


Adverse environmental conditions impose stress in the plant, like heat, cold, salinity, heavy metal, nutrient, droughtstress; play a very prominent role to limit the plant growth and development. During the exposure of these wide ranges of stresses, plant itself changing or developing at morphological, biochemical or molecular level. During the morphological changes, plant not only reduceduration of life but also reducing plant body skeletons like shortening of plant height, while the adjustment of ion transport, carbon metabolism and synthesis of osmoregulatory compound or compatible molecules like proline, glycine betaine, sorbitol takes place at molecular level. Different kinds of compatible solutes enable to the plant to tolerate abiotic stress. Proline is one the most effective compatible molecule that produces in the wide range of organism including plant under the adverse conditions. A transcriptional characteristic known as up and down regulation of gene expression may be observed during osmotic stress. These changes may be facilitating directly by various stress condition. The up regulation in the plants result, proline accumulation under osmotic stress while down regulation result deterioration process.
Agriculture, Biotic, Cadmium, Density, Energy, Forage
Proline is a water soluble amino acid and beneficial solute that accumulate in plant under different kinds of stresses such as drought, cold, heat, heavy metal, nutrient and salt stress. Initially the proline accumulation occurs in plant tissues in response to stresses that cause reduction of water result low water potential such as drought, salinity and freezing but also recorded in response to heavy metal toxicity(Anaytullah, 2007; Verslues et al., 2006; Sharma and Dietz, 2009 and Siddique and Dubey, 2017).The capacity of proline accumulation depending upon the level of stress and also can vary from one species to another species up to many foldin respect of control(Hayat et al., 2012 and Verbruggen and Hermans, 2008). Now a day, it is established that the accumulation of proline within plants under adverse or stress conditions play a significant roles for developing stress tolerance capacity and act as an osmoregulatory compound (Hare and Cress, 1997).Heat stress, salt stress and heavy metal stress is another important abiotic stress (Anaytullah et al., 2012; Munns and Tester, 2008 and Siddique and Dubey, 2017) which inversely affect the crop growth in term of biomass and yield of crop (Ahmad et al., 2012; Mantri et al., 2012 and Karur et al., 2012). In saline situation, Osmotic regulation is an important mechanism at cellular level by virtue plants accumulate many compatible solutes inside the cell like proline, polyols and betaine that play a an important role to mitigate the effect of salinity stress in barley plant(Hasegawa et al., 2000 and Chen et al., 2007). While, heat stress also reduce crop growth and accelerate the accumulation of different organic base biochemical compound i.e. proline, betaine and sugar by adapting specific strategies. However, the synthesis of proline under various kinds of stresses results stabilization process cell structure (Anaytullah et al., 2012; Kaushal et al., 2011and Ashraf and Foolad, 2007). Synthesis of osmolytes (osmoregulatory compound) especially proline enable plants to develop osmotic adjustment through osmoregulatory process under diverse environmental conditions. Therefore, it is well known with osmoprotectant compound which can stabilize the effect of stresses at cellular level (Aziz and Khan, 2001).
Role of proline in heat stress
Heat stress in term of increased temperature beyond the threshold level, have a detrimental effect on crop growth and their development around the world.The harmful effect of heat stress can be overcome by the development of thermo tolerance genotypes by using modern approaches. Therefore, it is urgent need to understand the response of plant against heat stress. In general, the term heat stress may be defined as increase in temperature beyond a threshold level for a period of time sufficient to cause permanent damage to plant growth and development. When temperature raises more than 40 0C, plants suffer with higher amount of water loss through stomata that reduced the chlorophyll content and ultimately rate of photosynthesis and respiration and enhance membrane damageat molecular level (Gosavi et al.,2014; Kumar et al., 2012 and Jagtap et al., 1998).A well known concept of increased temperature making imbalance between rate of photosynthesis and respiration in which rate of photosynthesis decrease due to decrease the activity of rubisco at carboxylation site and rate of photorespiration increased consistently as temperature goes beyond the threshold level (Fig.1)due to this reason, growth and development of plant is adversely affected therefore, the yield of crop plant reduced (Cen and Sage; Hall, 2001;Kurukulasuriya and Rosenthal,2013 and Siddique and Bose, 2015). To mitigate the effect of heat stress, genetic approaches may be one of tool which helps in developing thermo tolerance variety of plant (Wahid et al., 2007). Wide ranges of compound are known at present time which may help to protect the plant against heat stress injury within the plant by maintaining osmotic and turgor pressure that facilitate the movement of water within the plant cells. Such kind of compound is known as compatible solutes. Proline is one of the compounds that play a very crucial role for tolerance mechanism against various kind of stress (Shevyakova et al., 2009 and Anaytullah et al., 2012). Proline is not only acting as osmolytes but also have a three very fine role in inner side of plant i.e. metal chelator, antioxidant defense and signaling molecule (Hayat et al., 2012). Effect of heat stress in beginning stage appear at cellular level specially plasma membrane and biochemical reaction event (Sung et al., 2003 and Anaytullah et al., 2012).Increased proline content within the plant due to heat stress act asa osmoprotectant for plant cell (Kumar et al., 2012). The most popular biosynthetic pathway of proline synthesis takes place in cytoplasm is Glutamate via ¯-Glutamyl phosphate and Glutamyl- ¯-semialdehydeand initial two steps of proline biosynthesis is initiated byd-1-pyrroline-5-carboxylate synthetase (Parida et al., 2008). Proline accumulation due to stress play a multifunctional role within the plant like low molecular weight chaperons, stabilizing proteins and enzymes and scavenging reactive oxygen species (Hameed et al., 2012, Anaytullah et al., 2012).

(Source: Adopted from Hasanuzzaman et al., 2013)
Fig.1. Effect of heat stress on plant
Role of proline in salinity stress
Word “salinity”represents diverse form of problems related to the soil when excessive amount of salts gathering takes place. This diverse form of gathering of salt within soil may be dividing into two types especially alkaline and saline soils. The major root cause for the development of saline soil is the mismanagement of irrigation water in the soil because scarcity of drainage facility for leaching of salts resulting salty soil (IRRI, 2011; Iyenger and Reddy,1994). Therefore, salinity of soil plays a very wide role for better crop growth and development from initial stage of seed germination to the yield of crop. The intensity of side effect of salinity may be vary depending upon the amount of salt deposition in the soil and as well as tolerance level of particular crop (Lauchli and Grattan, 2007 and Munnset al., 2002). Various attempts were taken by the worker to find out some important selection criteria for salinity tolerance during early stage of sowing especially up to seedling establishment but initial screening or evaluation have no much importance in respect to yield under salinity stress condition (Igartua et al., 1994 and Munns et al., 2002). Production of reactive oxygen species i.e. radicals of H2O2,O2– and OH+increased under various kind of stress (i.e. salinity, heat, heavy metal, drought, high light intensity etc.) in which hydroxyl radical is one of the most reactive compound at molecular level(Fig.2). Therefore, these reactive oxygen species may contribute a lot as a damaging molecule under adverse environment (Apel and Hirt, 2004; Das and Roychoudhury, 2014; Hasegawa et al., 2000; Prasad et al., 1994). The primary strategy to mitigate the effect of salinity stress is the accumulation of compatible solutes within the plant(Sakamoto and Murata, 2000 and Hasegawa et al., 2000). In case of salt sensitive variety of barley, accumulation of osmolytes proline, glycine betaine and polyols along with NaCl2 induced K+efflux were found effective indicator of salt stress tolerance (Di Martino et al., 2003 and Chen et al., 2007).A comparative study between sensitive and tolerance genotype reported that the proline content increased under salt stress conditions while an average increase of proline was fivefold in root and twenty fold in leaf(Chen et al., 2007 and Munns, 2002) Instead of conventional role of compatible solutes those are synthesize during the salinity stress have multifunctional role to stabilize plant cell and their skeleton for growth and development via protecting photosystem II, structure of enzymes and regulating reactive oxygen species(Abbas et al., 2018; Zhu et al., 2001; Bohnert et al., 1995; Noiraud et al., 2001)(Fig. 3).
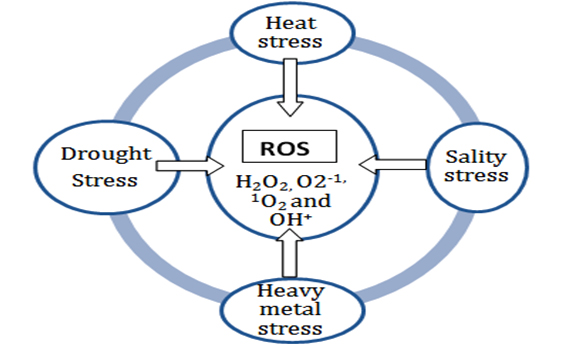
(Source: Adopted from Das and Roychoudhury, 2014)
Fig.2. Production of ROS in various kind of stress
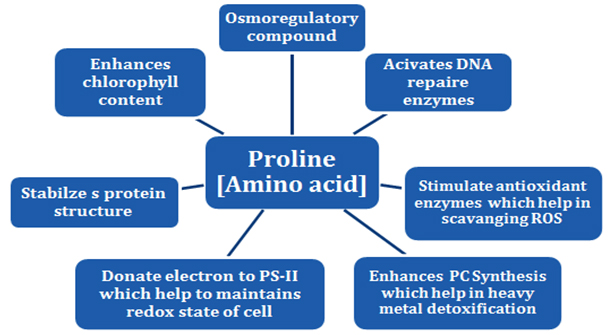
(Source: Adopted from Abbas, et al., 2018)
Fig. 3. Multifunctional role of proline under various kind of stress in plant
Role of proline in heavy metal stress
Industrialization, weathering of rocks and urbanization are the main source for causing heavy metal pollution in the soil and water that is one of the most important natural resources. Due to this activity, many harmful heavy metals such as Cu, Pb, Cd, As, Ni, Mn, Zn and others enter in the soil, water and air therefore; they cause serious damage to the plant and animal. Theses heavy metals are very useful for plant growth and development when they are in limited range but beyond the limit will be very harmful (Hall, 1981). Various methods have been used to make plants defensive against the toxicity of heavy metals including agronomical practices (Siripornadilsul et al., 2002). Many scientists believe that proline synthesis in the plant may be result of any kind of injury either through, heat, cold, salinity or heavy metal (Anaytullah et al., 2012; Anaytullah and Bose, 2007; Chen 2007and Lutts et al., 1996). Biosynthesis of proline accelerated when the plants are treated with toxic heavy metal content while the activity of SOD and membrane injury increased (Bose et al., 2008; Siddique and Dubey, 2017; Schat et al., 1997). Accumulation of heavy caused toxicity within the plant (a kind of stress) consequently proline synthesized and helps to protect the plant (Siddique and Dubey, 2017and Kavi Kishor et al., 1995).Various kind of stress commonly induced oxidative stress that lead to cause a process known as lipid per-oxidation thereby disruption takes place at cellular level especially plasma membrane therefore leakage of potassium from the plant cellwhile the finding of research also shows that exogenous application of proline not only suppress the heavy metal induced lipid per oxidation but also potassium leakage that may execute to provide protection to the plant cell (Mehta and Gaur, 1999; Seel et al., 1992 and Cuin and Shabala, 2007 and Hasanuzzaman et al., 2010b).
Finally, we can conclude that, now- a –days, people facing the daunting challenges for the remediation of heavy metals from our contaminated agricultural fields. The stresses viz. Heavy metal, Salinity and Heat, finally produce the Reactive Oxygen Species, which lead the membrane dame and diminishing the homeostasis of the plant cells. At the same time Proline play a significant role in the form of protectant/ osmolyte for defending the cross talk stresses in the plants.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
- Abbas, G., Murtaza, B., Bibi, I., Shahid, M., Niazi, N. K., Khan, M. I., Amjad, M., Hussain, M. and Natasha. Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects. International Journal of Environmental Research and Public Health, 2018; 15(59): 2-45
- Anaytullah, Bose, B and Yadav, R. S. PEG induced moisture stress: screening for drought tolerance in rice. Indian J. Plant Physiol., 2007; 12(1): 88-90.
- Anaytullah, Srivastava, A. K. and Bandana Bose. Impact of seed hardening treatment with nitrate salts on nitrogen and anti oxidant defense metabolisms in Triticum aestivum L. under different sowing conditions. Vegetos, 2012; 25 (1): 292-299.
- Ashraf, M. and Foolad, M. R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and Experimental Botany, 2007; 59(2): 206-216.
- Aziz, I., Khan, M. A. Effect of seawater on the growth, ion content and water potential of Rhizophora mucronata Lam. J. Plant Res., 2001; 114: 369–373.
- Bohnert, H. J., Nelson, D. E. and Jensen, R. G. Adaptations to environmental stresses. The Plant Cell, 1995; 7: 1099–1111
- Cen, Y. P., Sage, R. F. The regulation of Rubisco activity in response to variation in temperature and atmospheric CO2 partial pressure in sweet potato.Plant Physiol., 2005; 139(2): 979-90
- Chen, Z., Tracey, A. C., Zhou, M., Twomey, A., Bodapati, P. N. and Shabala, S. Compatible solute accumulation and stress mitigating effects in barley genotypes contrasting in their salt tolerance. J. Exp. Bot., 2007; 58: 4245–4255
- Cuin, T. A., and Shabala, S. Compatible solutes reduce ROS-induced potassium efflux in Arabidopsis roots. Plant Cell Environ., 2007; 30: 875–85
- Das, K. and Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Fron. Environ. Science, 2014; 2: 1-13
- Di Martino, C., Delfine, S., Pizzuto, R., Loreto, F. and Fuggi, A. Free amino acids and glycine betaine in leaf osmoregulation of spinach responding to increasing salt stress. New Phytologist, 2003; 158: 455–463.
- Gosavi, G. U., Jadhav, A. S., Kale, A. A., Gadakh, S. R., Pawar, B. D. and Chimote, V. P. Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (Sorghum bicolor) at seedlings stage, Indian Journal of Biotechnology, 2014; 13: 356-363
- Hall, A. E., Crop Responses to Environment. CRC Press LLC, Boca Raton, Florida 2001.
- Hare, P. D., and Cress, W. A. Metabolic implications of stress induced proline accumulation in plants. Plant Growth Regulation, 1997; 21: 79-102.
- Hasanuzzaman, M., Hossain, M. A. and Fujita, M. Selenium in Higher Plants: Physiological Role, Antioxidant Metabolism and Abiotic Stress Tolerance.J PlantSci., 2010b; 5: 354–375
- Hasanuzzaman, M., Nahar, K., Alam, M. M., Roychowdhury, R. and Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013; 14: 9643-9684
- Hasegawa, P. M., Bressan, R. A., Zhu, J. K., and Bohnert, H. J. Plant cellular and molecular responses to high salinity. Annual Review of Plant Physiology and Plant Molecular Biology, 2000; 51: 463-499.
- Hayat, S., Hayat, Q., Alyemeni, M. N., Wani, A. S. Pichtel, J., Ahamad, A. Role of proline under changing environments. Plant Signal and Behavior, 2012; 7(11): 1456-1466.
- Jagtap, V., Bhargava, S., Streb, P. and Feierabend, J, Comparative effect of water, heat and light stress on photosynthetic reactions in Sorghum biolor (L.) Moench, Journal of Experimental Botany, 1998; 49: 1715-1721
- Kaushal, N., Gupta, K., Bhandhari, K., Kumar, S., Thakur, P. and Nayyar, H. Proline induces heat tolerance in chickpea (Cicer arietinumL.) plants by protecting vital enzymes of carbon and antioxidative metabolism. Physiol. Mol. Biol. Plants. 2011; 17(3): 203–213.
- Kavi Kishor, P. B., Hong, Z., Miao, G. H., Hu, C. A. and Verma, D. P. S. Over expression of D”-pyrroline-S-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiology, 1995; 108: 1387–1394
- Kumar, S., Gupta, D. and Nayyar, H. Comparative response of maize and rice genotypes to heat stress: Status of oxidative stress and antioxidants, Acta Physiology of Plant, 2012; 34: 75-86
- Kurukulasuriya, P., and Rosenthal, S. Climate change and agriculture: A review of impacts and adaptations. Retrieved from Washington, D.C.: 2013; https://openknowledge.worldbank.org/handle/10986/16616 License: CC BY 3.0 IGO
- Lauchli, A. and Grattan, S. R. Plant growth and development under salinity stress. (eds.) Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops, 2007; 1–32
- Lipiec, J., Doussan, C., Nosalewicz, A., and Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. [yes]. International Agrophysics, 2013; 27: 463-477.
- Lutts, S., Kinet, J. M. and Bouharmont, J. Effects of various salts and of mannitol on ion and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) callus cultures. Journal of Plant Physiology, 1996; 149: 186–195
- Noiraud, N., Maurousset, L. and Lemoine R. Transport ofpolyols in higher plants. Plant Physiology and Biochemistry, 2001; 3: 717–728
- Munns, R. Comparative physiology of salt and water stress. Plant, Cell & Environment, 2002; 25(2): 239-250
- Munns, R. and Tester, M. Mechanisms of salinity tolerance. Annual Review of Plant Biology, 2008; 59: 651-681.
- Parida, A. K., Dagaonkar, V. S., Phalak, M. S. and Aurangabadkar, L. P. Differential responses of the enzymes involved in proline biosynthesis and degradation in drought tolerant and sensitive cotton genotypes during drought stress and recovery, Acta Physiology of Plant, 2008; 30: 619-627
- Sakamoto, A. and Murata, N. Genetic engineering of glycine betaine synthesis in plants: current status and implications for enhancement of stress tolerance. Journal Experimental Botany, 2000; 51: 81–88.
- Sharma, S. S. and Dietz, K. J. The relationship between metal toxicity and cellular redox imbalance. Trends in Plant Science, 2009; 14: 43-50.
- Siddique A. and Dubey, A. K. Phyto-toxic eûect of heavy metal (CdCl2) on seed germination, seedling growth and antioxidant defence metabolism in wheat (Triticum aestivum L.) variety HUW-234. International Journal of Bio-resource and Stress Management, 2017; 8(2): 261-267.
- Siripornadulsil, S., Traina, S., Pal, D., Verma, V. and Sayre, R. T. Molecular Mechanisms of Proline-Mediated Tolerance to Toxic Heavy Metals in Transgenic Microalgae. The Plant Cell, 2002; 14: 2837-2847.
- Sung, D. Y., Kaplan, F., Lee, K. J. and Guy, C. L. Acquired tolerance to temperature extremes. Trends Plant Sci., 2003; 8: 179–187
- Verbruggen, N. and Hermans, C. Prolineaccumulationinplants:areview. Amino acids, 2008; 35(4): 753-759.
- Verslues, P. E., and Bray, E. A. Role of abscisic acid (ABA) and Arabidopsis thaliana ABA-insensitive loci in low water potential-induced ABA and proline accumulation. Journal of Experimental Botany, 2006; 57: 201-212.
- Vinocur, B. and Altman, A. Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Curr. Opin. Biotechnol., 2005; 16: 123–132
- Wahid, A. and Close, T. J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant., 2007; 51: 104–109
- Zhu, J. K. Cell signaling under salt, water and cold stresses. Current Opinion in Plant Biology, 2001; 4: 401–406.
© The Author(s) 2018. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.