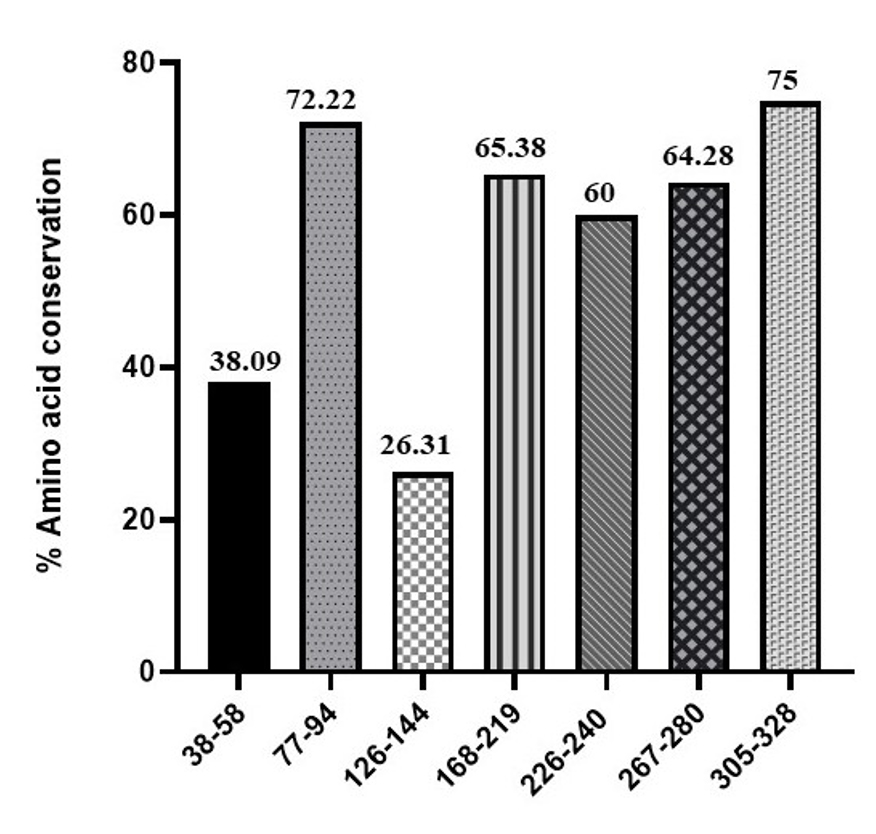
Figure 4. Amino acid sequence conservation in the B-cell epitopes of OmpA among the considered Enterobacteriaceae pathogens. The percent amino acid conservation in the B-cell epitopes was depicted in percent (%)
Antibody cross reactivity provides cross protective immune response against a broad range of closely related pathogens.31-33 Shared epitopes because of amino acid sequence homology among species is the underlying factor for this. Enterobacteriaceae pathogens such as E. coli, Salmonella, Shigella, Proteus and K. pneumoniae are responsible for a wide range of gastrointestinal and other mucosal infections.1-5 Moreover, the family members of Enterobacteriaceae share similarities in the antigenic properties. Significantly, the Enterobacteriaceae OMPs are strong immunogens and also highly conserved with greater amino acid identity.18-21,34 The OMPs play a crucial role in the adherence of bacteria to the surface of host cells. Therefore, prevention or blocking of bacterial adhesion to cell surface will be a promising approach for immune therapy. The antibodies directed against these OMPs will prevent the establishment of infection.
Hence, application of these OMPs in vaccine development and passive immune therapies might confer cross protection against multiple pathogens of Enterobacteriaceae by producing cross reactive antibodies in the host. Our recent study has explored the likelihood of conserved epitopes in the crude OMP of Salmonella and other Enterobacteriaceae pathogens which were revealed with a potent Sal-6 monoclonal antibody (mAb).18 This mAb was raised using crude extracts of OMP proteins recovered from the S. typhimurium cells. Whereas the identity of the OMP to which Sal-6 reacts was not deduced.
In this study, we explored the antibody mediated cross reactivity against the Enterobacteriaceae pathogens as a primary attempt to uncover an important OMP, i.e., OmpA as broad-spectrum epitope vaccine for serious bacterial infections caused by Enterobacteriaceae pathogens. Our results show that the polyclonal antibodies that are raised against rOmpA of S. typhimurium are reactive towards E. coli, Shigella, Salmonella, Proteus, and K. pneumoniae
(Figure 1c). The high degree of amino acid sequence conservation with ~ 66% of identity indicates the presence of conserved B-cell epitopes (Figure 3 and 4) Because of highly conserved B-cell epitopes in OmpA, the polyclonal antibodies are reactive against standard Enterobacteriaceae strains used in this study (Figure 1c). In support with the few other studies,18,21,34 the initial finding in our study is critical to study the cross protective efficiency of OmpA as a subunit vaccine candidate against Enterobacteriaceae infections.
However, T-cell mediated immune response is also very crucial in generating immune response against the pathogen. We also presume the occurrence of several T-cell epitopes in OmpA with high sequence conservation among Enterobacteriaceae. But this presumption should be investigated with appropriate in vitro and in vivo experiments.
Finally, our work resulted in the identification of several conserved B-cell epitopes in the OmpA which are responsible for antibody mediated cross reactivity against the Enterobacteriaceae strains.
The rOmpA could be explored as a potential sub unit immunogen against multiple pathogens of Enterobacteriaceae. Additionally, the antibodies against OmpA could be used as an alternative for antibiotics to treat infections caused by these prominent Enterobacteriaceae pathogens. However, in vitro experiments and pre-clinical evaluation in mice model are highly needed to accomplish the immunogenicity and safety of OmpA for immunoprophylaxis against broad range of Enterobacteriaceae infections.
Finally, our work resulted in the identification of several conserved B-cell epitopes in the OmpA which are responsible for antibody mediated cross reactivity against the Enterobacteriaceae strains. The rOmpA could be explored as a potential subunit immunogen against multiple pathogens of Enterobacteriaceae. Additionally, the antibodies against OmpA could be used as an alternative for antibiotics to treat infections caused by these prominent Enterobacteriaceae pathogens. However, in vitro experiments and preclinical evaluation in mice model are highly needed to accomplish the immunogenicity and safety of OmpA for immunoprophylaxis against broad range of Enterobacteriaceae infections.
ACKNOWLEDGMENTS
HBK, and SLS are thankful to Vignan’s Foundation for Science, Technology and Research (VFSTR) for their support. PNR is thankful to Dr. V. Saleem Basha, Principal, SKR & SKR Govt. Degree College for Women (A), Kadapa, for his support and encouragement. SSM is thankful to ECR research grant provided by Department of Science and Technology, Govt of India (ECR/2016/000685).
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
PNR and SSM designed the study. HBK, SSL and DV performed experiments. HBK performed bioinformatics analysis. HBK, SSL and DV wrote the manuscript. PNR and SSM reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
The study was funded through ECR Research Grant by the Department of Science and Technology, Govt of India (ECR/2016/000685).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study utilized BALB/c mice as animal model. The animals were bred and housed at animal house facility (Reg. No.: 2046/PO/ReBi/S/18/CPCSEA) of Vignan’s Foundation for Science, Technology and Research (VFSTR). Necessary permission to use animals was provided by the Institutional Animal Ethical Committee (IAEC) of VFSTR. The protocols and procedures performed on the animals in the study abide by the guidelines laid by the Committee for Control and Supervision of Experiments on Animals (CCSEA), Govt. of India.
- Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2(2):123-140.
Crossref - Kelmani CR, Chidre PR. Shigellosis: A Conformity Review of the Microbiology, Pathogenesis and Epidemiology with Consequence for Prevention and Management issues. J Pure Appl Microbiol. 2018;12(1):405-417.
Crossref - Zha L, Garrett S, Sun J. Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer. Diseases. 2019;7(1):28.
Crossref - Hamilton AL, Kamm MA, Ng SC, Morrison M. Proteus spp. as Putative Gastrointestinal Pathogens. Clin Microbiol Rev. 2018;31(3):e00085-17.
Crossref - Paczosa MK, Mecsas J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol Mol Biol Rev. 2016;80(3):629-661.
Crossref - Nataro JP, Kaper JB. Diarrheagenic Escherichia coli [published correction appears in Clin Microbiol Rev 1998;11(2):403]. Clin Microbiol Rev. 1998;11(1):142-201.
Crossref - Poolman JT, Wacker M. Extraintestinal Pathogenic Escherichia coli, a Common Human Pathogen: Challenges for Vaccine Development and Progress in the Field. J Infect Dis. 2016;213(1):6-13.
Crossref - Ragupathi NKD, Sethuvel DPM, Inbanathan FY, Veeraraghavan B. Accurate differentiation of Escherichia coli and Shigella serogroups: challenges and strategies. New Microbes New Infect. 2017;21:58-62.
Crossref - Mintz E, Chaignat C. Shigellosis. In: Control of Communicable Diseases Manual. 6th ed. Washington, DC; 2008.
- Centers for Disease Control and Prevention. General Information | Shigella – Shigellosis. CDC. Published August 3, 2016. Archived from the original on April 16, 2017. Accessed April 20, 2017. https://www.cdc.gov
- Baylis C, Uyttendaele M, Joosten H, Davies A. The Enterobacteriaceae and their significance to the food industry. ILSI Europe Report Series. 2011:1-48
- Schaffer JN, Pearson MM. Proteus mirabilis and Urinary Tract Infections. Microbiol Spect. 2015.
Crossref - Senior BW, Leslie DL. Rare occurrence of Proteus vulgaris in faeces: a reason for its rare association with urinary tract infections. J Med Microbiol. 1986;21(2):139-144.
Crossref - Centers for Disease Control. National Nosocomial Infections Study Report. Atlanta, GA: Centers for Disease Control; 1974.
- Ahmad TA, El-Sayed LH, Haroun M, Hussein AA, El Ashry el SH. Development of immunization trials against Klebsiella pneumoniae. Vaccine. 2012;30(14):2411-2420.
Crossref - Huang Y, Li J, Gu D, et al. Rapid Detection of K1 Hypervirulent Klebsiella pneumoniae by MALDI-TOF MS. Front Microbiol. 2015;6:1435.
Crossref - Alkofide H, Alhammad AM, Alruwaili A, et al. Multidrug-Resistant and Extensively Drug-Resistant Enterobacteriaceae: Prevalence, Treatments, and Outcomes – A Retrospective Cohort Study. Infect Drug Resist. 2020;13:4653-4662.
Crossref - Reddy PN, Makam SS, Kota RK, et al. Functional characterization of a broad and potent neutralizing monoclonal antibody directed against outer membrane protein (OMP) of Salmonella typhimurium. Appl Microbiol Biotechnol. 2020;104(6):2651-2661.
Crossref - Babu L, Uppalapati SR, Sripathy MH, Reddy PN. Evaluation of Recombinant Multi-Epitope Outer Membrane Protein-Based Klebsiella pneumoniae Subunit Vaccine in Mouse Model. Front Microbiol. 2017;8:1805.
Crossref - Yang Y, Wan C, Xu H, Wei H. Identification and characterization of OmpL as a potential vaccine candidate for immune-protection against salmonellosis in mice. Vaccine. 2013;31(28):2930-2936.
Crossref - Liu EY, Chen JH, Lin JC, et al. Cross-protection induced by highly conserved outer membrane proteins (Omps) in mice immunized with OmpC of Salmonella Typhi or OmpK36 of Klebsiella pneumoniae. Vaccine. 2022;40(18):2604-2611.
Crossref - Delcour AH. Outer membrane permeability and antibiotic resistance. Biochim Biophys Acta. 2009;1794(5):808-816.
Crossref - Aguilella VM, Queralt-Martin M, Aguilella-Arzo M, Alcaraz A. Insights on the permeability of wide protein channels: measurement and interpretation of ion selectivity. Integr Biol (Camb). 2011;3(3):159-172.
Crossref - Peddayelachagiri BV, Paul S, Makam SS, et al. Functional characterization and evaluation of in vitro protective efficacy of murine monoclonal antibodies BURK24 and BURK37 against Burkholderia pseudomallei. PLoS One. 2014;9(3):e90930.
Crossref - Roy Chowdhury A, Sah S, Varshney U, Chakravortty D. Salmonella Typhimurium outer membrane protein A (OmpA) renders protection from nitrosative stress of macrophages by maintaining the stability of bacterial outer membrane. PLoS Pathog. 2022;18(8):e1010708.
Crossref - Nie D, Hu Y, Chen Z, et al. Outer membrane protein A (OmpA) as a potential therapeutic target for Acinetobacter baumannii infection. J Biomed Sci. 2020;27(1):26.
Crossref - Kota RK, Kolla HB, Reddy PN, Kalagatur NK, Samudrala SK. Immunoinformatics analysis and evaluation of recombinant chimeric triple antigen toxoid (r-HAB) against Staphylococcus aureus toxaemia in mouse model. Appl Microbiol Biotechnol. 2021;105(21-22):8297-8311.
Crossref - Kota RK, Srirama K, Reddy PN. IgY antibodies of chicken do not bind staphylococcal binder of immunoglobulin (Sbi) from Staphylococcus aureus. Annal Microbiol. 2019;69:531-540.
Crossref - Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193(1):265-275.
Crossref - Jespersen MC, Peters B, Nielsen M, Marcatili P. BepiPred-2.0: improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017;45(W1):W24-W29.
Crossref - Fairlie-Clarke KJ, Shuker DM, Graham AL. Why do adaptive immune responses cross-react? Evol Appl. 2009;2(1):122-131.
Crossref - Cywes-Bentley C, Skurnik D, Zaidi T, et al. Antibody to a conserved antigenic target is protective against diverse prokaryotic and eukaryotic pathogens. Proc Natl Acad Sci U S A. 2013;110(24):E2209-E2218.
Crossref - Hoffmann-Veltung H, Anabire NG, Ofori MF, et al. Analysis of allelic cross-reactivity of monoclonal IgG antibodies by a multiplexed reverse FluoroSpot assay. Elife. 2022;11:e79245.
Crossref - Kolla HB, Makam SS, Reddy PN. Mapping of conserved immunodominant epitope peptides in the outer membrane porin (Omp) L of prominent Enterobacteriaceae pathogens associated with gastrointestinal infections. J Genet Eng Biotechnol. 2023;21(1):146.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.