


ISSN: 0973-7510
E-ISSN: 2581-690X
This study investigated the exopolysaccharide (EPS) producing ability of an endophytic fungus GloS2. It involved examining various aspects, including identifying and cultivating endophytic fungi, producing and characterizing EPS, optimizing the production process, and evaluating their antioxidant properties. The fungus GloS2 (Fusarium sp.) was optimized for EPS synthesis through one variable at a time (OVAT) and response surface methodology (RSM) to determine the optimal culture environment and growth medium components, resulting in the maximum EPS production of 5.39 gL-1. Furthermore, the EPS exhibited significant antioxidant activity with a greater IC50 value (15.05 ± 0.51 µg mL-1) in the hydroxyl radical scavenging assay. The Fourier Transform Infrared Spectral analysis indicates the presence of chemical groups i.e., O-H, C-H, C-O-H, and C-O. Additionally, gas chromatography-mass spectrometry (GC-MS) analysis revealed that erythrose, fructose, and mannose were the major monosaccharide components, with a molar proportion of nearly 1:1:1 and a molecular mass of ~1.12 x 105 Da. Moreover, the structural characterization of EPS has confirmed the existence of α-glycosidic linkage along with β-D-galactopyranosyl moieties and substituted glucose residues.
Endophytic Fungi, Heteropolysaccharide, Process Optimization, in vitro Antioxidant
The domain of microbial exopolysaccharide (EPS) and its associated bioactivity in pharmaceuticals has been studied for a long ago. These are majorly novel microbial metabolites with sustainable utilities and are less toxic to contemporary synthetic chemicals. EPS from novel sources, e.g. fungal endophytes of ethnomedicinal plants are potent contributors to the field of biotechnology and come with a variety of bioactivities in both medicine and agriculture. EPS from fungal endophytes of various structures, and requires particular media composition and growth conditions.
EPSs are the secondary metabolites produced by various fungi under different growth conditions. These complex and distinct macromolecules exhibit various utilities i.e., antioxidative, immunomodulatory, antitumor, antiaging, and hypoglycemic.1
Endophytes are a type of fungus that resides in a mutualistic association with plants during a phase of their life cycle. They can remain inside the plant tissues without leading to immediate signs of infection or visible disease development.2 These fungi are a reservoir of distinctive bioactive secondary metabolites, including alkaloids, quinones, phenolic acids, steroids, tannins, and terpenoids. These compounds may exhibit antibacterial, anti-insect, anticancer, and other effects.
Antioxidants play a vital role in safeguarding health by providing the initial defence against oxidative damage and effectively scavenging free radicals.3 They protect cell function against homeostatic disturbance, including those from septic shock, aging and oxidative stress. Although aerobic microorganisms need oxygen to survive, it can lead to harmful reactive oxygen species.4 ROS and reactive nitrogen species can lead to oxidative damage in cellular components. Developing new antioxidant drugs is vital for combating ROS and preventing chronic disease.5 Recently endophytic fungi have gained attention for their ability to produce bioactive compounds with anticancer potential. Endophytic fungus Curvularia geniculata isolated from Phyllanthus niruri showed strong anticancer and antioxidant properties, particularly effective against HepG2 cell lines.6
Optimizing microbial metabolite biosynthesis is vital for industrial applications, and the effectiveness of fermentation processes is affected by parameters like as the media composition, starting pH, incubation period, and temperature. Structure-function connection of fungal EPS is critical since they have considerable antioxidant capabilities and act as free radical scavengers to attenuate oxidative damage.7 Exopolysaccharides produced by Lasiodiplodia theobromae have shown significant antioxidant and antimicrobial activity enhancement.8
The flocculation capability of EPS is a crucial aspect of biopolymer applications.9 Bio-flocculants are biopolymers that induce flocculation by creating bridges between themselves and other particles. This feature leads to the aggregation and precipitation of suspended particles. Hence, there is a shift toward substituting synthetic polymers with bio-flocculants due to their easy biodegradability, eco-friendly application in wastewater treatment processes, and negligible toxicity to humans and the environment. These attributes have positioned biopolymers as a focal point of interest for numerous researchers in the current era.10
Here optimization of EPS production from endophytic fungi Fusarium sp. GloS2 has been performed using “one variable at a time” (OVAT) approach and response surface methodology (RSM) technique. Subsequently, the characterization and antioxidative activity of the isolated EPS was investigated.
Isolation of endophytic fungi
Healthy Globba marantina plants were collected from Ghatshila, East Singhbhum, of the Jharkhand district (latitude 22.5873°N and longitude 86.4744°E). Plant parts were rinsed thoroughly with tap water followed by the surface sterilization procedure. After that stem portions were placed on water agar plates at 27°C in a BOD incubator.11 After 4-7 days, fungal mycelium appeared from the tissues and was inoculated to a PDA (Potato Dextrose Agar) plate for optimum growth.
ITS-based molecular characterization of the endophytic fungal isolate
After 7 days of incubation, the reproductive structure of the endophytic fungi was studied microscopically (Leica DM 3000). Finally, rDNA molecular-based gene sequencing analysis of the universal ITS1 and ITS4 sequences was performed following standard procedures.12 The ITS sequences were aligned to the NCBI GenBank. Utilizing MEGA 11 software, multiple sequence alignment was performed and similar sequences were included within the alignment which was done through CLUSTALW for phylogenetic tree preparation.13
Production and Optimization of EPS
Production of EPS
The experiment was conducted using a 250 mL Czapek Dox Broth (CDB) broth. The microorganism was initially maintained in 100 mL of 6.0 pH at 37°C in an incubator for 8 to 10 days. Fungal spore suspensions were grown in conical flasks after 8 to 10 days. After centrifugation of fungal biomass, the filtrate and fermentation broth were recovered. The fungal biomass was dried at 80°C (12 hours) and then weighed. To separate the cells, the fungal culture broth was centrifuged for 10 minutes at 10,000 rpm. The crude EPS was recovered after centrifugation, freeze-dried, and stored in double volumes of cold ethanol.14 Quantitative sugar and protein analysis was performed using the standard methodology of previous work.15,16 Monosaccharide analysis was done after lyophilization of the viscous EPS.
Optimization by the OVAT method
The culture parameters and medium content had a substantial impact on the synthesis of EPS. Despite prior research indicating that the F. solani SD5 strain produces the most polysaccharides,17 it is critical to understand that the Fusarium sp. GloS2 isolate may have unique growth needs for producing a significant amount of EPS. As a result, there is a need to investigate and comprehend the variables that favor large EPS generation from GloS2.
Therefore, the OVAT method was applied for optimization of the production process. Various carbon sources (2 g 100 mL-1), such as glucose, dextrose, galactose, fructose, maltose, sucrose, and galactose, were utilized simultaneously in PDB media to study the necessity of supplementary nutrients for the development of mycelium and EPS production. After that, two nutrients of varying concentrations were administered to the culture media that were successful in EPS and biomass development. Some metal ions (0.5 g 100 mL-1), such as NaNO3, KH2PO4, and NaCl, were also added to the culture media to determine how efficient they were at producing EPS and biomass.
Optimization using Response Surface Methodology (RSM)
RSM optimization was performed by analyzing the results of the OVAT experiments. A three-level (-1, 0, 1) BBD approach was used. Of the six critical factors involved in EPS production, only four-glucose concentration (GC), yeast extract concentration (YEC), pH of fermentation media (MpH), and fermentation time (FT) were used for RSM-BBD analysis. The four mentioned parameters (Supplementary Table 1) were applied in three different approaches (-1, 0, and +1, which indicate values lower than the optimum, optimum, and higher than the optimum, respectively). The second-order polynomial equation YEPS= β0 +∑β1x1 +∑βijxixj +∑βiix2i can estimate the proper relationship between the factors. YEPS represents the dependent variable, independent factors are indicated through xi and xj, bi, bii, and bij are the linear, quadratic, and interaction coefficients, respectively.
FTIR spectroscopic assessments
Infrared spectra were captured using KBr pellets in an FTIR spectrophotometer (PerkinElmer FT-IR C98747) between 400 and 4000 cm-1.18
XRD assessments
To evaluate the physical properties of the EPS, X-ray diffraction (XRD) was carried out with a powder diffractometer (RIGAKU MINIFLEX II).19 Dry powder was applied to the sample tray, and scanning was conducted over a range of 2q angles (20-70°C).
Monosaccharide analysis
For monosaccharide analysis, viscous polysaccharides were lyophilized, and then the dried polysaccharide was analyzed following the protocol of De and Nag.20 In brief, 100 mg of dried samples were prepared using methanol, ribitol, and norleucin. Afterwards, the mixture was heated to 70°C (15 minutes), centrifuged, and then the supernatant was treated with methoxyamine HCL followed by trimethylsiloxane (TMS). The sample was then injected into GC-MS for monosaccharide analysis. For running this program, the DB-5 Ultra Inert column was used, followed by the described protocols.21
Molecular weight determination
Gel permeation chromatography (GPC) was employed to determine the molecular weight (MW) of the EPS. This determination employed a Sepharose-6B column (65 cm x 2 cm) with dH2O as the eluent (flow rate of 4 mL min-1) and dextran (40, 70, and 200 kDa) as a reference.22
Advanced spectroscopic analysis
The basic characteristics of EPS were determined via 1D 1H and 13C 400 MHz NMR spectra. The lyophilized samples were solubilized in D2O for analysis of NMR. The spectra were captured on a Bruker AscendTM 400 and analyzed with Bruker TopSpin 4.4.0 software.
Flocculation property
The flocculation capacity of EPS was assessed by the technique of Devi et al.,23 with minimal changes. Concisely, the solution of EPS was stirred with 1% CaCl2 and 0.5% kaolin suspension (pH 7.0). The resultant compound was vortexed for 2 min then the uppermost surface of the sample was retrieved and quantified at 550 nm using a UV-Vis spectrophotometer. Flocculation activity (FA) of EPS was evaluated using the following formula: a (%) = {(a–b) / b} x100, where ‘a’ and ‘b’ are the OD values of the control (dH2O) and EPS samples, accordingly.
Water holding capacity (WHC) analysis
EPS was assessed according to the earlier published methodology with slight modifications.24 For this test, in a microcentrifuge tube, 15 mg of EPS sample was mixed with Milli-Q water and then weighed. Then the sample was kept at ambient temperature for 24 hours. Then incubation, the mixture was centrifuged and then the supernatant was discarded. The tubes were reweighed, and the WHC calculation was carried out using the given formula.
WHC = Water attached weight (mg) / primary sample weight (mg) x 100.
Screening for antioxidant activity
Common free radical producers, such as DPPH (2,2-diphenyl-1-picrylhydrazyl), ABTS (2,2¹-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) were used to assess the capacity of EPS to combat oxidative stress. H2O2 (hydrogen peroxide), and FRAP (ferric ion reducing antioxidant power) assays were also performed. Ascorbic acid served as the reference antioxidant. For this evaluation, the spectrophotometric method was utilized.25
The antioxidant activity of EPS was assessed following the procedure of Blois et al.,26 Rajurkar et al.,27 Winterbourne28 and Oyaizu.29 The results were calculated using the following formula [(Ab – As) /Ab] x 10.
Statistical analysis
All trials were executed in triplicate, and the results are expressed as the means ± standard errors (SE). Data analysis was conducted by Prism Graph Pad software version 8.2 (San Diego, California, USA). Additionally, Minitab (version 20.2) software was utilized for the response surface methodology experiment, which was based on Box Behnken Design (BBD).
EPS producer GloS2 is identified as Fusarium sp.
4 endophytic fungi were obtained and the one with the highest EPS production was further studied for its identification. The mycelia of isolate lack any special structure was confirmed as Fusarium sp. GloS2 with a GenBank Acc. No.- OP379518. To determine its phylogenetic position, we employed the MEGA 11 program utilizing the obtained ITS. The resulting phylogenetic tree is shown in Figure 1. To capture microscopic images, polarized microscopic equipment was utilized. These images revealed the sterile mycelial structure and provided top and bottom views of the plate morphology (Figure 1).
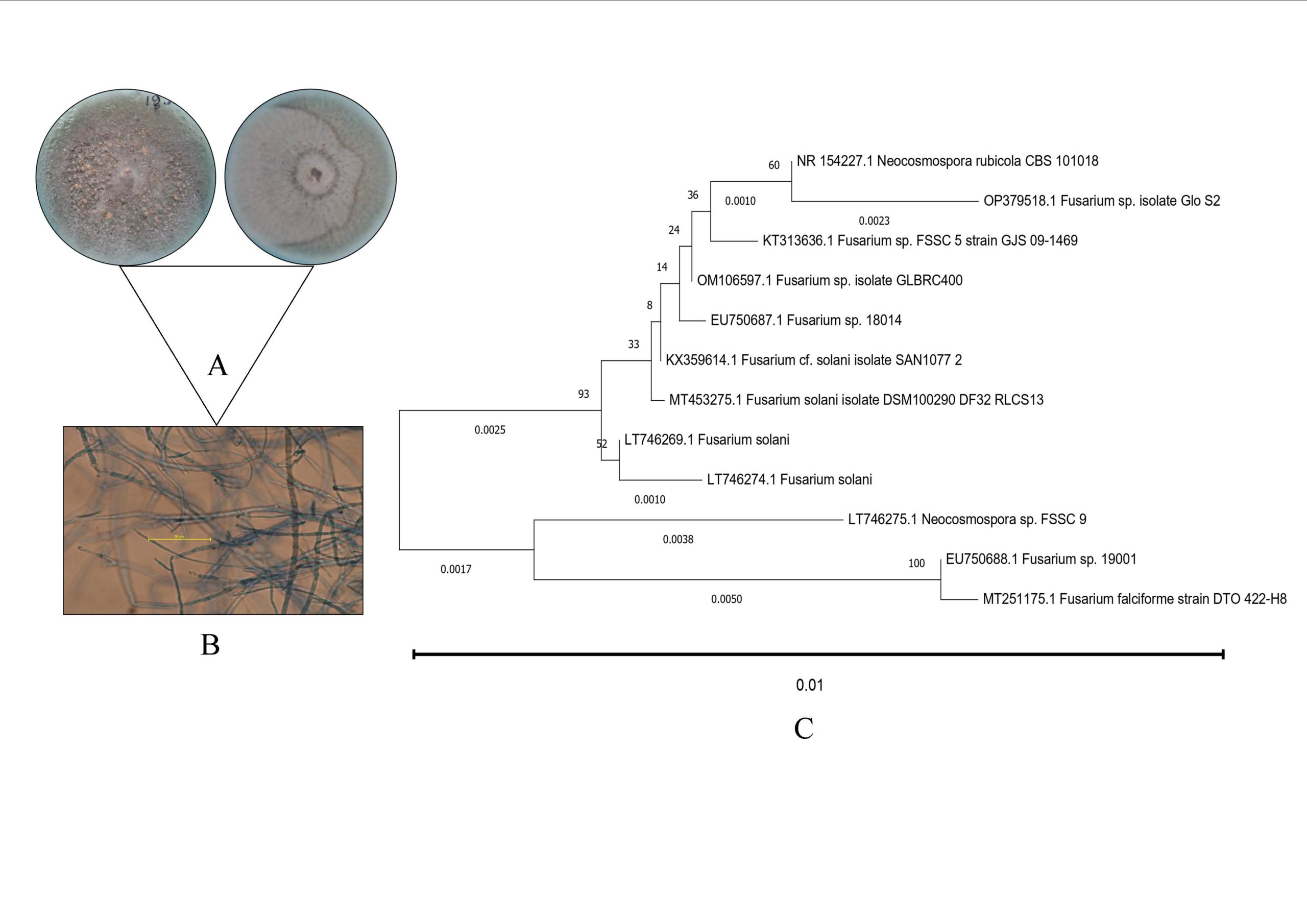
Figure 1. Plate morphology of the endophytic isolate (a) microscopic image of the endophytic isolate (b) and evolutionary relationship of Fusarium sp. GloS2 with several related fungi (c)
Production of and optimization of the production parameters
The OVAT optimization findings indicated that a 5 gL-1 GC, 0.4 gL-1 YEC, an MpH of 7, an 8-day FT, and a temperature of 35°C resulted in the highest EPS production of GloS2 (Table).
Table:
Effects of various parameters on the growth of GloS2
| Parameters | Impacting components | Concentration of components (g L-1) | Fungal cell mass (g L-1) | EPS obtained (g L-1) |
|---|---|---|---|---|
| Fermentation time (in day) | 2 | – | 4.03 ± 0.20 | 0.86 ± 0.02 |
| 4 | – | 4.57 ± 0.10 | 1.02 ± 0.05 | |
| 6 | – | 6.55 ± 0.02 | 1.52 ± 0.01 | |
| 8 | – | 8.98 ± 0.03 | 1.68 ± 0.02 | |
| 10 | – | 8.94 ± 0.01 | 1.49 ± 0.04 | |
| Incubation temperature (ᵒC) | 24 | – | 6.22 ± 0.02 | 1.18 ± 0.36 |
| 28 | – | 7.79 ± 0.04 | 2.02 ± 0.02 | |
| 30 | – | 6.34 ± 0.03 | 1.52 ± 0.02 | |
| 35 | – | 8.79 ± 0.34 | 2.77 ± 0.01 | |
| 37 | – | 8.22 ± 0.54 | 1.55 ± 0.03 | |
| Initial medium pH | 4 | – | 6.98 ± 0.06 | 2.80 ± 0.02 |
| 5 | – | 7.27 ± 0.01 | 0.68 ± 0.03 | |
| 6 | – | 8.98 ± 0.05 | 1.38 ± 0.22 | |
| 6.5 | – | 7.87 ± 0.04 | 3.44 ± 0.03 | |
| 7 | – | 8.19 ± 0.03 | 3.89 ± 0.02 | |
| Additional carbon source | Amylose | 1 | 3.50 ± 0.11 | 3.54 ± 0.03 |
| Fructose | 1 | 6.50 ± 0.04 | 3.02 ± 0.07 | |
| Dextrose | 1 | 8.40 ± 0.07 | 4.87 ± 0.07 | |
| Sucrose | 1 | 8.70 ± 0.01 | 3.87 ± 0.13 | |
| Glucose concentration | Glucose | 2 | 8.12 ± 0.20 | 3.02 ± 0.10 |
| 4 | 8.72 ± 0.07 | 3.22 ± 0.08 | ||
| 5 | 8.90 ± 0.03 | 4.87 ± 0.05 | ||
| 6 | 9.50 ± 0.17 | 4.32 ± 0.05 | ||
| 7 | 9.67 ± 0.05 | 4.00 ± 0.03 | ||
| 8 | 8.90 ± 0.15 | 3.59 ± 0.01 | ||
| Nitrogen source | Yeast extract | 0.3 | 2.50 ± 0.02 | 4.02 ± 0.02 |
| NH4NO3 | 0.3 | 3.20 ± 0.01 | 3.18 ± 0.08 | |
| Yeast extract concentration | 0.1 | – | 2.90 ± 0.07 | 2.98 ± 0.21 |
| 0.3 | – | 4.10 ± 0.02 | 3.87 ± 0.01 | |
| 0.4 | – | 4.89 ± 0.09 | 4.98 ± 0.23 | |
| 0.5 | – | 4.89 ± 0.02 | 3.57 ± 0.05 | |
| 0.6 | – | 5.02 ± 0.07 | 3.00 ± 0.07 | |
| Different metal ions | NaCl | 0.05 | 2.20 ± 0.12 | 1.20 ± 0.04 |
| KCl | 0.05 | 2.90 ± 0.10 | 1.50 ± 0.01 | |
| MgCl2 | 0.05 | 2.87 ± 0.08 | 0.97 ± 0.03 | |
| CaCl2 | 0.05 | 2.65 ± 0.05 | 1.30 ± 0.02 |
Following OVAT optimization, the study used the RSM approach with a tri-model BBD design (Box Behnken design), with four replicates at the center point for critical parameters such as the amount of glucose, amount of yeast concentration, fermentation media pH, and fermentation time. This model aimed to investigate the most suitable EPS production process. Supplementary Tables shows the experimental design, including expected and measured data related to EPS production. The expected response of Y to EPS synthesis by Fusarium sp. GloS2 was defined to the coded factors as detailed below:
YEPS = -116.88 + 49.65 YEC + 4.970 GC + 26.82 MpH + 6.034 FT – 52.05 YEC*YEC- 0.5409 GC*GC – 2.0221 MpH*MpH – 0.7181 FT*FT – 1.458 YEC*GC- 0.505 YEC*MpH + 0.183 YEC*FT – 0.0530 GC*MpH + 0.2162 GC*FT- 0.0450 MpH*FT
To evaluate the degree of fit of the RSM to the experimental data, a regression analysis was conducted (Supplementary Table 2). The F statistics of the model (291.91) are significant. To assess the goodness of fit of the regression equation, the determinant coefficient (R2Adj) of the model was checked. An R2Adj value of 99.14% was obtained from the RSM-generated second-order polynomial equation (Supplementary Table 1). This value indicates that both the obtained and assumed values are in proper alignment. The model is good from its fitness point of view, as the calculated F statistics related to ANOVA is 21.55 and the model is accurate in depicting any equation related to EPS production. The model was also consistent, as the P value for lack of fit was 0.005. Each model term was regarded as significant if the Probability value of the probability was within a range of 0 to 0.05. The interactions between glucose and yeast extract were the most important (P < 0.05). Therefore, YEC and FT (P < 0.05) were correlated, and a three-dimensional surface plot was generated utilizing the information from the constructed model. This 3D plot provides an idea of the possible interactions required for the optimum synthesis (Supplementary Figure 1). The 3D plots have significant quadratic surfaces compared to the linear planes. The RSM-BBD yielded a maximum production of 5.39 g L-1 using the said parameters.
FTIR analysis
The FTIR spectra of GloS2 within the 400- 4500 cm-1 range are shown in Figure 2. The FTIR spectra, as depicted in Figure 3, revealed the potential chemical groups present in the structure of EPS. A prior study illustrated that EPS encompasses a variety of functional groups.30 The prominent and wide absorption bands observed at 3300 cm-1 in the EPS spectrum are associated along with the presence of hydroxyl (O-H)functional groups.31 In addition, bands at 2982 cm-1 suggest alkyl C-H bonds inside the carbon ring, which is a typical characteristic of polysaccharides.32 This finding is similar to previous findings on polysaccharide architectures.33 Additionally, the absorption peak in the 1000-1200 cm-1 is a distinctive characteristic of sugar derivatives. The unique properties of the EPS observed at 800-1500 cm-1 region are generally determined by the vibrational properties of the glucose unit. Polysaccharides are specifically found to have bands that range from 800 to 1200 cm-1. The peak underneath 1000 cm-1 is frequently attributed to phosphate group bending vibrations. Thus, the presence of a phosphate group may be indicated by the small peak at 628.75 cm-1.34 Additionally, distinctive bands of EPS were observed in the 800-1200 cm-1 range.
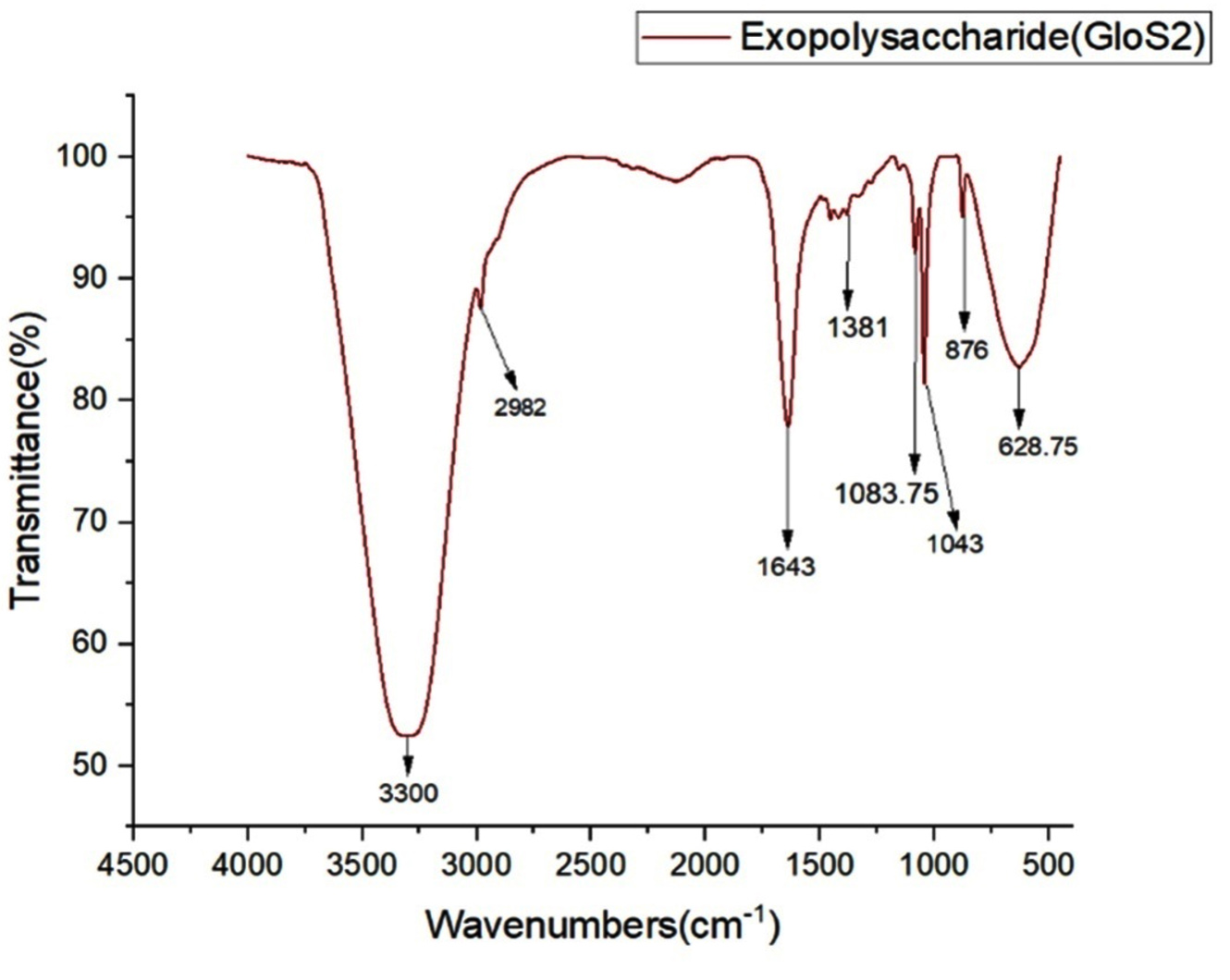
Figure 2. FTIR spectroscopy of GloS2 EPS
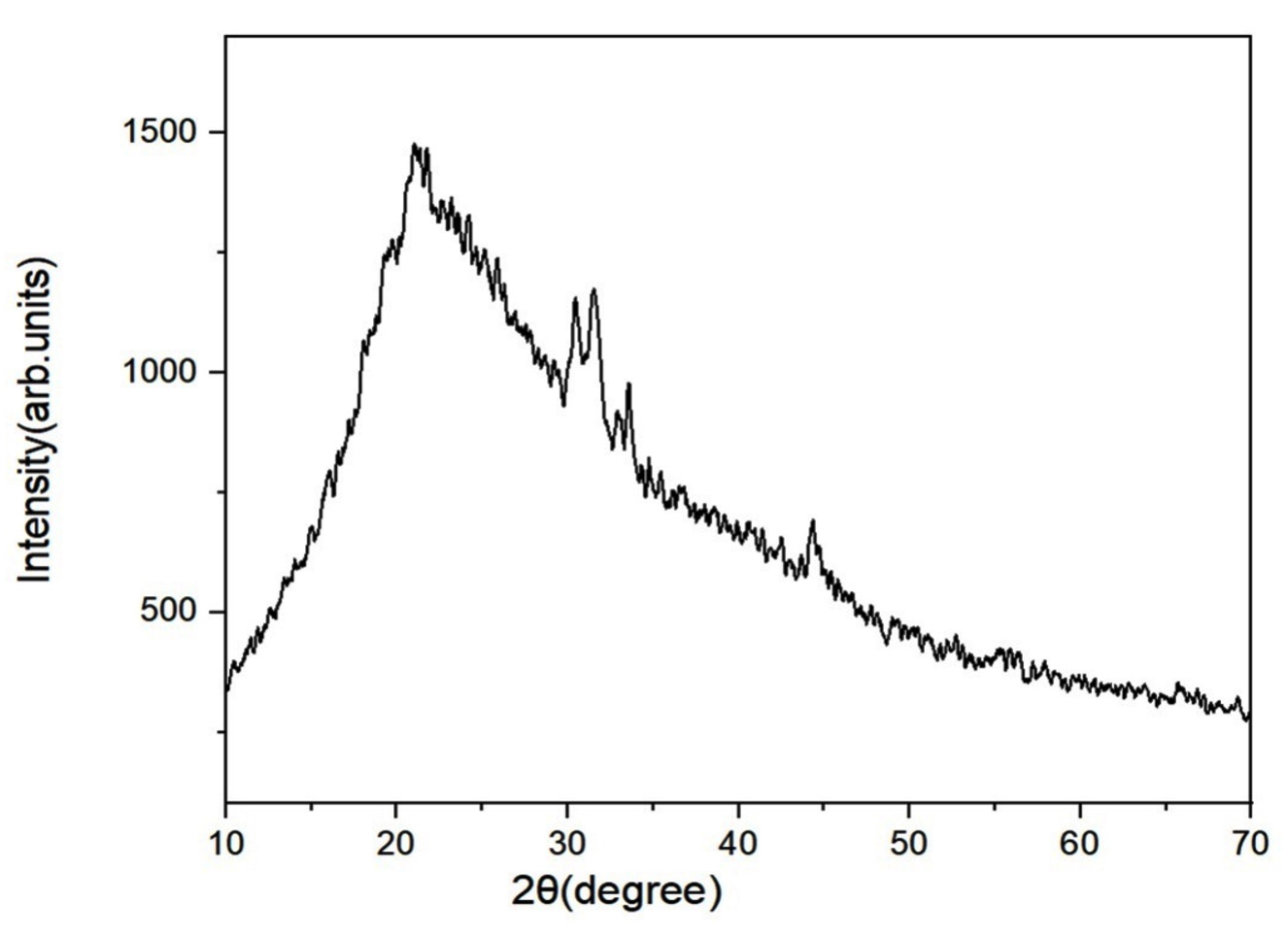
Figure 3. X-ray diffraction (XRD) analysis of GloS2 EPS
XRD analysis
It is a robust tool to assess the amorphous and crystalline properties of materials in both qualitative and semiquantitative ways. The XRD pattern of GloS2 EPS (Figure 3) shows distinct peaks in the 2q range of 20-50. The existence of wide peaks can be attributed to the polymer structure.
Analysis of monosaccharide compositions
The monosaccharide components are erythrose (RT-50.65 min), fructose (RT-57.61 min), and mannose (RT-58.69 min). Fructose is the most abundant component compared to sugars. So, it is a heteropolysaccharide type of EPS.
Molecular weight determination
EPS is composed of erythrose (with a retention time of 50.65), fructose (RT-57.61), and mannose (RT-58.69) in a 1:1:1 ratio with a molecular weight of ~ 1.12׳ 105 Da. Each repeating unit of EPS contains one erythrose, fructose, or mannose unit. Therefore, two hundred thirty-three repeating units are present in the EPS.
NMR analysis
In this study, we used 1H and 13C NMR spectra for EPS analysis. Signals within the range of 5.3 to 3.3 ppm (1H) and 60 to 103 ppm (13C) are characteristics of polysaccharides.35 The 1H NMR spectra are shown in Figure 4A. Four signals from a-anomeric proton at 5.31, 5.30, 5.12 and 5.11 ppm and it’s confirmed that a-glycosidic linkage existed in this EPS.36 The chemical shift from 4.1 to 3.3 ppm has indicated the a-glycosidic rings.37A signal in the range of 5.12 was attributed to galacturonate acid residues.38 13C NMR spectra showed in Figure 4B, the anomeric carbon signals at 103,92 ppm. Also, a single spectrum at 92.11 indicated that it was a-pyranose, similar to the polysaccharide from Cordyceps sinensis.39 The C2-C6 signals were observed within the region of 60-92 ppm. The downfield chemical shift at about 76.3 stated that it was (1 4,6)-b-D-galactopyranosyl moiety, similar to what was seen with the EPS from Fusarium solani.22 The chemical shift at 62.34 ppm represented the unsubstituted C6 and attributed at about 69.59 ppm was represented as the substituted of C4 glucose residue.40
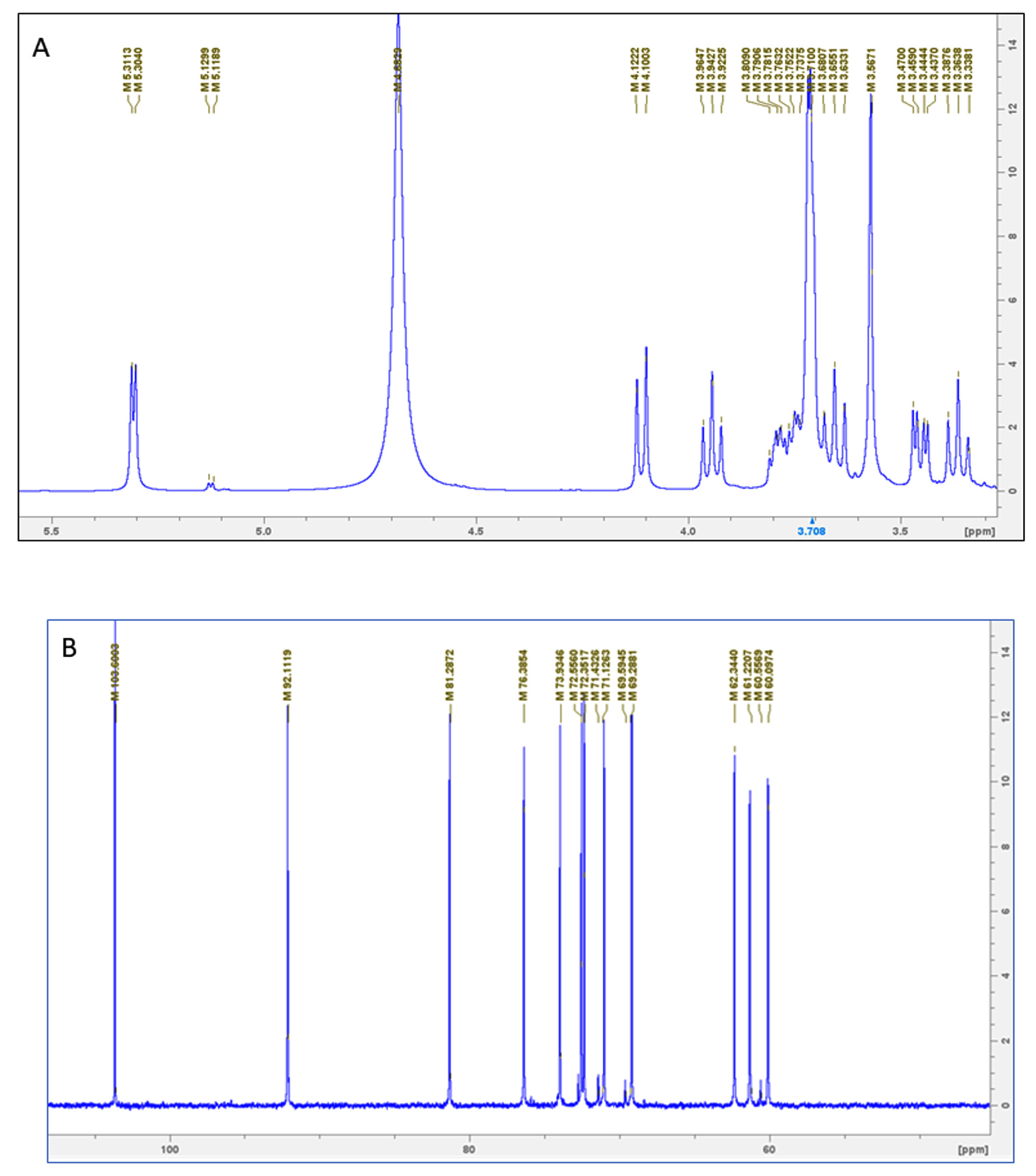
Figure 4. NMR spectrum analysis of GloS2 (a) 1H NMR (b) 13C NMR
Flocculation activity
The EPS showed significant flocculation activity (Supplementary Table 3). The highest flocculating activity (88.37%) of the EPS was observed at a concentration of 40 mgL-1 in comparison to the equal amount of standard clay suspension.
Water holding capacity (WHC) analysis
The hygroscopic nature of EPS is reflected in its water-holding capacity (WHC), which is related to the existence of hydrogen bonds within the polymer substances, enabling the retention of water.41 The WHC of GloS2 EPS was 188.07 ± 1.33%.
Antioxidative activity of GloS2-EPS
The IC50 value for the hydroxyl radical scavenging ability of GloS2 was 15.05 ± 0.51 µg mL-1 (Supplementary Table 4). However, compared to ascorbic acid (AsA), EPS showed greater scavenging activity.
The IC50 value DPPH radical scavenging was 29.21 ± 0.41 µg mL-1, which is equivalent to that of AsA (31.93 ± 0.68 µg mL-1). Therefore, a comparison shows that the scavenging ability of EPS is moderate against DPPH radicals (Supplementary Table 4).
In the ABTS radical assay, antioxidants can contribute electrons or hydrogen atoms, influencing the color of ABTS, and the antioxidant capacity of the sample was estimated (Supplementary Table 4). GloS2 had a scavenging capacity of 25.35 ± 0.086 µg mL-1 against ABTS radicals. The results indicated that EPS had greater scavenging ability than AsA.
Reducing ability was assessed in the reducing power assays by the reaction change of Fe3+ to Fe2+. The EPS of GloS2 showed a greater absorbance value than the standard, indicating a stronger reductive potential and electron donor capacity for free radical stabilization. IC50 value was 73.44 ± 1.32 µg mL-1 (Supplementary table 4). Furthermore, the investigation revealed that the antioxidant potency of the sample simultaneously depended on the concentration.
With several uses in both medicine and agriculture, endophytic fungi have long been reliable sources of potent bioactive chemicals.42 These endophyte-derived secondary metabolites have proven to be viable alternatives to common antibacterial and antioxidant substances. In particular, EPS exhibits considerable potential and has excelled in this domain.43 Extracellular polymeric substances from endophytic fungi have gained widespread recognition for their potent antioxidative properties. They hold the promise of replacing currently used components in this role. To realize their full potential for pharmaceutical applications, it is imperative to maximize their production. To attain the optimum yield of EPS, a variety of optimization strategies are being used.
The fermentation duration is the first crucial factor in ensuring maximum production of EPS. The highest yield of EPS was observed after 8 incubation days in the current study. Table shows the daily fluctuations in fungal growth and related EPS production. EPS production gradually increased between days 2 and 8 but then abruptly declined. The decline in EPS production may be due to EPS mobilization by the organism, possibly influenced by EPS hydrolase along with depletion of the carbon source also reduced broth viscosity supported this belief. An extended incubation period also promotes EPS lyase activity, reducing EPS levels.44 The data also revealed similar incubation durations for EPS production.45,46
The synthesis of EPS is significantly influenced by the temperature either enzymatic stimulation or inhibition. Furthermore, both EPS and biomass production were reduced at certain temperatures. The synthesis of any microbial metabolite is particularly sensitive to the pH of the medium.47 pH plays a vital role in enzymatic activity for high EPS production. The maximum biomass and EPS were achieved in this investigation at an initial medium pH of 7 (3.890 g L-1). D-glucose was shown as the most efficient carbon source for achieving maximal synthesis of EPS. The production shows a robust correlation with the addition of carbon sources.45 Experiments with varying glucose concentrations (from 2% to 5%) revealed that biomass and EPS production increased simultaneously within this range. However, the best concentration for generating the greatest amount of EPS was 5% glucose, which produced a yield of 4.876 gL-1. Furthermore, nitrogen was shown to be an essential ingredient for the formation of EPS and biomass. High amounts of EPS and biomass were produced by yeast extract and NH4NO3, with the most favorable amount of yeast extract concentrate being 0.4 gL-1, which produced EPS at a rate of 4.98 gL-1. Moreover, metal ions are key to biological functions by acting as catalysts in essential enzyme reactions. In this study, EPS production was greater in KCl-supplemented media than in NaCl– and MgCl2–supplemented media. These results emphasize that metal ions may increase cell membrane permeability and increase EPS secretion. Alternatively, these ions could act as cofactors for key enzymes essential to EPS production.17 Also this finding suggests the role of metal ions, glucose content, and nitrogen sources in the optimization.25 However, the OVAT optimization technique played a significant role in selecting various culture conditions and growth components that improved the yield of EPS produced by Fusarium sp. GloS2.
From the FTIR data, it is anticipated that the EPS contains different chemical groups. The occurrence of these distinct chemical groups influences the bioactivities of EPS.48 XRD results revealed the crystalline structure of the EPS. These findings are consistent with prior studies,49 indicating that biopolymers have crystalline areas, even though most biopolymers are characterized as amorphous in the literature.
According to GC-MS analysis, EPS were shown to be heteropolysaccharides consisting of monosaccharides. Three distinct forms of the monosaccharides, erythrose, fructose, and mannose, were discovered to be present in Fusarium sp. GloS2 (molecular weight ~ 1.12 x 105 Da). The unique configuration of EPS frequently determines their bioactivity. The endophytic fungi Colletotrichum alatae LCS1 and Fusarium sp. A14 has been shown to possess similar heteropolysaccharides (molecular weight ~3.29 x 105 Da and ~ 2.4 x 3 104 Da), which comprise similar kinds of sugar moieties. These heteropolysaccharides demonstrate significant bioactivity.25,35 Moreover, both proton and carbon NMR spectra of EPS revealed the existence of a-glycosidic linkage along with galacturonic acid residues.36 Also, this finding highlighted the complex and unique structure of EPS with notable b-D-galactopyranosyl moieties and substituted glucose residues. A recent study showed that exopolysaccharides of Lactobacillus delbrueckii ssp. bulgaricus exhibited strong antioxidant activity mainly due to its high galactose content. Glycosidic linkage and branching patterns enhanced this activity by improving flexibility and water solubility. These characteristics are crucial for effective radical scavenging.50 Research showed that antioxidant activity of exopolysaccharides might depend on low molecular weight, high branch degree, different linkages and complex conformation. In vitro antioxidant activity revealed that polysaccharides (extracted from Tricholoma lobayense) made up of galactopyranosyl moieties exhibited antioxidant activities.51
The flocculation activity of the EPS is concentration-dependent (Supplementary Table 3). A comparable level of flocculation activity was observed for EPS isolated from MRL SM 8 (Fusarium mangiferae).52 Functional group and molecular weight significantly improve flocculation, and higher doses can decrease the efficiency of flocculants.53 A comparable progression in flocculation activity with increasing EPS dosage was previously found for Lysinibacillus fusiformis KMNTT 10.54 The current flocculation study revealed that the EPS generated by Fusarium strains had a wide range of flocculation efficiencies, making them potentially useful in the sludge treatment, food, and mining industries. The WHC capacity of GloS2 was high, surpassing other EPS sources like Bacillus licheniformis and Gloeocapsa gelatinosa.19,55
DPPH radical and H2O2 scavenging assays assess the oxidative stress resistance ability of natural substances.56 Previous research revealed that EPS derived from Fusarium solani SD5 has considerable DPPH radical scavenging ability.57 Isolated EPS was therefore predicted to donate hydrogen ions, resulting in the decolourization of violet-coloured DPPH to a colourless product.57 The superoxide radical scavenging capacity is also important in antioxidant research.56 Recent discoveries showed that different kinds of exopolysaccharides isolated from endophytic fungi exhibited antioxidative activities, e.g. Cyclocarva paliurus,58 Armillaria mellea,59 Fusarium equiseti,60 Fusarium culmorum,61 Pilidiella guizhouensis.62 Different types of approaches are taken to fight against problems of malignancy, tumor, and cytotoxicity, and EPS from GloS2 is just a nature-based sustainable solution to that.63 The antioxidant capacity of GloS2, as determined by DPPH, ABTS, H2O2, and FRAP radical capturing, was contrasted against that of ascorbic acid in this study. The remarkable IC50 value of EPS shows its potential as a substitute for synthetic antioxidants.
The EPS from GloS2 is found to be an effective in vitro antioxidant (in terms of free radical scavenging). Still, its in vivo activity must be assessed for further therapeutic use and dose selection. Their oral dosage-based application in the liver, and kidney tissues of Wistar rats or Swiss albino mice as per the body weight will elucidate the potent radical scavenging activity in animal systems and will ease up things for further pharmaceutical utilisation. Antioxidative potentialities of two novel EPS from endophytic fungi of medicinal plants exhibit strong repro-protective activity in vivo and support our present investigational outcomes.64
Polysaccharides possess distinct therapeutic qualities, including antibacterial, antioxidant, anticancer, and antiproliferative effects. This study revealed that Endophytic Fusarium sp. GloS2 exhibits a diverse composition of EPSs comprising various monosaccharide units, as well as in vitro antioxidative properties. However, further in vivo testing is essential to accurately ascertain its antioxidative potential. These findings hold significant promise for the pharmaceutical industry. Moreover, they fuel the enthusiasm of endophyte biologists to explore novel, naturally biodiverse regions or biomes in search of such unique endophytic fungi. These fungi serve as effective sources of functional EPS, opening up new avenues for research and application.
Additional file: Additional Table S1-S4 and Figure S1.
ACKNOWLEDGMENTS
The authors acknowledge the USIC (University Sophisticated Instrument Center) of Vidyasagar University for support with instrumental facilities.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
SS and HKS performed the work. SS and HKS wrote the manuscript. SC and DB supervised the project and critically reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This research work was financially supported by the Savitribai Jyotirao Phule Single
Girl Child Fellowship of UGC- Fellowship Number-(ID:202223-UGCES-22-GE-WES-F-SJSGC-3863).
DATA AVAILABILITY
The datasets generated and/or analysed during the current study are available in the NCBI GenBank Acc. No.- OP379518 (https://www.ncbi.nlm.nih.gov/).
ETHICS STATEMENT
Not applicable.
- Zhu K, Zhang Y, Nie S, et al. Physicochemical properties and in vitro antioxidant activities of polysaccharide from Artocarpus heterophyllus Lam. pulp. Carbohydr Polym. 2017;155:354-361.
Crossref - Stone JK, Bacon CW, White Jr JF. An overview of endophytic microbes: endophytism defined. Microbial Endophytes. 2000;25:17-44.
Crossref - Jomova K, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, Valko M. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol.2024;98(5):1323-1367.
Crossref - Prakash CR, Raja S, Saravanan G, Kumar PD, Selvam TP. Synthesis and evaluation of antioxidant activities of some novel isatin derivatives and analogs. Asian Journal of Research in Pharmaceutical Science. 2011;1(4):140-143.
- Selvam TP, Kumar PV, Kumar AS. Synthesis and anti-oxidant activity of novel 6, 7, 8, 9 tetra hydro-5H-5-(2′-hydroxy phenyl)-2-(4′-substituted benzylidine)-3-(4-nitrophenyl amino) thiazolo quinazoline derivatives. Res Biotechnol. 2010;26:1(1):38-48.
- Kalimuthu AK, Parasuraman P, Sivakumar P, et al. In silico, in vitro screening of antioxidant and anticancer potentials of bioactive secondary metabolites from an endophytic fungus (Curvularia sp.) from Phyllanthus niruri L. Environ Sci Pollut Res. 2022;29(32):48908-48925.
Crossref - Khatua S, Paul S, Acharya K. Mushroom as the potential source of new generation of antioxidant: a review. Res J Pharm Technol. 2013;6(5):496-505.
- Calegari GC, Santos VAQ, Teixeira SD, Barbosa-Dekker AM, Dekker RFH, da Cunha MAA. Sulfonation of (1→6)-b-D-glucan (lasiodiplodan) and its antioxidant and antimicrobial potential. J Pharm Pharmacol. 2017;5(12):850-863.
Crossref - Jiang JQ. The role of coagulation in water treatment. Curr Opin Chem Eng. 2015;8:36-44.
Crossref - Nouha K, Kumar RS, Balasubramanian S, Tyagi RD. Critical review of EPS production, synthesis and composition for sludge flocculation. J Environ Sci. 2018;66:225-245.
Crossref - Schulz B, Wanke U, Draeger S, Aust HJ. Endophytes from herbaceous plants and shrubs: effectiveness of surface sterilization methods. Mycol Res. 1993;97(12):1447-1450.
Crossref - White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Academic Press, Inc. 1990:315-322.
Crossref - Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38(7):3022-3027.
Crossref - Rasulov BA, Yili A, Aisa HA. Biosorption of Metal Ions by Exopolysaccharide Produced by Azotobacter chroococcum XU1. Journal of Environmental Protection.2013;04(09):989-993.
Crossref - DuBois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28(3):350-356.
Crossref - Lowery GH, Newman RJ. Notes on the ornithology of southeastern San Luis Potosi. The Wilson Bulletin. 1951;63(4):315-22.
- Mahapatra S, Banerjee D. Optimization of a bioactive exopolysaccharide production from endophytic Fusarium solani SD5. Carbohydr Polym. 2013;97(2): 627-634.
Crossref - Santra HK, Banerjee D. Broad spectrum bioactivity of a novel b-glucan rich heteropolysaccharide, Pestalopine isolated from endophytic fungi Pestalotiopsis chamaeropsis CEL6. Arch Microbiol. 2023;205(8):284.
Crossref - Insulkar P, Kerkar S, Lele SS. Purification and structural-functional characterization of an exopolysaccharide from Bacillus licheniformis PASS26 with in-vitro antitumor and wound healing activities. Int J Biol Macromol. 2018;120(Pt B):1441-1450.
Crossref - De B, Nag G. Metabolic diversity in the grains of Indian varieties of rice. C R Biol. 2014;337(4):283-293.
Crossref - Santra HK, Banerjee D. Drought alleviation efficacy of a galactose rich polysaccharide isolated from endophytic Mucor sp. HELF2: A case study on rice plant. Front Microbiol. 2023;13:1064055.
Crossref - Mahapatra S, Banerjee D. Structural elucidation and bioactivity of a novel exopolysaccharide from endophytic Fusarium solani SD5. Carbohydr Polym. 2012;90(1):683-689.
Crossref - Devi PB, Kavitake D, Shetty PH. Physico-chemical characterization of galactan exopolysaccharide produced by Weissella confusa KR780676. Int J Biol Macromol. 2016;93(Part A):822-828.
Crossref - Ahmed Z, Wang Y, Anjum N, Ahmad A, Khan ST. Characterization of exopolysaccharide produced by Lactobacillus kefiranofaciens ZW3 isolated from Tibet kefir-Part II. Food Hydrocolloids. 2013;30(1):343-350.
Crossref - Santra HK, Banerjee D. Production, optimization, characterization and drought stress resistance by b-Glucan-Rich heteropolysaccharide from an endophytic fungi Colletotrichum alatae LCS1 isolated from clubmoss (Lycopodium clavatum). Front Fungal Biol. 2022;2:796010.
Crossref - Blois MS. Antioxidant determinations by the use of a stable free radical. Nature. 1958;181(4617):1199-1200.
Crossref - Rajurkar NS, Hande SM. Estimation of phytochemical content and antioxidant activity of some selected traditional Indian medicinal plants. Indian J Pharm Sci. 2011;73(2):146-151.
Crossref - Winterbourn CC. Myeloperoxidase as an effective inhibitor of hydroxyl radical production. Implications for the oxidative reactions of neutrophils. J Clin invest. 1986;78(2):545-550.
Crossref - Oyaizu M. Studies on products of browning reaction– antioxidative activities of products of browning reaction prepared from glucosamine.The Japanese Journal of Nutrition and Dietetics. 1986;44(6):307-315.
Crossref - Chambi D, Romero-Soto L, Villca R, et al. Exopolysaccharides production by cultivating a bacterial isolate from the hypersaline environment of Salar de Uyuni (Bolivia) in pretreatment liquids of steam-exploded quinoa stalks and enzymatic hydrolysates of curupau sawdust. Fermentation. 2021;7(1):33.
Crossref - Chawananorasest K, Saengtongdee P, Kaemchantuek P. Extraction and characterization of tamarind (Tamarind indica L.) seed polysaccharides (TSP) from three difference sources. Molecules. 2016;21(6):775.
Crossref - Romdhane MB, Haddar A, Ghazala I, Jeddou KB, Helbert CB, Ellouz-Chaabouni S. Optimization of polysaccharides extraction from watermelon rinds: Structure, functional and biological activities. Food Chem. 2017;216:355-364.
Crossref - Hu X, Li D, Qiao Y, et al. Purification, characterization and anticancer activities of exopolysaccharide produced by Rhodococcus erythropolis HX-2. Int J Biol Macromol. 2020;145:646-654.
Crossref - Sahana TG, Rekha PD. A bioactive exopolysaccharide from marine bacteria Alteromonas sp. PRIM-28 and its role in cell proliferation and wound healing in vitro. Int J Biol Macromol. 2019;131:10-8.
Crossref - Pan F, Hou K, Li DD, Su TJ, Wu W. Exopolysaccharides from the fungal endophytic Fusarium sp. A14 isolated from Fritillaria unibracteata Hsiao et KC Hsia and their antioxidant and antiproliferation effects. J Biosci Bioeng. 2019;127(2):231-240.
Crossref - Corsaro MM, De Castro C, Naldi T, Parrilli M, Tomas JM, Regue M. 1H and 13C NMR characterization and secondary structure of the K2 polysaccharide of Klebsiella pneumoniae strain 52145. Carbohydr Res. 2005;340(13):2212-2217.
Crossref - Chen Y, Mao WJ, Yan MX, et al. Purification, chemical characterization, and bioactivity of an extracellular polysaccharide produced by the marine sponge endogenous fungus Alternaria sp. SP-32. Mar Biotechnol. 2016;18(3):301-313.
Crossref - Peng H, Wang N, Hu Z, et al. Physicochemical characterization of hemicelluloses from bamboo (Phyllostachys pubescens Mazel) stem. Industrial Crops and Products. 2012;37(1):41-50.
Crossref - Nie SP, Cui SW, Phillips AO, et al. Elucidation of the structure of a bioactive hydrophilic polysaccharide from Cordyceps sinensis by methylation analysis and NMR spectroscopy. Carbohydr Polym. 2011;84(3):894-899.
Crossref - Mende S, Rohm H, Jaros D. Influence of exopolysaccharides on the structure, texture, stability and sensory properties of yoghurt and related products. Int Dairy J. 2016;52:57-71.
Crossref - Zhu FM, Du B, Li J. Effect of ultrafine grinding on physicochemical and antioxidant properties of dietary fiber from wine grape pomace. Food Sci Technol Int. 2014;20(1):55-62.
Crossref - Chen C, Xin K, Liu H, et al. Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat. Sci Rep. 2017;7(1):41564.
Crossref - Sun L, Yang Y, Wang R, et al. Effects of exopolysaccharide derived from Pantoea alhagi NX-11 on drought resistance of rice and its efficient fermentation preparation. Int J Biol Macromol. 2020;162:946-55.
Crossref - Pham PL, Dupont I, Roy D, Lapointe G, Cerning J. Production of exopolysaccharide by Lactobacillus rhamnosus R and analysis of its enzymatic degradation during prolonged fermentation. Appl Environ Microbiol. 2000;66(6):2302-2310.
Crossref - Gao Z, Yang J, Wu J, et al. Structural characterization and in vitro evaluation of the prebiotic potential of an exopolysaccharide produced by Bacillus thuringiensis during fermentation. LWT. 2022;163:113532.
Crossref - Mahapatra S, Banerjee D. Production and structural elucidation of exopolysaccharide from endophytic Pestalotiopsis sp. BC55. Int J Biol Macromol. 2016;82:182-91.
Crossref - Prete R, Alam MK, Perpetuini G, Perla C, Pittia P, Corsetti A. Lactic acid bacteria exopolysaccharides producers: a sustainable tool for functional foods. Foods. 2021;10(7):1653.
Crossref - Mao YH, Song FL, Xu YX, et al. Extraction, characterization, and platelet inhibitory effects of two polysaccharides from the Cs-4 fungus. Int J Mol Sci. 2022;23(20):12608.
Crossref - Philippini RR, Martiniano SE, Marcelino PRF, Chandel AK, dos Santos JC, Da Silva SS. Production of b-glucan exopolysaccharide lasiodiplodan by Lasiodiplodia theobromae CCT 3966 from corn bran acid hydrolysate. Appl Microbiol Biotechnol. 2021;105(6):2319-2332.
Crossref - Tang W, Dong M, Wang W, et al. Structural characterization and antioxidant property of released exopolysaccharides from Lactobacillus delbrueckii ssp. bulgaricus SRFM-1. Carbohydr Polym. 2017;173:654-664.
Crossref - Chen Y, Li XH, Zhou LY, et al. Structural elucidation of three antioxidative polysaccharides from Tricholoma lobayense. Carbohydr Polym. 2017;157:484-492.
Crossref - Mittholiya S, Meena BR, Chittora D, Yadav J, Jain T, Sharma K. Exopolysaccharides production and characterization of Fusarium mangieferae from mango (Mangifera indica) with some commercial potential applications. Journal of Postharvest Technology. 2022;10(3):102-112.
Crossref - Biswas JK, Banerjee A, Sarkar B, et al. Exploration of an extracellular polymeric substance from earthworm gut bacterium (Bacillus licheniformis) for bioflocculation and heavy metal removal potential. Appl Sci. 2020;10(1):349.
Crossref - Mathivanan K, Chandirika JU, Vinothkanna A, Govindarajan RK, Meng D, Yin H. Characterization and biotechnological functional activities of exopolysaccharides produced by Lysinibacillus fusiformis KMNTT-10. J Polym Environ. 2021;29(6):1742-51.
Crossref - Gongi W, Pinchetti JLG, Cordeiro N, Ouada HB. Extracellular polymeric substances produced by the Thermophilic Cyanobacterium Gloeocapsa gelatinosa: Characterization and assessment of their antioxidant and metal-chelating activities. Marine Drugs. 2022;20(4):227.
Crossref - Chen Y, Xue Y. Purification, chemical characterization and antioxidant activities of a novel polysaccharide from Auricularia polytricha. Int J Biol Macromol. 2018;120(Pt A):1087-1092.
Crossref - Mahapatra S, Banerjee D. Evaluation of in vitro antioxidant potency of exopolysaccharide from endophytic Fusarium solani SD5. Int J Biol Macromol. 2013;53:62-66.
Crossref - Tao X, Chen X, Dong R, et al. Characterization and antioxidant properties of three exopolysaccharides produced by the Cyclocarya paliurus endophytic fungus. Int J Biol Macromol. 2024;271(Pt 1):132110.
Crossref - Su P, Qiu H, Liang L, et al. The antioxidant activity of polysaccharides from Armillaria gallica. Front Nutr. 2024;14(11):1277877.
Crossref - Prathyusha AMVN, Sheela GM, Bramhachari PV. Chemical characterization and antioxidant properties of exopolysaccharides from mangrove filamentous fungi Fusarium equiseti ANP2. Biotechnol Rep. 2018;19:e00277.
Crossref - Jaroszuk-Scisel J, Nowak A, Komaniecka I, et al. Differences in production, composition, and antioxidant activities of exopolymeric substances (EPS) obtained from cultures of endophytic Fusarium culmorum strains with different effects on cereals. Molecules. 2020;25(3):616.
Crossref - Zhang J, Yang B, Chen H. Identification of an endophytic fungus Pilidiella guizhouensis isolated from Eupatorium chinense L. and its extracellular polysaccharide. Biologia. 2020;75:1707-1715.
Crossref - Theivendren P, Kunjiappan S, Hegde YM, et al. Importance of protein kinase and its inhibitor: a review. Protein Kinases: Promising Targets for Anticancer Drug Research. IntechOpen. 2021;22:75-100.
Crossref - Saha S, Ghosh A, Santra HK, Banerjee D, Chattopadhyay S. Corrective role of endophytic exopolysaccharides from Clerodendrum infortunatum L. on arsenic-induced ovarian steroidogenic dysfunction and associated inflammatory responses. Int J Biol Macromol. 2024;282(Pt 1):136795.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.