


ISSN: 0973-7510
E-ISSN: 2581-690X
Plant Growth Promoting Rhizobacteria (PGPR) play an essential role in enhancing plant growth, health and yield. In this study, we describe the isolation of PGPR associated with the olive tree (Olea europaea L.) grown in three Moroccan regions of Zouala, Errachidia, and Essaouira. In these regions, we isolated 95 PGPRs from rhizosphere of Olive trees, 78% of them were characterized by their tolerance to a salinity of 4-11%. We also found that 39% of these PGPRs were phosphate solubilizing bacteria (PSB) with a solubilization ability greater than 100 µg/mL. In fact, Pantoea agglomerans (MRC_ZO_17) and Enterobacter ludwigii (MRC_ZO_97), showed the highest phosphate solubilization rates of ~450 µg/mL and ~196 µg/mL, respectively. In addition to their ability to solubilize phosphate, various isolates had the ability to produce Indole-3-acetic acid (IAA). For instance, E. ludwigii (MRC_ZO_97) had an IAA production of ~60.48 g/mL. In the region of Zouala, characterized by relatively higher salinity and lower rate of organic matter, Firmicutes isolates account for 87% of the isolated rhizobacteria. Interestingly, we found that the olive tree-associated PGPRs vary significantly between the three sampled regions. Several rhizobacteria isolated in this study are excellent candidates for formulation as bioinocula for plants in saline soils.
PGPR, Rhizosphere Soil, Diversity, Olive, Phosphate Solubilization, Abiotic Stress
The olive tree (Olea europaea L.) is one of the oldest cultivated crop species. It was domesticated around 2000 BC and since then, it has played a pivotal socioeconomic role, especially in the Mediterranean region, which accounts for 95% of olive production worldwide.1,2 It is the only member of the Oleaceae family that yields edible fruits, and it stands second oil crop after the palm.1,2 Olive oil presents an important source of nutrients and health benefits.3 The demand for olive oil continues to grow and thus the pressure on olive production has also increased.4,5 In Morocco, which is one of the most important olive producers among the Mediterranean countries, the olive crop occupies a surface of more than 560,000 ha and contributes about 5% of the Moroccan GDP.6
To face the growing demands for olive production in Morocco and other countries, two main strategies are followed: (a) expanding the olive-growing lands and (b) increasing the yield per area unit. Despite substantial efforts to expand olive production surface such as assisting farmers in adopting new irrigation technologies and modern farming practices to cultivate new lands the expansion of production areas remains unsatisfactory. This is primarily due to several limitations, including soil salinization, water scarcity, and the high cost of chemical fertilizers. Therefore, emphasizing the enhancement of yield per unit area emerges as a more practical and sustainable approach to concentrate on. This strategy necessitates a deeper comprehension of O. europaea’s biology and cannot be accomplished solely through the application of chemical fertilizers and pesticides. For instance, when essential elements, such as nitrogen (N), phosphate (P), and potassium (K) are supplied by chemical fertilizers, a huge amount of them can be lost due to leaching or immobilization.7,8 However, continuous application of chemical fertilizers and pesticides can lead to deterioration of the microbiology, texture, and fertility of the treated soils, negatively affects the plant performance. Besides the accumulation of numerous toxic elements in the phreatic napes.9,10 Nevertheless, chemical fertilizers could be substituted with organic amendments that have shown significant economic and environmental benefits.1 One very old and simple way to enhance soil potential is to mix different soils to remedy nutrients and as it was described by Theophrastus (372–287 BC) “to add heart” to the soil.11
Recently, research has put the spotlight on a different aspect that is very implicated directly and indirectly involved in the availability and uptake of nutrients and plant immunity, which is the plant microbiome, notably the rhizobacteria.12-14 The rhizosphere was first described by the agronomist Hiltner in 1904 as the region of the interface between the plant root system and its surrounding bulk soil. This interface is a very complex ecosystem that is characterized by a large number of diversified bacteria. Among these bacteria, Plant growth-promoting Rhizobacteria (PGPR) are of particular interest,15 because of their ability to facilitate the immobilization and absorption of Nitrogen (N), Phosphorus (P), and Potassium (K) without the need for mineral fertilizers,16 Interestingly, many species of PGPR are directly involved in the biosynthesis of the phytohormones Indole-3-acetic acid (IAA), cytokinins, and gibberellins that modulate plant growth and root architecture.17,18 Furthermore, PGPR can also secrete a variety of beneficial compounds, including cell wall degrading enzymes, antibiotics, and antifungals that directly influence plant-pathogen interactions and improve plant immunity.18-20 Moreover, once inoculated in a Rhizosphere, PGPRs can persist for a long time and in fact, the plant naturally encourages their establishment in a very symbiotic interaction. Therefore, PGPR can be a great source of biofertilizers, biostimulants, and biocontrol agents that should be incorporated into organic olive production.
This study focuses on the isolation, characterization, and molecular analysis of bacteria inhabiting the rhizosphere of cultivated olive trees across three distinct regions in Morocco. Emphasis is placed on assessing their potential to be categorized as Plant Growth-Promoting Rhizobacteria (PGPR) and, by extension, their suitability as bioinoculants for soil enhancement.
Study sites, sample collection and bacterial isolation
Soil samples were collected from the rhizosphere of olive trees grown at several sites within three southern regions of Morocco; Zouala region in the Ziz Valley (31° 47’N, 04° 14’ W); Errachidia region (31° 51’ N, 04° 16’ W) and Essaouira region (31° 17N, 9° 27’W) (Figure 1). Soil samples were placed in sterile boxes then transported to the laboratory four days later for further analysis. Each soil sample weighing 1g was resuspended in 9mL of sterile saline solution and utilized for bacterial isolation. The soil suspensions were gently stirred for 1 hour at room temperature, before being serially diluted with sterile saline solution. 100 µL of each dilution was spread on nutrient agar plates supplemented with 50 µg/mL of the antifungal Amphotericin B and incubated for 48±2 hours at 28°C. From each of the dilution plates exhibiting bacterial growth, colonies were chosen and streaked onto nutrient agar plates to establish pure cultures. For the purpose of acquiring ample biomass, subsequent to this step, each individual isolate was introduced into nutrient broth and allowed to incubate for 24+2 hours. Finally, the isolates were cryopreserved in a 40% (v/v) glycerol solution at -80°C.

Figure 1. Location of sampling sites. A: Errachidia, B: Zouala, C: Essaouira
Qualitative and quantitative estimation of phosphate solubilization
To evaluate the phosphate solubilization ability, qualitative tests were performed using Pikovskaya’s agar plates.21 The plates contain tricalcium phosphate as the sole phosphate source (P) and bromophenol blue to visualize phosphate solubilization. After 5 days of incubation at 28°C, colonies that showed a solubilization zone on the agar plates were considered to exhibit phosphate solubilization traits. Quantitative phosphate solubilization tests were performed in Pikovskaya’s liquid (PVK) medium.21 Selected isolates were cultivated in Erlenmeyer flasks containing 100 mL of liquid PVK medium supplemented with 0.1 g of phosphate rock (Phosphate mines, Khouribga, Morocco). Prior to its addition to the PVK medium, the phosphate rock was ground to a particle size of 0.02 mm. All cultures had an initial optical density (OD600= 0.05) in order to standardize the cell density of all isolates. The tested isolates were inoculated and grown in PVK medium for 48±2 hours at 28°C with continuous shaking at 180 rpm. At the end of each incubation, the pH of the growth medium was measured and the bacterial cultures were centrifuged at 13,000 x g for 10 minutes. The supernatants obtained were filter-sterilized using a 0.2 µm filter (Millipore). Soluble Phosphate concentrations in each supernatant were determined at 405 nm using the Vanadomolybdo-phosphoric Acid colorimetric method.22 Sterile PVK medium supplemented by phosphate rock incubated for 48h was used as a blank control. A KH2PO4 standard curve with known concentrations was used to calculate soluble phosphate concentrations.
Indole acetic acid (IAA) production
To determine the amounts of IAA produced by each isolate, a colorimetric technique was performed using Van Urk Salkowski reagent following Salkowski’s method.23 Bacterial cultures were prepared following the procedure described above and then cultivated in YEM liquid cultures containing 500 µg/mL of Tryptophan for 24h at 28°C in a shaking incubator at 180 rpm. Fully grown cultures were centrifuged at 10,000g for 10 min and the obtained supernatants were filtered using a 0.2 µm Millipore filter. From each supernatant 1 mL was mixed with 2 mL of Salkowski reagent (50 ml, 35% of Perchloric Acid, 1 ml 0.5 M FeCl3 solution) and kept in the dark for 30 min. The development of pink color is indicative of IAA production. OD values were recorded at 530 nm and the concentrations of IAA produced by each culture were determined against a standard curve built using known commercial IAA (Sigma Aldrich).
Ammonia (NH3) and Hydrogen cyanide (HCN) production
Tests of Ammonia and Hydrogen cyanide were carried out according to the methods described by Cappuccino and Sherman24 and Lorck,25 respectively.
Exopolysaccarides (EPS) production
For each isolate, a 5mm-diameter paper disc was inoculated with 2 microliters of liquid culture. These discs were then placed onto the surface of an agar medium (composed of 2% yeast extract, 1.5% K2HPO4, 0.02% MgSO4, 0.0015% MnSO4, 0.0015% FeSO4, 0.003% CaCl2, 0.0015% NaCl, 1.5% agar), supplemented with 10% saccharose and adjusted to a pH of 7.5.26 The production of EPS was determined by the size of the halo around the isolate-containing discs and their visible slime appearance. This EPS production was confirmed through the precipitation of a portion of the mucoid substance from the slime, which was mixed with 2 ml of absolute ethanol, resulting in a precipitate indicating the presence of EPS.27
Siderophores production
Production of siderophores by various isolates was quantified according to the Schwyn and Neilands method28 modified by Alexander and Zuberer.29 Cultures were cultivated in an iron-free medium composed of (in g.L-1): K2HPO4 ‘0.5’; MgSO4 7H2O ‘0.2’; (NH4)2SO4 ‘1.0’; Sodium glutamate ‘1.0’; NaCl ‘0.1’; Mannitol ’10.0’. After 7 days of incubation at 28°C in a rotary shaker, the cultures were centrifuged at 10,000g for 20 min and 1 mL of each supernatant was placed in a hemolysis tube pre-treated with HCl. Then they were mixed with Chrome Azurol S (CAS) reagent and incubated for 30 min in the dark. If present, siderophores remove iron from the dye complex and cause a decrease in the intensity of the blue color measured at 630 nm. The production of siderophores was measured in percentage units of siderophores calculated as ((Ar – As) × Ar – 1) × 100 ) where Ar is the absorbance of reference (media + CAS assay solution) and As is the absorbance of the sample.30
Potassium Solubilization
The qualitative test of potassium solubilization was carried out using Aleksandrov agar plate medium.31 The plates were then incubated at 28°C for 5 days. Colonies that showed a solubilization zone on the agar plates were considered to be positive.
Stress Tolerance Evaluation
The tolerance of tested isolates toward abiotic stress such as temperature and salinity were studied. Salt tolerance was tested in nutrient broth medium supplemented with (1%, 2%, 3% and up to 12% NaCl, w/v). Following incubation at 28°C for 24h, each isolate was streaked on an agar medium and growth was visually evaluated. The temperature tolerance of the isolates was investigated through streaking it onto nutrient agar plates, followed by incubation at specific temperatures: 28°C, 35°C, 40°C, 45°C, 55°C, and 60°C. The incubation duration was 24 hours.
DNA extraction, REP-PCR, and 16S rRNA
Total genomic DNA from each isolate was extracted and purified using the method described by Atashpaz et al.32 A NanoDrop 2000 was used to evaluate the purity and concentration of the isolated DNA.
The REP-PCR profiling was performed on 205 isolates using primers REP1R 5’-IIIICGICGICATCIGGC3’ and REP1I 5’- ICGICTTATCIGGCCTAC-3’. The thermocycling protocol comprises an initial denaturation at 95°C (3 min), 40 cycles of denaturation at 94°C (30 sec), hybridization at 40°C (45 sec) and extension at 72°C (1 min), followed by a final extension at 72°C (5 min). The amplicons were analyzed by electrophoresis in a 2% agarose gel at 70 volts for 90 min. After migration, the fingerprints were examined visually. From a group of isolates that had the same DNA fingerprint and physiological and biochemical characteristics, a single isolate was selected for 16S rRNA identification. A total of 95 isolates were retained and their 16S rRNA sequences were amplified using the forward (27F 5’-AGTTTGATCMTGGCTCAG-3’) and reverse (1492R 5’-GGTTACCTTGTTACG ACTT-3’) primers. The PCR program was as follows: initial denaturation at 95°C for 3 min, 30 cycles of 94°C for 3 min, 94°C for 30 s, 55°C for 45 s, 72°C for 1 min and 72°C for 8 min. All amplified gene fragments were sequenced in UATRS-CNRST, Rabat, Morocco. The sequences were corrected using Chromas software and were assembled using DNA Dragon: (Sequence Contig DNA assembly software”) and finally submitted to the GenBank Database (NCBI).

Figure 2. Cluster analysis of rhizobacteria isolated from Zouala region. The classification displayed here is based on their biochemical and physiological traits with the unweighted pair-group method using arithmetic averages (UPGMA), with identification, similarity (%) and accession number (GenBank database), designed using GraphPad Prism 8 software
This study presents the results of isolates from different regions: Zouala, Errachidia, and Essaouira. The outcomes for Zouala isolates, identified by the prefix MRC_ZO, are displayed in (Figure 2). Likewise, results pertaining to isolates from the Errachidia region, characterized by the prefix MRC_ES, are depicted in (Figure 3). Lastly, findings from the Essaouira region, with the prefix MRC_ES, are illustrated in (Figure 4).

Figure 3. Cluster analysis of rhizobacteria isolated from Errachidia region. Classification displayed here is based on their biochemical and physiological traits with the unweighted pair-group method using arithmetic averages (UPGMA), with identification, similarity (%) and accession number (GenBank database).
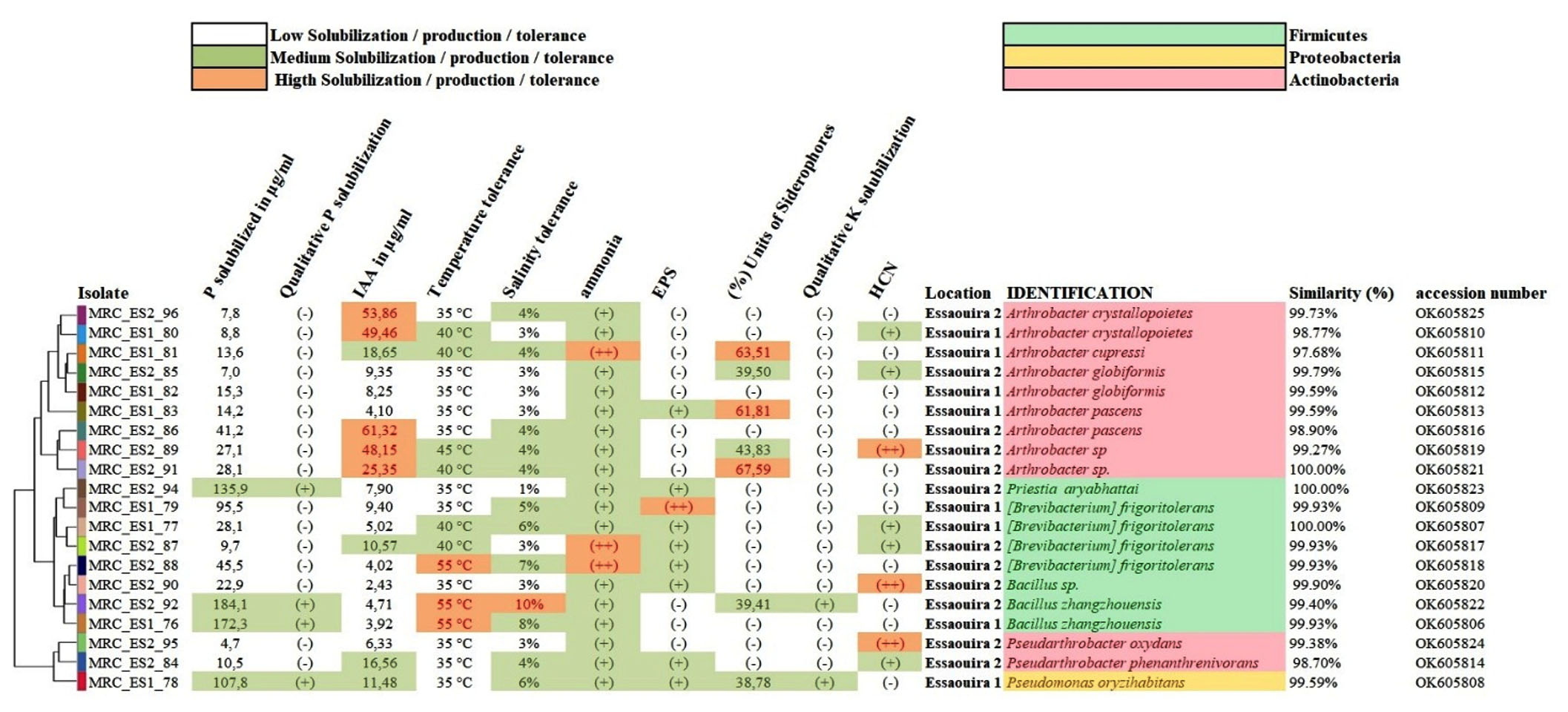 Figure 4. Cluster analysis of rhizobacteria isolated from Essaouira region. Classification displayed here is based on their biochemical and physiological traits with the unweighted pair-group method using arithmetic averages (UPGMA), with identification, similarity (%) and accession number (GenBank database)
Figure 4. Cluster analysis of rhizobacteria isolated from Essaouira region. Classification displayed here is based on their biochemical and physiological traits with the unweighted pair-group method using arithmetic averages (UPGMA), with identification, similarity (%) and accession number (GenBank database)

Figure 5. Qualitative activity of Olive rhizosphere-associated bacteria. A: The clear halo indicates the solubilization of phosphate on the PVK agar plate. B: Clear halo indicates solubilization of potassium on Aleksandrov agar plate. C: brown coloration of filter paper indicates HCN production. D: halo with slime appearance around the isolate indicates EPS production. E: confirmation of EPS production by the formation of a white precipitate in absolute ethanol. F: development of brown to yellow color indicates ammonium production
Qualitative and quantitative estimation of phosphate solubilization
During the qualitative test of P solubilization, all tested isolates were able to grow on PVK agar plates and only 30% of them were able to form a solubilization halo (Figure 5). The quantitative test showed that all isolates were able to solubilize the phosphate rock with solubilization values ranging from 4.7µg/mL to 447µg/mL and a pH ranging between 3,99 and 7,27. Interestingly about 39% and 2.1% of the tested isolates were able to solubilize more than 100µg/mL and 200 µg/mL, respectively. The lowest solubilization value was recorded in the isolate MRC_ES_95 with a final pH of 6,65, and the highest value was observed in the isolate MRC_ZO_17 with a pH of 3,99.
Indole acetic acid production
The IAA production test showed that all isolates were able to produce this phytohormone with a production range between 2,43 µg/mL and 61,32 µg/mL scored by the isolates MRC_ES_90 and MRC_ES_86, respectively. However, the number of isolates with a high to moderate production rate was low, as only 3.2% of the isolates were able to produce more than 50 mg/mL, 23.1% of the isolates produced between 10 µg/mL and 50 µg/mL and the rest (73.7%) produced less than 10 µg/mL of IAA.
Siderophores production
The production of siderophores is revealed by measuring the variation of color at 630 nm after the addition of the (CAS) reagent. Results were fluctuating between 0 and 79.8 percentage units. The vast majority of olive tree rhizobacteria (81% of isolates) did not produce siderophores. Conversely, 18.9% exhibited siderophore production above 25 percentage units, with 11.7% exceeding 40 percentage units and 7.4% surpassing 60 percentage units. Notably, the isolate MRC_ZO_19 showcased the most elevated outcome (79.8 percentage units), followed by isolate MRC_ER_50 (70.8 percentage units).
Ammonia and hydrogen cyanide production
All the tested isolates scored positive for ammonia production and 16.9 % of them were characterized with high levels of ammonia production, while 83.1% of them had low to moderate ammonia production. Regarding hydrogen cyanide production, 54 isolates scored positive with only 4 isolates having a good amount of HCN production.
Potassium solubilization and exopolysaccharides production
Among the 95 isolates tested for potassium solubilization, 12 isolates showed the formation of a solubilization halo and only 1 isolate (MRC_ZO_17) exhibited an important halo. Regarding exopolysaccharides production, 26 isolates were able to produce the exopolysaccharide, of which 17 showed moderate production and 9 showed good production.
Stress tolerance test
All isolates were tested for their tolerance to salinity with NaCl concentrations ranging from 1% to 12%. The results showed that 36.8% of the isolates grew at NaCl concentrations between 1% and 4%, 46.4% of the isolates grew at concentrations from 5% to 8%, and finally 16.8% of them were able to grow at NaCl concentrations between 8% and 11%. Interestingly, two isolates (MRC_ZO1_12 and MRC_ZO3_41) were able to tolerate 11% NaCl. With respect to the different incubation temperatures, 54.7% of the isolates were able to grow at temperatures above 45 °C and 44.2% of them grew at temperatures between 35 °C and 40°C.
Molecular analysis
A total of 205 bacterial isolates were isolated from the analyzed soil samples. Out of these, 95 isolates (46 from Zouala, 29 from Errachidia, and 20 from Essaouira) were selected based on their REP-PCR profiling results. After comparing the 16S rRNA sequences with BLAST homologs in GenBank, As displayed in Table 1, there are numerous genera, with the genus Bacillus predominating.
Table (1):
Taxonomic distribution of the 95 rhizobacteria isolated from the Zouala, Errachidia and Essaouira regions
| Order | Family | Genus | Number of bacterial isolates | |||
|---|---|---|---|---|---|---|
| Zouala | Errachidia | Essaouira | Total | |||
| Bacillales | Bacillaceae | Bacillus | 27 | 13 | 3 | 43 |
| Brevibacterium | 7 | 2 | 4 | 13 | ||
| Priestia | 4 | 1 | 1 | 6 | ||
| Cytobacillus | 2 | – | – | 2 | ||
| Pseudomonadales | Pseudomonadaceae | Pseudomonas | – | 10 | 1 | 11 |
| Moraxellaceae | Acinetobacter | 1 | – | – | 1 | |
| Enterobacteriaceae | Enterobacter | 1 | – | – | 1 | |
| Pantoea | 1 | – | – | 1 | ||
| Micrococcales | Micrococcaceae | Arthrobacter | 1 | – | 9 | 10 |
| pseudatrthrobacter | 2 | 2 | 2 | 6 | ||
| Sinomonas | – | 1 | – | 1 | ||
Plant growth-promoting Rhizobacteria has been subjected to a huge number of studies across multiple crops to study their potential use as a sustainable and eco-friendly alternative to chemical fertilizers. So far, multiple important PGPR isolates have been identified and belong to a variety of genera, including Acinetobacter, Agrobacterium, Arthrobacter, Azotobacter, Bacillus, Burkholderia, Enterobacter, Pantoea, Pseudomonas and Rhizobium.15,33 The diversity of the rhizosphere microbiome is very enormous and is influenced by both the surrounding plants and environmental factors, such as the type of soil. In the present study, samples were collected from two different types of climates, arid (Errachidia and Zouala) and semi-arid (Essaouira). These three targeted regions are known for their traditional olive tree agriculture in small-scale farming systems with low to almost no irrigation and chemical inputs. Regarding soil characteristics, the Zouala region has a high rate of salinity and a very low rate of nitrogen and organic matter while the Essaouira region has a low rate of phosphorus (Table 2). The objective of this study is to choose elite PGPR isolates for the development of a biofertilizer and bioprotective bio-inoculum, while also establishing a bio-bank of rhizosphere bacteria for future applications. Additionally, this research acts as a supplementary extension to the work conducted by Lahsini et al.34 wherein each region was represented only by five isolates.
Table (2):
Physicochemical characteristics of the soil of the olive trees of the three regions Zouala, Errachidia, and Essaouira
Clay |
Sand |
Silt |
Classification |
pH (H2O) |
pH (Clk) |
Water |
Salinity |
OM |
N total |
C/N |
P |
K |
Fe |
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
% |
% |
% |
% |
dS/m |
% |
% |
ppm |
ppm |
ppm |
|||||
Zouala |
25.20 |
34.54 |
40.26 |
Loam |
7.8 |
7.8 |
10.92 |
8.83 |
0.83 |
0.058 |
8.29 |
13 |
860 |
3 |
Errachidia |
20.60 |
32.07 |
47.33 |
Loam |
8.3 |
8.1 |
18.26 |
0.87 |
2.34 |
0.162 |
8.32 |
63 |
640 |
5 |
Essaouira |
58.1 |
7.22 |
34.68 |
Clay |
7.9 |
7.0 |
11.30 |
0.18 |
3.83 |
0.266 |
8.27 |
2 |
280 |
4 |
As expected, the rhizobacterial profiles of the three regions were different with a predominance of Bacillaceae in the Zouala region, a co-dominance of Bacillaceae and Pseudomonadaceae in the region of Errachidia and finally, a co-dominance of Bacillaceae and Micrococcaceae in the Essaouira region. The predominance of firmicutes in the Zouala region with 87% firmicutes could be explained by its high soil salinity. Several studies have reported the adaptation of firmicutes to high salt concentrations through the production of endospores under stress conditions.35 Priest36 reported that firmicutes are often dominant in soils with poor organic matter. And despite its high salinity and lack of organic matter, the Zouala rhizosphere conceals an important diversity compared to the two other regions.
Barriuso et al. reported that 95% of gram-positive bacteria in the rhizosphere belong to the genus Bacillus and 5% belong to the genus Arthrobacter and Frankia while the majority of gram-negative bacteria belonging to the genus Pseudomonas.37 Similarly, our results showed the presence of Bacillus in the three studied regions, no pseudomonas was isolated in the region of Zouala. The Essaouira region was characterized by the codominance of Bacillaceae and Micrococcaceae. The differences observed in the microbial profile of the three regions are due to several factors such as the nature and composition of the soil, climate, and even agricultural practices.
In the present study, a total of 13 Brevibacterium frigoritolerans were isolated and identified from the 3 regions showing an important intraspecific diversity with a temperature tolerance ranging from 35°C to 55°C, a salinity tolerance ranging from 3% to 9% and their P solubilization varied from 8.4µg/mL to 95.5 µg/mL. Brevibacterium frigoritolerans and Brevibacterium halotolerans, reclassified nowadays as Bacillus halotolerans,38 were first isolated from the Moroccan arid soils by Delaporte and Sasson,39 and they seem to be typical indigenous species of arid Moroccan soils. Several studies have shown the effectiveness of Brevibacterium frigoritolerans and Bacillus halotolerans as bioremediants.40-42 We were able to isolate from the Zouala region two Bacillus halotolerans isolates with salinity tolerance ranging from 9% to 10%, a temperature tolerance of 55°C and good production of exopolysaccharides EPS.
These EPS are an important group of high-molecular-weight complex polymers composed of sugar moieties and form the major component of bacterial biofilm.43 Our results showed that 27.3% of the tested isolates have the ability to produce EPS, particularly those of the genus Bacillus. The exopolysaccharides produced by PGPR bind to Na+ cations and are known to increase plant tolerance to salinity and drought.43 During the temperature tolerance test, most Bacillus isolates produced EPS under heat stress with a secretion peak of secretion scored at 45°C. The presence of a viscous liquid in the iron-free medium after 7-day of incubation following stress related to lack of iron was also noted. Under stress conditions, these bacterial secretions keep the surrounding microenvironment hydrated and full of nutrients and metabolites that could improve the physicochemical characteristics of the soil and thus enhance the tree’s performance.
IAA secreted by PGPR is reported to significantly improve root growth parameters.44 Among the rhizobacteria we isolated from the three Moroccan regions, 26.3% showed good IAA production with an amount greater than 10 µg/mL. In our study, the best three were Arthrobacter pascens (MRC_ES2_86), Enterobacter ludwigii (MRC_ZO2_97), and Arthrobacter crystallopoietes (MRC_ES2_96). Noting that among the 13 bacteria that produce a quantity greater than 20µL/mL, 8 of them belong to Arthrobacter genus, which is not surprising, Actinobacteria are the main known producers of IAA.45-47 Nevertheless, our results showed that those isolates exhibited low Phosphate solubilization. In contrast, Bacillus isolates scored high performance as PSB and their IAA production fluctuated between (2.43 and 11.40 µg/mL), which is considered to be relatively low. Furthermore, other studies have reported significant IAA production by the genera Pseudomonas and Enterobacter and their beneficial effect on plant growth.48-50
A number of PGPRs are known for their ability to produce siderophores, which have a strong affinity to Fe3+ and other heavy metals. This characteristic is very useful to biologically detoxify contaminated soils by a huge number of heavy metals such as (Ag+, Al3+, Cd2+, Co2+, Cr2+, Cu2+, Eu3+, Fe3+, Ga3+, Hg2+, Mn2+, Ni2+, Pb2+, Sn2+, Tb3+, Tl+, and Zn2+). Some of these metals are essential for plant pathogens, and thus their aggregation by siderophores is a limiting factor for their proliferation. Among the 95 tested isolates in this study, 18 of them showed the ability to produce siderophores and thus they could be used as biocontrol agents and as a tool to help alleviate stress related to heavy metals.51-53 Furthermore, 41 of the bacterial isolates are capable of producing Hydrocyanic acid (HCN). They are considered to play a key role in the biological control of soil pathogens,54 by inhibiting the cytochrome oxidase and other metalloenzymes leading of the disturbance to the pathogens’ energy and thus their death.55
Phosphorus is an essential mineral for plant growth and development, it is considered the second limiting nutrient in the soil and is regularly supplemented, usually as a chemical fertilizer.56 Some PGPRs have the ability to transform inorganic P that is normally not assimilable by plants into an easily assimilated form. During this study, all tested stains displayed the ability to solubilize P. The MRC_ZO_17 isolate, identified as Pantoea agglomerans, showed an excellent solubilization capacity and high tolerance to salinity up to (8% NaCl w/v) which is in concordance with other reports.57,58 Furthermore, almost all tested isolates belonging to the genus Pseudomonas exhibited a solubilization halo on PVK agar plates, which indicates their ability to produce organic acids. Gram-negative bacteria use organic acids most likely Gluconic acid to lower the pH as part of the phosphate solubilization.59 However, some bacteria use other mechanisms such as the secretion of some enzymes like phosphatases in order to dissolve the phosphate.60,61 Indeed, in our study, only 26.6% of isolates belonging to the genus Bacillus presented a halo of solubilization. Interestingly, the stains MRC_ER2_58 and MRC_ER2_62 showed good solubilization activity, 197.1 µg/mL and 190.8 µg/mL, respectively, without exhibiting any solubilization halo in PVK agar plates. During the phosphate solubilization assays, we used a Moroccan phosphate rock as a source of inorganic phosphate. Tchuisseu Tchakounte et al. reported that the Moroccan phosphate rock is the most recalcitrant source of phosphate compared to 4 other rocks originating from Algeria, Cameroon, Mali, and Mexico. Only a few isolates of the genera Bacillus and Arthrobacter were able to solubilize the Moroccan phosphate rock.62 This indicates that our rhizobacteria isolated from the rhizosphere of Moroccan olive trees display a very good efficiency in terms of solubilization of rock phosphate.
Potassium is also an essential element for plant growth. Approximately 90 to 98% of soil potassium is fixed in silicate minerals, making it very difficult or unavailable for plant uptake.8 Our results revealed that 12 isolates showed the formation of a solubilization halo on Aleksandrov agar plate, 8 isolates belonging to the genus Pseudomonas, 2 to the genus Bacillus, 1 to the genus Enterobacter, and 1 to the genus Pantoea, which is in agreement with previous studies.8,63,64
To sum up, the microbial profiles of olive tree rhizospheres vary across the three studied regions. However, all three regions contained a good number of isolates that were tolerant to high concentrations of salinity. the genus Arthrobacter was characterized by a good production of IAA and siderophores, as well as a low P solubilization compared to other genera. The genus Bacillus was characterized by good P solubilization, low IAA production, good tolerance to salinity/temperature, and produced EPS. The isolates MRC_ZO_17 and MRC_ZO_97 isolated herein and identified as Pantoea agglomerans and Enterobacter ludwigii, respectively, are stress tolerant and possess potential PGP traits, indicating that they have a promising potential use as an inoculum to improve the oil yield and olive quality in saline and/or arid soils.
ACKNOWLEDGMENTS
The authors thank the Hassan II Academy of Science and Technology of Morocco for the financial support (Rhizolive project).
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Bizos G, Papatheodorou EM, Chatzistathis T, Ntalli N, Aschonitis VG, Monokrousos N. The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants. 2020;9(6):743.
Crossref - Porfirio S, da Silva MDRG, Cabrita MJ, Azadi P, Peixe A. Reviewing current knowledge on olive (Olea europaea L.) adventitious root formation. Sci Hortic. 2016;198:207-226.
Crossref - Persuric ASI, Damijanic AT. Connections between Healthy Behaviour, Perception of Olive Oil Health Benefits, and Olive Oil Consumption Motives. Sustainability. 2021;13(14):7630.
Crossref - Baldoni L, Belaj A. Olive. In: Vollmann J, Rajcan I, eds. Oil Crops. Handbook of Plant Breeding. Springer; 2010:397-421.
Crossref - Mili S, Bouhaddane M. Forecasting Global Developments and Challenges in Olive Oil Supply and Demand: A Delphi Survey from Spain. Agriculture. 2021;11(3):191.
Crossref - El Mouhtadi I, Agouzzal M, Guy F. L’olivier au Maroc. OCL. 2014;21(2):D203.
Crossref - Alori ET, Glick BR, Babalola OO. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front Microbiol. 2017;8:971.
Crossref - Zhang C, Kong F. Isolation and identification of potassium-solubilizing bacteria from tobacco rhizospheric soil and their effect on tobacco plants. Appl Soil Ecol. 2014;82:18-25.
Crossref - Hegedus M, Toth-Bodrogi E, Nemeth S, Somlai J, Kovacs T. Radiological investigation of phosphate fertilizers: Leaching studies. J Environ Radioact. 2017;173:34-43.
Crossref - Kang J, Amoozegar A, Hesterberg D, Osmond DL. Phosphorus leaching in a sandy soil as affected by organic and inorganic fertilizer sources. Geoderma. 2011;161(3-4):194-201.
Crossref - Rashad M, Hafez M, Popov AI, Gaber H. Toward sustainable agriculture using extracts of natural materials for transferring organic wastes to environmental-friendly ameliorants in Egypt. Int J Environ Sci Technol. 2022;20:7417-7432.
Crossref - Adedeji AA, Haggblom MM, Babalola OO. Sustainable agriculture in Africa: Plant growth-promoting rhizobacteria (PGPR) to the rescue. Sci Afr. 2020;9:e00492.
Crossref - Berendsen RL, Pieterse CMJ, Bakker PAHM. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012;17(8):478-486.
Crossref - Gupta K, Dubey NK, Singh SP, Kheni JK, Gupta S, Varshney A. Plant Growth-Promoting Rhizobacteria (PGPR): Current and Future Prospects for Crop Improvement. In: Yadav AN, Singh J, Singh C, Yadav N, eds. Current Trends in Microbial Biotechnology for Sustainable Agriculture. Environ Microbial Biotechnol. 2021:203-226.
Crossref - Mekonnen H, Kibret M. The roles of plant growth promoting rhizobacteria in sustainable vegetable production in Ethiopia. Chem Biol Technol Agric. 2021;8(1):15.
Crossref - Mali SD, Attar YC. Formulation of cost-effective agro residues containing potassium solubilizing bacterial bio-inoculants using response surface methodology. Biocatal Agric Biotechnol. 2021;35:102113.
Crossref - Goswami D, Thakker JN, Dhandhukia PC. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Tejada Moral M, ed. Cogent Food Agric. 2016;2(1).
Crossref - Riaz U, Murtaza G, Anum W, Samreen T, Sarfraz M, Nazir MZ. Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides. In: Hakeem KR, Dar GH, Mehmood MA, Bhat RA, eds. Microbiota and Biofertilizers. 2021:181-196.
Crossref - Diabankana RGC, Afordoanyi DM, Safin RI, Nizamov RM, Karimova LZ, Validov SZ. Antifungal Properties, Abiotic Stress Resistance, and Biocontrol Ability of Bacillus mojavensis PS17. Curr Microbiol. 2021;78(8):3124-3132.
Crossref - Suresh P, Vellasamy S, Almaary KS, Dawoud TM, Elbadawi YB. Fluorescent pseudomonads (FPs) as a potential biocontrol and plant growth promoting agent associated with tomato rhizosphere. J King Saud Univ Sci. 2021;33(4):101423.
Crossref - Pikovskaya R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya. 1948;17:362-370.
- Tandon HLS, Cescas MP, Tyner EH. An Acid-Free Vanadate-Molybdate Reagent for the Determination of Total Phosphorus in Soils. Soil Sci Soc Am J. 1968;32(1):48-51.
Crossref - Gordon SA, Weber RP. Colorimetric Estimation of indoleacetic Acid. Plant Physiol. 1951;26(1):192-195.
Crossref - Cappuccino JC, Sherman N. Ammonia production. Microbiol Lab Man 3rd Ed Benjamin Cummings Pub Co Redw City CA USA. 1992:125-179.
- Lorck H. Production of hydrocyanic acid by bacteria. Physiol Plant. 1948;1(2):142-146.
Crossref - Kavamura VN, Santos SN, da Silva JL, et al. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol Res. 2013;168(4):183-191.
Crossref - Paulo EM, Vasconcelos MP, Oliveira IS, et al. An alternative method for screening lactic acid bacteria for the production of exopolysaccharides with rapid confirmation. Food Sci Technol. 2012;32(4):710-714.
Crossref - Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987;160(1):47-56.
Crossref - Alexander DB, Zuberer DA. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol Fertil Soils. 1991;12(1):39-45.
Crossref - Payne SM. [25] Detection, isolation, and characterization of siderophores. Methods in Enzymology. 1994;235:329-344.
Crossref - Rajawat MVS, Singh S, Tyagi SP, Saxena AK. A Modified Plate Assay for Rapid Screening of Potassium-Solubilizing Bacteria. Pedosphere. 2016;26(5):768-773.
Crossref - Atashpaz S, Khani S, Barzegari A, et al. A robust universal method for extraction of genomic DNA from bacterial species. Microbiology. 2010;79(4):538-542.
Crossref - Kumawat KC, Nagpal S, Sharma P. Potential of plant growth-promoting rhizobacteria-plant interactions in mitigating salt stress for sustainable agriculture: A review. Pedosphere. 2022;32(2):223-245.
Crossref - Lahsini AI, Sallami A, Ait-Ouakrim EH, et al. Isolation and molecular identification of an indigenous abiotic stress-tolerant plant growth-promoting rhizobacteria from the rhizosphere of the olive tree in southern Morocco. Rhizosphere. 2022;23:100554.
Crossref - Boroujeni YK, Boroujeni VN, Rastegari AA, Yadav N, Yadav AN. Soil Microbes with Multifarious Plant Growth Promoting Attributes for Enhanced Production of Food Crops. In: Yadav AN, ed. Soil Microbiomes for Sustainable Agriculture. Sustainable Development and Biodiversity. 2021;27:55-83.
Crossref - Priest FG. Isolation and Identification of Aerobic Endospore-Forming Bacteria. In: Harwood CR, ed. Bacillus. Springer. 1989:27-56.
Crossref - Barriuso J, Ramos Solano B, Lucas JA, Lobo AP, Garca-Villaraco A, Gutirrez Maero FJ. Ecology, Genetic Diversity and Screening Strategies of Plant Growth Promoting Rhizobacteria (PGPR). In: Ahmad I, Pichtel J, Hayat S, eds. Plant-Bacteria Interactions. Wiley-VCH Verlag GmbH & Co. KGaA; 2008:1-17.
Crossref - Ben-Gad D, Gerchman Y. Reclassification of Brevibacterium halotolerans DSM8802 as Bacillus halotolerans comb. nov. Based on Microbial and Biochemical Characterization and Multiple Gene Sequence. Curr Microbiol. 2017;74(1):1-5.
Crossref - Delaporte B, Sasson A. Study of bacteria from arid soils of Morocco: Brevibacterium haloterans n. sp. and Brevibacterium frigoritolerans n. sp. Comptes Rendus Hebd Seances Acad Sci Ser Sci Nat. 1967;264(18):2257-2260.
- Jariyal M, Gupta VK, Mandal K, Jindal V. Brevibacterium frigoritolerans as a Novel Organism for the Bioremediation of Phorate. Bull Environ Contam Toxicol. 2015;95(5):680-686.
Crossref - Jariyal M, Jindal V, Mandal K, Gupta VK, Singh B. Bioremediation of organophosphorus pesticide phorate in soil by microbial consortia. Ecotoxicol Environ Saf. 2018;159:310-316.
Crossref - Deng Z, Jiang Y, Chen K, Gao F, Liu X. Petroleum Depletion Property and Microbial Community Shift After Bioremediation Using Bacillus halotolerans T-04 and Bacillus cereus 1-1. Front Microbiol. 2020;11:353.
Crossref - Deka P, Goswami G, Das P, et al. Bacterial exopolysaccharide promotes acid tolerance in Bacillus amyloliquefaciens and improves soil aggregation. Mol Biol Rep. 2019;46(1):1079-1091.
Crossref - Vessey JK. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 2003;255(2):571-586.
Crossref - Li M, Li T, Zhou M, et al. Caenorhabditis elegans Extracts Stimulate IAA Biosynthesis in Arthrobacter pascens ZZ21 via the Indole-3-pyruvic Acid Pathway. Microorganisms. 2021;9(5):970.
Crossref - Manulis S, Shafrir H, EpteinA E, Barash I. Bissynthesis of indole-3-acetic acid via the indole-3-acetarnide pathway in Streptomyces SPP. Microbiology. 1994:140(5):634-642.
Crossref - Nafis A, Raklami A, Bechtaoui N, et al. Actinobacteria from Extreme Niches in Morocco and Their Plant Growth-Promoting Potentials. Diversity. 2019;11(8):139.
Crossref - Suzuki S, He Y, Oyaizu H. Indole-3-Acetic Acid Production in Pseudomonas fluorescens HP72 and Its Association with Suppression of Creeping Bentgrass Brown Patch. Curr Microbiol. 2003;47(2):138-143.
Crossref - Panigrahi S, Mohanty S, Rath CC. Characterization of endophytic bacteria Enterobacter cloacae MG00145 isolated from Ocimum sanctum with Indole Acetic Acid (IAA) production and plant growth promoting capabilities against selected crops. South Afr J Bot. 2020;134:17-26.
Crossref - Zhang BX, Li PS, Wang YY, et al. Characterization and synthesis of indole-3-acetic acid in plant growth promoting Enterobacter sp. RSC Adv. 2021;11(50):31601-31607.
Crossref - Braud A, Hannauer M, Mislin GLA, Schalk IJ. The Pseudomonas aeruginosa Pyochelin-Iron Uptake Pathway and Its Metal Specificity. J Bacteriol. 2009;191(11):3517-3525.
Crossref - Ashraf S, Ali Q, Zahir ZA, Ashraf S, Asghar HN. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotoxicol Environ Saf. 2019;174:714-727.
Crossref - Wang YJ, Huang W, Li YQ, Yu FB, Penttinen P. Isolation, Characterization and Evaluation of a High- Siderophore-Yielding Bacterium from Heavy Metal Contaminated Soil. Environmental Sci Poll Res. 2021:30.
Crossref - Voisard C, Keel C, Haas D, Defago G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. EMBO J. 1989;8(2):351-358.
Crossref - Blumer C, Haas D. Mechanism, regulation, and ecological role of bacterial cyanide biosynthesis. Arch Microbiol. 2000;173(3):170-177.
Crossref - Azziz G, Bajsa N, Haghjou T, et al. Abundance, diversity and prospecting of culturable phosphate solubilizing bacteria on soils under crop-pasture rotations in a no-tillage regime in Uruguay. Appl Soil Ecol. 2012;61:320-326.
Crossref - Son HJ, Park GT, Cha MS, Heo MS. Solubilization of insoluble inorganic phosphates by a novel salt- and pH-tolerant Pantoea agglomerans R-42 isolated from soybean rhizosphere. Bioresour Technol. 2006;97(2):204-210.
Crossref - Saadouli I, Mosbah A, Ferjani R, et al. The Impact of the Inoculation of Phosphate-Solubilizing Bacteria Pantoea agglomerans on Phosphorus Availability and Bacterial Community Dynamics of a Semi-Arid Soil. Microorganisms. 2021;9(8):1661.
Crossref - Oteino N, Lally RD, Kiwanuka S, et al. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front Microbiol. 2015;6.
Crossref - Suleman M, Yasmin S, Rasul M, Yahya M, Atta BM, Mirza MS. Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLOS ONE. 2018;13(9):e0204408.
Crossref - Mazumdar D, Saha SP, Ghosh S. Isolation, screening and application of a potent PGPR for enhancing growth of Chickpea as affected by nitrogen level. Int J Veg Sci. 2020;26(4):333-350.
Crossref - Tchuisseu Tchakounte GV, Berger B, Patz S, Fankem H, Ruppel S. Community structure and plant growth-promoting potential of cultivable bacteria isolated from Cameroon soil. Microbiol Res. 2018;214:47-59.
Crossref - Sarikhani MR, Oustan S, Ebrahimi M, Aliasgharzad N. Isolation and identification of potassium releasing bacteria in soil and assessment of their ability to release potassium for plants. Eur J Soil Sci. 2018;69(6):1078-1086.
Crossref - Khan YM, Pirdashti H, Rahimian H, Nematzadeh G, Ghajar Sepanlou M. Potassium solubilising bacteria (KSB) isolated from rice paddy soil: from isolation, identification to K use efficiency. Symbiosis. 2018;76(1):13-23.
Crossref
© The Author(s) 2023. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.