


ISSN: 0973-7510
E-ISSN: 2581-690X
Escherichia coli harbors various virulence factors responsible for diarrhea and other diseases in neonatal calves. The antimicrobial resistance (AMR) among the diarrheagenic and non-diarrheagenic E. coli may impose a potential health hazard among the calves and make the antibacterial therapy ineffective. The virulence gene expression among AMR strains plays a crucial role in establishing diarrheal disease with therapeutic complications. This study investigates the diversity of E. coli isolates from neonatal calves. E. coli isolated from the fecal samples were subsequently subjected to pathotyping and virulotyping using the PCR technique. AMR profiling was done by phenotypic and genotypic methods. The ERIC pattern of E. coli pathotypes was compared and correlated with the pattern of AMR and virulence traits. 179 isolates were obtained from 158 fecal samples collected from neonatal calves. Forty-two isolates were pathotypable; predominated by shigatoxigenic E. coli. Seventy-four isolates were ESBL producers. Nineteen isolates were carbapenem resistant and 68 isolates exhibited quinolone resistance. blaAmpC, blaVIM and qnrS were the predominant genes detected among ESBL, carbapenem and quinolone resistance genes, respectively. The present study indicated diverse virulence profiles and the presence of significant ESBL, carbapenem and fluoroquinolone resistance among E. coli pathotypes from calves. The analysis of resistance profiling indicates the risks associated with the indiscriminate antibiotic use among farm animals, which may pose a potential health hazard to livestock owners.
E. coli, Virulence Genes, Pathotypes, Antimicrobial-resistance, Diarrhea
The antibiotics industry started facing a major setback with an upsurge of multidrug-resistant microbes from the early 1960s.1 The infections induced by antimicrobial-resistant bacteria have a major impact on public health.2 Both pathogenic and commensal organisms may be resistant to the many antimicrobial agents, but the latter is regarded as a potential reservoir of the resistance markers.3 Most members belonging to the genus Escherichia are reported to be commensals.4 Although commensal strains of E. coli rarely cause infections, they can act as reservoirs of resistance genes (RG) that may be transferable to other bacteria, leading to resistant infections.5,6 Antimicrobial resistance (AMR) emerges due to the frequent and indiscriminate use of antimicrobials resulting in the increased shedding of resistant E. coli by the affected individual.7 Also, there is an age-dependent trend in the carriage of resistant genes by fecal E. coli as most of the isolates from calves are significantly more resistant and often multidrug-resistant, compared to that from older cattle.8,9 Production of beta-lactamase enzymes by the bacteria is the most important mechanism by which they acquire resistance to the commonly used b-lactam antibiotics like cephalosporins and penicillins. E. coli are mostly recognized as Extended-spectrum Beta-Lactamases (ESBLs) producers.10 Also, carbapenem resistance in E. coli isolates from livestock may increase the risk of dissemination to humans through contact with animals. Carbapenemase enzymes and efflux pumps primarily mediate carbapenem resistance.11 There are incidences of co-resistance to other antimicrobial drugs along with carbapenem resistance.12 Fluoroquinolone resistance has also been reported in E. coli and fluoroquinolone-resistant E. coli is often resistant to all other main classes of available antimicrobials such as gentamicin, tetracycline, ampicillin, chloramphenicol, and trimethoprim/sulfamethoxazole.13
Though the majority of the fecal E. coli isolates are found to be non-pathogenic commensals, as a result of acquiring virulence and RG, some of the commensals can evolve into pathogenic strains.14 Pathogenic and commensal E. coli are frequently isolated from diarrheic cases of neonatal calves. Diarrhea is an important cause of economic losses in the dairy industry. The pathotypes of Escherichia coli come under the category of Diarrhegenic E. coli (DEC), which is recognized as the most important bacterial cause of diarrhea, leading to high mortality in calves.15 The various pathotypes of DEC include Enterotoxigenic E. coli (ETEC), Enteropathogenic E. coli (EPEC), Enterohemorrhagic E. coli (EHEC), Shiga-like toxin-producing E. coli (STEC), Enteroinvasive E. coli (EIEC), Enteroaggregative E. coli (EAEC) and Diffusely Adherent E. coli (DAEC).16 Cattle may act as a reservoir of STEC/EHEC, but they do not develop the systemic disease due to the lack of Stx receptor. However, they can survive and propagate inside the host intestine with the help of immunomodulation and intestinal colonization.17,18 EHEC is frequently shed by cattle and approximately 75% of the human outbreaks are associated with cattle or bovine-derived products.19 Among these, Few strains have features of different pathotypes, making them possibly more virulent hybrid pathogenic strains.16 The number of virulence factors (VF) carried by the pathogenic E. coli strains plays a crucial role in the colonization and contributes to the pathogenicity.20
Very few studies have investigated the association between the virulence and resistance pattern of E. coli isolates from neonatal calves in India. Therefore, it is customary to study the distribution and occurrence of virulence genes (VG) and their relationship with resistance patterns. With this background, the present study was conducted on virulotyping and AMR profiling of E. coli associated with neonatal calves.
Faecal sampling, isolation and identification
A total of n = 158 fecal samples were collected from the neonatal calves (up to the age of one month) of two different farms (multiple times from the Cattle and buffalo farm (CBF), ICAR-IVRI, Bareilly, Uttar Pradesh, and two times from the Instructional dairy farm (IDF), GBPUAT, Pantnagar, Uttarakhand). Out of this, 65 were diarrheic (watery to pasty consistency with or without blood) samples and 93 were from non-diarrheic animals. The samples were inoculated in buffered peptone water and incubated overnight at
37°C for enrichment. The enriched culture was further streaked on Eosin methylene blue (EMB) agar plates and incubated at 37°C for 24 hours. Colonies revealing characteristic metallic sheen on EMB agar were subjected to biochemical characterization by using a HiMViC kit (Himedia, India).
Pathotyping of E. coli isolates
The E. coli isolates were further pathotyped by multiplex PCR.21 DNA was extracted using QIAamp DNA Mini Kit following standard protocol. Multiplex PCR was optimized for the detection of different pathotypes viz., EPEC (eae and bfp), EHEC (eae, stx1, stx2 and hly), STEC (stx1, stx2, and eae), EAEC (aggR and pic), ETEC (elt, esta, and estb) and EIEC (ipaH and invE). Details of the primers used for pathotyping are given in Supplementary Table 1. ATCC 25922 (E. coli) was used as negative control.
Table (1):
Prevalence of Resistance Genes in E. coli Isolates. ESBL resistance genes were present among 93.24% of phenotypically resistant E. coli isolates, whereas quinolone resistance and carbapenem resistance genes were found among 47.05% and 10.52% of resistant isolates
| Phenotypic resistance (No. of isolates) | Genotypic resistance among phenotypically resistant isolates | Resistance genes | Presence among phenotypically resistant isolates | Presence among phenotypically susceptible isolates |
|---|---|---|---|---|
| ESBL resistance (74 isolates) | 69 (93.24%) | blaAmpC (153) | 68 (91.89%) | 85 (80.95%) |
| blaCTX-M (59) | 45 (60.81%) | 14 (13.33%) | ||
| blaTEM (43) | 17 (22.97%) | 26 (24.76%) | ||
| Carbapenem resistance (19 isolates) | 2 (10.52%) | blaVIM (3) | – | 3 (1.87%) |
| blaNDM (2) | 2 (10.52%) | – | ||
| blaBIC (1) | – | 1 (0.62%) | ||
| Quinolone resistance (68 isolates) | 32 (47.05%) | qnrS (82) | 27 (39.70%) | 55 (49.54%) |
| qnrB (13) | 6 (8.82%) | 7 (6.30%) | ||
| aac(6′)-Ib-cr (6) | 6 (8.82%) | – | ||
| qepA (3) | 2 (2.94%) | 1 (0.90%) |
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing of the E. coli isolates was done using 18 antibiotics (9 different classes) using Kirby Bauer disc diffusion method as per CLSI guidelines (CLSI, 2018).22 The Combination disc diffusion method was used for phenotypic detection of ESBL producing E. coli as per CLSI recommendation. An increase of 5 mm or more in the zone of inhibition of the disc containing a combination of antibiotic and clavulanic acid, than the disc containing the antibiotic alone was indicating of ESBL production. Phenotypic resistance against carbapenem and quinolones was also checked. The following antimicrobial agents at concentrations mentioned were used for antimicrobial susceptibility testing: Streptomycin (10 µg), Gentamicin (10 µg), Kanamycin (30 µg), Amikacin (30 µg), Cefotaxime (30 µg), Ceftazidime (30 µg), Colistin (10 µg), Enrofloxacin (10 µg), Ciprofloxacin (5 µg), Ampicillin (10 µg), Meropenem (10 µg), Ertapenem (10 µg), Doripenem (10 µg), Imipenem (10 µg), Tetracycline (30 µg), Cotrimoxazole (25 µg), Sulpadiazine (100 µg), Trimethoprim (10 µg).
Additionally, ESBL production, carbapenem and quinolone resistance among the E. coli isolates were determined genotypically using PCR. Eight genes were targeted for the determination of ESBL production. The protocol included a multiplex PCR for the detection of 5 different alleles of the blaCTX gene (blaCTX-M1, blaCTX-M2, blaCTX-M8, blaCTX-M9, and blaCTX-M25)23 and three uniplex PCRs for detecting blaAmpC, blaTEM and blaSHV genes. Detection of carbapenem RG included two sets of multiplex PCRs: blaIMP, blaVIM, blaSPM & blaOXA-23 and blaOXA-48, blaKPC, blaBIC & blaNDM respectively.24,25 A multiplex PCR targeting eight different genes (qnrA, qnrB, qnrC, qnrD, qnrS, oqxAB, qepA and aac-(6′)-Ib-cr) was used for detecting quinolone resistance in E. coli isolates.26 Details of the primers used for genotypic AMR detection are given in Supplementary Table 1. E. coli ATCC 25922, E. coli ATCC 35218, E. coli ATCC 2496, K. pneumoniae ATCC 700603, and K. pneumoniae NCTC 13440 (K. pneumoniae) were used as reference strains.
Virulence gene detection
All the E. coli isolates were investigated for 14 different VG (lpfAO113, efa1, katP, mat, fimC, hrA, iss, ibeA, ompA, traT, chuA, iroN, ehxA and iha) by PCR. Three uniplex PCRs were used for detecting lpfAO113, efa1 and katP genes. Two multiplex PCRs for detecting two sets of genes mat, fimC, hrA & iss, and ibeA, ompA, traT, chuA & iroN, respectively and one duplex PCR targeting ehxA and iha genes were also performed. Details of the primers used for virulotyping are given in Supplementary Table 1.
Determination of E. coli molecular heterogeneity
E. coli isolates were genotyped using Enterobacterial Repetitive Intergenic Consensus (ERIC) PCR.27 The PCR condition was as follows: denaturation for 3 min at 95°C, followed by 35 cycles of 95°C for 0.5 min, 51.2°C for 1 min, and 72°C for 2 min, followed by a final extension for 5 min at 72°C. Gel images were captured after electrophoresis, and the dendrogram was constructed using GelJ and visualized using FigTree v1.4.4. Details of the primers used for the ERIC PCR are given in Supplementary Table 1.
All the PCR reactions were carried out in 25 µL consisting of 10 X PCR buffer (Thermo Scientific), 25 mM (final concentration in 25 µL) each of dATP, dGTP, dTTP and dCTP (Thermo Scientific), 0.625 U of DNA Taq polymerase (Thermo Scientific), 1 µl of DNA template and the final volume was made up to 25 µl by adding nuclease-free water. Concentrations of all the primers used are given in the supplementary Table 1. Apart from this, 2.5 mM MgCl2 was added to the reaction mixture for the multiplex PCRs.
Isolation of E. coli
179 isolates were recovered from 158 samples (77 isolates from CBF, IVRI and 102 from IDF, GBPUAT). 42 (23.46%) E. coli isolates were diarrheagenic pathotypes with STEC being the most prevalent pathotype (n = 18/179; 10.05%), followed by EPEC (n = 15/179; 8.37%, EHEC (n = 7/179; 3.9%) and EAEC (n = 2/179; 1.11%).
Phenotypic and genotypic antimicrobial resistance profiling
The AMR profiling revealed a high occurrence of multidrug-resistant (MDR) E. coli isolates among neonates. 135 (75.41%) isolates were MDR, among which 102 were commensal E. coli. Resistance to ceftazidime (84.35%; n = 151) and cefotaxime (67.59%; n = 121) was highest and the lowest resistance was shown to ertapenem (6.70%; n = 12) and doripenem (6.14%; n = 11). None of the E. coli isolates were resistant to meropenem and colistin. The complete resistance profile among E. coli isolates is depicted in Figure 1. Out of 179 isolates, 74 (41.34%) were phenotypic ESBL producers, 68 (37.98%) were quinolone-resistant and 19 (10.61%) were carbapenem-resistant. Out of 74 ESBL producers, 69 exhibited the presence of at least one gene for ESBL production. The predominant ESBL gene, blaAmpC was present among 85.47% (n = 153/179) of the isolates. 91.89% (n = 68/74) of phenotypic ESBL producers were positive for the ESBL gene, blaAmpC followed by blaCTX-M (60.81%; n = 45/74) and blaTEM (22.97%; n = 17/74). The M1 allele group was the predominant type among blaCTX-M positive isolates. Six isolates had carbapenem RG and out of 19 resistant isolates, only two carried carbapenem RG (blaNDM among two isolates). 68 isolates were resistant phenotypically to quinolone drugs. qnrS (39.70%; n = 27/68) was the predominant gene followed by qnrB, aac(6’)-lb-cr (8.82 % each; n = 6/68 ) and qepA (2.94%; n=2/68). The carriage of RG by phenotypically resistant and susceptible E. coli isolates is depicted in Table 1.
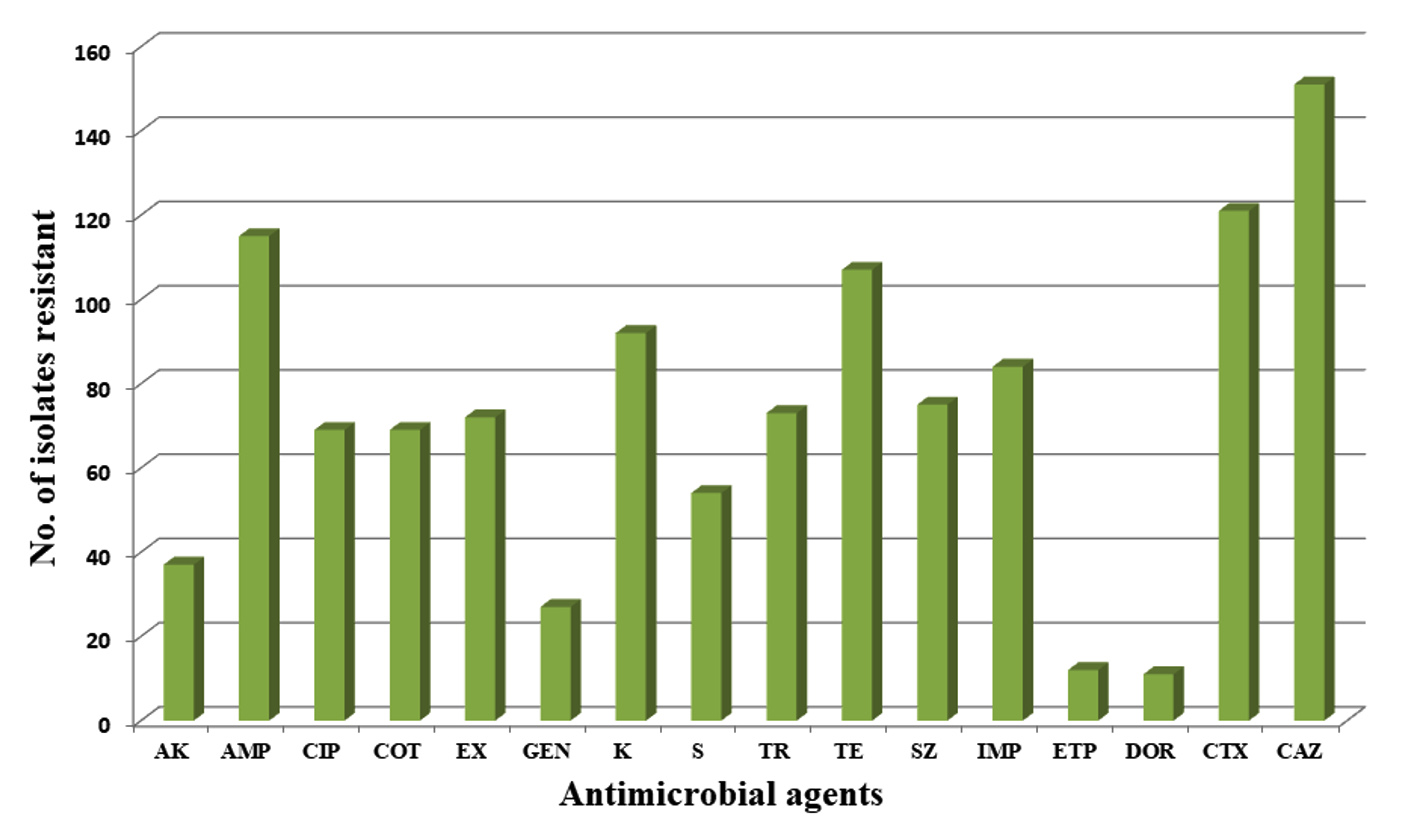
Figure 1. Number of E. coli isolates resistant to various drugs in the study. The highest resistance was found against the cephalosporin class [ceftazidime (84.35%; n = 151) and cefotaxime (67.59%; n = 121)] and the lowest resistance was shown against carbapenem class of drugs [ertapenem (6.70%; n = 12) and doripenem (6.14%; n = 11)]. No resistance was detected against meropenem and colistin
Virulence gene profiling
The distribution and presence of 14 VG were investigated. The incidence of the ompA gene was highest and detected among 99.44% (n = 178/179) of the isolates, followed by fimC
(n = 161/179; 89.94%) and mat (n = 160/179; 89.38%). ibeA was detected only in one isolate. All the isolates screened in this study carried at least one virulence gene. 30.16% (n = 54/179) of the isolates carried seven or more VG. Out of 42 pathotypes, the majority (83.33%; n = 35/42) had six or more VG. The ehxA and katP genes were present exclusively among pathotypes. The distribution of VG among pathotypes and isolates from diarrheic animals is shown in Table 2.
Table (2):
Prevalence of Virulence Genes in E. coli Isolates. The occurrence of the virulence genes was marginally higher among isolates from diarrheic calves than the healthy ones except for mat, iha, ehxA and efa1. ehxA and katP were present exclusively among pathotypes. All STEC isolates had traT, iha, mat and ompA genes. traT, iha, efa1, mat, ompA and ehxA were possessed by all the EHEC pathotypes
| Virulence genes | Health status | Isolate type | Total | |||||
|---|---|---|---|---|---|---|---|---|
| Diarrheic | Non-diarrheic | STEC | EPEC | EHEC | EAEC | Non-typable isolates | ||
| Total No. of isolates (179) | 76 | 103 | 18 | 15 | 7 | 2 | 137 | |
| ompA (Outer membrane protein A) | 76 (100%) | 102 (99.02%) | 18 (100%) | 15 (100%) | 7 (100%) | 2 (100%) | 136 (99.27%) | 178 |
| fimC (Chaperone protein FimC) | 70 (92.10%) | 91 (88.34%) | 17 (94.44%) | 12 (80%) | 4 (57.14%) | 1 (50%) | 127 (92.70%) | 161 |
| mat (meningitis-associated and temperature-regulated fimbria) | 65 (85.52%) | 95 (92.23%) | 18 (100%) | 15 (100%) | 7 (100%) | 1 (50%) | 119 (86.86%) | 160 |
| traT (Transfer genes) | 54 (71.05%) | 70 (67.96%) | 18 (100%) | 12 (80%) | 7 (100%) | 1 (50%) | 86 (62.77%) | 124 |
| lpfAO133 (Long polar fimbria) | 38 (50%) | 41 (39.80%) | 7 (38.88%) | 8 (53.33%) | 3 (42.85%) | – | 61 (44.52%) | 79 |
| iha (Iron-regulated gene homologue adhesin) | 19 (25%) | 27 (26.21%) | 18 (100%) | 13 (86.66%) | 7 (100%) | – | 8 (5.83%) | 46 |
| hrA (Heat-resistant agglutinin) | 15 (19.73%) | 20 (19.41%) | 4 (22.22%) | – | – | 1 (50%) | 30 (21.89%) | 35 |
| efa1 (EHEC factor for adherence) | 13 (17.10%) | 21 (20.38%) | 8 (44.44%) | 11 (73.33%) | 7 (100%) | 1 (50%) | 7 (5.10%) | 34 |
| iroN (catecholate-siderophore) | 16 (21.05%) | 13 (12.62%) | 9 (50%) | 2 (13.33%) | – | – | 18 (13.13%) | 29 |
| chuA (E. coli heme- utilization) | 17 (22.36%) | 12 (11.65%) | 3 (16.66%) | – | – | 1 (50%) | 25 (18.24%) | 29 |
| iss (Increased serum survival) | 13 (17.10%) | 14 (13.59%) | 7 (38.88%) | 2 (13.33%) | – | 1 (50%) | 17 (12.40%) | 27 |
| ehxA (entero-haemolysin) | 8 (10.52%) | 17 (16.5%) | 6 (33.33%) | 12 (80%) | 7 (100%) | – | – | 25 |
| katP (Catalase peroxidase) | 8 (10.52%) | 11 (10.67%) | 3 (16.66%) | 10 (66.66%) | 6 (85.71%) | – | – | 19 |
Molecular fingerprinting
Genotyping was carried out using ERIC-PCR. PCR fingerprints were used to generate a dendrogram to understand the genetic relatedness among isolates. Two major clusters could be observed among the isolates, with most pathotypes falling under one cluster. Mostly, isolates included in a single cluster were from the same farm. The isolates within each cluster did not show any correlation for the health status designated. The related genotypes within the clusters showed a limited correlation concerning genotypic AMR and VF. Genetic diversity analysis of E. coli isolates using ERIC-PCR fingerprints-based dendrogram is depicted in Figure 2. The analysis shows that E. coli isolates recovered from the calves may indicate the predominant genetic lineages in the farm E. coli population. Also, the isolates recovered from diarrheic calves may not be probably involved in clinical diarrhea in all cases.
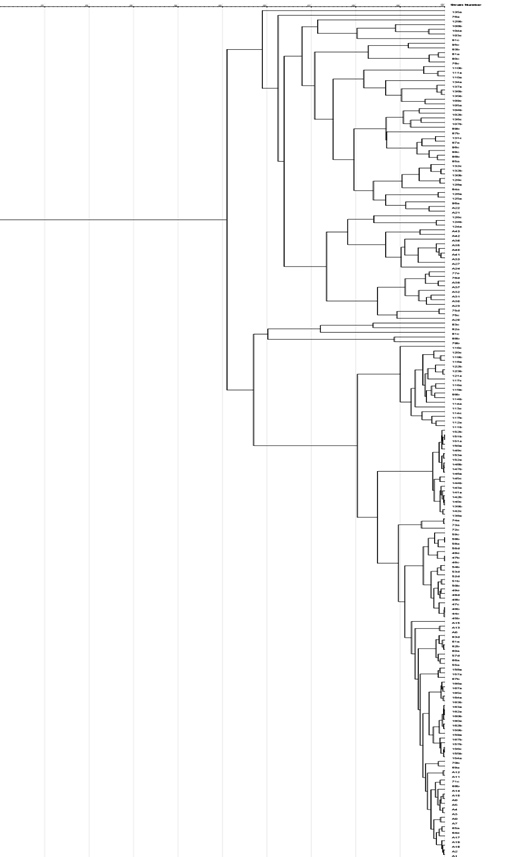
Figure 2. Dendrogram created using the ERIC pattern showing genetic relatedness among E. coli isolates. Two major clusters with most pathotypes falling under one cluster. Most isolates in a single cluster were from the same farm
Statistical analysis
A heatmap was constructed with hierarchical clustering using R software v.4.3.2 with “pheatmap” package v1.0.12. A correlation plot was prepared using “corrplot” package v0.92 by calculating Spearman’s rank correlation coefficient. A heatmap constructed with a hierarchical clustering algorithm identified numerous instances of pathotype-based clustering and locality-based clustering among the isolates. The heatmap is shown in Figure 3. The correlation plot revealed a high correlation among resistance to ceftazidime, cefotaxime, ESBL production, and the presence of blaAmpC gene (p < 0.05) (Figure 4).
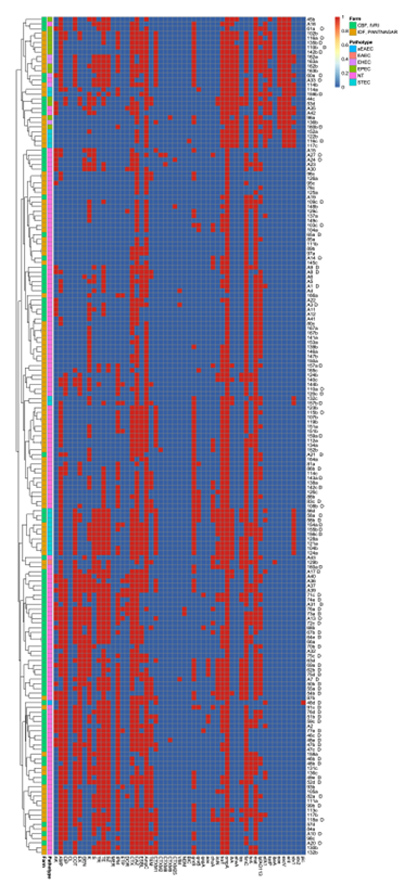
Figure 3. Heat map representing resistance and virulence genes among isolates of various pathotypes obtained from the two farms. Several occurrences of pathotype-based clustering and locality-based clustering were identified among the E. coli isolates

Figure 4. Diagrammatic representation of the correlation between the virulence and resistance genes. A high correlation among resistance to ceftazidime, cefotaxime, ESBL production, and the presence of blaAmpC gene was revealed
AMR is a major concern of public health importance. Carbapenem-resistant and ESBL-producing bacteria under Enterobacteriaceae have been listed as the ‘Priority 1: CRITICAL’ pathogens according to the WHO global priority pathogens list of antibiotic-resistant bacteria. Among these, E. coli is the most common producer of ESBL as well as the most common etiological agent of diarrhea and septicemia among neonatal calves. Diarrhea in young calves is a major concern due to the multi-factorial nature of the disease and hence it is imperative to identify the etiological agent and other risk factors associated with the calf diarrhea so that appropriate prevention and control measures may be implemented. In our study, STEC was the predominant pathotype identified. The most prevalent pathotypes detected from IDF, GBPUAT and CBF, IVRI were STEC and EPEC, respectively. Many studies were conducted throughout the world on the pathotyping of E. coli showing variations among predominance of pathotypes.28-31 The variation in the pathotypes and their predominance among calves may be due to the geographical locations, managemental practices, vaccinations and hygienic measures adopted at farms.32,33
The antibiogram revealed a higher occurrence of multidrug-resistant E. coli (75.41%) among the neonatal calves which indicates the indiscriminate antibiotic use in the farm. A similar study from India reported an incidence of 69.81% of MDR strains among E. coli.34 The resistance was highest against ceftazidime (84.35%), followed by cefotaxime (67.59%), ampicillin (64.24%) and tetracycline (59.77%). Similar findings were reported by Batabyal et al.,35 from India. 102 commensal E. coli isolates were found to be MDR. The isolation of resistant commensal E. coli from healthy animals can be considered as an indicator of long-term resistance among the animal populations.36
A total of 74 isolates were phenotypic ESBL producers and out of which, 69 were genotypically positive. blaAmpC was the predominant ESBL gene (91.89%). A similar study was conducted by Ibrahim et al.,37 A high percentage of CTX-M genes (32.8%) among ESBL-producing E. coli was reported by Schmid et al.,38 Few of the non-ESBL phenotypes also carried ESBL genes indicating the role of such isolates in the dissemination of ESBL resistance in the herd without its phenotypic expression. The minor differences between the genotypic and phenotypic ESBL producers may be attributed to the presence of other genes encoding ESBL resistance.
There has been a consistent increase in carbapenem usage nowadays due to the gradual rise in the ESBL resistance. In our study, 19 isolates were carbapenem-resistant but the expression of the resistance gene was much lower. Only six isolates (three blaVIM, two blaNDM and one blaBIC) were positive for the RG of carbapenem. In a similar study, 29.3% of isolates were carbapenem-resistant and the blaVIM gene could be detected only in one isolate.39 There may be the existence of new variants of carbapenemases being present among the E. coli population which could not be detected by PCR-based gene identification methods. Whole genome-based approaches may be an alternative to characterize all genetic elements involved among resistant isolates. 68 isolates were phenotypically quinolone resistant. Among these, 47.05% were genotypically positive for the quinolone resistance gene and qnrS (39.70%) was the predominant resistance gene among resistant isolates. the higher incidence of qnrS and qnrB genes was reported by other researchers also.40
We found a higher predominance of MDR, quinolone resistance and carbapenem resistance among commensals. These commensal bacteria may act as a reservoir of RG that may be transferable to other susceptible bacteria.7 The isolation of resistant commensal E. coli from healthy animals can be considered as an indicator of long-term resistance among the animal populations.36
To designate a pathotypic E. coli as virulent and potentially pathogenic, it is imperative to perform the virulotyping.41 In India, studies on the virulotyping of E. coli strains from neonatal calves are very limited. Out of fourteen VG investigated, ompA (adhesin and invasin, participate in biofilm formation) was the predominant virulence gene (99.44%) followed by fimC (89.94%) and mat (89.38%) genes involved in cellular adhesion. ibeA and katP genes had the lowest incidence (in one and 19 isolates respectively). More than 90% of fimC-positive and less than 12% of katP-positive E. coli strains from calves were reported from Sweden and agree with our findings.42 Likewise, an analogous study reported ompA among 100% of the commensal E. coli and EPEC strains, and
0% and 20.5% hrA among EPEC and commensal E. coli, respectively which is similar to our findings.20 A higher rate of occurrence of the traT gene among E. coli strains from diarrheic (56-70%) than the non-diarrheic feces (20-40%) is following our findings.43 Variations in results may be attributed to the location of the farms, the number of isolates, and the VG studied. The marginally higher occurrence of some of the VG among isolates from diarrheic calves indicates the role of such genes in the pathogenicity of diarrhea. The ehxA and katP genes were exclusively associated with pathotypes. 100% of STEC had traT, iha, mat and ompA gene. The traT, iha, efa1, mat, ompA and ehxA genes were possessed by all the EHEC pathotypes. 86.66% (n = 13/15) of EPEC contained iha gene and traT, fimC & ehxA were present in 80% (n = 12/15) of EPEC isolates. Higher occurrence of traT, iha, mat and ompA, efa1, ehxA genes among pathotypes indicates increased risk and severity of infections caused by the pathotypes and the importance of these genes in establishing more severe infection.
Pathotype-based clustering & locality-based clustering and a high correlation among resistance to ceftazidime, cefotaxime, ESBL production & presence of blaAmpC gene were revealed among the isolates under the study Phylogenetic analysis revealed two major clusters with the most pathotypes falling under same cluster. However, there was lesser correlation of related genotypes to AMR and VF. This may indicate the role of horizontal transfer in maintaining the VF and AMR determinants which could be maintained in genetically unrelated genotypes. Virulence markers like traT, iss, ehxA and katP have been reported to be carried by plasmids.44-46 Both ESBL and carbapenemase encoding genes are commonly located on mobile genetic elements enabling their dissemination. The findings almost coincide with other such investigations that focused on the genetic resemblance of E. coli isolates from different sources.47-49
Our study envisages the virulotyping and AMR profiling of the Escherichia coli isolates recovered from neonatal calves. Most of the commensal E. coli under the study were MDR, indicating long-term resistance among the animal populations. The higher occurrence of RG among phenotypically resistant isolates signifies the association between phenotypic and genotypic resistance. The resistance profile indicated a high risk associated with indiscriminate antimicrobial use in animals. The predominance of STEC isolates carrying major VF substantiates their role in calf infections and may act as a potential source for human transmission. The presence of certain VG exclusively among pathotypes reveals the importance of such VF in the severity of infections. The study also revealed pathotype-based and locality-based clustering among the isolates and a high correlation of ESBL resistance and ESBL genes but a lesser correlation of related genotypes to AMR and VF. The study may be helpful in the development of a prophylactic vaccine against major pathotypes causing diarrhea and mortality among calves.
Additional file: Additional Table S1.
ACKNOWLEDGMENTS
The authors would like to thank the Indian Council of Agricultural Research (ICAR), New Delhi, for funding the All India Network Programme on Neonatal Mortality in Farm Animals.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
A and PT designed and conceptualized the study. VA and SSN did the sampling and isolation of E. coli. AV and SI performed pathotyping, antimicrobial sensitivity testing and virulence gene profiling. KS and PT helped generate and analyze the ERIC profile. PC and VKC supervised the study. AV, SSN, and TSA wrote the manuscript. PD, MSK and BK edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This study was carried out in part with financial assistance from the Indian Council of Agricultural Research (ICAR), New Delhi (Project No: 1005643).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript and/or the Supplementary files.
ETHICS STATEMENT
Not applicable.
- Phoenix DA, Dennison SR, Harris F. Antimicrobial peptides: Their history, evolution, and functional promiscuity. Antimicrobial Peptides. 2013:1-37.
Crossref - Wernli D, Haustein T, Conly J, Carmeli Y, Kickbusch I, Harbarth S. A call for action: the application of the international health regulations to the global threat of antimicrobial resistance. PLoS Med. 2011;8(4):e1001022.
Crossref - Shoemaker NB, Vlamakis H, Hayes K, Salyers AA. Evidence for extensive resistance gene transfer among Bacteroides spp. and other genera in the human colon. Appl Environ Microbiol. 2001;67(2):561-568.
- Tenaillon O, Skurnik D, Picard B, Denamur E. The population genetics of commensal Escherichia coli. Nat Rev Microbiol. 2010;8(3):207-217.
Crossref - Sorum H, Sunde M, Resistance to antibiotics in the normal flora of animals. Vet Res. 2001;32(3-4):227-241.
Crossref - Summers AO. Generally overlooked fundamentals of bacterial genetics and ecology. Clin Infect Dis.2002;34(Suppl 3):S85-S92.
Crossref - Van den Bogaard AE, Stobberingh EE, Epidemiology of resistance to antibiotics: links between animals and humans. Int J Antimicrob Agents. 2000;14(4):327-335.
Crossref - Berge AC, Hancock DD, Sischo WM, Besser TE. Geographic, farm, and animal factors associated with multiple antimicrobial resistance in fecal Escherichia coli isolates from cattle in the western United States. J Am Vet Med Assoc. 2010;236(12):1338-1344.
Crossref - Yamamoto S, Iwabuchi E, Hasegawa M, et al. Prevalence and molecular epidemiological characterization of antimicrobial-resistant Escherichia coli isolates from Japanese black beef cattle. J Food Prot.2013;76(3):394-404.
Crossref - Alizade H, Fallah F, Ghanbarpour R, et al. Genotyping of ESBL producing uropathogenic and diarrheagenic Escherichia coli in southeast of Iran. Infect Disord Drug Targets. 2015;15(2):118-124.
Crossref - Nordmann P, Naas T, Poirel L. Global spread of carbapenemase-producing Enterobacteriaceae, Emerg Infect Dis. 2011;17(10): 1791–1798.
Crossref - Webb HE, Bugarel M, Den Bakker HC, et al. Carbapenem-resistant bacteria recovered from faeces of dairy cattle in the high plains region of the USA, PLoS One. 2016;11(1):0147363.
Crossref - Kuntaman K, Lestari ES, Severin JA. Fluoroquinolone-resistant Escherichia coli, Indonesia, Emerg Infect Dis. 2005;11(9):1363–1369.
Crossref - de Sousa CP. Escherichia coli as a specialized bacterial pathogen. Rev Biol Cienc Terra. 2006;2(2):341-352.
- Windeyer MC, Leslie KE, Godden SM, Hodgins DC, Lissemore KD, LeBlanc SJ. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age, Prev Vet Med. 2014;113(2):231-240.
Crossref - Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent Advances in Understanding Enteric Pathogenic Escherichia coli, Clin Microbiol Rev. 2013;26(4):822-880. .
Crossref - Menge C, Wieler LH, Schlapp T, Baljer G. Shiga toxin 1 from Escherichia coli blocks activation and proliferation of bovine lymphocyte subpopulations in vitro, Infect Immun. 1999;67(5):2209-2217.
Crossref - Etcheverria AI, Padola NL. Shiga toxin-producing Escherichia coli: factors involved in virulence and cattle colonization, Virulence. 2013;4(5):366-372.
Crossref - Nguyen Y, Sperandio V. Enterohemorrhagic E. coli (EHEC) pathogenesis, Front Cell Infect Microbiol. 2012;2:90.
Crossref - Nowak K, Fahr J, Weber N, et al. Highly diverse and antimicrobial susceptible Escherichia coli display a naive bacterial population in fruit bats from the Republic of Congo, PLoS One. 2017;12(7):e0178146.
Crossref - Antikainen J, Tarkka E, Haukka K, Siitonen A, Vaara M, Kirveskari J. New 16-plex PCR method for rapid detection of diarrheagenic Escherichia coli directly from stool samples, Eur J Clin Microbiol Infect Dis. 2009;28(8):899-908.
Crossref - CLSI. Performance standards for antimicrobial susceptibility testing, 28th edn. CLSI supplement M100. Clinical and Laboratory Standards Institute, Wayne. 2018.
- Woodford N, Fagan EJ, Ellington MJ. Multiplex PCR for rapid detection of genes encoding CTX-M extended spectrum b-lactamases, J Antimicrob Chemother. 2006;57(1):154-155.
Crossref - Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes, Diagn Microbiol Infect Dis. 2011;70(1):119-123.
Crossref - Paul D, Ingti B, Bhattacharjee D, et al. An unusual occurrence of plasmid-mediated blaOXA-23 carbapenemase in clinical isolates of Escherichia coli from India, Int J Antimicrob Agents, 2017;49(5):642-645.
Crossref - Ciesielczuk H, Hornsey M, Choi V, Woodford N, Wareham DW. Development and evaluation of a multiplex PCR for eight plasmid-mediated quinolone-resistance determinants, J Med Microbiol. 2013;62(12):1823-1827.
Crossref - Adamus-Bialek W, Wojtasik A, Majchrzak M, Sosnowski M, Parniewski P. (CGG) 4-based PCR as a novel tool for discrimination of uropathogenic Escherichia coli strains: comparison with enterobacterial repetitive intergenic consensus-PCR, J Clin Microbiol. 2009;47(12):3937-3944.
Crossref - Wieler LH, Sobjinski G, Schlapp T, et al. Longitudinal prevalence study of diarrheagenic Escherichia coli in dairy calves. Berl Munch Tierarztl Wochenschr. 2007;120(7):296-306.
- Wani SA, Hussain I, Beg SA, et al. Diarrhoeagenic Escherichia coli and salmonellae in calves and lambs in Kashmir: absence, prevalence and antibiogram, Rev Sci Tech Off Int Epiz.2013;32(3):833-840.
Crossref - Shahrani M, Dehkordi FS, Momtaz H. Characterization of Escherichia coli virulence genes, pathotypes and antibiotic resistance properties in diarrheic calves in Iran. Biol Res. 2014;47:1-13.
Crossref - Dhaka P, Vijay D, Vergis J, et al. Genetic diversity and antibiogram profile of diarrheagenic Escherichia coli pathotypes isolated from human, animal, foods and associated environmental sources, Infect Ecol Epidemiol. 2016;6(1):31055.
Crossref - Cho YI, Han JI, Wang C, et al. Case control study of microbiological etiology associated with calf diarrhea, Vet Microbiol. 2013;166(3-4):375-385.
Crossref - El-Seedy FR, Abed AH, Yanni HA, El-Rahman SAAA. Prevalence of Salmonella and E. coli in neonatal diarrheic calves, Beni-Suef Univ J Basic Appl Sci. 2016;5(1):45-51.
Crossref - Srivani M, Reddy YN, Subramanyam KV, Reddy MR, Rao TS. Prevalence and antimicrobial resistance pattern of Shiga toxigenic Escherichia coli in diarrheic buffalo calves, Vet World. 2017;10(7):774.
Crossref - Batabyal K, Banerjee A, Pal S, et al. Detection, characterization, and antibiogram of extended spectrum beta-lactamase Escherichia coliisolated from bovine milk samples in West Bengal, India. Vet World. 2018;11(10):1423-1427.
Crossref - Franklin A, Acar J, Anthony F, et al. Antimicrobial resistance: harmonisation of national antimicrobial resistance monitoring and surveillance programmes in animals and in animal-derived food. RevSci Tech Off Int Epiz. 2001;20 (3):859-866.
Crossref - Ibrahim DR, Dodd CER, Stekel DJ, Ramsden SJ, Hobman JL. Multidrug resistant, extended spectrum b-lactamase (ESBL)-producing Escherichia coli isolated from a dairy farm.FEMS Microbiol Ecol. 2016;92(4):fiw013.
Crossref - Schmid A, Hormansdorfer S, Messelhausser U, Kasbohrer A, Sauter-Louis C, Mansfeld R. Prevalence of extended-spectrum b-lactamase-producing Escherichia coli on Bavarian dairy and beef cattle farms, Appl Environ Microbiol. 2013;79(9):3027-3032.
Crossref - Murugan MS, Sinha DK, Kumar ORV, et al. Epidemiology of carbapenem-resistant Escherichia coli and first report of blaVIM carbapenemases gene in calves from India. Epidemiol Infect. 2019;147:e159.
Crossref - Amador P, Fernandes R, Prudencio C, Duarte I. Prevalence of Antibiotic Resistance Genes in Multidrug-Resistant Enterobacteriaceae on Portuguese Livestock Manure. J Antibiot. 2019;8(1):23.
Crossref - Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2(2):123-140.
Crossref - de Verdier K, Nyman A, Greko C, Bengtsson B, Antimicrobial resistance and virulence factors in Escherichia coli from Swedish dairy calves. Acta Vet Scand. 2012;54(1):2.
Crossref - Montenegro MA, Bitter-Suermann D, Timmis JK, et al. traT gene sequences, serum resistance and pathogenicity-related factors in clinical isolates of Escherichia coli and other gram-negative bacteria. Microbiol. 1985;131(6):1511-1521.
Crossref - Binns MM, Mayden J, Levine RP. Further characterization of complement resistance conferred on Escherichia coli by the plasmid genes traT of R100 and iss of ColV, I-K94. Infect Immun. 1982;35(2):654-659.
Crossref - Cookson AL, Bennett J, Thomson-Carter F, Attwood GT. Molecular subtyping and genetic analysis of the enterohemolysin gene (ehxA) from Shiga toxin-producing Escherichia coli and atypical enteropathogenic E. coli. Appl Environ Microbiol. 2007;73(20):6360-6369.
Crossref - Lim JY, Yoon JW, Hovde CJ. A brief overview of Escherichia coli O157: H7 and its plasmid O157. J Microbiol Biotechnol. 2010;20(1):5-14.
- Prabhu V, Isloor S, Balu M, Suryanarayana VVS, Rathnamma D. Genotyping by ERIC-PCR of Escherichia coli isolated from bovine mastitis cases. Indian J Biotechnol. 2010;09(3):298-301. http://nopr.niscpr.res.in/handle/123456789/9888
- Dhanashree B, Mallya SP. Molecular typing of Enteropathogenic Escherichia coli from diarrheagenic stool samples. J Clin Diagn Res. 2012;6(3):400-404.
Crossref - Awadallah MA, Ahmed HA, Merwad AM, Selim MA, Occurrence, genotyping, shiga toxin genes and associated risk factors of E. coli isolated from dairy farms, handlers and milk consumers. Vet J. 2016;217:83-88.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.