


ISSN: 0973-7510
E-ISSN: 2581-690X
Helicobacter pylori outer inflammatory protein A (OipA) has been found to associate with inflammation that is similar to cag pathogenicity island (cagPAI). However, the roles of the presence of oipA gene involving inflammatory responses in vivo need to be clarified. We investigated the association of oipA and cagPAI on the expression of pro-inflammatory cyotkine genes (IL-1b and IL-8) in gastric tissues of Thai gastroduodenal patients. We detected the oipA and cagPAI genes in 35.56% and 68.89%, respectively. The oipA “on” status was mostly found (93.75%) in oipA-positive samples. We observed higher levels of IL-1b and IL-8 gene expression in oipA-positive tissues (with “on” status), similar to those with cagPAI-positive tissues. Interestingly, samples positive for both oipA and cagPAI genes showed significantly higher levels of IL-1b and IL-8 gene expression, when compared with tissues single-positive for either oipA or cagPAI, or double-negative for these two genes. We conclude that H. pylori induces IL-1b and IL-8 gene expression via oipA-dependent mechanisms. Furthermore, synergy in the presence of both oipA and cagPAI genes associated with increased IL-1b and IL-8 gene expression levels in gastric tissues, which suggested that oipA plays a critical role in the H. pylori pathogenesis.
H. pylori, oipA, cag PAI, IL-1b, IL-8, inflammation
Helicobacter pylori (H. pylori) is a Gram-negative microaerophilic bacterium and a causative agent in various gastroduodenal diseases in humans. The association between H. pylori infection and gastric cancer led to its classification as a class I carcinogen 1. Infection with this bacterium has been found to stimulate of various pro-inflammatory cytokines from gastric cells such as interleukin-1 (IL-1), IL-6 and IL-8, that is play important roles in gastric inflammation 2. The most important H. pylori virulence factor is the cag pathogenicity island (cagPAI) which encodes a type 4 secretory system (T4SS) and immunodominant antigen of CagA. Strains expressing the cagPAI have been associated with a more severe inflammatory response than that induced by cagPAI-negative strains. H. pylori has been reported to depend upon both T4SS- and CagA-dependent mechanisms to induce IL-8 production in gastric cells, resulting in increased inflammation3,4. It has also been shown that studies of gastric biopsies from patients infected with H. pylori cag-positive strains induce significantly more IL-1a and IL-1b than do cag-negative strains 5. Therefore, several studies have suggested that the H. pylori cagPAI is a putative marker for virulent strains of H. pylori that are associated with severe gastroduodenal diseases, especially gastric adenocarcinoma6,7. Interestingly, however, some IL-1b and IL-8 responses have also been found to be independent of the cagPAI mechanisms8,9. Therefore, we speculated that several H. pylori virulence factors may be involved together in promoting inflammation and disease severity.
Bacterial adherence to gastric epithelial cells is a critical step in H. pylori pathogenesis. H. pylori contains a large family of outer membrane proteins (OMPs), comprising 32 members involved in adhesion, immunostimulatory functions and micropore formation10,11. The outer inflammatory protein A (OipA), or HopH, encodes one of the outer membrane proteins, with the oipA gene located approximately 100 kb from the cag PAI in the H. pylori genome12. Aside from promoting bacterial adherence to gastric epithelial cells, OipA (status “on”) increases inflammatory responses by inducing IL-8 production and regulating its secretion through the PI3/Akt pathway in these cells13. Using purified OipA protein was shown to cause cell cytotoxicity and apoptotic cell death by modulation of Bax/Bcl-2 levels in the AGS human gastric epithelial cell line, indicating that H. pylori OipA may play a pathogenic role by damaging host cells14,15. However, previous studies on the role of OipA-induced inflammation have been reported and revealed that mutagenesis of the H. pylori oipA gene did not alter IL-8 production in gastric cells in vitro nor in animal model studies16,17. Currently, the effect of H. pylori OipA on inflammatory responses of IL-8 is still controversial, in particular its role in vivo. Additionally, the association of OipA with the expression of IL-1b has not yet been examined. We hypothesized that H. pylori OipA may associate with pro-inflammatory responses by regulating the expression of IL-1b and IL-8 in the human gastric cells in vivo. To investigate this question, the purpose of the current study was to determine the association between the oipA and cagPAI gene status of H. pylori infections with IL-1b and IL-8 gene expression levels in the gastric tissue samples from Thai subjects with gastroduodenal disease (gastritis and peptic ulcer disease).
Patients and Sample Collection
Gastric tissues were obtained from subjects undergoing gastroduodenal examination at the unit of endoscopy medicine, Supprasittiprasong Hospital, Ubon Ratchathani, Thailand. H. pylori infection status of the subjects was determined by the Rapid Urease Test kit (RUT). A total of 45 gastric tissues were collected for this study from subjects with confirmed gastritis (GT=35) and peptic ulcer disease (PUD=10). Ethic committee approval was received for this study from The Institutional Review Board of Mahidol University (COA.NO. 2016/157.0912). Written inform consent was obtained from patients who participated in this study.
Genomic DNA Extraction and PCR
Genomic DNA was extracted from RUT positive gastric tissues using DNA purification kits (RBC Real Genome, RBCBioscience), according to the manufacturer’s protocol. Isolated DNA was solubilized by Tris-EDTA (TE) buffer and kept at -20oC until polymerase chain reaction (PCR) was performed.
PCR was used to detect H. pylori using primers specific for 16SrRNA, oipA, and cagPAI. All primer sequences were selected from published data with slight modifications, as shown in Table 118-20. The PCR were prepared in a volume of 25 µl using a ready-to-use PCR master mix (OnePCR Ultra®, Bio-Helix Co., LTD. Taiwan), containing 0.5 µM of each primer. PCR amplifications were performed in an automated thermal cycler (BioRad T100TM, USA) with the following cycle conditions: 16SrRNA: 35 cycles of 30 s at 94°C, 30 s at 60°C, and 30 s at 72°C; cagPAI: 35 cycles of 1 min at 94 °C, 1 min at 57 °C, and 1 min at 72°C; and oipA: 35 cycles of 1 min at 94°C, 1 min at 57°C, and 1 min at 72°C. PCR products were analyzed on 1.5% agarose gels and visualized under a UV illuminator. H. pylori oipA amplicons were subsequently sequenced to determine their functional status by observing the stop codons of a signal-peptide coding region 12.
Table (1):
Primers used in the study.
Genes |
Primer sequences |
|---|---|
16SrRNA |
F: GCGACCTGCTGGAACATTAC |
R: CGTTAGCTGCATTACTGGAGA |
|
oipA |
F: GTTTTTGATGCATGGGATTT |
R: GTGCATCTCTTATGGCTTT |
|
cagPAI |
F: ACATTTTGGCTAAATAAACGCTG |
R: TCTCCATGTTGCCATTATGCT |
|
IL-1β |
F: GCACGATGCACCTGTACGAT R: CACCAAGCTTTTTTGCTGTGAGT |
IL-8 |
F: ACTGAGAGTGATTGAGAGTGGAC |
R: AACCCTCTGCACCCAGTTTTC |
|
18SrRNA |
F: CGGCGACGACCCATTCGAAC |
R: GAATCGAACCCTGATTCCCCGTC |
RNA Extraction and mRNA Quantification
Total RNA was extracted from RUT positive tissues using RiboZolTM (Amresco, VWR Company, USA), according to the manufacturer’s protocol. RNA pellets were solubilized with RNAse-free distilled water. The RNA yield and quality were measured by spectrophotometer. RNA (1 µg) was reverse transcribed using SuperScriptTM III Reverse Transcriptase (Invitrogen), as described by the manufacturer. IL-1b and IL-8 gene expression were quantified by the SYBR Green assay on a ThermalCycler96® Real-time PCR (Roche Molecular Systems, Inc.) with an initial denaturation at 95°C, followed by 40 cycles of 95°C for 30 s and 60°C for 30 sec, and normalized to 18SrRNA. The primer sequences for IL-1b, IL-8 and 18SrRNA genes are listed in Table 121,22. Gene expression levels were quantified using the DDCT-method.
Data Analysis
All data were performed using GraphPad Prism software (version 5.0) and significance evaluated with the Student’s t-test. A value of p < 0.05 was considered statistically significant.
As shown in table 2, 45 RUT positive samples were confirmed to harbor H. pylori infection by PCR amplification. A 401-bp PCR product, indicating the presence of the oipA gene, was obtained in 16 (35.56%) of these subjects. No significant differences between the gastritis and PUD were detected. Subsequently, the oipA functional status (“on” / “off”) was determined by PCR-based sequencing. We found that in 15 of 16 (93.75%) subjects, the oipA status was “on”.
Table (2):
The prevalence of oipA and cagPAI genes in Thai subjects with gastroduodenal disease.
| Genes | Patient groups | Total (n=45) |
|
|---|---|---|---|
| Gastritis (n=35) |
PUD (n=10) |
||
| oipA+ | 13 (37.14%) | 3 (30%) | 16 (35.56%) |
| oipA+ status on | 12 (92.31%) | 3 (100%) | 15 (93.75%) |
| cag PAI+ | 22 (62.86%) | 9 (90%) | 31 (68.89%) |
The cagPAI-positive samples was found 68.89% (31/45) of gastric tissues of Thai gastroduodenal subjects (Table 2). The vital components of cagPAI, cagA and cagE, were also found in the cagPAI positive samples (data not shown). There were no significant differences between the gastritis and PUD. The frequency distributions of the combined genotypes of H. pylori cagPAI and oipA in the 45 samples of Thai subjects are presented in table 3. We clustered a total 4 different cagPAI / oipA genotype combinations, out of which the most prevalent genotypes was cagPAI+ / oipA–, but the genotype of cagPAI–/oipA– was not found in PUD.
Table (3):
Distribution of H. pylori cagPAI and oipA combined genotypes in Thai subjects with gastroduodenal disease.
| Gene combinations | Patient groups | Total (n=45) |
|
|---|---|---|---|
| Gastritis (n=35) |
PUD (n=10) |
||
| cagPAI– / oipA– | 10 (28.57%) | 0 (0%) | 10 (22.22%) |
| cagPAI+ / oipA– | 12 (34.29%) | 7 (70%) | 19 (42.22%) |
| cagPAI– / oipA+ | 3 (8.57%) | 1 (10%) | 4 (8.89%) |
| cagPAI+ / oipA+ | 10 (28.57%) | 2 (20%) | 12 (26.67%) |
In order to study the association of H. pylori oipA and cagPAI genes with inflammatory responses, we determined IL-1b and IL-8 gene expression levels in the gastric tissues of Thai subjects with gastroduodenal disease. As shown in fig. 1 and 2, IL-1b and IL-8 gene expression levels in samples containing single-positive gene status of cagPAI (cagPAI+/oipA–) were significantly higher than in those containing double-negative genes (cagPAI–/oipA–) (p<0.05). Similarly, samples containing single-positive oipA gene status (cagPAI–/oipA+) were found to have significantly increased IL-1b and IL-8 gene expression levels when compared with those with a double-negative gene status (cagPAI–/oipA–) (p<0.05), however, the levels were slightly lower than for the cagPAI+ samples (cagPAI+/oipA–). Our study, only one sample carrying oipA-functional “off” status was combined with the cagPAI-negative samples that showed the decreasing of IL-1b and IL-8 gene expression. Interestingly, we found that samples with double-positive gene status for oipA and cagPAI (cag PAI+/oipA+), had higher IL-1b and IL-8 levels compared with single-positive and double-negative samples. These results were consistent for subjects with gastritis and PUD.
H. pylori has been shown to stimulate the production of various pro-inflammatory cytokines from gastric epithelial cells 23. IL-1b is a pro-inflammatory cytokine that has been involved in pain, inflammation and autoimmune condition 24. The biological effects of IL-1b is involved to induce expression of many cytokine genes by either initiating their transcription or stabilizing their mRNA, including tumor necrosis factor a (TNF-a), IL-2, IL-6, IL-12, interferon a, b, and g, granulocyte-colony stimulating factor and macrophage-colony stimulating factor 25. The in vitro study revealed that H. pylori cagA and cagE mutant strains showed a marked decrease in the induction of IL-1b production of human peripheral blood mononuclear cells. It was suggested that the components of cagPAI more frequently associated with the production of IL-1b 26 , especially H. pylori requires the expression of cagE for the full induction of IL-1, IL-12, and TNF- a in the immature dendritic cells 27. However, the molecular mechanisms of H. pylori lipopolysaccharide (LPS) and vacuolating cytotoxin A (VacA) induced IL-1b have been determined 9,28. Thus, based on the previous data, we assumed that other components of H. pylori might be associated with the production of IL-1b.

Fig. 1. IL-1β geneexpression levels in the presence/absence of oipA and cagPAI genes in gastric tissues of subjects with gastritis (A) and PUD (B).
asignificant differences when compared with double-negative cagPAI / oipA gene status (p<0.05).
bsignificant differences when compared with single-positive oipA, cagPAI gene status, or double-negative for these two genes (p<0.05).
IL-8 is also a pro-inflammatory cytokine and is well known to be a chemoattractant for neutrophils and an activator of macrophages that plays important role in pro-inflammatory responses. This chemokine is also associated with cell proliferation, migration and survival of endothelial cells, thereby potentiating the epithelial-mesenchymal transition and survival of cancer cells, required for cancer progression 29. IL-8 has been found to play a dominant role in gastric epithelial cells exposed to H. pylori infection and in pathological processes leading to gastroduodenal disease 30. It was shown that H. pylori can also stimulate gastric epithelial cells to produce IL-8 through a CagA-dependent mechanism. In addition to the CagA protein, bacterial cell wall components, such as peptidoglycan and heptose-1,7-bisphosphate, are also translocated via the T4SS into host epithelial cells and recognized by the nucleotide-binding oligomerization domain receptor (NOD) and the TRAF-interacting protein with forkhead-associated domain (TIFA), respectively, to inducs NF-kB activation and up-regulation of pro-inflammatory immune responses 31,32. Although previous studies reported that H. pylori cagPAI mutant strains induced reduced levels of IL-8 secretion in vitro, other H. pylori components may also be involved 33. This suggests that although the cagPAI may promote clinical disease, other factors important for H. pylori pathogenicity remain to be investigated 34.
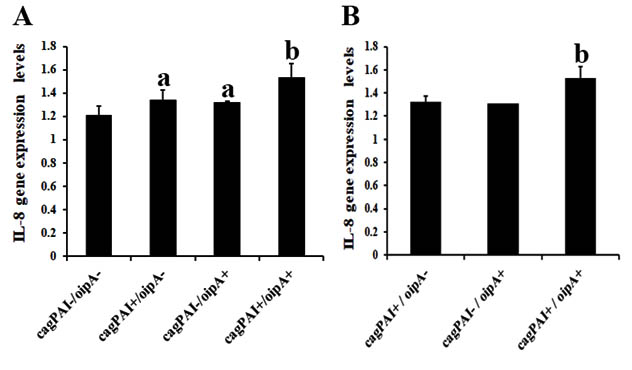
Fig. 2. IL-8 gene expression levels in the presence/absence of oipA and cagPAI genes in gastric tissues of subjects with gastritis (A) and PUD (B)
asignificant differences when compared with double-negative cagPAI / oipA gene status (p<0.05).
bsignificant differences when compared with single-positive oipA, cagPAI gene status, or double-negative for these two genes (p<0.05).
Approximately 4% of the H. pylori genome is composed of genes encoding OMPs. Among them, OipA has emerged as another virulence factor involved in promoting PUD and gastric cancer 12. Previous in vitro study showed that IL-8 production was significantly reduced in epithelial cells stimulated with H. pylori oipA mutant strains on a cag-positive background, indicating the important role of OipA in IL-8 production12 Conversely, OipA-positive strains on a cag-negative background, did not induce higher IL-8 amounts than OipA-negative strains. These results suggest that the OipA protein alone is not able to stimulate IL-8 secretion in AGS cells, but that OipA expression seems to support Cag-induced cytokine secretion 35. However, the effect of OipA on IL-8 secretion in vitro might not be sufficient to explain the exact role of OipA under physiological conditions in vivo. Additionally, the association of OipA on the expression of IL-1b has not been extensively studied. Thus, we evaluated the association of the oipA gene, in the presence or absence of the cagPAI, on IL-1b and IL-8 gene expression levels in gastric tissues of subjects with gastroduodenal disease. We found that IL-1b and IL-8 gene expression levels in gastric tissues was dependent on the cagPAI gene (cagPAI+/oipA–). We also found that tissue samples positive for the oipA gene (cagPAI–/oipA+) also exhibited higher levels of IL-1b and IL-8 gene expression, when compared with those double-negative for the oipA and cagPAI genes. Our data indicated that increasing levels of IL-1b and IL-8 gene expression might be associated with the oipA gene. Additionally, the highest levels of IL-1b and IL-8 gene expression were found in the tissue samples that were positive for both oipA and cagPAI, when compared to single-positive gene and double-negative genes of oipA and/or cagPAI. It is possible that the association of oipA and cagPAI might be associated with heightened levels of IL-1b and IL-8 gene expression in gastric tissues. The results of the present study are consistent with the findings of previous in vitro studiy which showed that H. pylori oipA mutant bacteria displayed reduced levels of cell adherence and induced less IL-8 secretion in the AGS cell line. Furthermore, these mutant bacteria were unable to translocate CagA into gastric epithelial cells 36. Taken together, these data suggest that H. pylori OipA might involve in bacterial adherence leading to IL-1b and IL-8 production through direct signaling and that, moreover, this OMP might co-operate with the cagPAI to induce inflammation.
Our study investigate that H. pylori oipA gene associates with increased IL-1b expression in gastric tissues of subjects with gastroduodenal disease, as well as IL-8 expression. Our findings conclude that IL-1b and IL-8 gene expression levels were dependent on the H. pylori oipA gene. In addition, synergistic effects of oipA combined with cagPAI have been found to associate with increased levels of IL-1b and IL-8 gene expression. These data suggest that OipA might be an important H. pylori virulence factor associated with inflammation. However, the molecular mechanisms of oipA-induced IL-1b expression should be further determined.
ACKNOWLEDGMENTS
This study was supported by the grant of Thailand research Fund (MRG5980069). We kindly thank the staff at Endoscopy Medicine, Sapprasittiprasong Hospital, for gastric tissue collection. We acknowledge Dr. Richard L. Ferrero (Hudson Institute of Medical Research) for review and English editing. Finally, we would like to thank Central Laboratory, Amnat Charoen Campus, Mahidol University, for the instruments used in this study.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
- NIH Consensus Conference. Helicobacter pylori in peptic ulcer disease. NIH Consensus Development Panel on Helicobacter pylori in Peptic Ulcer Disease. JAMA 1994;272(1):65-69.
- Noach LA, Bosma NB, Jansen J, Hoek FJ, van Deventer SJ, Tytgat GN. Mucosal tumor necrosis factor-alpha, interleukin-1 beta, and interleukin-8 production in patients with Helicobacter pylori infection. Scand J Gastroenterol 1994;29(5):425-429.
- Kusters JG, van Vliet AH, Kuipers EJ. Pathogenesis of Helicobacter pylori infection. Clin Microbiol Rev 2006;19(3):449-490.
- Tegtmeyer N, Lind J, Schmid B, Backert S. Helicobacter pylori CagL Y58/E59 mutation turns-off type IV secretion-dependent delivery of CagA into host cells. PLoS One 2014;9(6):e97782.
- Peek RM, Jr., Miller GG, Tham KT, Perez-Perez GI, Zhao X, Atherton JC, et al. Heightened inflammatory response and cytokine expression in vivo to cagA+ Helicobacter pylori strains. Lab Invest 1995;73(6):760-770.
- Figura N, Guglielmetti P, Rossolini A, Barberi A, Cusi G, Musmanno RA, et al. Cytotoxin production by Campylobacter pylori strains isolated from patients with peptic ulcers and from patients with chronic gastritis only. J Clin Microbiol 1989;27(1):225-226.
- Blaser MJ, Perez-Perez GI, Kleanthous H, Cover TL, Peek RM, Chyou PH, et al. Infection with Helicobacter pylori strains possessing cagA is associated with an increased risk of developing adenocarcinoma of the stomach. Cancer Res 1995;55(10):2111-2115.
- Beswick EJ, Bland DA, Suarez G, Barrera CA, Fan X, Reyes VE. Helicobacter pylori binds to CD74 on gastric epithelial cells and stimulates interleukin-8 production. Infect Immun 2005;73(5):2736-2743.
- Luo JJ, Li CY, Liu S, Yu W, Tang SY, Cai HL, et al. Overexpression of Helicobacter pylori VacA N-terminal fragment induces proinflammatory cytokine expression and apoptosis in human monocytic cell line through activation of NF-kappaB. Can J Microbiol 2013;59(8):523-533.
- Tomb JF, White O, Kerlavage AR, Clayton RA, Sutton GG, Fleischmann RD, et al. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 1997;388(6642):539-547.
- Alm RA, Bina J, Andrews BM, Doig P, Hancock RE, Trust TJ. Comparative genomics of Helicobacter pylori: analysis of the outer membrane protein families. Infect Immun 2000;68(7):4155-4168.
- Yamaoka Y, Kwon DH, Graham DY. A M(r) 34,000 proinflammatory outer membrane protein (oipA) of Helicobacter pylori. Proc Natl Acad Sci U S A 2000;97(13):7533-7538.
- Tabassam FH, Graham DY, Yamaoka Y. Helicobacter pylori-associated regulation of forkhead transcription factors FoxO1/3a in human gastric cells. Helicobacter 2012;17(3):193-202.
- Teymournejad O, Mobarez AM, Hassan ZM, Noori S, Moazzeni SM, Khoramabadi N. Cloning, Expression, Purification and Toxicity Evaluation of Helicobacter pylori Outer Inflammatory Protein A. Indian J Microbiol 2013;53(4):391-394.
- Teymournejad O, Mobarez AM, Hassan ZM, Talebi Bezmin Abadi A. Binding of the Helicobacter pylori OipA causes apoptosis of host cells via modulation of Bax/Bcl-2 levels. Sci Rep 2017;7(1):8036.
- Dossumbekova A, Prinz C, Mages J, Lang R, Kusters JG, Van Vliet AH, et al. Helicobacter pylori HopH (OipA) and bacterial pathogenicity: genetic and functional genomic analysis of hopH gene polymorphisms. J Infect Dis 2006;194(10):1346-1355.
- Franco AT, Johnston E, Krishna U, Yamaoka Y, Israel DA, Nagy TA, et al. Regulation of gastric carcinogenesis by Helicobacter pylori virulence factors. Cancer Res 2008;68(2):379-87.
- Gramley WA, Asghar A, Frierson HF, Jr., Powell SM. Detection of Helicobacter pylori DNA in fecal samples from infected individuals. J Clin Microbiol 1999;37(7):2236-2240.
- Kauser F, Hussain MA, Ahmed I, Ahmad N, Habeeb A, Khan AA, et al. Comparing genomes of Helicobacter pylori strains from the high-altitude desert of Ladakh, India. J Clin Microbiol 2005;43(4):1538-1545.
- Akopyants NS, Clifton SW, Kersulyte D, Crabtree JE, Youree BE, Reece CA, et al. Analyses of the cag pathogenicity island of Helicobacter pylori. Mol Microbiol 1998;28(1):37-53.
- Tsai KW, Lai HT, Tsai TC, Wu YC, Yang YT, Chen KY, et al. Difference in the regulation of IL-8 expression induced by uropathogenic E. coli between two kinds of urinary tract epithelial cells. J Biomed Sci 2009;16:91.
- Bhat IA, Naykoo NA, Qasim I, Ganie FA, Yousuf Q, Bhat BA, et al. Association of interleukin 1 beta (IL-1beta) polymorphism with mRNA expression and risk of non small cell lung cancer. Meta Gene 2014;2:123-133.
- Maekawa T, Kinoshita Y, Matsushima Y, Okada A, Fukui H, Waki S, et al. Helicobacter pylori induces proinflammatory cytokines and major histocompatibility complex class II antigen in mouse gastric epithelial cells. J Lab Clin Med 1997;130(4):442-449.
- Ren K, Torres R. Role of interleukin-1beta during pain and inflammation. Brain Res Rev 2009;60(1):57-64.
- El-Omar EM. The importance of interleukin 1beta in Helicobacter pylori associated disease. Gut 2001;48(6):743-747.
- Khamri W, Walker MM, Clark P, Atherton JC, Thursz MR, Bamford KB, et al. Helicobacter pylori stimulates dendritic cells to induce interleukin-17 expression from CD4+ T lymphocytes. Infect Immun 2010;78(2):845-853.
- Galgani M, Busiello I, Censini S, Zappacosta S, Racioppi L, Zarrilli R. Helicobacter pylori induces apoptosis of human monocytes but not monocyte-derived dendritic cells: role of the cag pathogenicity island. Infect Immun 2004;72(8):4480-4485.
- Basak C, Pathak SK, Bhattacharyya A, Mandal D, Pathak S, Kundu M. NF-kappaB- and C/EBPbeta-driven interleukin-1beta gene expression and PAK1-mediated caspase-1 activation play essential roles in interleukin-1beta release from Helicobacter pylori lipopolysaccharide-stimulated macrophages. J Biol Chem 2005;280(6):4279-4288.
- Yuan A, Chen JJ, Yao PL, Yang PC. The role of interleukin-8 in cancer cells and microenvironment interaction. Front Biosci 2005;10:853-865.
- Crabtree JE, Wyatt JI, Trejdosiewicz LK, Peichl P, Nichols PH, Ramsay N, et al. Interleukin-8 expression in Helicobacter pylori infected, normal, and neoplastic gastroduodenal mucosa. J Clin Pathol 1994;47(1):61-66.
- Viala J, Chaput C, Boneca IG, Cardona A, Girardin SE, Moran AP, et al. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat Immunol 2004;5(11):1166-1174.
- Stein SC, Faber E, Bats SH, Murillo T, Speidel Y, Coombs N, et al. Helicobacter pylori modulates host cell responses by CagT4SS-dependent translocation of an intermediate metabolite of LPS inner core heptose biosynthesis. PLoS Pathog 2017;13(7):e1006514.
- Boonyanugomol W, Chomvarin C, Hahnvajanawong C, Sripa B, Kaparakis-Liaskos M, Ferrero RL. Helicobacter pylori cag pathogenicity island (cagPAI) involved in bacterial internalization and IL-8 induced responses via NOD1- and MyD88-dependent mechanisms in human biliary epithelial cells. PLoS One 2013;8(10):e77358.
- Jenks PJ, Megraud F, Labigne A. Clinical outcome after infection with Helicobacter pylori does not appear to be reliably predicted by the presence of any of the genes of the cag pathogenicity island. Gut 1998;43(6):752-758.
- Odenbreit S, Swoboda K, Barwig I, Ruhl S, Boren T, Koletzko S, et al. Outer membrane protein expression profile in Helicobacter pylori clinical isolates. Infect Immun 2009;77(9):3782-3790.
- Horridge DN, Begley AA, Kim J, Aravindan N, Fan K, Forsyth MH. Outer inflammatory protein a (OipA) of Helicobacter pylori is regulated by host cell contact and mediates CagA translocation and interleukin-8 response only in the presence of a functional cag pathogenicity island type IV secretion system. Pathog Dis 2017;75(8).
© The Author(s) 2018. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.