


ISSN: 0973-7510
E-ISSN: 2581-690X
Microbial inoculants are microorganisms including bacteria and fungi that are isolated from their natural habitats, cultured under controlled conditions, and applied to plants or soils to promote specific beneficial outcomes. These biological agents have emerged as an effective agrotechnical approach to benefit sustainable farming practices. Despite the growing number of studies on plant growth-promoting rhizobacteria (PGPR) and fungi (PGPF), a comprehensive synthesis quantifying their global effects and examining moderating factors remains lacking. Existing literature predominantly examines specific regions, crops, or microbial taxa, limiting our general understanding of how inoculant perform under diverse agroecological conditions. To fill this gap, we conducted a systematic review that integrated (i) mapping of research trends in PGPR and PGPF studies and (ii) quantitative synthesis of experimental evidence from 280 published reports across 68 countries. The analysis evaluated the overall impact of microbial on plant performance, soil enzyme activities, and nutrient uptake, while identifying the key moderators influencing these outcomes. Our results showed that most studies were carried out in greenhouse and lab conditions with India, China, Pakistan, Iran and Turkey leading in publication volume (148; 57.1%). Over 90% of the articles processed were indexed by Scopus and/or Web of Science databases. The efficacy microbial inoculants was context-dependent. For example, positive responses to inoculation were more pronounced in herbaceous plants than in shrubs and trees. The efficacy and benefits also varied according to the type of inoculant and the functional group of the host plant.
Microbial Inoculants, Rhizobacteria, Fungi, Plant Growth Promoting Microbes
Recent research statistics have revealed that demand for food has led to intensive non-sustainable agricultural practices particularly dependent on agrochemicals such as pesticides and chemical fertilizers.1-3 Despite the advancements in precision agriculture for optimizing or minimizing fertilizer use, the overuse of chemical fertilizers persists,4 which consequently led to significant negative environmental and social impacts.3 Current challenge in agricultural productivity remains unresolved despite the technological advances.5 This has driven growing interest in testing and applying beneficial microbes as a sustainable solution in agriculture. PGPF and PGPR are important cluster of beneficial, root-colonizing microorganisms and represent the most extensively used in agricultural systems.6-8 PGPF encompass a diverse group of fungi that enhance plant growth and health through various mechanisms, including improved nutrient acquisition, stress tolerance, and disease suppression.9 Among these, Arbuscular Mycorrhizal Fungi (AMF) are the most significant economically and ecologically and the most extensively studied and well-characterized.10 AMF establish symbiotic relationships with approximately 80% of terrestrial plant species and their functional traits are often related to crop performance parameters.11 Through their extensive hyphal networks, they enhance nutrient uptake including phosphorus and improve plant tolerance to various environmental stresses.10,12,13 Fungal-specific mechanisms include also the production of glomalin and the formation of extensive hyphal networks (up to 1000-fold), facilitating access to nutrients in soil microsites otherwise unreachable by plant roots alone. Similarly, PGPR such as Azospirillum, Rhizobia, Bacillus, Pseudomonas, Burkholderia, Pantoea, and Enterobacter promote plant growth through various mechanisms including nitrogen fixation, phosphorus solubilization, production of phytohormones, and induction of systemic resistance against pathogens. They also mitigate abiotic stresses like drought and salinity by producing enzymes such as ACC deaminase, which reduces ethylene levels and enhances plant stress tolerance. Their role in sustainable agriculture is increasingly acknowledged as they offer eco-friendly alternatives to chemical fertilizers and pesticides. Collectively, PGPR and PGPF form an essential component of the soil microbiome and significantly influence both plant performance and soil health. Despite decades of research on both microbial groups, their relative effectiveness and global effects remain unclear. Several factors influencing their effectiveness such as plant type, environmental conditions, and strain specificity have not been systematically evaluated across the literature. The aim of this systematic review is to critically evaluate the effects of fungal and rhizobacterial inoculants on both plant and soil health within the broader context of sustainable agriculture. To address existing knowledge gaps the review synthesizes data from 280 peer-reviewed publications to provide a comprehensive comparison of PGPF and PGPR effects on plant growth, nutrient uptake, and environmental parameters. Specifically, this work (1) assesses the relative effectiveness of AMF and PGPR inoculation on key plant performance indicators, (2) identifies the environmental, biological, and methodological factors that modulate inoculation success, (3) evaluates the consistency of reported effects across different research contexts, and (4) provides evidence-based recommendations for the application of these beneficial microorganisms in sustainable agricultural systems.
Literature search and data compilation
A comprehensive literature search was conducted to identify peer-reviewed research articles addressing the effects of plant growth-promoting microorganisms, including rhizobacteria and fungi, on plant performance and soil parameters. The search was performed across three major scientific databases: Web of Science (Clarivate Analytics), Scopus (Elsevier) and Google Scholar. To ensure comprehensive coverage of the latest research findings, Google Scholar was used in addition to core databases (Figure 1). A range of keywords and search terms was employed to capture the diversity of relevant studies. For bacteria, the search focused on ‘‘rhizobacteria and plant growth promotion’’, ‘‘biological nitrogen fixation’’, ‘‘PGPR-mediated nutrient solubilization (P, K, and Zn)’’, ‘‘crop yield response to PGPR inoculation’’, ‘‘PGPR-based stress mitigation’’, and ‘‘field performance of PGPR inoculation’’. For fungi, the search emphasized ‘‘arbuscular mycorrhizal fungi (AMF) symbiosis’’, ‘‘nutrient uptake enhancement through PGPF inoculation’’, ‘‘mycorrhiza-induced stress tolerance’’, ‘‘soil aggregation and structure’’, ‘‘field performance of PGPF inoculation’’, and ‘‘efficiency of fungal root colonization under field conditions’’. The study screening follows the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines14 (Figure 1).
Study screening and selection
The initial search yielded a total of 317 articles. The inclusion criteria required that articles be peer-reviewed and written in English, and that they present empirical data on plant growth, nutrient uptake, nodulation, or mycorrhizal colonization influenced by the inoculation of plant growth-promoting microorganisms. All experimental studies conducted under controlled laboratory, greenhouse or open-field conditions were considered eligible. Conversely, publications that did not meet these standards were excluded. This included articles published in languages other than English, research with insufficient empirical data, and title/or abstract irrelevant of the topic. After the screening and quality assessment process, a total of 280 studies from 68 countries were retained for inclusion in the final systematic literature review (Figure 1).
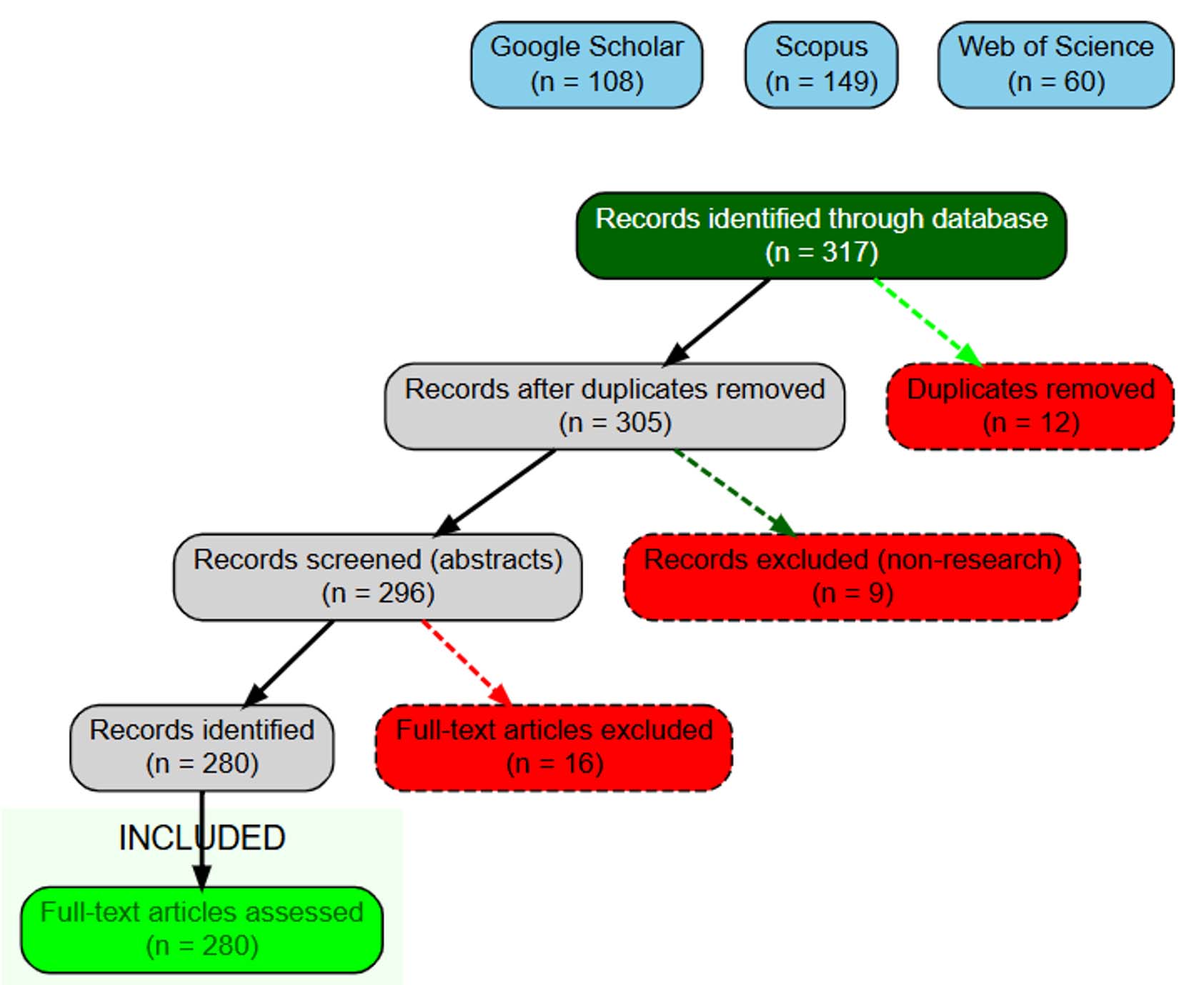
Figure 1. Process of article selection following the PRISMA flowchart of literature search
Data extraction and standardization
For each study, both quantitative and qualitative information were systematically extracted. Specifically, data were collected on plant growth metrics, including total dry biomass or shoot biomass when root data were unavailable. Information on symbiotic characteristics such as root colonization was collected. In addition, nutrient parameters particularly nitrogen (N), phosphorus (P), and potassium (K) concentrations in plant tissues were measured. Finally, contextual experimental details were gathered, including the crop species studied, the type of microbial inoculation applied (single or mixed), and the environmental conditions under which the experiments were conducted, whether in laboratory, greenhouse or field settings. Data from figures were extracted via the Get Data functionality available in Microsoft Excel (Microsoft Corporation, Redmond, WA, USA; version 365; https://getdata.sourceforge.net/download.html). All data were compiled in Microsoft Excel® spreadsheets for standardization and preliminary analysis. Subsequently, further analyses were carried out with the open-source software RStudio. The effect of microbial inoculation on plant biomass and other measured parameters was quantified using the Mean Difference (MD), calculated as the difference between the treatment and control group means.
Quantitative metadata mapping of microbial inoculant research
Geographical distribution of the research and trend of published papers
A total of 317 articles were retrieved and after screening for relevance 280 were reviewed in detail. These articles originated from 68 countries across the world, from 1992–2024 (Figure 2). Most of the studies were performed in Asia (148; 57.1%) with India, China, Pakistan, Iran and Turkey being the most represented. Europe accounted for 42 (16.2%) was mainly represented by Germany, Spain, Italy, France and UK. In contrast, Africa and South America (6.6% and 5.8%, respectively) are contributing but remain underrepresented compared to Asia/Europe. A total of 21 items were not included in this breakdown because the country was not specified in the corresponding studies.
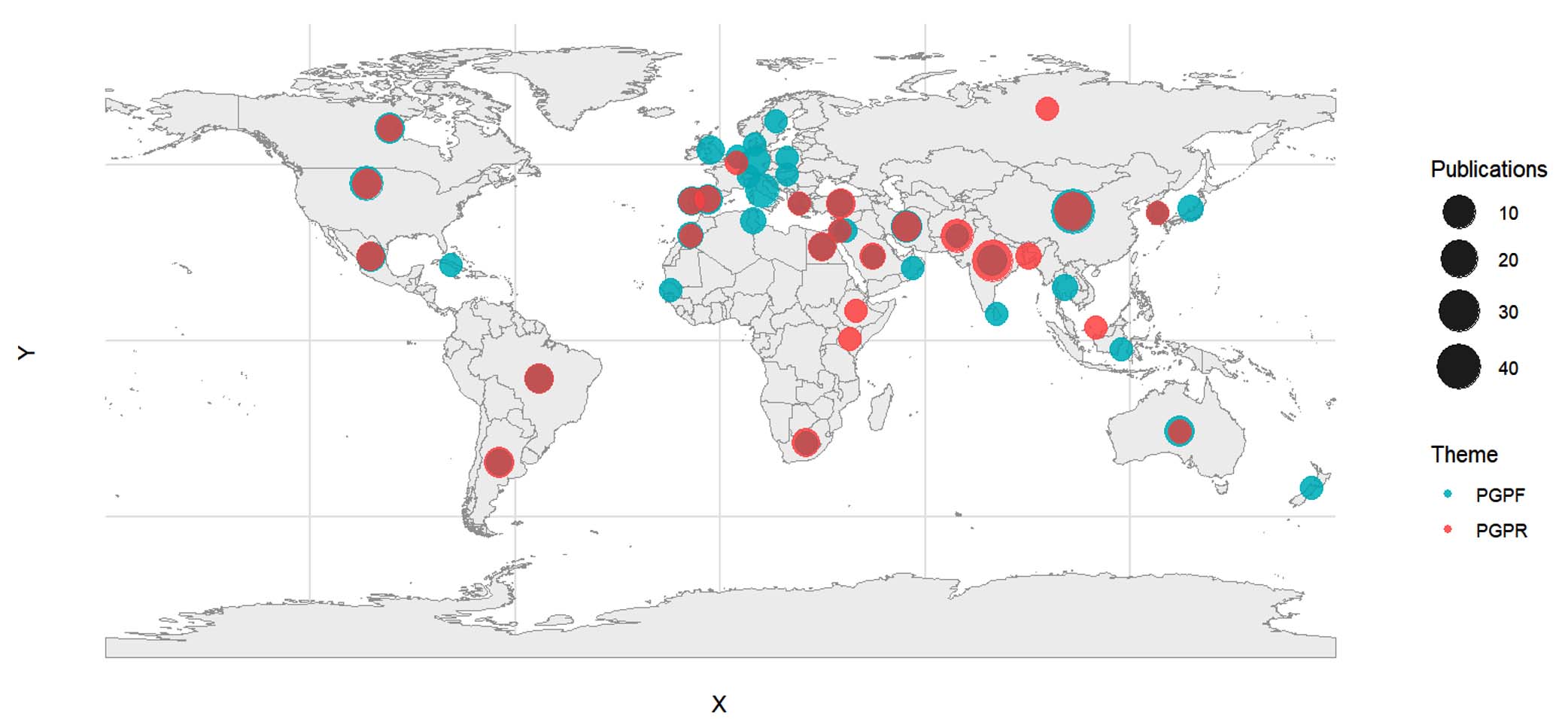
Figure 2. Map of study sites included in the meta-analysis
Journal visibility through major indexing databases
In addition to geographic patterns, the indexing profile of journals publishing microbial inoculant research reveals a strong presence in major scientific databases (Figure 3). Of the journals identified, 127 were indexed in Scopus, 120 in Web of Science (WOS), and 114 indexed in both Scopus and WOS, while only 10 journals were not indexed in either databases.
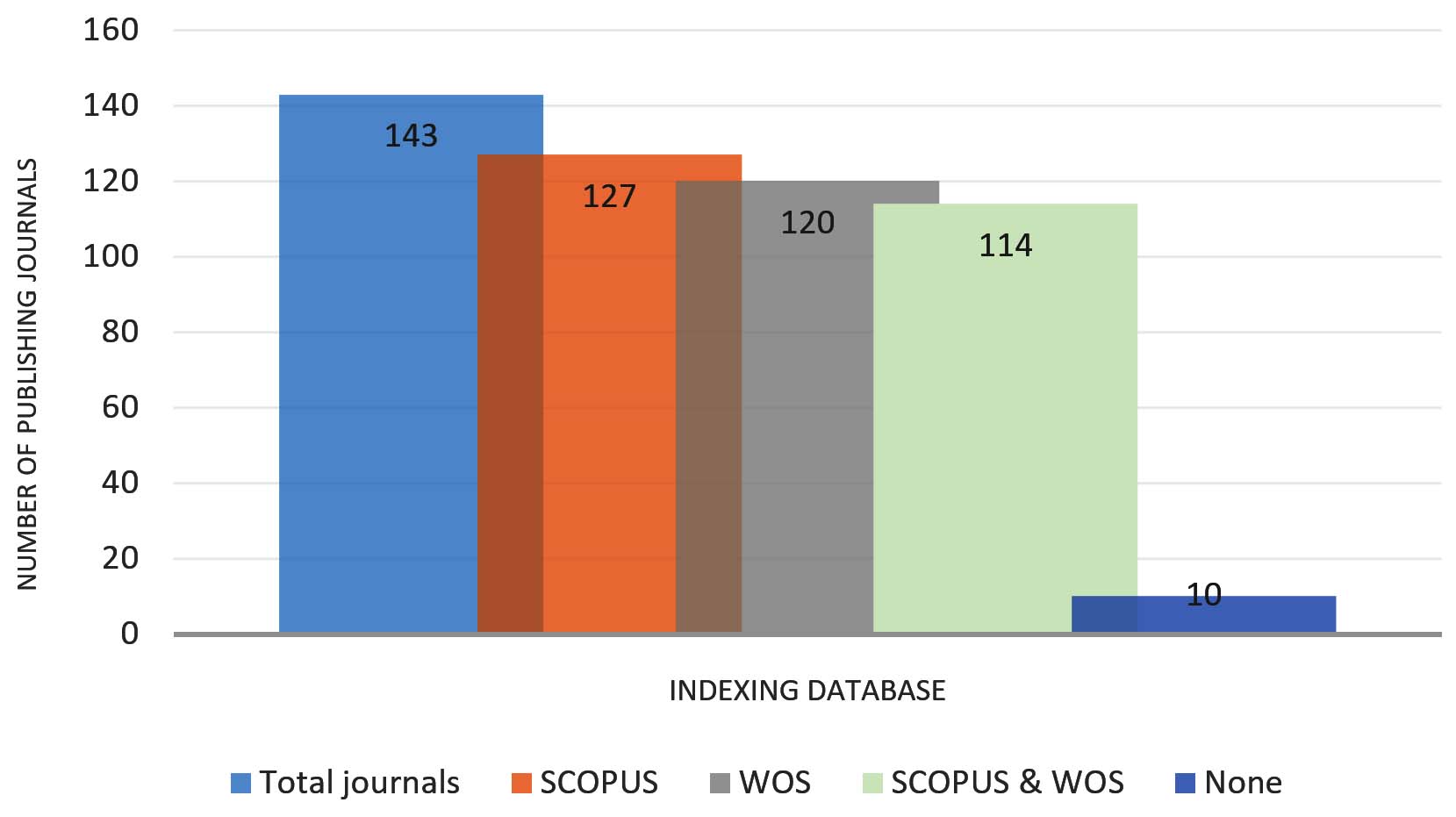
Figure 3. Indexing status of journals publishing microbial inoculant studies
Representation of Plant Families in Inoculant Studies
The analysis revealed that Poaceae were the most represented family, accounting for approximately 35.3%, followed by Solanaceae (15.4%) and Fabaceae (14.9%). Other notable families such as Rosaceae (4.1%), Brassicaceae (3.7%), Asteraceae (3.3%) and Cucurbitaceae (3.3%), appeared less frequently represented. Many families including but not limited to Apiaceae and Salicaceae showed very low representation (less than 1%). Overall, the group of non-legumes is much larger compared to legume represented by Fabaceae (Figure 4).
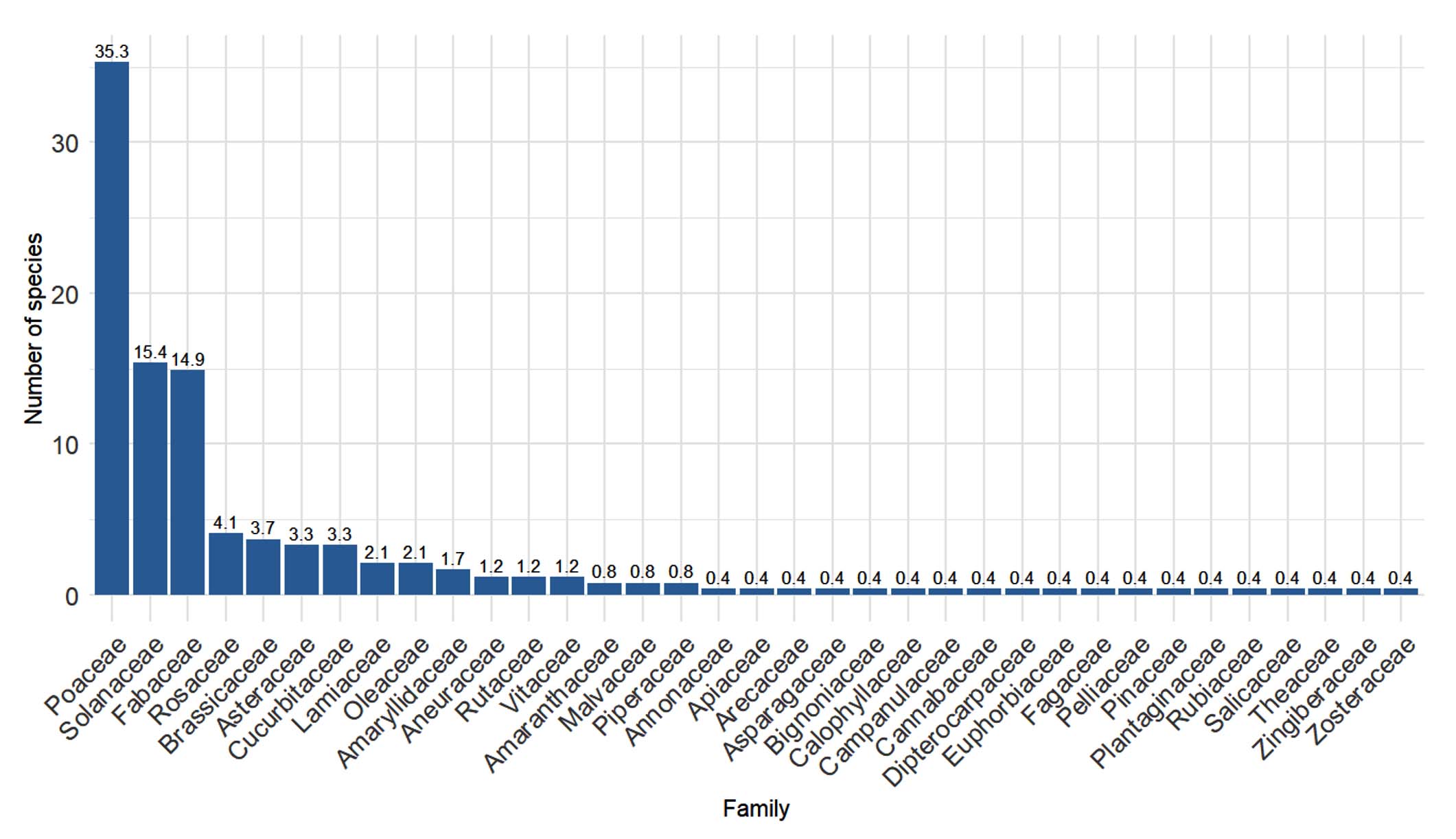
Figure 4. Species Count per Plant Family. The values above the histogram bars represent the percentage of each family
Distribution of experimental conditions, plant types, and strain groups in PGPF and PGPR studies
The systematic review shows that most studies on microbial inoculants for sustainable agriculture have been performed under controlled greenhouse or laboratory conditions (over 76% in both cases). These studies primarily focus on non-leguminous (81.8% for PGPF and 85.7% for PGPR) and herbaceous plants (76% for PGPF and 92.9% for PGPR). Moreover, the results showed the dominance of AMF for PGPF (94.2%) indicates a research concentration on well-known microbial taxa, while other potentially beneficial microbes and underrepresented plant systems (trees, shrubs, and legumes) remain insufficiently explored. For PGPR, studies focus on functions including, biological nitrogen fixation, phosphate solubilisation and others function such as bioremediating inoculants, phytostimulation or organic matter-decomposing inoculant accounting for 44.9% (Figure 5).
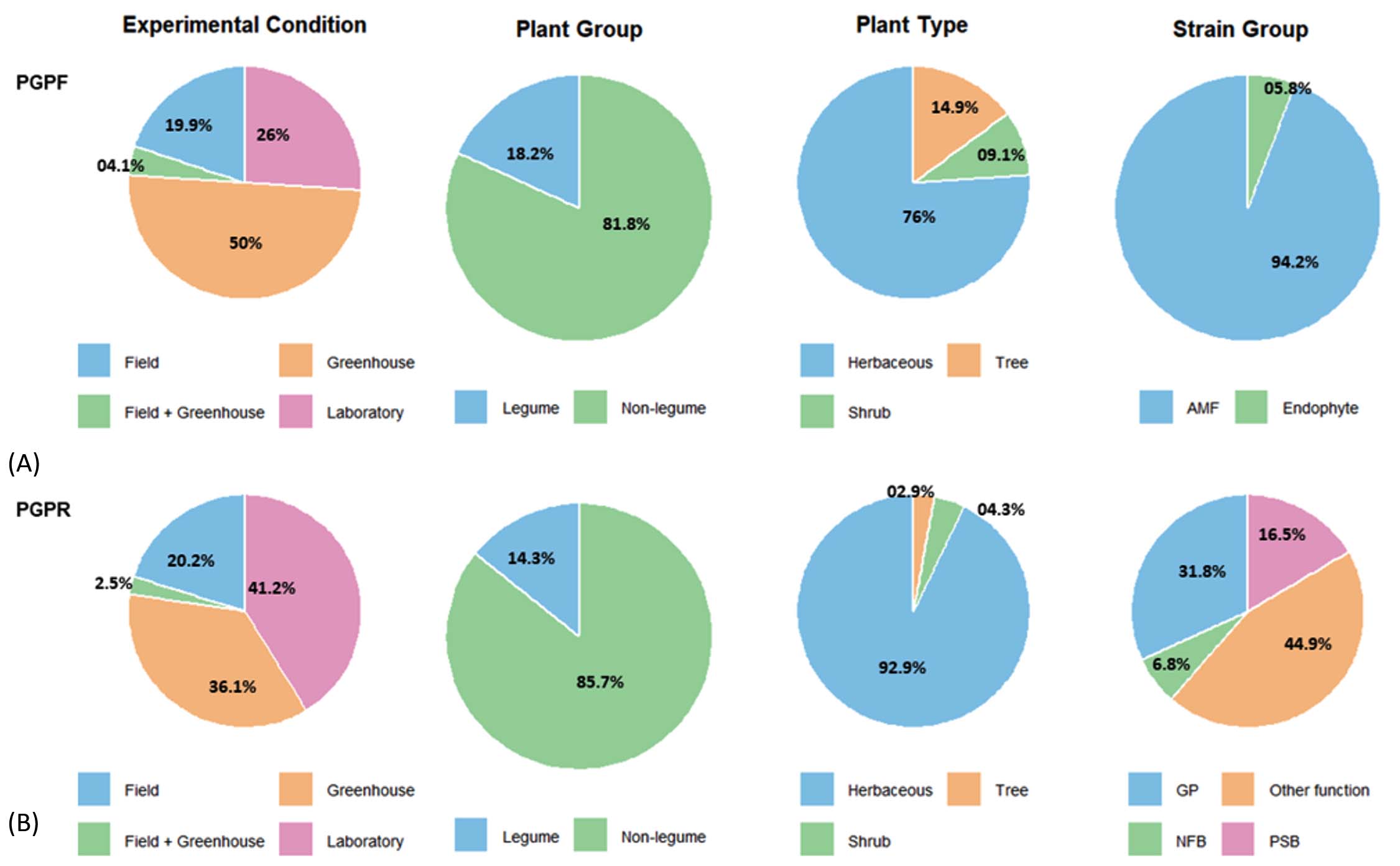
Figure 5. Comparative Analysis of Experimental Conditions and Plant-Strain Groups in PGPF (A) and PGPR (B) studies
Effects of PGPF and PGPR Strain Inoculation on Plant Biomass Production and Nutrient Accumulation
Figure 6 shows two radar plots comparing Control and Strain treatments for two groups: PGPR (A) and PGPF (B). These graphs visualize how each treatment influences various plant growth and nutrient uptake parameters (Figure 6A and B).
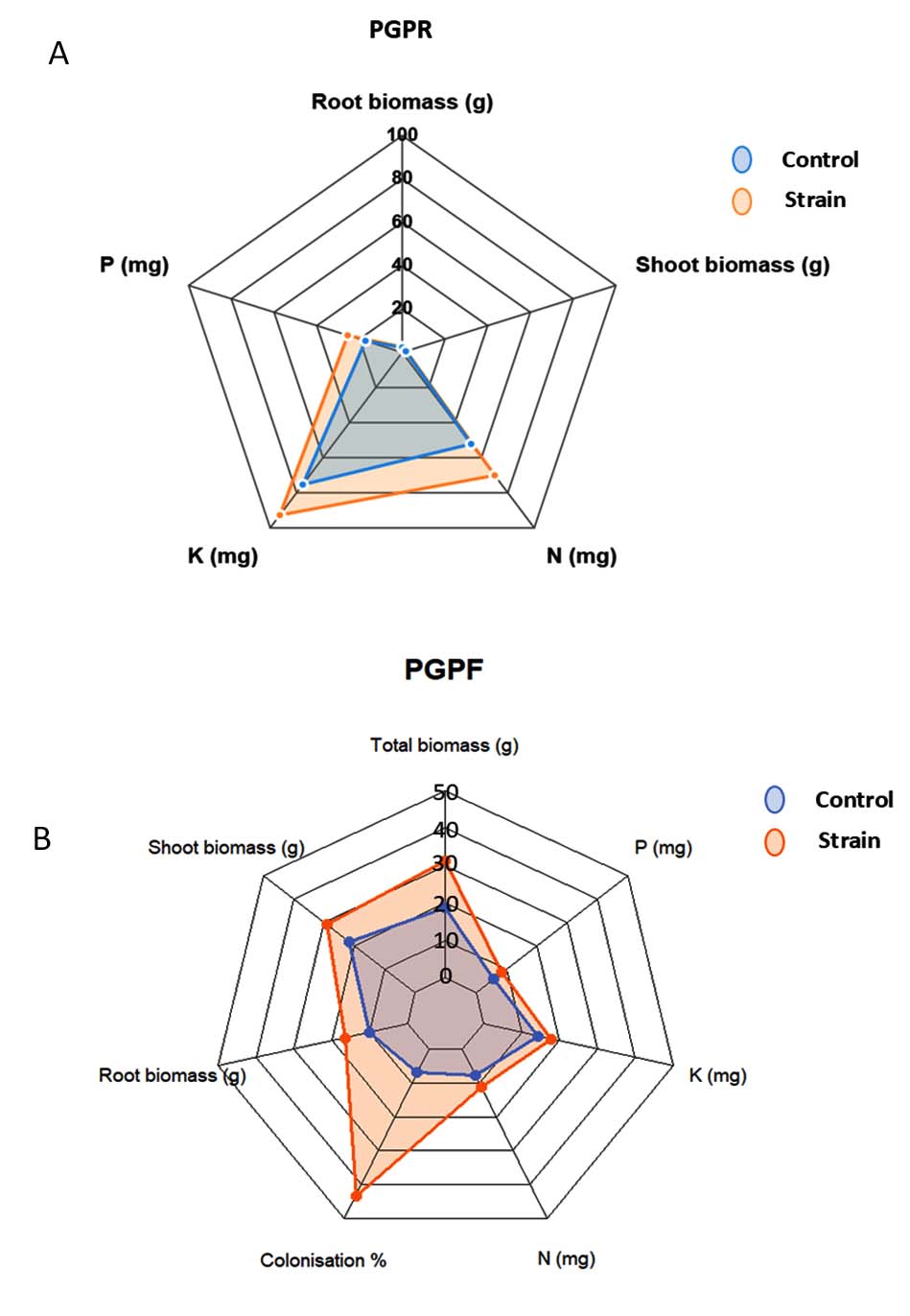
Figure 6. Radar chart – overall PGPR vs control (A) and PGPF vs control (B)
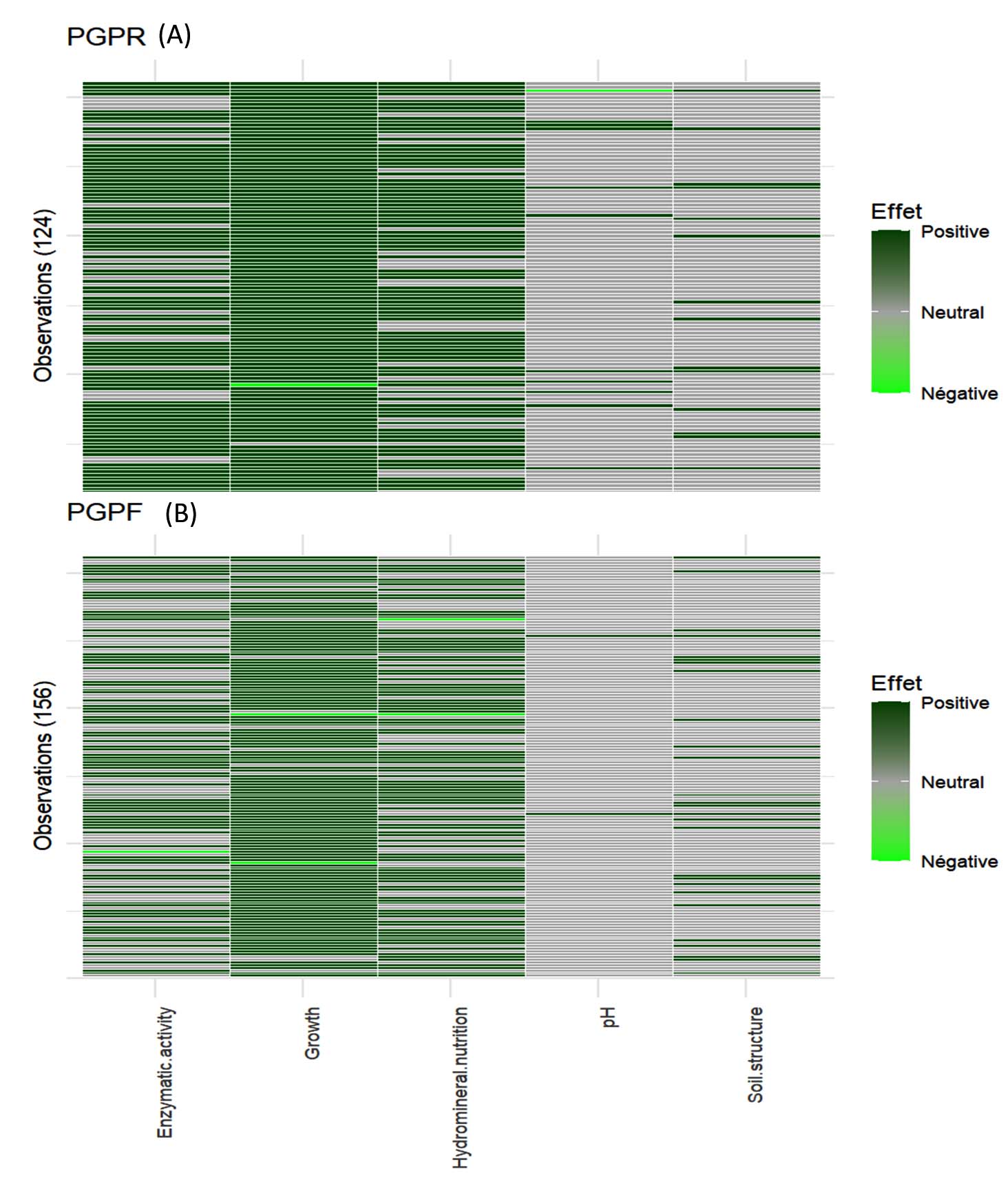
Figure 7. Heatmap effects of PGPR (A) and PGPF (B) inoculation across multiple trials on various environmental and soil parameters
For PGPF, plants inoculated with the strains showed greater growth than the control. Root biomass increased from 10-20 g, total biomass from 20-30 g, and shoot biomass from 21-31 g. The most notable difference in colonization, where the strains show a very high value compared to the Control (45% vs 8%) while N, P, and K show moderate improvements under the Strain from 8, 6, 10 mg to 15, 12 and 14 mg respectively. Conversely to PGPF, PGPR shows the most pronounced increases in N, P, and K (NPK) levels.
Effects of microbial strain inoculation on environmental and soil parameters
The Figure 7 shows two heatmaps illustrating the effects of microbial inoculations on different environmental and soil parameters. Enzymatic activity, growth, and hydromineral nutrition exhibited the strongest positive responses to PGPR and PGPF inoculation. In contrast, pH and soil structure remained largely unaffected, showing predominantly neutral responses. The PGPF show greater variability compared to PGPR (Figure 7).
The bibliometric analyses showed increasing interest in microbial inoculants during the last 30 years particularly in Asian and European countries. This is probably related to the high food demand, the need for sustainable agriculture and policy & commercial dynamics favouring bio-inputs in large agricultural sector. This trend is consistent with the findings of Sun et al.15 who have reported the rapid growth of biofertilizer industry and research in China. They showed that more than 10,000 products are registered with an annual production exceeding 35 million tons, and a market value of over USD 5.5 billion. Similar trends were reported for India from 1992-2022 by Patil et al.16 According to the latest report by IMARC Group, biofertilizer market was estimated at USD 143 million in 2024 and projected to grow further.17 Comparable growth has also been reported in Europe from January 2010 and February 2024 by Meena et al.18 Previously, Bashan et al.19 have estimated biostimulants at USD 6.00 Billion in 2016 and projected an annual rate of 13.8%, reaching more than USD 14.5 billion by 2023. Our analysis by focussing on different continents, climates, and soil types provide a robust and globally representative foundation for assessing the ecological and agronomic benefits of microbial inoculants effects on soil and plant performance. In addition, the journal indexing analysis indicates that most of publications are indexed in major databases such as Scopus and WOS. This supports that microbial inoculants are quantitatively growing in reputable outlets. This global interest in microbial inoculants may be related to their help in mitigating environmental impacts. Through enhanced enzymatic functions, they promote nutrient cycling, which supports plant growth and balanced mineral nutrition,20,21 which often requires complex analytical approaches to discern structural and functional roles of microbial taxa inhabiting the soil. To investigate these ecological properties, we characterized the assembly and soil functioning contribution of ectomycorrhizal. So, the enhanced root biomass and microbial exudation improve soil aggregation, leading to better water management and carbon stabilization.12,22 Therefore, these processes contribute to mitigating environmental impacts by reducing fertilizer dependence23,24 water dynamics, and adaptation to environmental stress. This study investigated the effects of environmental temperature 5, 15, and 25 °C. However, the limited influence on pH and soil structure suggests that these parameters change slowly and may require long-term management or combined interventions. Although microbial inoculants can induce acidification through secretion of secondary metabolites, these effects are typically confined to the rhizosphere at the microscale (i.e., in the immediate vicinity of roots and microbial colonies), and is therefore often insufficient to significantly alter bulk soil pH. In addition, soil buffering capacity may further mitigate detectable changes. Similar results were reported by Zhao et al.25 who found no significant differences in soil pH between several PGPR treatments and their respective controls. However, the soil structure was noticeably improved especially with PGPF. This observation is consistent to several studies,26,27 which report that AMF modify the rhizosphere by amassing glomalin protein which improve soil structure and increase the absorptive surface area and water holding capacity. The predominance of Poaceae, followed by Solanaceae and then Fabaceae, reflects current patterns in our dataset and is broadly consistent with global trends in inoculant research. Poaceae includes the world’s primary cereals: rice, wheat, maize, barley, sorghum, millet etc. These are the backbone of global food systems. Based on a meta-analysis from 2000-2010, showed the predominance of grasses including maize, rice, and wheat.28 Despite this prominence of cereals, the global trajectory of inoculant research has historically focused on legumes, which are key model systems for studying symbiotic nitrogen fixation and plant-microbe interactions. Legumes are extremely important, not only in agriculture broadly but especially in microbial inoculant research. They serve as key model systems for studying symbiotic nitrogen fixation and plant microbe interactions. Our results are in accordance with those of Peoples et al.29 for every tonne of shoot dry matter produced by crop legumes, the symbiotic relationship with rhizobia is responsible for fixing, on average on a whole plant basis shoots and nodulated roots, who showed that legumes play a major role in supplying N to support cereal production is in the rainfed mixed crop-livestock farming systems of Australia. Legumes have traditionally been the primary focus of inoculant research due to their unique and agriculturally symbiosis with rhizobia that fix biologically nitrogen.30 Additionally, The commercial history of inoculants particularly for rhizobia and the relatively predictable yield and N-benefit responses in many legume species have also concentrated research and product development on Fabaceae.30 By contrast, cereals and other non-legumes are typically the focus of research on free-living or associative diazotrophs and other PGPR, which tend to produce more variable effects and are thus less consistently represented in inoculation studies.31-33
Our multi-year, regional-scale investigation demonstrates that PGPR effects appear slightly more consistent than PGPF, likely due to their broader ecological adaptability and direct mechanisms of action (e.g., auxin production, siderophores, ACC deaminase, etc.). According to Lorenz et al.34 stimulating primary production and improving stress resistance. Climate change has several consequences worldwide that affect arable land and agriculture. Studies on plant-soil-microorganism interactions to enhance plant productivity and/or resistance to abiotic stress may open new perspectives. This strategy aims to make agricultural-relevant plant species able to complete their biological cycle in extreme soils with the help of inoculated or primed plant growth-promoting rhizobacteria Interest in this approach has remarkably increased with the aim of improving plant growth and yield under adverse conditions, such as drought and salinization. In contrast, PGPF often display higher variability in efficacy, potentially attributable to several factors. According to Adedayo & Babalola9 revealing functions that endeavor sustainability of the plants, are commonly referred to as ‘plant-growth-promoting fungi’ (PGPF). This variability is due to complex root-fungus-soil interactions and less fully resolved mechanisms. Most research on both microbial groups remains confined to controlled environments, particularly greenhouses, suggesting a need for more field-based validations to ensure practical applicability in real agricultural systems. Future research should aim to bridge this gap between laboratory or greenhouse findings and field applications to enhance the real-world impact of microbial bioinoculants.
The largest numbers of studies on microbial inoculants were done in Asia followed by Europe. The interest in this topic has increased significantly in the last 15 years, both PGPFs and PGPR inoculants increase plant nutrient content and growth, soil enzymes activities and structure with more pronounced effects of PGPR across the studies analyzed. Currently, most of studies still in control condition where the screening-based PGPR inoculum development mostly relies on specific functions (like auxin production, phosphate solubilization, nitrogen fixation, etc.) of pure culture isolates. Despite these promising findings, the strong geographical concentration of studies in Asia (57.1% of the dataset) highlights a limitation in the global representativeness of the results. Agricultural systems in these regions are often characterized by specific climatic conditions, soil properties, and cropping systems that may not fully reflect those of underrepresented regions such as Africa or Latin America. Given the context-dependent nature of microbial inoculant performance, caution is therefore needed when extrapolating these findings to a global scale.
Future research should focus on field scale in traditional cultivation and development of accessible techniques for inoculants production. Future research should solve the challenges of scaling up to field application efficiencies, as well as regulatory constraints, including stringent biosecurity requirements, high costs and complexity of strain registration, and the lack of harmonized approval frameworks across countries producing and commercializing these biofertilizers.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Kumari M, Swarupa P, Kesari KK, Kumar A. Microbial Inoculants as Plant Biostimulants: A Review on Risk Status. Life. 2022;13(1):12.
Crossref - Kumari P, Singh A, Kharwar RN. Chapter 18—Phytostimulation and ISR responses of fungi. In: Sharma VK, Shah MP, Parmar S, Kumar A, eds. Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology. Elsevier. 2021:459-473.
Crossref - Tilman D, Balzer C, Hill J, Befort BL. Global food demand and the sustainable intensification of agriculture. Proc Natl Acad Sci U S A. 2011;108(50):20260-20264.
Crossref - Brunelle T, Chakir R, Carpentier A, et al. Reducing chemical inputs in agriculture requires a system change. Commun Earth Environ. 2024;5(1):369.
Crossref - Chen B, Zou C, Zhang Y. et al. The current status, opportunities, challenges and coping strategies of sustainable agriculture. Discov Sustain. 2025;6:1282.
Crossref - Basu A, Prasad P, Das SN, et al. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability. 2021;13(3):1140.
Crossref - Bayranvand M, Akbarinia M, Jouzani GS, Gharechahi J, Kooch Y, Baldrian P. Composition of soil bacterial and fungal communities in relation to vegetation composition and soil characteristics along an altitudinal gradient. FEMS Microbiol Ecol. 2021;97(1):fiaa201.
Crossref - Bhullar M. Soil Application of Plant Growth-Promoting Fungi: A Sustainable Strategy For Agriculture. Ann Agric Sci Technol. 2019;8:1-8.
Crossref - Adedayo AA, Babalola OO. Fungi That Promote Plant Growth in the Rhizosphere Boost Crop Growth. J Fungi. 2023;9(2):239.
Crossref - Soumare A, Diop T, Manga A, e t al. Role of arbuscular mycorrhizal fungi and nitrogen fixing bacteria on legume growth under various environmental stresses. Int J Biosci. 2015;7:31-46.
Crossref - Zhang F, Zou D, Wang J, et al. Co-inoculation of rhizobia and AMF improves growth, nutrient uptake, and cadmium resistance of black locust grown in sand culture. Physiol Plant. 2024;176(2):e14205.
Crossref - Biswas D, Chakraborty AK, Srivastava V, Mandal A. Plant Growth Promoting Rhizobacteria (PGPR): Reports on Their Colonization, Beneficial Activities, and Use as Bioinoculant. Adv Agric. 2024;2024(1):8173024.
Crossref - Mohamed RR, Fahim ME, Soliman SMA. Development of hydrogel based on Carboxymethyl cellulose/poly(4-vinylpyridine) for controlled releasing of fertilizers. BMC Chem. 2022;16(1):52.
Crossref - Liberati A, Altman DG, Tetzlaff J, et al. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: explanation and elaboration. BMJ. 2009;339:b2700.
Crossref - Sun X, Xu Z, Zhang N, et al. Biofertilizer Industry and Research Developments in China: A Mini-Review. Microb Biotechnol. 2025;18(5):e70163.
Crossref - Patil M, Ajayakumar, Sagar, Suman L, Nandita B, Priyanka V. Biofertilizers in India: Growth Patterns and Export Competitiveness. Biological Forum – An International Journal. 2024;16(8):256-261
- IMARC Group. 2025. Biofertilizer Market Size, Share, Trends and Forecast by Type, Crop, Microorganism, Mode of Application, and Region, 2026-2034. Biofertilizers Market Size, Share And Trends Report | 2034. Accessed December 2, 2025.
- Meena DC, Birthal PS, Kumara TMK. Biostimulants for sustainable development of agriculture: a bibliometric content analysis. Discov Agric. 2025;3(1):2.
Crossref - Bashan Y, Prabhu SR, de-Bashan LE, Kloepper JW. Disclosure of exact protocols of fermentation, identity of microorganisms within consortia, formation of advanced consortia with microbe-based products. Biol Fertil Soils. 2020;56(4):443-445.
Crossref - Prieto-Rubio J, Garrido JL, Alcantara JM, Azcon-Aguilar C, Rincon A, Lopez-Garcia A. Ectomycorrhizal fungal network complexity determines soil multi-enzymatic activity. SOIL. 2024;10(1):425-439.
Crossref - Soumare A, Diédhiou AG, Arora NK, et al. Potential Role and Utilization of Plant Growth Promoting Microbes in Plant Tissue Culture. Front Microbiol. 2021;12:649878.
Crossref - Wang C, Kuzyakov Y. Rhizosphere engineering for soil carbon sequestration. Trends Plant Sci. 2024;29(4):447-468.
Crossref - Bindeliya, Zhang W, Li H. The Role of Synthetic Root Exudates in Modulating Soil Hydraulic Properties and Strengths Under Temperature Variations. Water. 2025;17(7):1033.
Crossref - Ollio I, Zornoza R, Gallego JC, Egea-Gilabert C, Fernandez JA, Lloret E. Combined microbial inoculation and reduced inorganic fertilization enhances diversity and functionality in potato rhizosphere microbiome at field scale. Chem Biol Technol Agric. 2025;12(1):1-21.
Crossref - Zhao G, Zhu X, Zheng G, et al. Development of biofertilizers for sustainable agriculture over four decades (1980–2022). Geography and Sustainability. 2024;5(1):19-28.
Crossref - Lopes MJ dos S, Dias-Filho MB, Gurgel ESC. Successful Plant Growth-Promoting Microbes: Inoculation Methods and Abiotic Factors. Front Sustain Food Syst. 2021;5.
Crossref - Manivannan S, Umadevi P. Plant Microbe Interface: The Plant Antimicrobial Peptides. In: Varma A, Tripathi S, Prasad R, eds. Plant Biotic Interactions : State of the Art Cham: Springer International Publishing; 2019:253-261.
Crossref - Li J, Wang J, Liu H, Macdonald CA, Singh BK. Application of microbial inoculants significantly enhances crop productivity: A meta-analysis of studies from 2010 to 2020. J Sustain Agric Environ. 2022;1(3):216-225.
Crossref - Peoples MB, Brockwell J, Herridge DF, et al. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis. 2009;48(1):1-17.
Crossref - Bullard GK, Roughley RJ, Pulsford DJ. The legume inoculant industry and inoculant quality control in Australia: 1953–2003. Aust J Exp Agric. 2005;45(3):127-140.
Crossref - Boubekri K, Soumare A, Lyamlouli K, Ouhdouch Y, Hafidi M, Kouisni L. Improving the efficiency of phosphate rocks combined with phosphate solubilizing Actinomycetota to increase wheat growth under alkaline and acidic soils. Front Plant Sci. 2023;14:1154372.
Crossref - Giller KE, James EK, Ardley J, Unkovich MJ. Science losing its way: examples from the realm of microbial N2-fixation in cereals and other non-legumes. Plant Soil. 2025;511(1):1-24.
Crossref - Sharma AK, Dwivedi SV, Devi J, et al. Botany, ethnomedicine, phytochemistry and pharmacology of Amaranthus spp.- a review. South African Journal of Botany. 2025;178:198-216.
Crossref - Lorenz C, Vitale E, Hay-Mele B, Arena C. Plant growth promoting rhizobacteria (PGPR) application for coping with salinity and drought: a bibliometric network multi-analysis. Plant Biol. 2024;26(5):777-788.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.