


ISSN: 0973-7510
E-ISSN: 2581-690X
Microorganisms and plants can produce indole-3-acetic acid (IAA) by mechanisms that either involve tryptophan or do not involve tryptophan. The control of Zone of inhibition (ZOI) ranged from 1.2 cm to 1.0 cm. A promising antagonistic isolate, Streptomyces sp. VSMKU1027, exhibited robust antagonistic activity against two significant soil-borne phytopathogenic fungi, namely Rhizoctonia solani and Fusarium oxysporum, outperforming the control. The morphology of both R. solani and F. oxysporum, including their sclerotium and spores, became condensed and indistinct, and mycelial disintegration was observed due to the action of VSMKU1027, in contrast to the control. The isolate significantly produced antimicrobial traits and hydrolytic enzymes except hydrogen cyanide and cellulase. Furthermore, the promising isolate VSMKU1027 was identified as Streptomyces sp. based on its morphological, physiological, and biochemical characteristics. In comparison to the control, the isolate VSMKU1027 demonstrated increased synthesis of IAA and the hydrolytic enzyme protease with the corresponding substrates. The production of IAA was optimized on the sixth day, at 35°C and pH 6.5. The highest generation of IAA was recorded, with an ISP2 and 0.4% concentration of L-tryptophan.
Streptomyces sp, Antifungal Activity, Antimicrobial Traits, Hydrolytic Enzymes, Pathogens, Optimization of IAA
Gram-positive bacteria known as actinomycetes have genomes that are largely composed of G + C nucleotides. This characteristic allowed them to endure harsh conditions and thrive in highly competitive environments. Like other plant growth-promoting rhizobacteria (PGPR), Streptomyces can enhance plant growth and nutrient uptake through their metabolites or specialized biological activities. These activities include the production of auxins and ammonia, iron chelation by bacterial siderophores, and phosphate solubilization.1,2 Plant growth-promoting rhizobacteria (PGPR) employ the synthesis of antibiotics, lytic enzymes, volatile compounds, and siderophores as key mechanisms to inhibit pathogens. These processes are implicated in the suppression of plant diseases.3 Numerous reports indicate that Streptomycetes in rhizospheric and endophytic soils have the capability to produce indole acetic acid, promoting plant growth and development. Research conducted to evaluate the antagonistic and plant growth-promoting effects of endophytic and soil actinobacteria yielded similar results.4,5 Streptomyces spp. are recognised for their production of hydroxamate-type siderophores, which compete with iron in the rhizospheric soil environment, thereby inhibiting the growth of phytopathogens. Furthermore, other groups of soil microbes, including Streptomyces spp., Trichoderma spp., Pseudomonas spp., and Bacillus spp., are proficient in producing diverse antimicrobial compounds such as phenazine and 2,4-diacetylphloroglucinol.6-10 Likewise, these microbes produce hydrolytic enzymes like chitinase, glucanase, and protease, which regulate various soil-borne phytofungal pathogens.11,12 This indicates that the production of siderophores by the isolate VSMKU1027 may play a role in inhibiting bacterial and fungal pathogens, thereby indirectly promoting plant growth.13,14
The rhizosphere, which is defined as the area surrounding a plant’s roots, hosts a diverse array of nutrient and energy sources, including carbon and organic compounds released by the roots themselves. Many soil-dwelling microorganisms, including PGPR like Streptomyces sp.,3 Pseudomonas sp.15,16 and Bacillus sp.11 enhance plant growth directly by providing and facilitating the uptake of various nutrients and modulating phytohormone levels. Plant growth-promoting rhizobacteria (PGPR) are naturally occurring, free-living bacteria that colonize the rhizosphere and enhance soil fertility, increase yield, promote plant growth, mitigate pathogens, as well as biotic and abiotic stressors.17,18 Plant growth-promoting rhizobacteria (PGPR) produce phytohormones like indoleacetic acid (IAA), cytokinins, and gibberellins,19 facilitate inorganic phosphate solubilization,20 engage in symbiotic nitrogen fixation,21 and generate antibacterial and antifungal compounds.22-24 These substances aid plants in suppressing phytopathogenic microorganisms.
Actinomycetes are extensively distributed in the plant rhizosphere and produce a variety of agroactive compounds. Due to their strong antibacterial properties and dominant sporophytic nature in soil, this group of bacteria has recently attracted considerable attention as plant growth promoters.25 Actinobacteria are capable of actively colonizing plant root systems, secreting various hydrolytic enzymes to break down a range of biopolymers, and forming spores to endure adverse environmental conditions.26 Actinobacteria, particularly Streptomyces, have demonstrated strong biocontrol activity against a wide range of phytopathogens.27 Additionally, actinobacteria can produce phytohormones like IAA, siderophores, and soluble phosphate, thereby promoting plant growth.20 Since the 1940s, actinomycetes have primarily been utilised in the pharmaceutical industry, but only a limited number have been developed into commercial products for agricultural use.28 Streptomycetes were once considered merely free-living soil organisms, but recent research is uncovering the importance of their complex interactions with plants and other organisms.29
Keyeo et al.30 reported that IAA, a naturally occurring auxin phytohormone containing an indole ring, is a metabolic byproduct of microorganisms’ processing of L-tryptophan, Among the plant growth-promoting bacteria are actinomycetes and fungi, which boost plant growth by producing IAA through mechanisms dependent on L-tryptophan.31 Some studies indicate that Streptomyces sp. is the leading genus among actinomycetes that produce IAA.4,32 IAA influences various physiological and developmental processes in plants, including embryogenesis, organogenesis, vascular differentiation, root and shoot development, trophic growth, and fruit development. In this context, the current study focused on the biological control of soil-borne phytofungal pathogens and optimization of IAA production by Streptomyces sp. VSMKU1027. This potential microbe VSMKU1027 was identified up to genus level based on its morphological and physiological characteristics, as well as the detection of genomic DNA and 16S rDNA encoding gene. Additionally, further characterization of IAA is ongoing.
Streptomyces sp. VSMKU1027 and soil-borne phytofungal pathogens
All the phytofungal pathogens, including Fusarium oxysporum and Rhizoctonia solani, along with the selected isolate VSMKU1027, were sourced from the Department of Microbial Technology at the School of Biological Sciences at Madurai Kamaraj University in Madurai, Tamil Nadu, India. Potato dextrose agar medium (composition: potato-200 g/L, glucose-20 g/L, agar-15 g/L) was used to culture R. solani and F. oxysporum. Plates were incubated at 28°C for three to ten days, and pathogen stock cultures were maintained on PDA slants at 4°C. For the isolation of VSMKU1027, ISP2 agar medium (composition: yeast extract-4 g/L, malt extract-10 g/L, dextrose-4 g/L, agar-20 g/L) was used with duplicate plates incubated for seven to ten days at 28°C. Actinomycete colonies were selected based on their morphological characteristics and purified using ISP-2 agar plates.33
Antagonistic activity of Streptomyces VSMKU1027 against phytofungal pathogens
The dual culture assay evaluated the antagonistic activity of VSMKU1027 against R. solani and F. oxysporum15 on PDA medium. VSMKU1027, grown for four days on ISP-2 agar, was streaked along the periphery of each PDA petri plates, positioned 3 cm away from the fungal disc. Following three and five days of incubation, test and control plates containing R. solani and F. oxysporum were inspected, and the growth of the fungal mycelium was measured using a meter scale.
Interaction between VSMKU1027 and phytofungal pathogens
The interaction between Streptomyces sp. VSMKU1027 and phytofungal pathogens such as R. solani and F. oxysporum was examined under a light microscope (Labomed, USA) to assess antifungal activity. Both pathogens and the antagonist isolate VSMKU1027 were co-inoculated on PDA plates for up to 5 days. Following the incubation period, a small portion of mycelium from both test and control plates was taken from the zone of inhibition interaction and stained with lactophenol cotton blue (Himedia, India) on a sterile glass slide. The slides were then observed under a microscope at 40X magnification to evaluate the effectiveness of VSMKU1027 against the phytofungal pathogens.34
Characterization of VSMKU1027
The chosen isolate, VSMKU1027, was obtained from the Department of Microbial Technology, School of Biological Sciences, Madurai Kamaraj University, Madurai-625021, Tamil Nadu, India Morphological observations and biochemical examinations were conducted to characterise various features such as Gram staining, morphology, catalase and oxidase activities, citrate and urea utilisation, nitrate reduction, indole synthesis, MR-VP tests, TSI reactions, carbohydrate utilisation, and nitrogen source utilisation for the identification of VSMKU1027. The isolate VSMKU1027 was kept in long-term storage at
-80°C with 30% glycerol stock for further research endeavors.8
Production of antimicrobial traits
Production of Siderophore
The positive control in this experiment involved using Pseudomonas fluorescens CHAO strain, while the ISP2 broth medium supplemented with 2 µl mol filter-sterilized ferrous sulfate served as the negative control. VSMKU1027 cells were harvested from the medium by centrifugation at 4,000 g for 10 minutes. Subsequently, 1 mL of filter-sterilised supernatant was mixed with 1 ml of chrome-azurol S solution (Himedia, India). The presence of siderophores was indicated by a colour change from blue to reddish-brown within 15 minutes of incubation.34
Hydrogen cyanide (HCN) production
VSMKU1027 was cultivated in nutrient sucrose medium (NSM) supplemented with 4.4% glycine as a fresh isolate. The petri dish lid was covered with sterilized Whatman No. 1 filter paper that had been soaked in a solution of 1% picric acid and 2% sodium carbonate. The Petri dish plates were firmly sealed with parafilm and incubated for 48 hours to stop gas exchange from occurring inside of them. The creation of HCN is indicated by the filter paper changing from yellow to orange in depiction.35
Phosphate solubilization
The isolate VSMKU1027 was inoculated punctiformly on Pikovskaya’s agar12 with minor modifications. For 48 hours, the plates were incubated at 28 ± 2°C. When compared to the control, the formation of a clear zone surrounding the colony was thought to be beneficial for the solubilization of insoluble phosphate.
Hydrolytic enzyme production
The young and pure isolate VSMKU1027 was streak on the nutrient agar medium supplemented with 1% of respective substrates for the production of hydrolytic enzymes like chitinase, cellulase, gelatinase, protease, amylase, and pectinase assay was performed.36
Optimization of Indole-3-acetic acid production
The production of plant growth hormone IAA was optimized from the selected isolate VSMKU1027 by various abiotic stresses in this present study.
Production of IAA in different L-tryptophan concentrations, days, and medium
Selected isolate VSMKU1027 observed the production of various L-tryptophan concentrations and days on IAA production. For 10 days, the isolate VSMKU1027 was cultured in 50 milliliters of ISP-2 broth supplemented with varying concentrations of tryptophan (about 1 to 1.5%), at pH 7.0, 37°C, and 120 revolutions per minute in a shaker.4 Several media, including ISP2, yeast malt broth, minimum medium, Bennett’s broth, and Tryptic soy broth, were used to optimize the generation of IAA by VSMKU1027. After incubating at 35°C and 120 rpm for seven days, the formation of IAA was detected.
Production of IAA at different temperatures and pH
To assess the effect of temperature and pH, Streptomyces sp. VSMGT1027 produced IAA across a range of temperatures from 5°C and 40°C and pH values from 4 and 10. The isolated was cultured for ten days in 50 µl of ISP-2 broth, supplemented with approximately 0.7% tryptophan, at 37°C and pH 7.0, on a shaker set to 120 rpm.12
Antagonistic potential of Streptomyces sp. VSMKU1027
Numerous bacterial and fungal diseases that affect both flowering and nonflowering plants result in large losses in food production globally, with major economic consequences. Plant pathogenic fungi are the primary cause of agricultural losses among the organisms responsible for these illnesses,37 as they infect a diverse variety of cultivated plants. In light of this, the current investigation examined the antifungal activity of an indigenous isolate of Streptomyces sp. VSMKU1027, which demonstrated varying degrees of zone of inhibition in comparison to the control against two economically significant soil-borne fungal pathogens, R. solani and F. oxysporum (Table 1). Both soil fungi are notable for being extensively dispersed diseases that can seriously harm economically important crops by producing, sheath blight, vascular wilt, root rot, fruit and flower rot, and finally plant death.7 Various Streptomyces spp. have been recognized as bio-control agents against numerous plant pathogens, demonstrating their capability to produce bioactive compounds that suppress or inhibit the mycelial growth of several fungi.38 Secondary metabolites may be used in addition to or instead of chemical pesticides, and their microbiological application in crop protection has gained significance in recent years.15
Table (1):
Antagonistic activity of Streptomyces sp. VSMKU1027 against soil-borne fungal plant pathogens
No. |
Soil-borne fungal plant pathogens |
Zone of Inhibition (cm) |
|---|---|---|
1 |
R. solani |
1.2±0.15 |
2 |
F. oxysporum |
1.5±0.15 |
Research has concentrated on employing Streptomyces spp. as biocontrol agents to manage various bacterial and fungal-induced plant diseases.33,37 Bacteria of the genus Streptomyces spp. can control a variety of bacterial and fungal plant diseases through diverse mechanisms. Their action mechanism has been described as the synthesis of volatile and non-volatile antibiotics, enzymes that break down cell walls, hyperparasitism on pathogenic organisms, growth stimulation in plants, and the creation of systemic resistance in the host plant.39
Interaction between Pathogens and Streptomyces sp. VSMKU1027
Given this context, the present study has investigated the antifungal activity of indigenous isolate Streptomyces sp. VSMKU1027, which exhibited significant inhibition against two economically important soil-borne fungal pathogens, R. solani and F. oxysporum, as evidenced by varying levels of zone of inhibition compared to the control (Table 1). Various Streptomyces spp. have been recognized as bio-control agents against numerous plant pathogens, demonstrating their capability to produce bioactive compounds that suppress or inhibit the mycelial growth of several fungi.39 Secondary metabolites may be used in addition to or instead of chemical pesticides, and their microbiological application in crop protection has gained significance in recent years.15
Mycelia of R. solani were collected from the outermost zone of a three-day-old dual culture. Light microscopy observations revealed that, unlike the control, the mycelia of R. solani had disintegrated, the mycelium exhibited swelling, and no fruiting bodies were formed on the R. solani test plant (Figure 1a, 1b, 1c and 1d). Moreover, the isolate VSMKU1027 significantly altered the morphology of F. oxysporum, encapsulated hyphae causing it to become extensively branched and coiled. Additionally, the control mycelium displayed a different shape, with the mycelial tips expanding to form a spherical structure (Figure 2).

Figure 1. Interaction between soil-born fungal plant pathogens and R. solani, F. oxysporum and Streptomyces sp. VSMKU1027 visualized under light microscope (Magnification at 20X); 1a: Compact mass of R. solani mycelium (Control plate), 1b: R. solani mycelium disintegration (Test plate). 1c: F. oxyporum mass and dense mycelium (Control plate); 1d: F. oxysporum mycelia and hyphae were encapsulated by Streptomyces sp. VSMKU1027 (Test plate)
The secretion of exogenous compounds, such as cell-wall degrading enzymes and antibiotics, which visibly harm and disintegrate R. solani hyphae under light microscopy, is believed to be responsible for inhibiting R. solani mycelial growth in the presence of VSMGT1014. Compared to the control, interactions between VSMGT1014 and the pathogen resulted in abnormalities such as deformation and swelling of hyphae.3,40 In contrast to the control, interactions between VSMGT1014 and the pathogen resulted in abnormalities such as morphological deformities and hyphal swelling. This observation aligns closely with previous research.37 In the current study, pathogens like R. solani and F. oxysporum were observed under the light microscope exhibiting dissolution along with some swelling.41 This phenomenon may be attributed to the action of lytic enzymes such as protease and chitinase, or to secondary metabolites produced by VSMGT1027.
Identification of selected isolates VSMKU1027
The isolate VSMKU1027’s peach color and rough surface on different ISP mediums. On the ISP2 medium, the isolate displayed a punch of aerial substrate mycelium, short-looped chain spores, and aerial mycelium. Under a light microscope (20X), morphological features of the isolate VSMKU1027 spore chains were observed to be short chains resembling the flexuous sporophores group of Streptomyces (Figure 2).
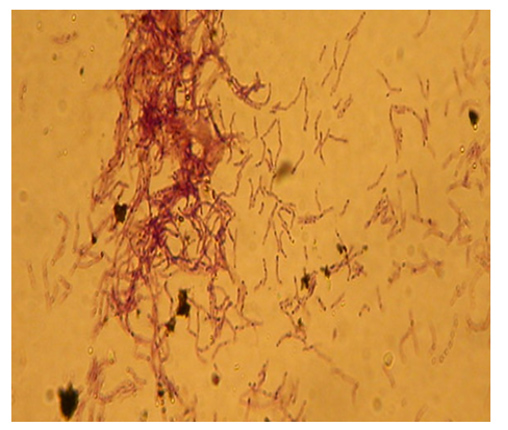
Figure 2. Spore morphology of VSMKU1027 visualized under Light microscope (Magnification at 20X)
In comparison to the control, the isolate VSMKU1027 displayed favorable results for the Gram’s reaction, catalase, and oxidase. In addition, urease synthesis, citrate utilization, casein, gelatin liquefaction, and gelatin hydrolysis were all significantly higher in the isolate VSMKU1027 than in the control. Regarding H2S, Voges Proskauer, methyl red, indole, nitrate reduction, arabinose, and mannitol utilization, the isolate VSMKU1027 exhibited a negative reaction. In comparison to the control, the strain VSMKU1027 used the majority of simple sugars, including lactose, glucose, fructose, and sucrose, as the only carbon source, excluding arabinose and mannitol. VSMKU1027 grew to its maximum on the ISP2 medium that contained glycerol and starch (Table 2). According to our findings, certain researchers discovered. The findings showed that the isolates is capable of using a wide range of substrates, including glycerol, starch, lactose, inositol, rhamnose, raffinose, and maltose.3,42-43 The actinobacterium’s smooth rough surface of the colony nature, form of the spores, and colony and mycelial nature were extremely consistent with other research findings.40,44
Table (2):
Streptomyces sp. VSMKU1027 morphological, physiological, and biochemical characteristics
Morphology and Biochemical analysis |
Results |
|---|---|
Nature of mycelium |
Aerial and thin clumped mycelium |
Sores |
L short-looped chain spores |
Colony morphology |
Peach color with Rough surface |
Grams staining |
+ |
Odor |
Soil |
Mycelium |
Thin clumped mycelium |
ISP2 medium |
Favorable medium for growth |
Texture |
Dry, Powdery |
Catalase |
+ |
Oxidase |
+ |
Voges Proskauer test |
– |
Methyl red |
– |
Indole |
– |
Urease production |
+ |
H2S production |
– |
Citrate utilization |
+ |
Casein |
+ |
Gelatin liquefaction |
+ |
Gelatin hydrolysis |
+ |
Starch hydrolysis |
+ |
Nitrate reduction |
– |
Spores |
+ |
Acid production with various sugars |
|
Sucrose |
+ |
Lactose |
+ |
Arabinose |
– |
Glucose |
+ |
Fructose |
+ |
Mannitol |
– |
Glycerol |
+ |
Given their ability to produce spores, these filamentous, Gram-positive rhizobacteria offer an added advantage for their potential as formulation-based agents to enhance plant growth.45-47 As reported by Harikrishnan et al.3 and Al-Askar et al.,48 specific Streptomyces species have been successfully developed as potential biocontrol agents and formulated to combat fungal phytopathogens across various crops, For instance, Mycostop, a commercially available wettable formulation containing spores and mycelium of Streptomyces griseoviridis, has been extensively used in North America and Europe to protect vegetables and ornamental crops.49
Production of antimicrobial traits and Hydrolytic enzyme by Streptomyces sp. VSMKU1027
Compared to the control, except for hydrogen cyanide, the isolate VSMKU1027 was much more capable of producing siderophores and phosphate solubilization. Analogously, isolate VSMKU1027 demonstrated exceptional hydrolytic enzyme synthesis, including chitinase, protease, gelatinase, amylase, and pectinase, in their respective substrate-amended mediums. In contrast to the control, the isolate VSMKU1027 did not produce cellulase (Table 2). The production of antibiotics, lytic enzymes, volatile compounds, and siderophores constitutes a primary mechanism through which plant growth-promoting rhizobacteria (PGPR) inhibit pathogens.50 Strain VSMGT010143 was found to produce IAA, siderophores, volatile compounds, and hydrolytic enzymes including chitinase, cellulase, protease, gelatinase, amylase, pectinase, and solubilized phosphorus. Moreover, rhizospheric Streptomyces spp. exhibit plant growth-promoting traits (PGPT), including enzyme secretion, organic acid production, phosphate solubilization, and siderophore synthesis. These PGPTs contribute to enhanced plant growth and the management of soil-borne bacterial and phytofungal diseases through mechanisms such as systemic acquired resistance and induced systemic resistance, providing protection against diverse biotic and abiotic stresses (Table 3). Previous studies have demonstrated that Streptomyces strains with these attributes can enhance plant growth. For instance, S. violaceusniger AC12AB was found capable of producing siderophores, nitrogen fixation, phosphate solubilisation, and intermediate amino acid production.51 In field trials, it significantly increased potato yields by up to 26.8%. Additionally, Chouyia et al.52 found that barley plants infected with S. roseocinereus MS1B15, which produces IAA, solubilize phosphate, and fixes nitrogen, exhibited substantial increases in shoot and spike length.
Table (3):
Production of Antimicrobial metabolites and hydrolytic enzymes by the isolate VSMKU1027
Antimicrobial metabolites and hydrolytic Production |
Results |
|---|---|
Siderophore |
+ |
Phosphate solubilization |
+ |
Hydrogen cyanide |
– |
IAA |
+ |
Chitinase |
+ |
Gelatinase |
+ |
Protease |
+ |
Amylase |
+ |
Cellulase |
– |
Pectinase |
+ |
Production of IAA in different L-tryptophan concentrations by Streptomyces sp. VSMKU1027
IAA is a significant component of the auxin family of plant hormones. Streptomyces species are the predominant IAA-producing actinobacteria that also enhance plant development.53-55 Actinomycetes and fungi are among the genera of bacteria that promote plant growth; they enhance plant growth by producing IAA through processes that rely on L-tryptophan. According to certain research, Streptomyces sp. is the predominant genus among actinomycetes that produce IAA. IAA influences various physiological and developmental processes in plants, including embryogenesis, organogenesis, vascular differentiation, root and shoot development, trophic growth, and fruit development. The availability of substrate(s), growth phase, and varied physicochemical circumstances impact IAA production, which varies greatly throughout species. The availability of substrate(s), growth phase, and varied physicochemical circumstances impact IAA production, which varies greatly among the various rhizobacterial species. All the habitat microbes were secreted IAA through a tryptophan-dependent biosynthesis pathway. Various concentrations of L- tryptophan ranging from 0.1 to 1% were tested for the production of IAA. Spectrophotometric analysis revealed a gradual increase in the IAA production with increasing L-tryptophan concentration. The highest IAA production, reaching 132.36 µg/mL, was observed when the medium contained 0.4% in ISP2 medium, surpassing other media and the control (Figure 3). This observation aligns with the understanding that actinomycetes can produce the phytohormone auxin IAA when provided with an appropriate precursor like L-tryptophan.55 In addition to that our results highly coincides the previous report. In our study, high concentrations of L-tryptophan supplementation adversely affected the level of IAA production. However, at low concentrations of L-tryptophan supplementation, IAA production could reach up to 27.3 µg/mL compared to the control.
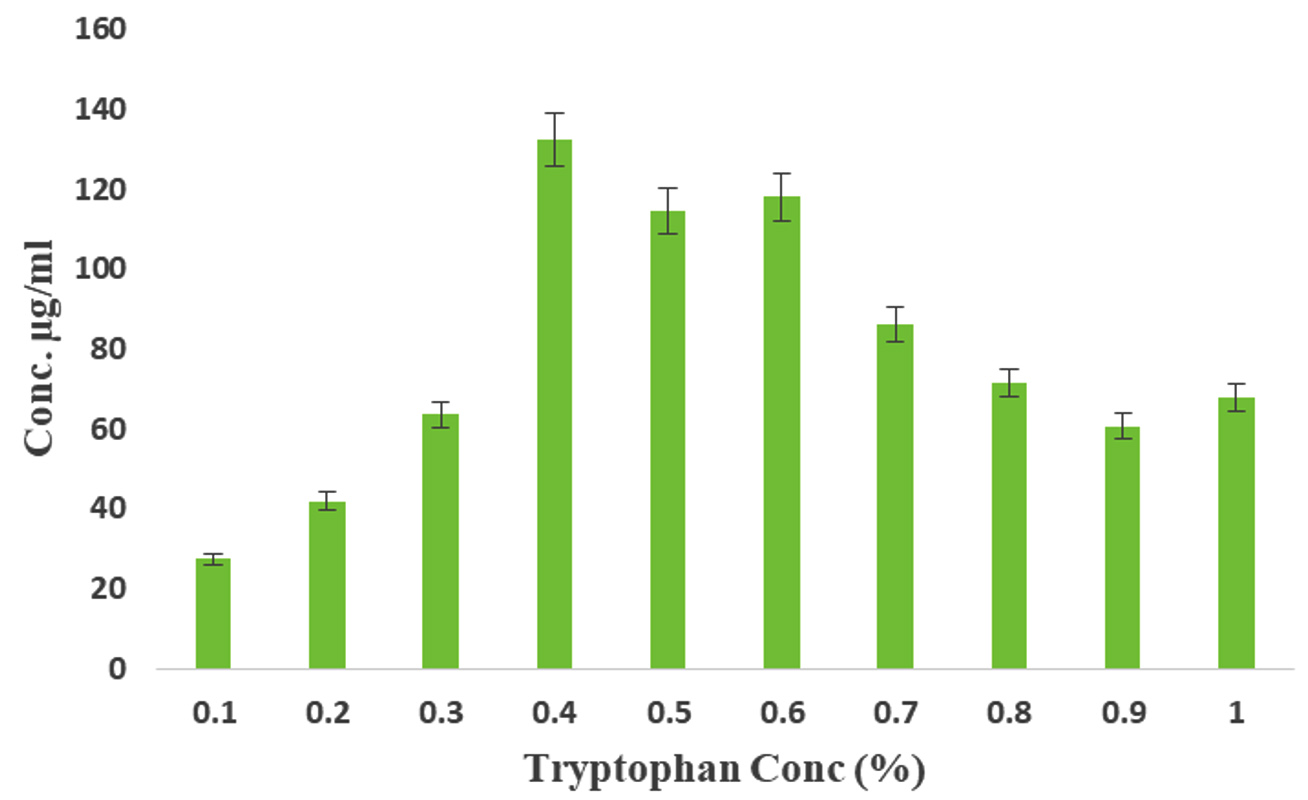
Figure 3. Production of IAA by Streptomyces sp. VSMKU1027 at various concentrations of L-Tryptophan
Production of IAA on different days, medium, temperature, and pH by Streptomyces sp. VSMKU1027
The effect of IAA production was monitored over a period of 10 days, with the highest production observed on the 6th day of incubation (93.4 µg/mL) (Figure 4). Kaur and Manhas56 reported that almost coincides of our results, because the maximum production IAA was 80.06 µg/mL by S. hydrogenans DH16 on 5th day. According to previous findings, the oxidase and peroxidase synthesis that degrades IAA may be the cause of the attenuation in the IAA levels.57 Likewise, among the five different media tested, ISP2 medium yielded the highest amount of IAA production (68.13 µg/mL) (Figure 5). In contrast, the lowest amount of IAA production, at 19.8 µg/mL, was observed in Bennett’s broth medium compared to the control. The maximum production of IAA by Streptomyces sp. VSMKU1027, reaching 66.5 µg/mL, was observed at pH 6.5. The results indicated a gradual increase in IAA production by VSMKU1027 from pH 4 to pH 6.5, whereas a significant decrease in production was noted at pH 8-10 compared to the control (Figure 6). Similarly, the highest IAA production (81.4 µg/mL) was recorded at 35°C. However the IAA production was slowly reduced at 40°C compared to control (Figure 7). The influence of pH on IAA synthesis demonstrated that S. hydrogenans DH16 produced the highest amount of IAA at a neutral pH, consistent with previous studies.54,58 Production decreases at acidic or highly alkaline pH levels because Streptomyces spp. are unable to thrive in such extreme pH conditions. The optimal temperature for IAA formation was found to be
30°C, aligning with Aldesuquy et al.59 study indicating maximal IAA production by Streptomyces spp. at 25-30°C. IAA produced by rhizobacteria primarily influences the root system, enhancing its size, weight, number of lateral roots, and soil contact area. This mechanism not only enhances plant development and yield but also facilitates nutrient uptake and acquisition in the soil.
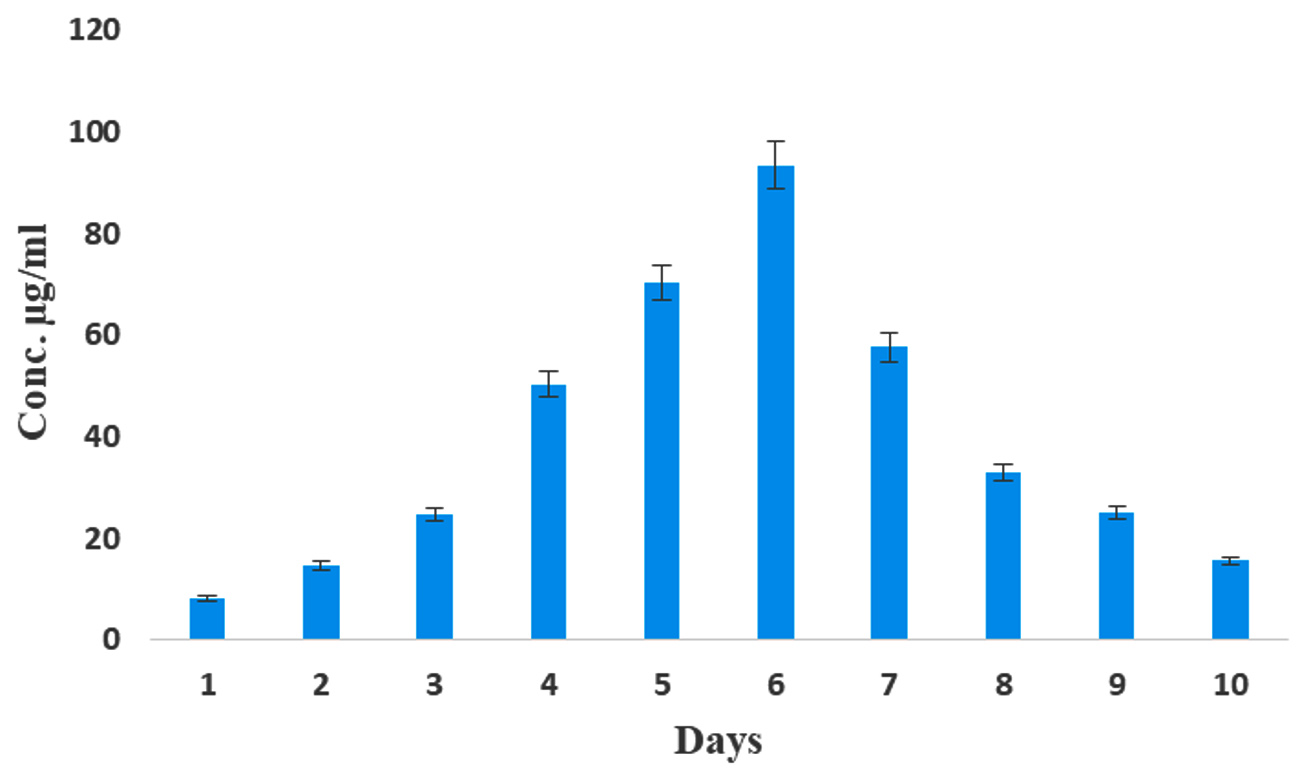 Figure 4. Production of IAA by Streptomyces sp. VSMKU1027 at various incubation days
Figure 4. Production of IAA by Streptomyces sp. VSMKU1027 at various incubation days
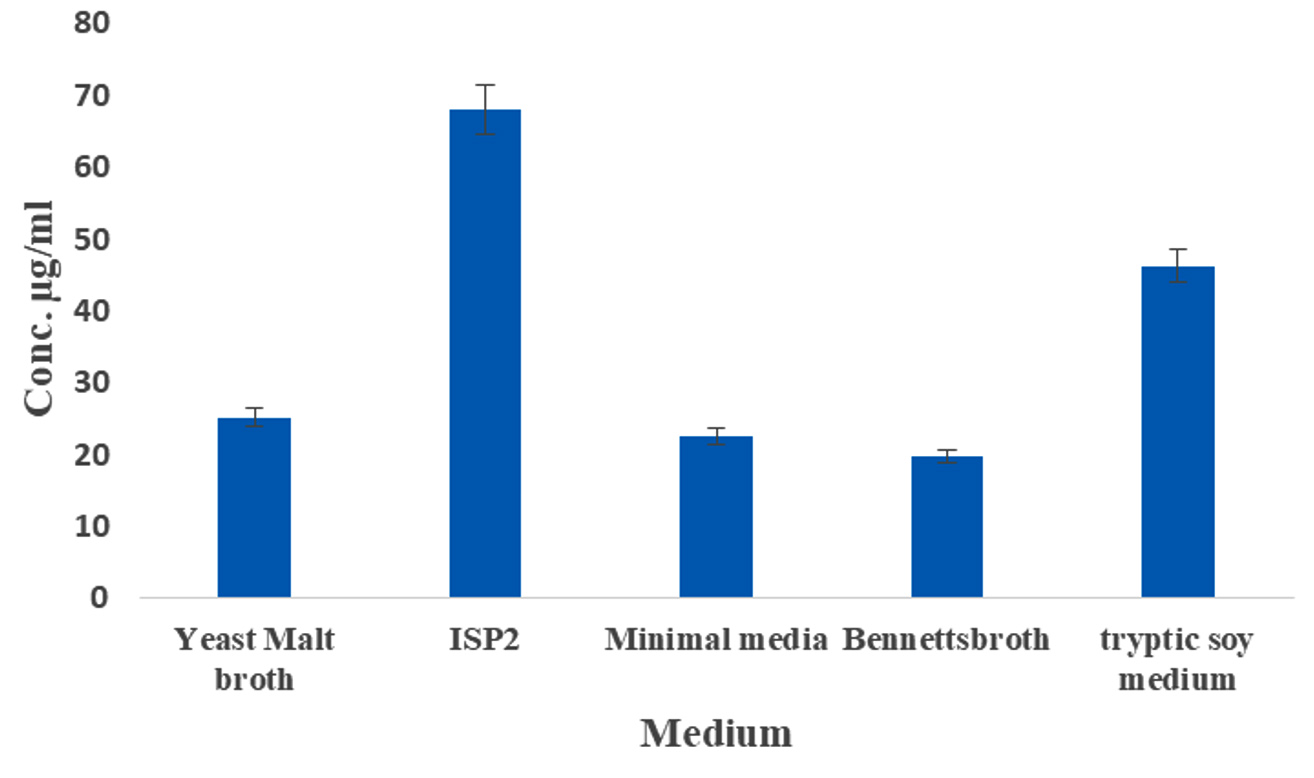 Figure 5. Production of IAA by Streptomyces sp. VSMKU1027 at different medium
Figure 5. Production of IAA by Streptomyces sp. VSMKU1027 at different medium
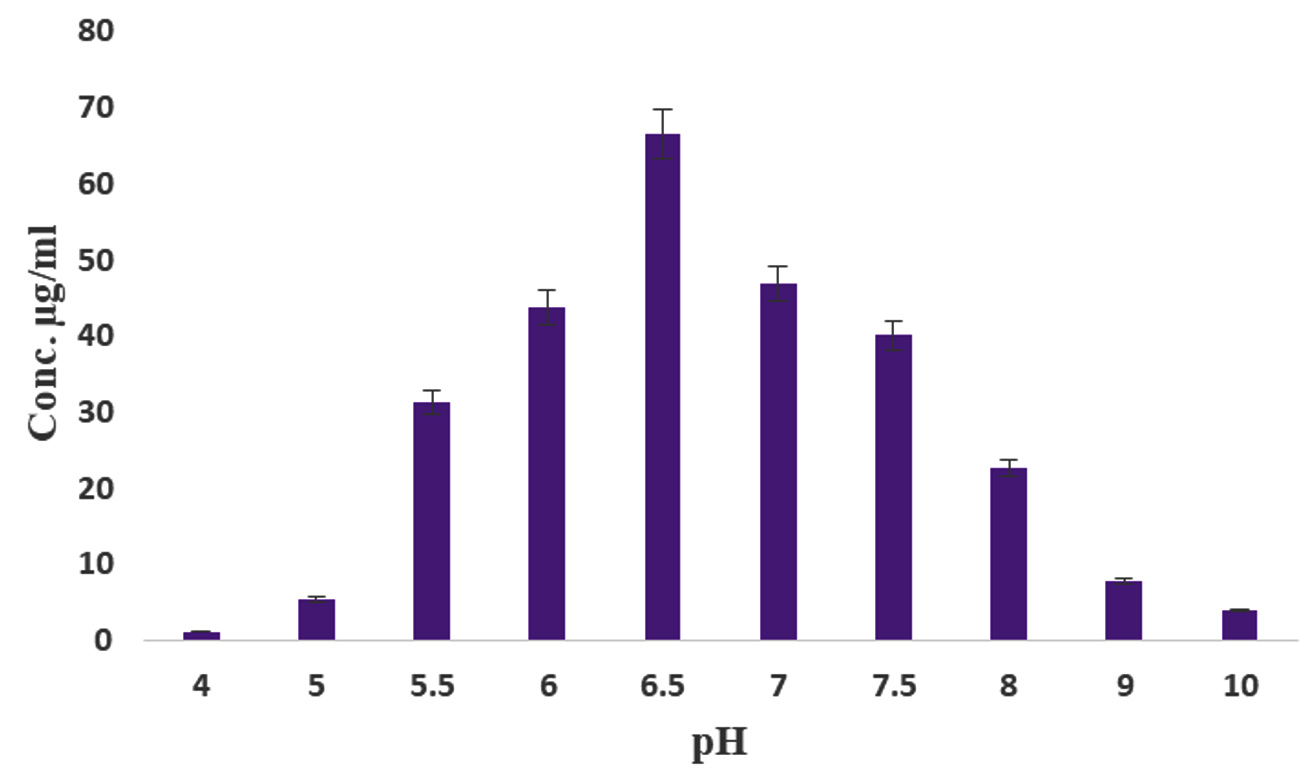 Figure 6. Production of IAA by Streptomyces sp. VSMKU1027 at different pH
Figure 6. Production of IAA by Streptomyces sp. VSMKU1027 at different pH
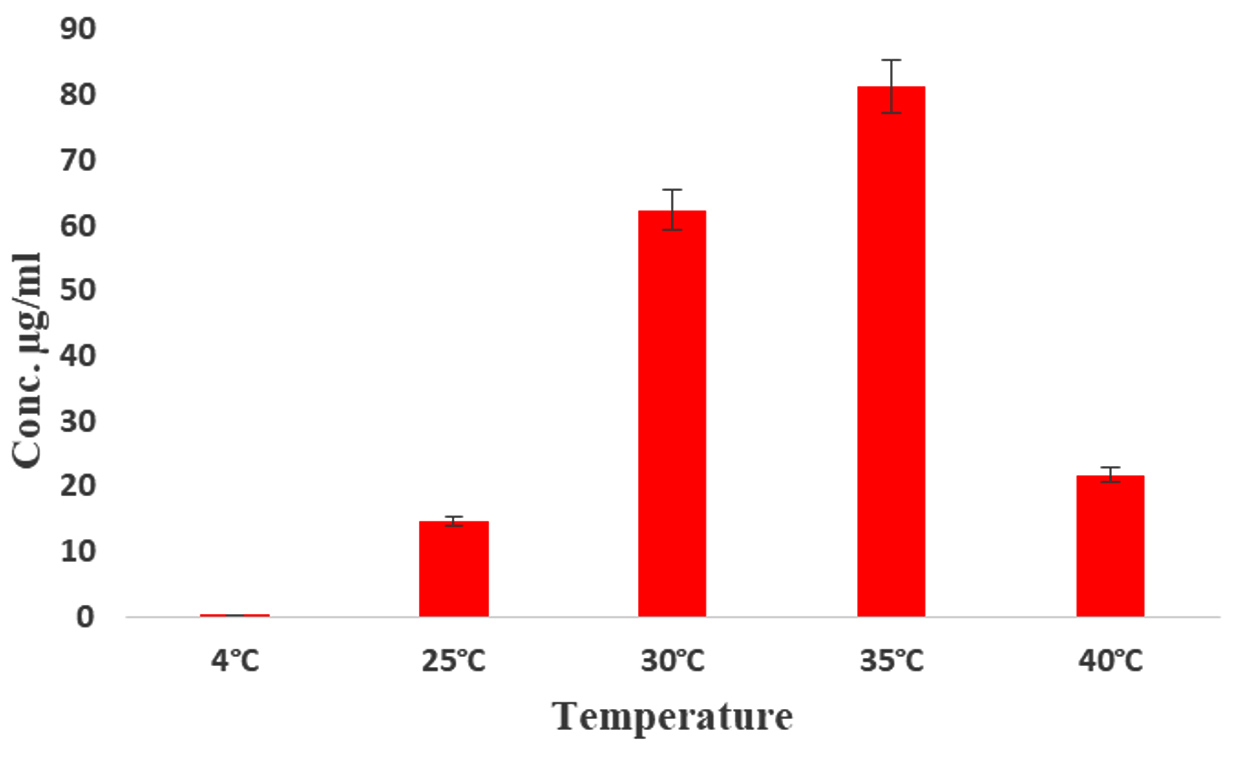 Figure 7. Production of IAA by Streptomyces sp. VSMKU1027 at different temperatures
Figure 7. Production of IAA by Streptomyces sp. VSMKU1027 at different temperatures
According to recent research, the native isolate Streptomyces sp. VSMKU1027 may be able to boost plant development and function as a biocontrol agent by using naturally occurring strains of bioactive actinomycetes. For the purpose of creating agricultural inoculants that protect plants from a variety of biotic and abiotic challenges, this makes it an invaluable bio resource.
ACKNOWLEDGMENTS
The authors would like to thank the Chairperson, School of Biological Sciences, Madurai Kamaraj University, for providing laboratory facilities. The authors would also like to thank MKU-RUSA for providing financial support.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
VVD and VS designed the study. VVD performed investigation. VVD performed results interpretations. KCMER, RSA, PS and VS validated the data. VS performed supervision. VVD wrote the manuscript. SG and VS reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This study was funded by MKU-RUSA with File No.011/RUSA/MKU/2020-2021.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Olanrewaju OS, Glick BR, Babalola OO. Mechanisms of action of plant growth promoting bacteria. World J Microbiol Biotechnol. 2017;33(11):1-6.
Crossref - Suarez-Moreno ZR, Vinchira-Villarraga DM, Vergara-Morales DI, et al. Plant-growth promotion and biocontrol properties of three Streptomyces spp. isolates to control bacterial rice pathogens. Front Microbiol. 2019;10:422554.
Crossref - Harikrishnan H, Shanmugaiah V, Balasubramanian N, Sharma MP, Kotchoni SO. Antagonistic potential of native strain Streptomyces aurantiogriseus VSMGT1014 against sheath blight of rice disease. World J Microbiol Biotechnol. 2014;30(12):3149-3161.
Crossref - Harikrishnan Ha, Shanmugaiah V, Balasubramanian N. Optimization for production of Indole acetic acid (IAA) by plant growth promoting Streptomyces sp VSMGT1014 isolated from rice rhizosphere. Int J Curr Microbiol Appl Sci. 2014;3(8):158-171.
- Kaur T, Sharma D, Kaur A, Manhas RK. Antagonistic and plant growth promoting activities of endophytic and soil actinomycetes. Arch Phytopathol Plant Prot. 2013;46(14):1756-1768.
Crossref - Gomathinayagam S, Persaud SA, Rekha M. Comparative study of biological agents, Trichoderma harzianum and Trichoderma viride for controlling brown spot disease in rice. Journal of Biopesticides. 2012;5:28-32.
- Harikrishnan H, Shanmugaiah V, Nithya K, et al. Enhanced production of phenazine like metabolite produced by Streptomyces aurantiogriseus VSMGT1014 against rice pathogen, Rhizoctonia solani. J Basic Microbiol. 2016;56(2):153-161.
Crossref - Nithya K, Vellasamy S, Natesan B, Sharma MP, Al Farraj DA, Elshikh MS. Characterization of antifungal metabolite phenazine from rice rhizosphere fluorescent Pseudomonads (FPs) and their effect on sheath blight of rice. Saudi J Biol Sci. 2020;27(12):3313-3326.
Crossref - Suresh P, Varathraju G, Shanmugaiah V, Almaary KS, Albadawi YV, Mubarak A. Partial purification and characterization of 2, 4-diacetylphloroglucinol producing Pseudomonas fluorescens VSMKU3054 against bacterial wilt disease of tomato. Saudi J Biol Sci. 2021;28(4):2155-2167.
Crossref - Suresh P, Rekha M, Gomathinayagam S, et al. Characterization and assessment of 2, 4-Diacetylphloroglucinol (DAPG)-producing Pseudomonas fluorescens VSMKU3054 for the Management of Tomato Bacterial Wilt Caused by Ralstonia solanacearum. Microorganisms. 2022;10(8):1508.
Crossref - Shanmugaiah V, Mathivanan N, Balasubramanian N, Manoharan PT. Optimization of cultural conditions for production of chitinase by Bacillus laterosporous MML2270 isolated from rice rhizosphere soil. Afr J Biotechnol. 2008;7(15):2562-2568.
- Varatharaju G, Nithya K, Suresh P, et al. Biocontrol properties and functional characterization of rice rhizobacterium Pseudomonas sp. VsMKU4036. J Pure Appl Microbiol. 2020;14(2):1545-56.
Crossref - Shanmugaiah V, Harikrishnan H, Al-Harbi NS, et al. Facile synthesis of silver nanoparticles using Streptomyces sp. VSMGT1014 and their antimicrobial efficiency. Dig. J Nanomater Biostruct. 2015;10(1):179-187.
- Sobanbabu G, Oviya R, Meena B, Vijayasamundeeswari A, Shanmugaiah V, Ramamoorthy V. Evaluation of phyllosphere bacterial biocontrol agents for the suppression of rice foliar diseases. J Phytopathol. 2024;172(2):e13300.
Crossref - Shanmugaiah V, Mathivanan N, Varghese B. Purification, crystal structure and antimicrobial activity of phenazine 1 carboxamide produced by a growth promoting biocontrol bacterium, Pseudomonas aeruginosa MML2212. J Appl Microbiol. 2010;108(2):703-711.
Crossref - Kalimuthu R, Suresh P, Varatharaju G, Balasubramanian N, Rajasekaran KM, Shanmugaiah V. Isolation and characterization of Indole acetic acid [IAA] producing tomato Rhizobacterium pseudomonas sp VSMKU4050 and its potential for plant growth promotion. Int J Curr Microbiol Appl Sci. 2019;8(06):443-455.
Crossref - Vessey JK. Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil. 2003;255(2):571-586.
Crossref - Kumar R. Role of microRNAs in biotic and abiotic stress responses in crop plants. Appl Biochem Biotechn. 2014;174(1):93-115.
Crossref - Marques APGC, Pires C, Moreira H, Rangel AOSS, Castro PML. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol Biochem. 2010;42(8):1229-1235.
Crossref - Jeon JS, Lee SS, Kim HY, Ahn TS, Song HG. Plant growth promotion in soil by some inoculated microorganisms. J Microbiol. 2003;41(4):271-276.
- Khan AG. Role of soil microbes in the rhizospheres of plants growing on trace metal contaminated soils in phytoremediation. J Trace Elem Med Biol. 2005;18(4):355-364.
Crossref - Lucy M, Reed E, Glick BR. Applications of free living plant growth-promoting rhizobacteria. Antonie van leeuwenhoek. 2004;86(1):1-25.
Crossref - Barriuso J, Ramos Solano B, Lucas JA, et al. Ecology, genetic diversity and screening strategies of plant growth promoting rhizobacteria (PGPR). Plant bacteria interactions: Strategies and techniques to promote plant growth. 2008;26:1-7.
Crossref - Majeed A, Abbasi MK, Hameed S, Imran A, Rahim N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front Microbiol. 2015;6:132438.
Crossref - Franco-Correa M, Quintana A, Duque C, Suarez C, Rodriguez MX, Barea JM. Evaluation of actinomycete strains for key traits related with plant growth promotion and mycorrhiza helping activities. Appl Soil Ecol. 2010;45(3):209-217.
Crossref - Alexander M. Introduction to soil microbiology. Soil Sci. 1978;125(5):331.
Crossref - Wang W, Li X, Wang J, Xiang S, Feng X, Yang K. An engineered strong promoter for streptomycetes. Appl Environ Microbiol. 2013;79(14):4484-4492.
Crossref - Minuto A, Spadaro D, Garibaldi A, Gullino ML. Control of soilborne pathogens of tomato using a commercial formulation of Streptomyces griseoviridis and solarization. Crop Protection. 2006;25(5):468-475.
Crossref - Seipke RF, Song L, Bicz J, et al. The plant pathogen Streptomyces scabies 87-22 has a functional pyochelin biosynthetic pathway that is regulated by TetR-and AfsR-family proteins. Microbiology. 2011;157(9):2681-2693.
Crossref - Keyeo F, Ai’shah ON, Amir HG. The effects of nitrogen fixation activity and phytohormone production of diazotroph in promoting growth of rice seedlings. Biotechnology. 2011;10(3):267-273.
Crossref - Khanna M, Solanki R, Lal R. Selective isolation of rare actinomycetes producing novel antimicrobial compounds. Int J Adv Biotechnol Res. 2011;2(3):357-375.
- Anwar S, Ali B, Sajid I. Screening of rhizospheric actinomycetes for various in-vitro and in-vivo plant growth promoting (PGP) traits and for agroactive compounds. Front Microbiol. 2016;7:203732.
Crossref - Harikrishnan H, Shanmugaiah V. Streptomyces sp. VSMGT1014-mediated antifungal activity against fungal plant pathogens. Prospects in bioscience: addressing the issues, Springer. 2013:335-341.
Crossref - Shanmugaiah V, Ramesh S, Jayaprakashvel M, Mathivanan N. Biocontrol and plant growth promoting potential of Pseudomonas sp. MML2212 from the rice rhizosphere. Mitteilungen aus der Biologischen Bundesanstalt fur Land- und Forstwirtschaft. 2006;408:320-324
- Suresh P, Vellasamy S, Almaary KS Dawoud PM, Elbadawi YB. Fluorescent pseudomonads (FPs) as a potential biocontrol and plant growth promoting agent associated with tomato rhizosphere. J King Saud Univer Sci. 2021;33(4):101423.
Crossref - Charulatha R, Harikrishnan H, Manoharan PT, Shanmugaiah V. Characterization of groundnut rhizosphere Pseudomonas sp. VSMKU 2013 for control of phytopathogens. Microbiological Research In Agroecosystem Management. Springer. 2013:121-127.
Crossref - Newitt JT, Prudence SMM, Hutchings MI, Worsley SF. Biocontrol of cereal crop diseases using streptomycetes. Pathogens. 2019;8(2):78.
Crossref - Suresh P, Shanmugaiah V, Rajagopal R, Muthusamy K, Ramamoorthy V. Pseudomonas fluorescens VSMKU3054 mediated induced systemic resistance in tomato against Ralstonia solanacearum. Physiol Mol Plant Pathol. 2022;119:101836.
Crossref - Bressan W, Figueiredo JEF. Efficacy and dose-response relationship in biocontrol of Fusarium disease in maize by Streptomyces spp. Eur J Plant Pathol. 2008;120(3):311-316.
Crossref - Dong D, Li M, Zhang T, et al. Antagonistic Activity of Streptomyces alfalfae 11F against Fusarium Wilt of Watermelon and Transcriptome Analysis Provides Insights into the Synthesis of Phenazine-1-Carboxamide. Plants. 2023;12(22):3796.
Crossref - Talwinder K, Riveka R, Manhas RK. Biocontrol and plant growth promoting potential of phylogenetically new Streptomyces sp. MR14 of rhizospheric origin. AMB Express. 2019;9(125):125.
Crossref - Marimuthu S, Karthic C, Mostafa AA, Al-Enazi NM, Abdel-Raouf N, Sholkamy EN. Antifungal activity of Streptomyces sp. SLR03 against tea fungal plant pathogen Pestalotiopsis theae. J King Saud Univer Sci. 2020;32(8):3258-3264.
Crossref - Le KD, Yu NH, Park AR, Park DJ, Kim CJ, Kim JC. Streptomyces sp. AN090126 as a biocontrol agent against bacterial and fungal plant diseases. Microorganisms. 2022;10(4):791.
Crossref - Pantoja-Guerra M, Burkett-Cadena M, Cadena J, Dunlap CA, Ramírez CA. Lysinibacillus spp.: An IAA-producing endospore forming-bacteria that promotes plant growth. Anto Van Leeuw. 2023 Jul;116(7):615-30.
- Emmert EAB, Handelsman J. Biocontrol of plant disease: a (Gram-) positive perspective. FEMS Microbiol Lett. 1999;171(1):1-9.
Crossref - Shobha G, Kumudini BS. Antagonistic effect of the newly isolated PGPR Bacillus spp. on Fusarium oxysporum. Int J Appl Sci Eng Res. 2012;1(3):463-474.
Crossref - Khan S, Srivastava S, Karnwal A, Malik T. Streptomyces as a promising biological control agents for plant pathogens. Front Microbiol. 2023;14:1285543.
Crossref - Al-Askar AA, Baka ZA, Rashad YM, et al. Evaluation of Streptomyces griseorubens E44G for the biocontrol of Fusarium oxysporum f. sp. lycopersici: ultrastructural and cytochemical investigations. Ann Microbiol. 2015;65:1815-1824.
Crossref - Tahvonen R, Avikainen H. The biological control of seed-borne Alternaria brassicicola of cruciferous plants with a powdery preparation of Streptomyces sp. Agric Food Sci. 1987;59(3):199-208.
Crossref - Mao J, Tang Q, Zhang Z, et al. Streptomyces radiopugnans sp. nov., a radiation-resistant actinomycete isolated from radiation-polluted soil in China. Int J Syst Evol Microbiol. 2007;57(11):2578-82.
Crossref - Sarwar A, Latif Z, Zhang S, et al. A potential biocontrol agent Streptomyces violaceusniger AC12AB for managing potato common scab. Front Microbiol. 2019;10:202.
Crossref - Chouyia FE, Romano I, Fechtali T, et al. P-solubilizing Streptomyces roseocinereus MS1B15 with multiple plant growth-promoting traits enhance barley development and regulate rhizosphere microbial population. Front Plant Sci. 2020;11:559833.
Crossref - Narayana K, Peddikotla P, Krishna PS, Venketeswarlu Y, Vijayalakshmi M. Indole-3-acetic acid production by Streptomyces albidoflavus. J Biol Res. 2009;11:49-55.
- Khamna S, Yokota A, Lumyong S. Actinomycetes isolated from medicinal plant rhizosphere soils: diversity and screening of antifungal compounds, indole-3-acetic acid and siderophore production. World J Microbiol Biotechnol. 2009;25(4):649-655.
Crossref - Myo EM, Ge B, Ma J, et al. Indole-3-acetic acid production by Streptomyces fradiae NKZ-259 and its formulation to enhance plant growth. BMC Microbiol. 2019;19(1):1-4.
Crossref - Kaur T, Manhas RK. Evaluation of ACC deaminase and indole acetic acid production by Streptomyces hydrogenans DH16 and its effect on plant growth promotion. Biocatal Agric Biotechnol. 2022;42:102321.
Crossref - Datta C, Basu PS. Indole acetic acid production by a Rhizobium species from root nodules of a leguminous shrub, Cajanus cajan. Microbiol Res. 2000;155(2):123-127.
Crossref - Goudjal Y, Toumatia O, Sabaou N, Barakate M, Mathieu F, Zitouni A. Endophytic actinomycetes from spontaneous plants of Algerian Sahara: indole-3-acetic acid production and tomato plants growth promoting activity. World J Microbiol Biotechnol. 2013;29(10):1821-1829.
Crossref - Aldesuquy HS, Mansour FA, Abo-Hamed SA. Effect of the culture filtrates of Streptomyces on growth and productivity of wheat plants. Folia Microbiologica. 1998;43(5):465-470.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.