


ISSN: 0973-7510
E-ISSN: 2581-690X
M. Gnanachitra1
Agro-residues constitute the largest reservoir of renewable carbon source on Earth. While their valorization has primarily focused on cellulose, hemicellulose is the second most abundant group of polysaccharides and remains underexploited. Glycosyl hydrolases (GHs) are key enzymes involved in the depolymerisation of plant biomass, and filamentous fungi are widely recognized as efficient producers of GHs enzymes. In the present study, an in-house isolate, Aspergillus fumigatus PSF1, was used to produce xylanase via solid-state fermentation (SSF) with wheat bran as the sole carbon source. Key process conditions, including moisture percent, inoculum load, temperature, and pH, were initially optimized using the One-Factor-at-a-Time (OFAT) method and subsequently statistically optimized using Response Surface Methodology (RSM). Under optimized conditions, xylanase activity reached 147.96 IU gds-1, a 1.29-fold increase relative to unoptimized conditions. The study demonstrates that systematic optimization of critical fermentation parameters significantly enhances xylanase productivity in SSF by A. fumigatus PSF1. The findings suggest that wheat bran is an effective low-cost substrate and highlight the potential of this fungal strain for scalable, high-yield xylanase production.
Response Surface Methodology, Solid-state Fermentation, Xylanase, Wheat Bran, Waste Valorization
Management of enormous amounts of waste generated by food, industrial, and particularly agricultural sources poses a serious global problem. In developing countries, where agriculture is a primary economic activity, the waste generated is usually burned, dumped, or landfilled.1 Approximately 1.3 billion tons of agricultural waste, valued at around $165 billion annually, are wasted worldwide, accounting for roughly one-third of total food production. 140 billion metric tons of waste originate from agriculture every year, of which half is disposed of in landfills or burned. Agricultural waste disposed of in landfills or burned pollutes ecosystems, emits harmful greenhouse gases, and endangers human health by releasing harmful substances.2
Agricultural residues are a primary renewable energy resource with negligible nutritional value for humans, produced mainly from wheat, rice, pearl millet, sorghum, barley, finger millet, sugarcane, potatoes, tubers, and pulses.3,4 These lignocellulosic biomasses are abundant, low-cost, and rich in cellulose, hemicellulose, and lignin, making them promising feedstocks for diverse bioprocessing applications, including biofuel production, enzyme synthesis via solid-state fermentation (SSF), and animal feed formulation.5 Among various lignocellulosic substrates, wheat bran is frequently reported as an excellent support and carbon source for fungal xylanase production, owing to its xylan content (18%-22%), favourable particle size, porosity, and balanced nutrient composition, which collectively support high enzyme yields under SSF.6 Furthermore, xylan has extensive applications in the production of bio-based products, including packaging materials, hydrogels, ethanol, lactic acid, furfural, etc. Xylan is also used in bioplastic formulations, medical applications, and value-added processes to produce xylitol, xylanase, and xylooligosaccharides, among other products.7,8 Valorization of wheat bran for xylanase and related bioproducts aligns with circular bioeconomy goals, transforming an abundant milling byproduct into a high-value biochemical input.
Xylanases, key hemicellulolytic enzymes that break β-1,4-glycosidic linkages in the xylan polymer, thereby enabling the restyling of biomass into fermentable sugars and value-added products.9,10 Xylanases are classified into GHs families, notably GH10 and GH11, with contributions from GH5, GH8, and GH43. GH11 xylanases are highly substrate-specific and efficient at cleaving β-1,4-glycosidic linkages in xylan, whereas GH10 xylanases often display broader substrate specificity and greater thermostability. The catalytic mechanism typically involves key glutamic and aspartic acid residues, and some xylanases operate via a double-displacement mechanism, thereby retaining the anomeric configuration.11 Galactose, arabinose, and 4-O-methylglucuronic acid are present in the main chain of xylan, which is made up of xylose. These glycosidic linkages can also be of the type β-(1→3), β-(1→6), α-(1→2), α-(1→3), and α-(1→6)12 Filamentous fungi, specifically Aspergillus spp., are widely preferred for xylanase production because of their robust secretory machinery, proliferation on low-cost residues, and the ability to yield extracellular enzyme titers suitable for industrial processes. SSF has gained prominence over submerged cultivation for such fungi as it mimics their natural habitat, enables high volumetric productivity, and can be implemented using inexpensive agricultural byproducts.13,14 However, xylanase productivity under SSF is strongly influenced by multiple interacting factors, including substrate moisture content, initial pH, incubation temperature, inoculum load, and supplementation with nitrogen and mineral sources, which cannot be efficiently optimized by traditional one-factor-at-a-time (OFAT) approaches.
In recent years, statistical designs have been widely applied to optimize xylanase production, enabling quantitative evaluation of factor interactions, reducing the number of experimental runs, and significantly enhancing enzyme titers. Dual step statistical strategies, combining screening and response surface methodology (RSM), have delivered several-fold improvements in xylanase yields in bacterial and fungal systems, highlighting their utility for developing economically viable bioprocesses.9 Nonetheless, there is limited information on the statistically optimized production of xylanase in A. fumigatus under SSF using wheat bran as a substrate despite the species’ recognized lignocellulolytic potential. Employing this strategy, this study was designed to enhance xylanase activity through SSF by optimizing key parameters, including moisture percentage, inoculum load, pH, and temperature, using the in-house A. fumigatus strain.
Substrate processing and culture preparation
Wheat bran was procured from the local wheat mill in Coimbatore. The collected wheat bran was shade-dried for two days to retain its physical and chemical properties. The shade-dried sample was sieved using a 200 µm sieve and stored at room temperature for further studies. A. fumigatus PSF1 was collected from the Biocatalysts Laboratory, Department of Agricultural Microbiology, Tamil Nadu Agricultural University. The culture exhibited a white, floccose mycelial mat that later turned greenish due to profuse conidial sporulation on Potato Dextrose Agar (PDA). The culture was stored at 4 °C on PDA plates and sub-cultured frequently onto fresh PDA plates.15
Substrate preparation and xylanase production
SSF was performed in 90 mm borosilicate glass petri-dishes filled with 10 g of untreated wheat bran supplemented with synthetic medium consisted of 2 gL-1 K2HPO4, 0.5 gL-1 KCl, 0.01 gL-1 FeSO4·7H2O, 0.15 gL-1 MgSO4·7H2O, 7 gL-1, KH2PO4, 1 gL-1 (NH4) SO4, and 1.2 gL-1 yeast extract at 50% moisture content and the pH was adjusted to 4.8 Prepared Petri dishes were autoclaved. Four discs of inoculum per plate were placed on sterile substrate and incubated at 40 °C for 10 days. The produced xylanase was extracted using a citrate-phosphate buffer (pH 3.4) by incubating at 40 °C for 30 min under shaking conditions, followed by filtration.16
Xylanase extraction
The 3,5-dinitrosalicylic acid (DNS) assay was used to measure xylanase activity.17 0.5 mL of enzyme extract was added to 1 mL of substrate (1% xylan in 50 mM sodium phosphate buffer, pH 6.5), mixed thoroughly, and incubated at 50 °C for 30 min. The activity was terminated by adding 1.5 ml of the DNS reagent, followed by boiling in a water bath for 5 min. Absorbance was measured at 540 nm using a UV-Vis spectrophotometer (Spectramax i3x, USA) to quantify the enzyme activity of the resulting solution. One international unit (IU) of xylanase activity was defined as the amount of enzyme that liberates 1 µmol of xylose per minute under the specified assay conditions.
Pre-optimization of process parameters for xylanase production
In the present study, culture and substrate conditions, including moisture percentage, inoculum load, temperature, and pH, were optimized to enhance xylanase production under SSF using an OFAT approach. The effect of moisture percent (60%, 65%, 70%, 75% and 80%), inoculum load (4, 5, 6, 7 and 8 disc per plate), temperature (32 °C, 34 °C, 36 °C, 38 °C, and 40 °C), and pH levels (4.0, 4.5, 5.0, 5.5, and 6.0) on xylanase produced from A. fumigatus PSF1 was studied.
Production optimization of xylanase by response surface methodology (RSM)
The Box-Behnken design (BBD) is well documented for evaluating the effects of individual factors and their interactions on experimental responses with considerably fewer experiments. This approach yields more accurate and reliable results, providing efficient, rapid process conditions by using second-order polynomial equations as response classifiers and highlighting the significance of key variables. The BBD approach drastically reduces the number of trials, making it highly advantageous for optimization.18 Xylanase production under SSF depends critically on variables such as inoculum load, substrate moisture content, temperature, and pH. The optimal response conditions for xylanase production using wheat bran were systematically identified using RSM. Four independent variables, such as moisture percent (%), inoculum load (disc per plate), temperature (°C), and pH, were utilised for optimization. The second-order model used to describe the influence of independent factors on the response variable was presented as
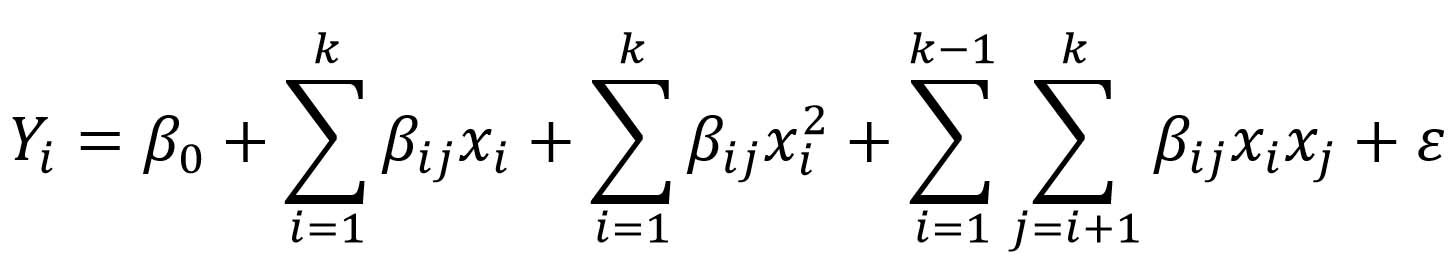 In this model, Yi denotes the output variable; x denotes the independent variable; β0 is the constant term; and βi, βii, and βij denote the linear, quadratic, and interaction regression coefficients, respectively. The β coefficients are calculated by the least-squares method
In this model, Yi denotes the output variable; x denotes the independent variable; β0 is the constant term; and βi, βii, and βij denote the linear, quadratic, and interaction regression coefficients, respectively. The β coefficients are calculated by the least-squares method
(Table 1). The BBD generated 29 experimental runs, comprising four variables, using Design-Expert software 10.0 (Stat-Ease, Inc., USA).19
Table (1):
Independent variables and levels of variation in Box-Behnken design (BBD)
| Study type | Response Surface | Subtype | Randomized | |||
|---|---|---|---|---|---|---|
| Design type | Box-Behnken | 29 Runs | ||||
| Design mode | Quadratic | |||||
| Factor | Name | Units | Type | Minimum | Maximum | Mean |
| A | Moisture percent | % | Numeric | 60 | 80 | 70 |
| B | Inoculum load | disc | Numeric | 4 | 8 | 6 |
| C | temperature | °C | Numeric | 32 | 40 | 36 |
| D | Initial pH | Numeric | 4 | 6 | 5 | |
| Response | Name | obs | Analysis | |||
| R1 | Xylanase | IU gds-1 | 29 | Polynomial | ||
Statistical analysis
Experiments were performed in triplicate; results were expressed as mean ± standard error. The RSM models and optimization procedures are constructed using Design-Expert software 10.0 (Stat-Ease, Inc., USA). Analysis of Variance (ANOVA) was also evaluated using Design-Expert software 10.0.
Pre-optimization of process parameters for xylanase production
Role of moisture percent on xylanase activity
Moisture percent is a crucial determinant in SSF, as excessive moisture alters substrate porosity and particle structure, whereas insufficient moisture increases water tension and reduces nutrient solubility. Therefore, optimizing the moisture content is crucial for maximizing enzyme yield.20 In the current study, the effect of five moisture levels on xylanase activity was evaluated. Xylanase activity elevated with rising moisture and achieved a maximum of 114.14 IU gds-1 at 70 % moisture level, beyond which activity declined significantly (Figure 1). Higher moisture levels limit oxygen diffusion and increase particle clumping, thereby restricting hyphal penetration. Similarly, Khanahmadi et al. reported a peak xylanase activity of 2070 U/g of fermented dry matter at 70% moisture, using wheat bran as the substrate under SSF by A. niger CCUG 3399.14 Yet another study reported maximum xylanase activity of 310 IU g-1 with wheat bran at 70% moisture, supporting the present findings.21
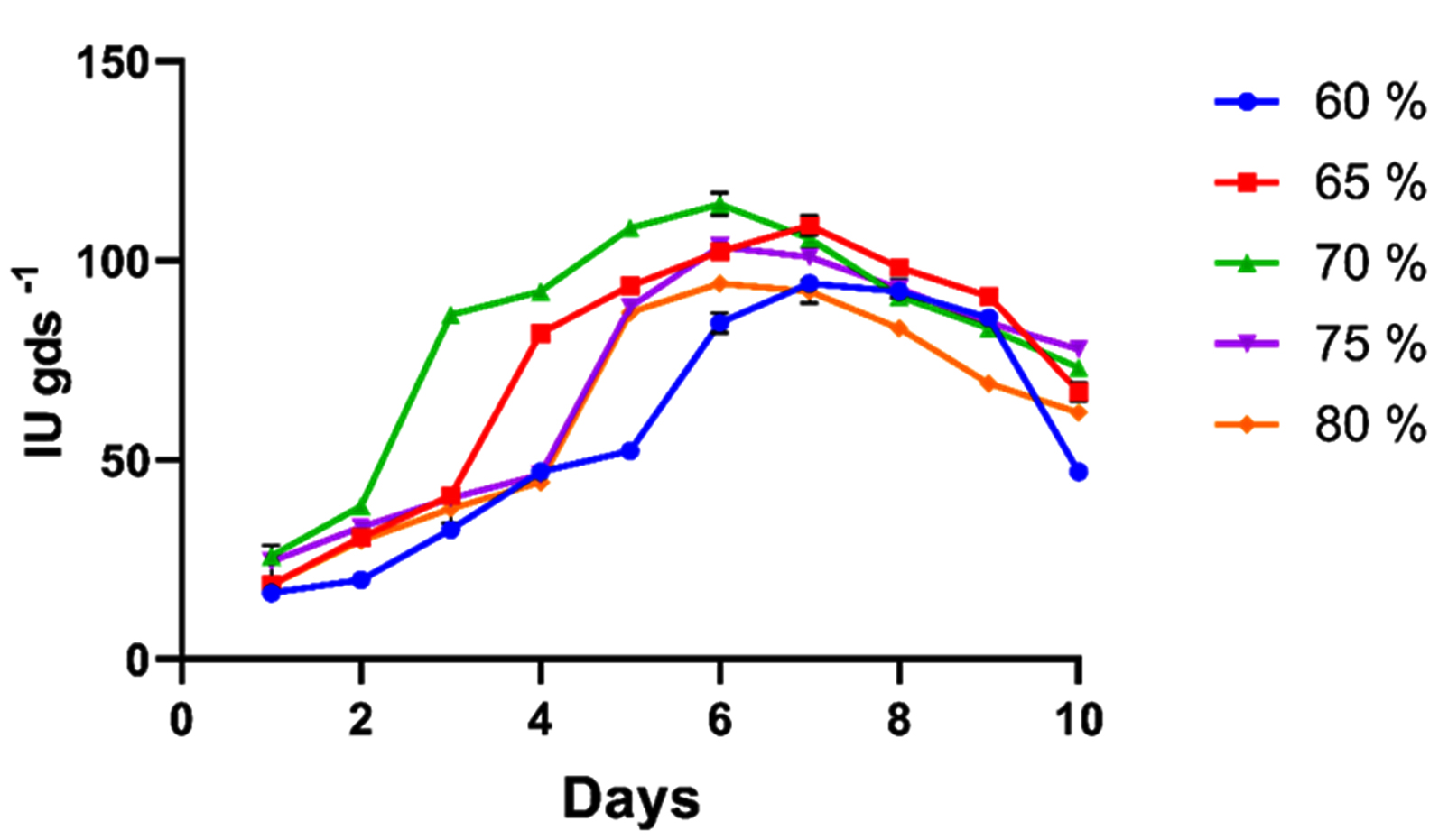
Figure 1. Pre-optimization studies for xylanase activity in solid-state fermentation at different moisture percentages (60, 65, 70, 75 and 80). All the data plotted indicates the mean of three replications accomplished by standard error (SE)
Role of inoculum load on xylanase activity
The effect of inoculum load on xylanase production showed that increasing the inoculum load from 4 to 7 discs (9 mm) significantly elevated xylanase production, with a maximum activity of 118.74 IU gds-1 (Figure 2). Further increase in inoculum load reduced activity, attributed to nutrient limitation. Conversely, a low inoculum load produced insufficient biomass to utilize available nutrients, resulting in reduced enzyme production effectively. Ramanjaneyulu and co-workers conducted a similar study to improve xylanase production by Fusarium sp. BVKT R2 and found the inoculum load of 6 agar discs (0.5 mm) as the optimum inoculum load, and higher loads led to a significant decline in activity.22
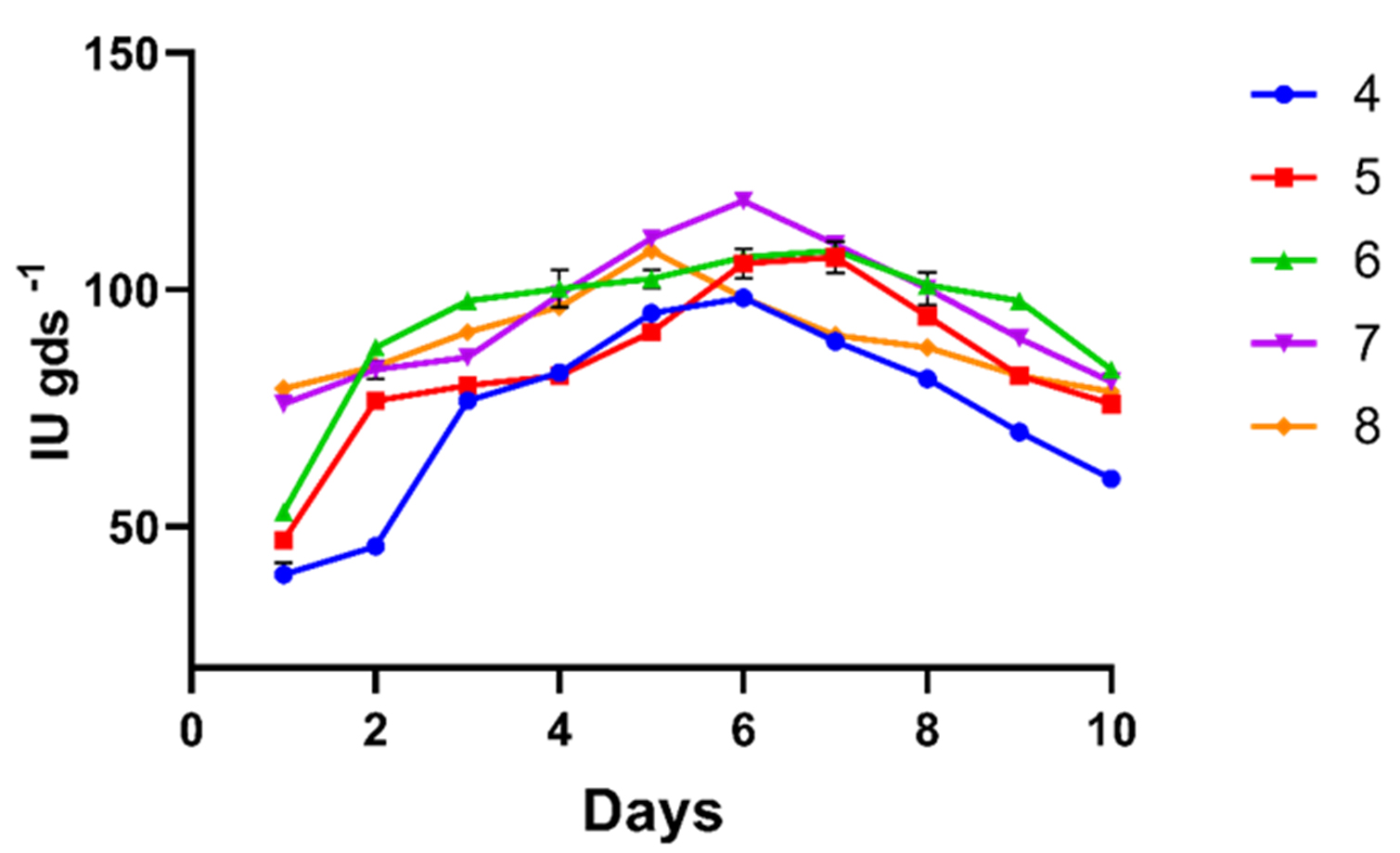
Figure 2. Pre-optimization studies for xylanase activity in solid-state fermentation at different inoculum loads in discs (4, 5, 6, 7, and 8). All the data plotted indicates the mean of three replications accomplished by standard error (SE)
Role of incubation temperature on xylanase activity
The influence of temperature on xylanase showed an increasing trend from 32-34 °C, reaching a maximum of 121.40 IU gds-1 at 36 °C. Further increase in temperature from 36-40 °C significantly reduced enzymatic activity (Figure 3). This trend in enzymatic activity at suboptimal temperatures is consistent with an Arrhenius-type relationship, in which elevated temperature promotes molecular motion and facilitates the reaction to overcome its activation energy barrier. At temperatures above the optimum, protein denaturation predominates, leading to enzyme deactivation and reduced activity.23 Another study on the optimization of xylanase production using wheat bran as a substrate with the A. niger strain BG also reported 36 °C as the optimal temperature within the range of 22-40 °C (4084.75 IU gds-1), which is in agreement with the current findings.24
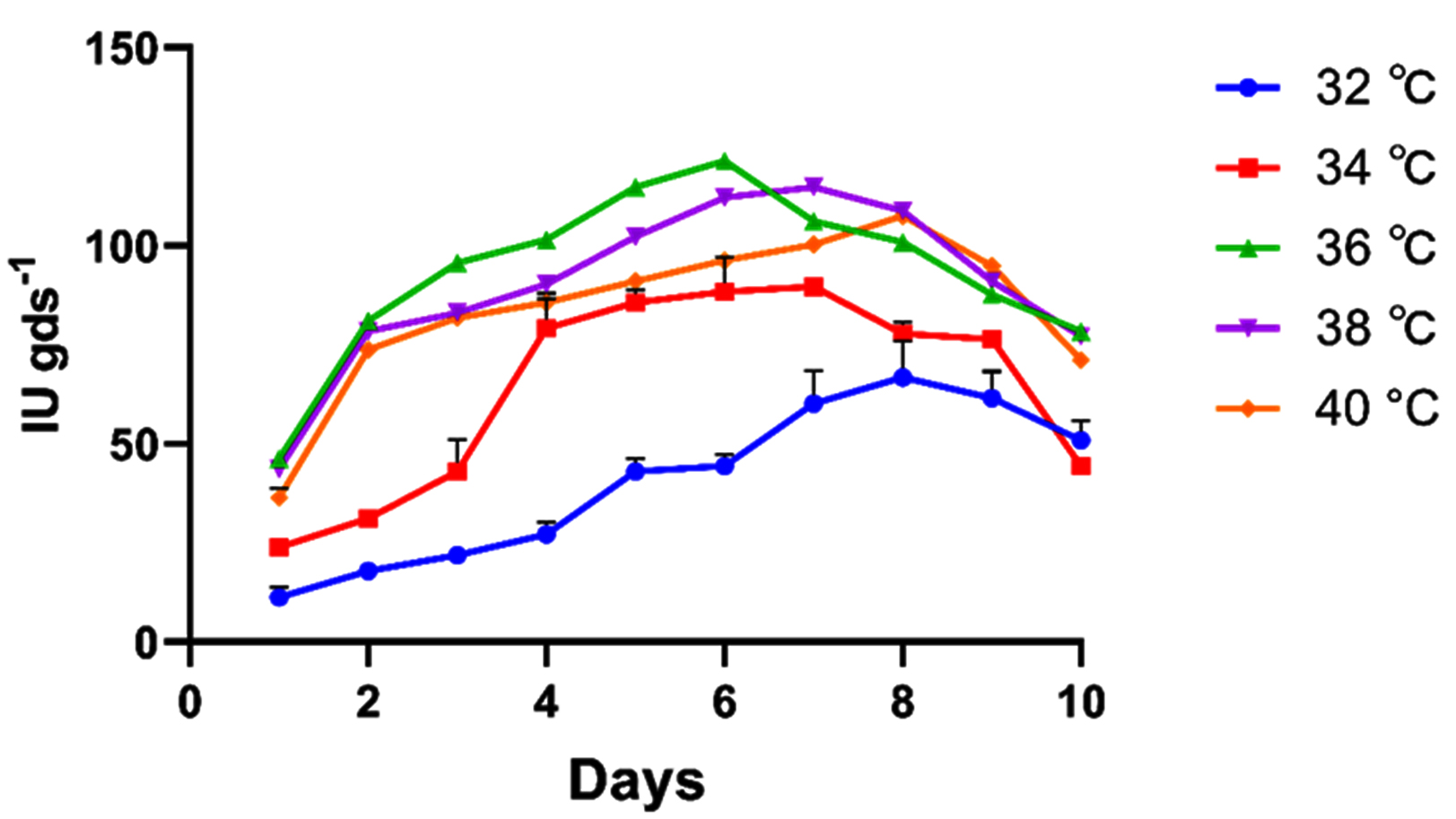
Figure 3. Pre-optimization studies for xylanase activity under SSF at different temperatures (32 °C, 34 °C, 36 °C, 38 °C, and 40 °C). All the data plotted indicates the mean of three replications accomplished by standard error (SE)
Role of pH on xylanase activity
The initial pH of the medium is a key determinant of enzyme synthesis by influencing enzyme stability, membrane transport, nutrient solubility, and the activity of regulatory proteins. The role of initial pH on xylanase was optimized. As depicted in Figure 4, the highest xylanase activity was achieved at pH 5.5 (140.23 IU gds-1). Enzyme activity declined markedly at acidic pH values (≤ 5.0) due to reduced growth and decreased enzyme stability. In contrast, the decline at pH 6.0 indicates that slightly alkaline conditions are also unsuitable for enzyme production. Similarly, a study on the production of xylanase from a white-rot fungus under SSF using wheat bran also concluded that pH 5.0 was optimal for maximum xylanase production.25
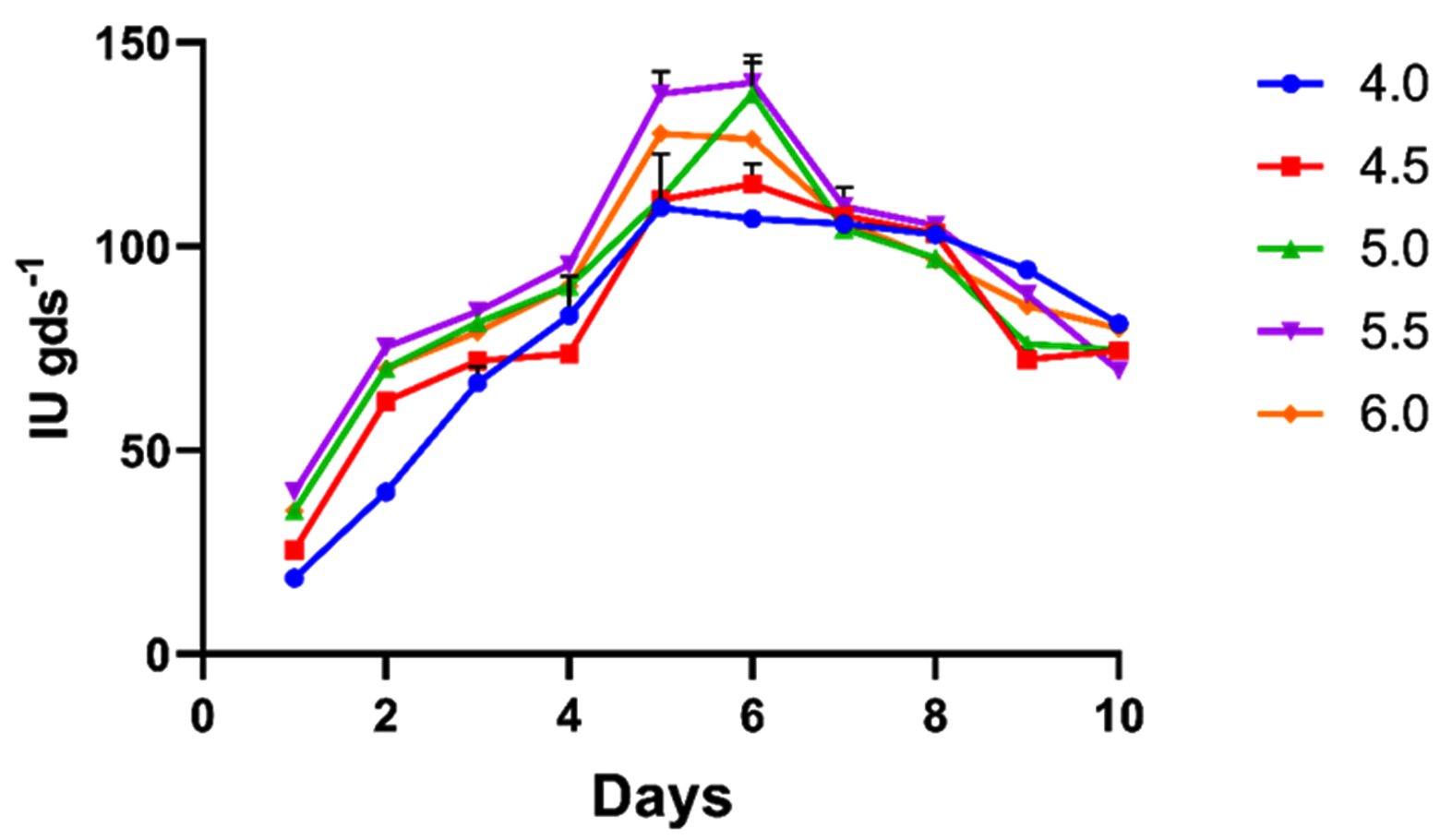
Figure 4. Pre-optimization studies for xylanase activity under SSF at different pH levels (4.0, 4.5, 5.0, 5.5, and 6.0)
All the data plotted indicates the mean of three replications accomplished by standard error (SE)
Statistical optimization of xylanase production using Response Surface methodology
A BBD comprising 29 runs was considered optimal for four significant parameters: moisture percentage, inoculum load, temperature, and pH source to achieve elevated production of the xylanase enzyme. The quadratic model yielded predicted and adjusted R2 values of 0.66 and 0.77, respectively, for xylanase activity. The predicted R2 closely matched the adjusted R2 suggesting that the model is reliable. Additionally, a precision value of 9.69 indicates a highly favorable signal-to-noise ratio. The second-order equation for xylanase optimization, expressed in terms of coded factors and derived from the BBD, is as follows:
Xylanase (IU gds-1) = 126.36- 1.90A+1.35B+1.56+8.57D-3.87AB-5.17AC-2.51AC+5.21BC+1.69BD+0.8731-6.32A2-3.39B2-18.56C2-14.29D2
The effects and significance of variable interactions on xylanase response in the quadratic model were evaluated using Analysis of variance (ANOVA). A model F-value of 7.78 indicates that the model is significant. A small F-value for the model is undesirable as it suggests that the variance is attributable to random, unexplained disturbances (noise). Among the interactions, D, C2, and D2 showed significant positive effects on the production process, with P-values <0.05. The Coefficient of variance (CV) value of 5.92% further demonstrated that the model possessed high precision and reliability (Table 2). The maximum xylanase activity of 147.96 IU gds-1 was obtained with 70% moisture, seven discs of inoculum load, under 36 °C, and pH 5.5 (Figure 5). Plotting the actual value curve as a function of the predicted values, which displays the points dispersed around the regression line confirmed the correlation (Figure 6). In a similar study, to elevate the xylanase activity, various physicochemical parameters were initially optimized using OFAT, then the xylanase production showed three folds increase after optimization through RSM with BBD by A. niger utilizing wheat bran as a sole carbon source.26 A concurrent study showed A. terreus GU227345.1 isolated from local bio waste sources enhanced xylanase activity to 80 IU gds-1 at optimized conditions of 6 g substrate, pH 6.0, 60% moisture, inoculum of 2.75 mL at 35 °C for 4 days.27 Another study reported elevated xylanase activity of 146 IU gds-1 using a fungi Fomes fomentarius under RSM optimized conditions.28 A recent study used RSM optimization to enhance the laccase production to 260 IU gds-1 using Fomes fomentarius.29 RSM is also used to optimize industrially important enzymes such as complex pectinolytic enzymes15 and lipases.30
Table (2):
Analysis of variance (ANOVA) for optimization of xylanase activity under solid-state fermentation
Source |
Sum of Squares |
df |
Mean Square |
F-value |
P-value |
|
|---|---|---|---|---|---|---|
Model |
14 |
332.18 |
7.78 |
0.0002 |
Significant |
|
A-Moisture percent |
4650.56 |
1 |
21.48 |
0.5028 |
0.4899 |
|
B-Inoculum load |
21.48 |
1 |
15.99 |
0.3743 |
0.5505 |
|
C-Temperature |
15.99 |
1 |
19.92 |
0.4663 |
0.5059 |
|
D-pH |
19.92 |
1 |
589.74 |
13.81 |
0.0023 |
|
AB |
589.74 |
1 |
48.60 |
1.14 |
0.3042 |
|
AC |
48.60 |
1 |
78.55 |
1.84 |
0.1965 |
|
AD |
78.55 |
1 |
12.33 |
0.2886 |
0.5995 |
|
BC |
12.33 |
1 |
83.93 |
1.96 |
0.1828 |
|
BD |
83.93 |
1 |
6.99 |
0.1636 |
0.6919 |
|
CD |
6.99 |
1 |
2.21 |
0.0518 |
0.8233 |
|
A2 |
2.21 |
1 |
166.08 |
3.89 |
0.0687 |
|
B2 |
166.08 |
1 |
48.74 |
1.14 |
0.3035 |
|
C2 |
48.74 |
1 |
1525.93 |
35.72 |
||
D2 |
1525.93 |
1 |
848.31 |
19.86 |
0.0005 |
|
Residual |
848.31 |
14 |
42.71 |
|||
Lack of Fit |
597.99 |
13 |
45.98 |
193.16 |
0.0563 |
Not significant |
Pure Error |
597.75 |
1 |
0.2380 |
|||
Cor Total |
0.2380 |
28 |
||||
R² |
0.88 |
C.V % |
5.92 |
|||
Adjusted R² |
0.77 |
Adeq Precision |
9.69 |
|||
Predicted R2 |
0.66 |
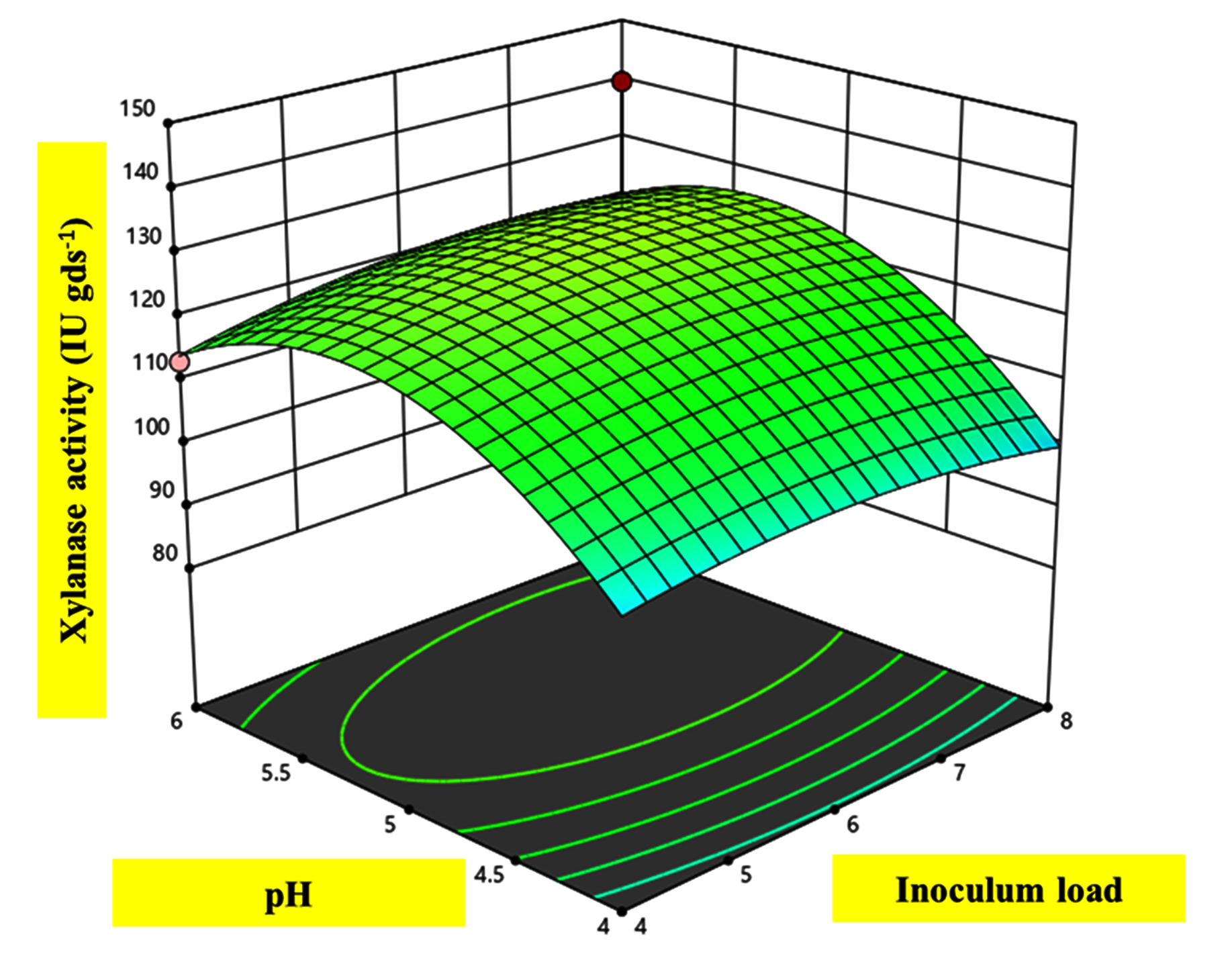
Figure 5. 3D surface plots for the response of xylanase activity (IU gds-1) due to physical and nutritional parameters: pH vs. inoculum load. The dependent effect is visualized by colors ranging from blue to red (blue, green, and red). Blue denotes least significant, green is moderately significant, and red is highly significant
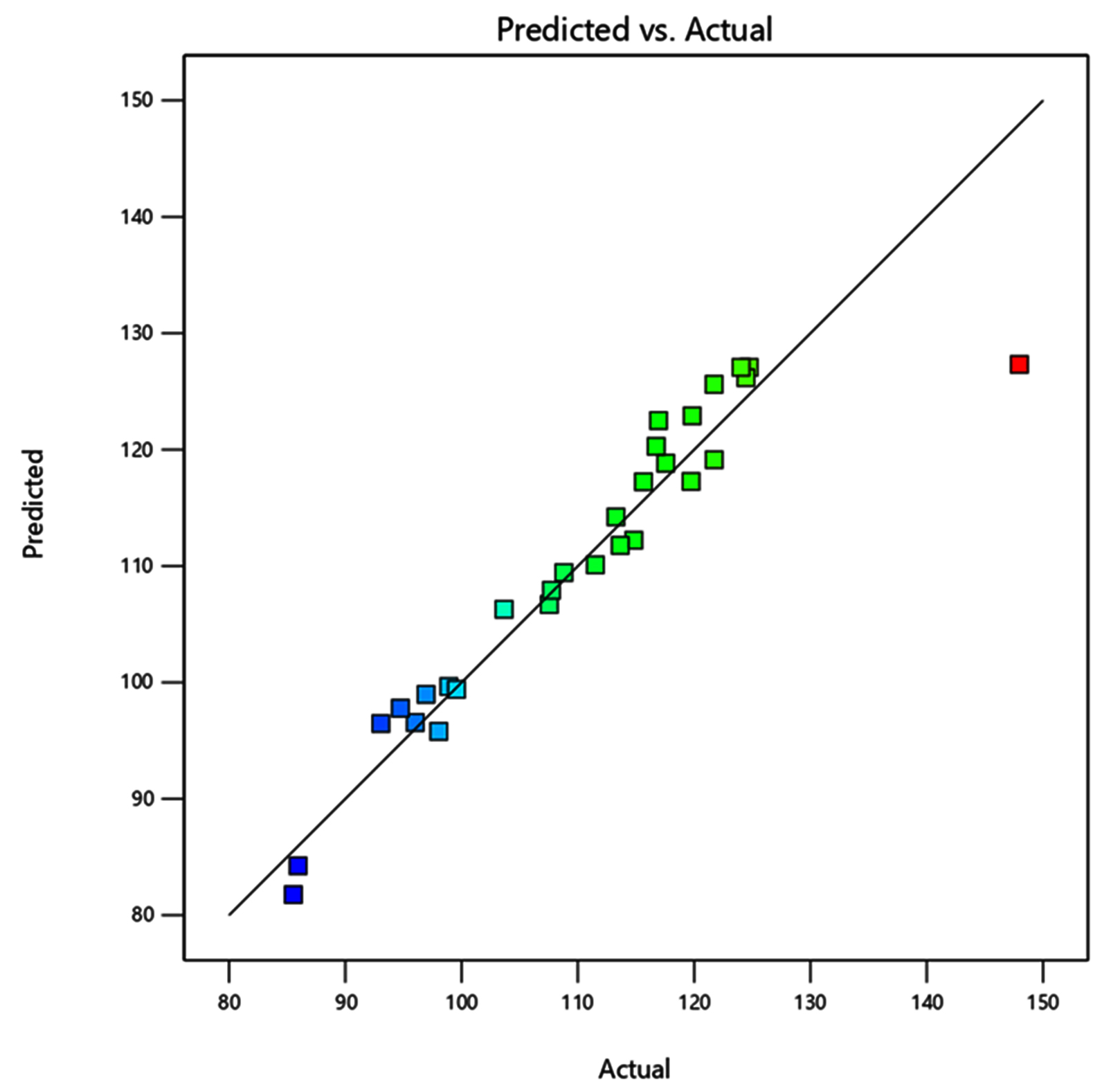
Figure 6. Correlation matrix between predicted value and actual values of xylanase activity. The interactive effect is represented with the color ranging from blue to red (blue, green, red); blue is least significant, green is moderately significant, and red is highly significant
This study demonstrates the production and optimization of xylanase from the in-house potential strain A. fumigatus PSF1 using wheat bran, nutrient-rich substrate, under SSF. Preliminary optimization was performed using OFAT. Subsequent optimization using RSM (BBD) significantly enhanced xylanase yield up to 1.29-fold compared to unoptimized conditions. The model exhibited good predictive capability, as indicated by acceptable R² values, significant model terms, and satisfactory experimental reliability. Optimal conditions included 70% moisture, an inoculum load of seven discs, incubation at 36 °C, and a pH of 5.5; these were experimentally validated and confirmed their suitability for maximizing enzyme output. The study’s findings emphasize the potential of A. fumigatus PSF1 as a promising fungal candidate for cost-efficient xylanase production from agro-residues. The optimized process provides a scalable strategy for sustainable enzyme production and the valorization of agricultural biomass, thereby contributing to broader goals of waste minimization and a circular bioeconomy.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support provided by the ICAR-CIRCOT Collaborative Research Project (CRP-TNAU 02) on Natural Fibers, entitled “Bioprocessing of Natural Fibers and Agro-Residues for the Production of Oligosaccharides and Starch”, awarded to Sivakumar Uthandi. The authors also acknowledge the financial assistance received through the DST-FIST Programme 2022 (TPN 83972).
The authors further express their sincere gratitude to the Metabolomic and Proteomic Analytical Facility (MPAF), Department of Agricultural Microbiology, Tamil Nadu Agricultural University, Coimbatore, India, for providing instrumentation facilities and analytical support that contributed to this research.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
This study was funded by ICAR-CIRCOT –CRP (CRP-TNAU 02).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Perwez M, Al Asheh S. Valorization of agro-industrial waste through solid-state fermentation: Mini review. Biotechnol Reports. 2025;45:e00873.
Crossref - Asim Z, Shahzad HMA, Ghodake G, Mahmoud KA, Almomani F, Rasool K. Transforming Agricultural Food Waste Into Bioplastics: Methods, Potential, and Technological Advances. Adv Sustain Syst. 2025;9(3):2400864.
Crossref - Taghizadeh-Alisaraei A, Tatari A, Khanali M, Keshavarzi M. Potential of biofuels production from wheat straw biomass, current achievements and perspectives: a review. Biofuels. 2023;14(1):79-92.
Crossref - Singh A, Tiwari R, Kour G, Kumar P, Dutt T. Enhanced Income Levels Increase Organic Waste Generation: A Regression-based Modelling of Associated Factors. Journal of Community Mobilization and Sustainable Development. 2025;20(1):158-166.
Crossref - Rani P, Singh B, Tiwari SK. Bacteriocin Production by Lactiplantibacillus plantarum LD1 in Solid-State Fermentation Using Lignocellulosic Substrates. Fermentation. 2025;11(4):233.
Crossref - Sanchez A, Oiza N, Artola A, et al. Solid-state fermentation: a review of its opportunities and challenges in the framework of circular bioeconomy. Afinidad J Chem Eng Theor Appl Chem. 2024;81(601):51-57.
Crossref - Macedo JVC, Abe MM, Sanvezzo PB, Grillo R, Branciforti MC, Brienzo M. Xylan-starch-based bioplastic formulation and xylan influence on the physicochemical and biodegradability properties. Polym Bull. 2023;80(7):8067-8092.
Crossref - Bueno D, Brienzo M. Production of bioplastics with chemical and enzymatic modificated xylan (lignin- and arabinose-free) from sugarcane bagasse. Biotechnol Sustain Mater. 2025;2(1):2.
Crossref - Kumari K, Nagar S, Goyal S, Maan S. Hyper xylanase production and potential of xylooligosaccharides formation from a novel Bacillus australimaris KS2. Biocatal Agric Biotechnol. 2023;54:102899.
Crossref - Vinuthana VH, Govindaraj O, Subramaniam S, Gnanachitra M, Uthandi S. Harnessing lignocellulosic biomass: Insights into pre treatment strategies and hydrolytic enzyme production. Ind Crops Prod. 2025;229:120986.
Crossref - Akram F, Safdar M, Shabbir I, Fatima T, Ikram-ul-Haq. Insight into the eminent biotechnological applications of xylanolytic enzymes for sustainable bioprocessing. 3 Biotech. 2025;15(8):249.
Crossref - Bueno D, de Freitas C, Brienzo M. Enzymatic Cocktail Formulation for Xylan Hydrolysis into Xylose and Xylooligosaccharides. Molecules. 2023;28(2):624.
Crossref - Bajaj BK, Khajuria YP, Singh VP. Agricultural residues as potential substrates for production of xylanase from alkali-thermotolerant bacterial isolate. Biocatal Agric Biotechnol. 2012;1(4):314-320.
Crossref - Khanahmadi M, Arezi I, Amiri MS, Miranzadeh M. Bioprocessing of agro-industrial residues for optimization of xylanase production by solid- state fermentation in flask and tray bioreactor. Biocatal Agric Biotechnol. 2018;13:272-282.
Crossref - Salim D, Anwar Z, Zafar M, Anjum A, Bhatti KH, Irshad M. Pectinolytic cocktail: Induced yield and its exploitation for lignocellulosic materials saccharification and fruit juice clarification. Food Biosci. 2018;22:154-164.
Crossref - Arumugam DP, Uthandi S. Optimization and characterization of laccase (LccH) produced by Hexagonia hirta MSF2 in solid-state fermentation using coir pith wastes (CPW). J Environ Manage. 2024;356:120625.
Crossref - Cui X, Li X, Li S, et al. Xylanase VmXyl2 is involved in the pathogenicity of Valsa mali by regulating xylanase activity and inducing cell necrosis. Front Plant Sci. 2024;15:1342714.
Crossref - Nassef HM, Ahmed HA, Bashal AH, et al. A novel Six Sigma approach and eco-friendly RP-HPLC technique for determination of pimavanserin and its degraded products: Application of Box–Behnken design. Rev Anal Chem. 2024;43(1):20230073.
Crossref - Box GEP, Behnken DW. Some New Three Level Designs for the Study of Quantitative Variables. Technometrics. 1960;2(4):455-475.
Crossref - Ali S, Noor P, Ahmad MU, et al. Kinetics of cellulase-free endo xylanase hyper-synthesis by Aspergillus niger using wheat bran as a potential solid substrate. BMC Biotechnol. 2024;24(1):69.
Crossref - El-Shishtawy RM, Mohamed SA, Asiri AM, Abu-bakr MJ, Ibrahim IH, Al-Talhi HA. Solid fermentation of wheat bran for hydrolytic enzymes production and saccharification content by a local isolate Bacillus megaterium. BMC Biotechnol. 2014;14(1):29.
Crossref - Ramanjaneyulu G, Sridevi A, Seshapani P, et al. Enhanced production of xylanase by Fusarium sp. BVKT R2 and evaluation of its biomass saccharification efficiency. 3 Biotech. 2017;7(5):351.
Crossref - Kabir MF, Ju LK. On optimization of enzymatic processes: Temperature effects on activity and long-term deactivation kinetics. Process Biochem. 2023;130:734-746.
Crossref - Azzouz Z, Bettache A, Djinni I, Boucherba N, Benallaoua S. Biotechnological production and statistical optimization of fungal xylanase by bioconversion of the lignocellulosic biomass residues in solid-state fermentation. Biomass Convers Biorefinery. 2024;10(15):e35496.
Crossref - Abena T, Simachew A. Production and characterization of acidophilic xylanase from wood degrading white rot fungus by solid-state fermentation of wheat straw. Heliyon. 2024;10(15). https://doi.org/10.1016/j.heliyon.2024.e35496
- Azzouz Z, Bettache A, Boucherba N, et al. Optimization of β-1,4-Endoxylanase Production by an Aspergillus niger Strain Growing on Wheat Straw and Application in Xylooligosaccharides Production. Molecules. 2021;26(9):2527.
Crossref - Raza A, Anwar Z, Zafar M. Production, optimization and purification of xylanase from isolated fungal strain Aspergillus terreus GU227345. 1 by screening of local biomass. Biomass Bioenergy. 2026;208:108872.
Crossref - Al-Sa’ady AJR. Determination of optimum conditions for xylanase production by Aspergillus niger using solid state fermentation. Iraqi J Sci. 2016;57(3):2220-2227.
- Riffat A, Anwar Z, Zafar M, Nadeem F, Mehmood T. Optimization of physicochemical parameters and characterization laccase enzyme produced by a novel strain of Fomes fomentarius through solid-state fermentation. Biomass Conv Bioref. 2022;14(3):3999-4006.
Crossref - Ali U, Anwar Z, Hasan S, et al. Bioprocessing and screening of indigenous wastes for hyper production of fungal lipase. Catalysts. 2023;13(5):853.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.