


Sugarcane (Saccharum officinarum L.) is a highly valuable agricultural crop, cultivated globally in tropical and subtropical regions, primarily for its sugar content. Out of the 110 countries that grow sugarcane, India and Brazil together account for half of the world’s total production. Being an annual crop, it is prone to many diseases. The major diseases that can affect the sugarcane are red rot, wilt, sett rot, grassy shoot and pokkah boeng. Among the diseases Colletotrichum falcatum, Fusarium sacchari, Ceratocystis paradoxa, Candidatus Phytoplasma and Fusarium fujikuroi pathogen leads to more yield loss. Traditional disease management strategies, such as chemical treatments, conventional methods and biological control, offer limited protection throughout the crop cycle and raise concerns regarding environmental impact and sustainability. In recent years, plant growth-promoting microorganisms (PGPMs) have emerged as a promising alternative, offering environmentally friendly solutions to enhance plant health and manage diseases. RNAi has been explored to manage various diseases caused by viruses, fungi, and bacteria. By silencing key pathogenicity genes in the causal organisms, sugarcane mosaic virus (SCMV), smut, and leaf scald. The integration of PGPMs with RNAi emerging biotechnological tools and transcriptomics presents a sustainable approach to disease control, potentially reducing the dependency on chemical pesticides and promoting eco-friendly agricultural practices.
Biocontrol, Plant Growth Promoting Microorganisms, RNAi, Sugarcane, Transcriptomics
Sugarcane is a commercial crop in the context of sugar production, playing a significant role in most global sugar production. Sugarcane is classified within the Poaceae and encompasses six species of perennial grass under the genus Saccharum. This classification highlights its botanical characteristics and its relationship to other grasses.1 The major diseases that leads to sugarcane yield loss are wilt (Fusarium sacchari), red rot (Colletotrichum falcatum), Pokkah Boeng (Fusarium verticillioides and Gibberella fujikuroi), sugarcane Grassy Shoot (Candidatus Phytoplasma), and pineapple sett rot (Ceratocystis paradoxa).2-7 These diseases are controlled by using chemical fertilizers but using excessive amount of agrochemicals, including pesticides and fertilizers, can lead to environmental problems.8 So, undertaking biological practices can be very helpful in developing more sustainable agricultural systems. Numerous bacteria have been utilized with efficiency to successfully control phytopathogenic fungi.9 The best method of preventing diseases rather than using chemicals is to plant healthy seedlings, use resistant varieties, and follow comprehensive disease management practices.10
This scholarly review holds significant importance as it furnishes an extensive comprehension of the manner in which PGPMs and novel biotechnological methodologies may be utilized to address the urgent challenge posed by sugarcane diseases. It outlines critical viewpoints on sustainable, economically practical, and groundbreaking tactics that are capable of redefining sugarcane agriculture, consequently strengthening its resilience, yield, and environmental preservation. And to understand the biological management practices and their benefits over chemical pesticides and the new RNA interference technology and transcriptomics to manage pathogen incidence.
Major diseases of sugarcane
Red rot
Collectotrichum falcatum, which causes red rot, is a major pathogen in sugarcane cultivation. This fungus is hemibiotrophic, which means that during its infection cycle, it alternates between the biotrophic (living within the host’s cells without causing immediate cell death) and necrotrophic (causing cell death and eventually feeding on the dead tissue) phases.11 It is first reported in India in 1895-1901 and also in tropical and subtropical regions.5 In the USA, Taiwan, Bangladesh, and Australia red rot is considered as a major setback.12 The disease incidence varies from 2-64%.5 The pathogen’s primary infection or mode of entry inside the plant is by the infected setts, and also by vectors it can easily spread from one field to the other field.13 In the 1980s, the popular sugarcane variety CoC671 in the various states of India became vulnerable to C. falcatum.14 The next important varieties-Co 997, Co 1148, Co 1158, Co 6304, Co 7805, CoC 671, CoC 92061, CoJ 64, CoS 8436, and CoSe 95422 were also severely affected.15
The symptoms of C. falcatum presents red lesions that are prominent on the central vein. As the disease progresses, these lesions develop a discolored center. Notably, in older leaves, the lesions can span the entire length of the vein.16 The symptoms and pathogen of red rot are given in Figure 1a-1b.
Pokkah Boeng
Pokkah Boeng is a term originating from the Javanese; in the year 1896, researchers Walker and Went worked on F. verticillioides and G. fujikuroi, which were identified as pathogenic organisms causing Pokkah Boeng disease in sugarcane.2 F. fujikuroi species complex (FFSC) is the airborne fungal disease known as Pokkah Boeng disease (PBD), which causes severe production losses in various kinds of sugarcane all over the world. F. sacchari, a species of FFSC, is linked to sugarcane withering and PBD.14 This disease causes economic yield loss either directly or through diverse fungal mycotoxins, which constitute a significant threat to the human health.17 This is an emerging disease that has been severely reducing crop yields in numerous developing and developed countries.18 The different researchers have identified the main species linked to it in different locations as F. sacchari, F. fujikuroi, F. proliferatum, F. verticillioides, and F. andiyazi.19 The occurrence of the disease decreased sugar production by 40.8-64.5% in the affected sugarcane plants.20 Furthermore, the disease incidence in Uttar Pradesh and Maharashtra differs based on the variety of sugarcane, ranging from 5% to 90%.21
In India F. sacchari and F. proliferatum were associated with the disease.22 The three phases of symptoms were visible in the various varieties.23 It can also spread through the seeds which were infected with fungus.
There are four stages of symptoms observed: chlorosis 1 and 2, the top decaying phase, and the knife cut phase. The predominant indicators of Pokkah Boeng disease involve the emergence of chlorotic patches near the lower portion of young leaves. Under conditions of high severity, the illness displays as a deformation of the stem, distinguished by both external and internal incisions that bear resemblance to cuts, as well as decay at the apex of the stem. Although the disease can manifest in many different ways from the typical symptoms in the field, the end effect is typically a deformed or broken stalk and top.24 The Pokkah Boeng symptoms are illustrated in Figure 1c-1d.
Wilt
Wilt has been associated with a significant increase in disease incidence in countries such as India and other nations in South Asia. Although wilt can be accompanied by red rot, it is noteworthy that wilt on its own possesses the capacity to inflict substantial harm in different parts of India. For instance, severe wilt was reported in 60% of disease cases in the Co 7717 variety, while 5-10% of cases were observed in CoJ 64, CoJ 79, and CoS 767 in Uttar Pradesh. Furthermore, major varieties in Bihar displayed severe wilt in conjunction with red rot. The anticipated losses of 3-6 tonnes per hectare translate to a loss of millions of tonnes and result in significant economic loss.25 F. sacchari its appearance in culture is similar to cotton, ranging from pink to purple.26 The primary source of infection is infected setts and crops left for ratooning. The pathogen can survive many years in the soil. The symptoms included wilting from the top and the whole plant dying and it was illustrated in Figure 1e-1f.

Figure 1. Symptoms and colony morphology of red rot, Pokkah Boeng and Wilt
(a) C. falcatum. (b) Collar rot symptoms tissues become red (red rot) (c) F. fujikuroi (d) Pokkah Boeng symptoms- Knife cut phase. (e) F. sacchari. (f) Wilt affected plants
Sett rot
The causal organism is Ceratocystis paradoxa. When the plant is infected by sett rot pathogen it leads to germination failure and it negatively affects the sugarcane crop growth7. The sett rot can also lead to total germination failure. The mode of infection is setts and soil-borne.27 The initial manifestations and indications of the disease encompass a water-saturated aspect observed at the extremities of the stems, leading to a subsequent reddening of the tissue. As the spread of infection progresses, the tissue transforms to a darker hue, revealing a cluster of dark spores within the stem. The process of colonization is characterized by the fermentation of host tissues, giving rise to a scent reminiscent of pineapple essence. This phenomenon is attributed to the release of ethyl acetate, a toxin that hinders the development of buds and roots.28
Grassy shoot of sugarcane
Candidatus Phytoplasma is an obligatory bacterial plant pathogen that lacks a cell wall, is not yet cultured, is spread by insects, and is linked to several plant disease signs.29 The symptoms of the SGS sickness include dwarfism and symptoms resembling a witch’s broom caused by an excessive number of tillers.30 This abundant growth produces number of tillers that are yellow and delicate to the touch, thin, narrow, and reduced in size.31 For a considerable amount of time, the challenges associated with cultivating phytoplasmas have hindered their detection and species identification, limiting their research to ecological studies, electron microscopy, and serological approaches.32
Control of Sugarcane Diseases
Physical control
The best method of preventing diseases is to plant healthy seeds, use resistant varieties, and follow comprehensive disease management practices.10 By using resistant cultivars, hazardous chemicals that could upset the natural balance and make some pests a problem are avoided.33 Integrated disease management combines numerous methods for effectively controlling diseases. This can involve biological control (using beneficial organisms), chemical control (using fungicides, if necessary), cultural techniques (crop rotation, optimum spacing), and other approaches.10
The method of immersing contaminated setts in hot water has been used to disinfect setts; the results demonstrated the efficacy of disinfection comparable to that of traditional chemical treatment which reduces the infection of F. fujikuroi.34 Planting of disease-free seedlings can reduce disease incidence. Frequently removing or “roguing out” unhealthy clumps involves eliminating plants showing disease symptoms. This is an essential step in stopping the SGS from spreading throughout the crop. It is important to note that insect vectors are recognized to be how phytoplasma infections are spread, emphasizing the importance of controlling these vectors.35 The pathogenic attack of F. fujikuroi may be effectively mitigated through the strategic application of a synergistic blend of diverse nutrients, including a formulation comprising NPK, ZnSO4, and FeSO4.36 Application of vermicompost can also decrease the attack of G. fujikuroi.37 The implementation of field sanitation methodologies, including the elimination and entombment of agricultural remnants, desiccated foliage, and stubble, among other practices before sowing, is imperative for the effective management of red rot.38 Sugarcane cultivation areas ought to be meticulously leveled, and adherence to hygienic agricultural practices is imperative. Systematic field evaluations and the removal of infected plants may significantly reduce the prevalence of red rot disease.39 Furthermore, given that the ailment is linked to an imbalance in soil nutrients, the management of fertilizers assumes critical importance.40
Chemical control
Even though chemical treatments cause environmental pollution it can rapidly increase the crop growth and decrease the disease incidence in the sugarcane. Within 15 days of Fusarium infection, two to three sprays are used to prevent the spread of the infection and minimize the cane’s yield and consistency loss.21 Application of the Propiconazole was practiced as a control management for pineapple sett rot.41 Debris infection can be prevented by soaking sugarcane setts in carbendazim and thiophanate methyl for 24 hours before planting.42 Fungicidal seed treatment with benomyl, thiram, benomyl + thiram, carbendazim + thiram, carboxin + thiram, fludioxonil, mancozeb, iprodione + triticonazole, prochloraz, thiophanate – methyl and ipconazole can reduce the Fusarium infestation.43 The fungicides benomyl, carbendazim, thiophanate-methyl, thiabendazole, and fuberidazole are examples of benzimidazole fungicides. Fungicides in a 0.3% suspension, including 50% carbendazim or 80% thiophanate-methyl, were applied to rice seeds to mitigate the Bakanae infections caused by the F. fujikuroi species complex (FFSC).44 The utilization of fungicides has been demonstrated to be essential for the proficient management of red rot in standing sugarcane within the Indian agricultural framework. Research grounded in observation has confirmed that numerous fungicides, including benomyl, tebuconazole, a mixture with 64% w/w mancozeb and 4% w/w metalaxyl-M, carbendazim, propiconazole, hexaconazole, carboxin, thiram, and azoxystrobin, effectively restricted the mycelial expansion of C. falcatum and showed efficacy against the red rot affliction.45
Biological control
Biological management can be very helpful in developing more sustainable agricultural systems. Numerous bacteria have been utilized with efficiency to successfully control phytopathogenic fungi.46 Both Bacillus species and Pseudomonas species have antagonistic qualities that can successfully restrict the growth of pathogens.47 Different Trichoderma spp. can decrease the Fusarium infection in numerous crops48 and additionally, it has several mechanisms that could reduce the spread of the disease.49 The various bacterial and fungal species controlling sugarcane diseases are illustrated in Table.
Table:
Bacteria species, crop, disease, and mode of action of plant growth promoting microorganism in sugarcane
Antagonistic Microrganism |
Disease |
Mode of action |
Ref. |
|---|---|---|---|
B. subtilis |
Pokkah Boeng, wilt |
Produce chitinase and glucanase |
47 |
Burkholderia |
Pokkah Boeng, wilt |
NR |
46 |
P. fluorescens |
Pokkah Boeng, red rot, wilt |
Antifungal activity and induced systemic resistance (ISR) |
57 |
Acinetobacter PK9 |
Red rot |
Antifungal activity |
62 |
K. pneumonia VRE36 |
Red rot |
Antifungal activity |
62 |
Bacillus sp. NH-217, B. subtilis NH-100, Bacillus sp. RSC 29 |
Red rot |
IAA production and phosphate solubilization |
60 |
O. intermedium TRD 14 |
Red rot |
NR |
62 |
Escherichia sp. VRE34 |
Red rot |
IAA production, Biological Nitrogen fixation and antagonistic properties |
62 |
P. putida strain NH-50 |
Red rot |
Pyoluteorin antibiotic |
70 |
B. subtilis TNAU |
Sett rot |
NR |
71 |
T. viride and T. harzianum |
Pokkah Boeng, Wilt |
Competition for food and space |
64 |
T. harzianum isolates Th1 and Th2 |
Wilt |
Hyperparasitism |
27 |
C. globosum |
Red rot |
NR |
72 |
T. asperullum, T. atroviride, T. harzianum and T. auroviride |
Red rot |
NR |
69 |
P. alvei |
Red rot |
NR |
69 |
T. asperellum TNAU |
Sett rot |
NR |
71 |
C. globosum Cg6 |
Sett rot |
NR |
71 |
Mechanism of action of biocontrol agents
Plant diseases have been successfully managed with the use of microbial inoculants.50 Plant growth and development are regulated by phytohormones, which also enable plants to withstand a variety of stresses.51 Certain rhizobacteria can generate phytohormones, including gibberellins, cytokinins, and abscisic acid, which can be helpful in plant growth processes, and increase N availability.52 However, the rhizobacteria’s synthesis of ABA is thought to be an indirect means of encouraging plant development.53 Indirect strategies of growth promotion include antibiosis, competition, and the development of lytic enzymes that can degrade fungal outer walls.54 The mechanism of action of biocontrol agents is illustrated in Figure 2.
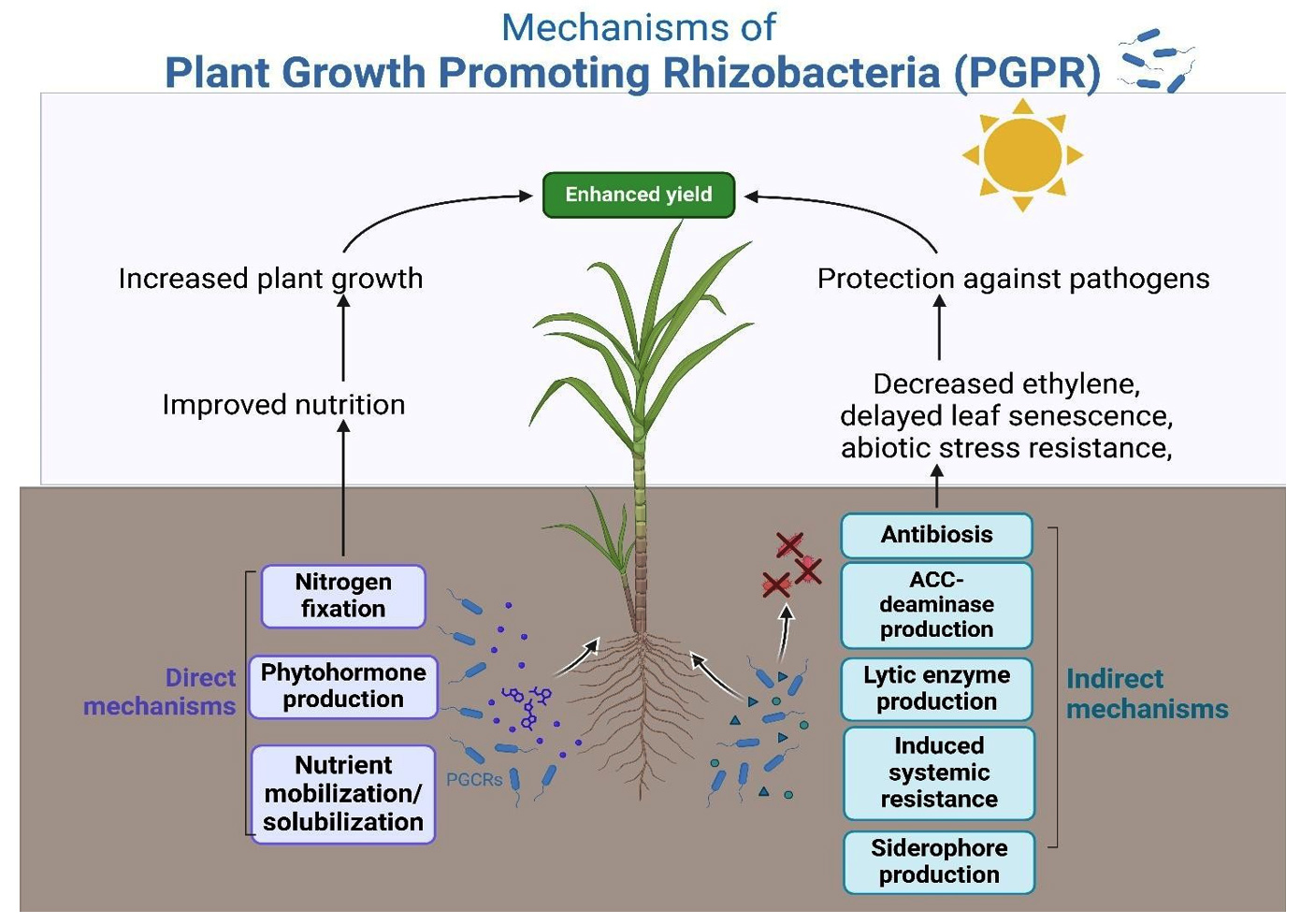
Figure 2. Mechanism of action of biocontrol agents against sugarcane diseases
Bacterial biocontrol agent to control sugarcane diseases
Certain Burkholderia species have been shown to have characteristics that inhibit disease and encourage plant development.55 Bacillus subtilis is a biocontrol agent that is successful in preventing F. verticillioides from proliferation.10 In sugarcane plants, the application of Bacillus decreased the prevalence of diseases by 45-49%.56 Pseudomonas fluorescens was used in a seed-based bioformulation that not only helped the plant grow but also reduced the prevalence of F. verticillioides, which causes disease in maize.57
The many PR-proteins were linked to the resistance against red rot that was produced by Pseudomonas.58 Acinetobacter, and Klebsiella pneumonia have antagonistic effects on sugarcane red rot.59 Bacillus strains that are antagonistic are preferable to other biocontrol agents in several aspects, including their widespread presence in soils, high sporulation, extended shelf life, and improved plant nutrition.60 The Bacillus amyloliquefaciens are potential antagonistic in the control of Pokkah Boeng in the sugarcane.61 Ochrobactrum intermedium (TRD14), Acinetobacter sp. (PK9), Bacillus sp. (RSC29 and KR91), and Escherichia sp. (VRE34) have been reported to exhibit inhibitory effects on the pathogen Colletotrichum falcatum, concurrently facilitating plant growth in sugarcane. These efficacious strains are utilized as significant bioinoculants for the integrated management of red rot disease in sugarcane.62
Fungal biocontrol agents
Trichoderma species have high antagonistic potential against various diseases in sugarcane. Plants that are subjected to Trichoderma spp. can develop systemic resistance, which is an improved defense state.63 Several scientists reported that T. viride and T. harzianum were the most efficient for preventing the infection of Pokkah Boeng disease.64 The pathogens F. fujikuroi and F. proliferatum can both have their maximum mycelial growth inhibited by the T. harzianum strain.63
In the rhizosphere, Trichoderma species effectively compete with Fusarium and other diseases for nutrients. Numerous enzymes that break down cell walls are produced by Trichoderma spp. These enzymes include massoia lactone, viridine, gliovirin, harzianic acid, alamethics, tricholine, peptaibols, and 6-penthyl-alpha-pyrone. These enzymes cause structural integrity to be disrupted and cell lysis to result from the breakdown of Fusarium and other fungi’s cell walls. Trichoderma species produce secondary metabolites having antifungal qualities. Heptilide acid and other bioactive substances are among them. Such antibiotic compounds are secreted, and they prevent Fusarium and other infections from growing.65 By producing both specific and nonspecific antibiotics such trichodemin, trichodermol, harzianun, and harzianolide, trichoderma are proven to eliminate plant pathogens.66
Trichoderma can inhibit the disease incidence of red rot in sugarcane. Soaking the setts in T. harzianum and then in farmyard manure can reduce the red rot infection. Combined inoculation of Trichoderma spp. (T. asperullum, T. atroviride, T. harzianum and T. auroviride) along with Paenibacillus alvei also effectively used as biocontrol agent for management of red rot in sugarcane.67-69
New trends in sugarcane disease management
RNA interference technology
Highlighting RNA interference technology as a key aspect of the investigation into Fusarium infection is grounded in its capacity to be an efficient method for managing fungal pathogens like Fusarium, responsible for noteworthy agricultural declines. RNA interference serves to precisely inhibit genes involved in pathogenicity or those that grant resistance to antifungal drugs. Through the silencing of critical genes within Fusarium, researchers can diminish the organism’s capacity to infect plants or disseminate, thus providing a more precise and environmentally sustainable alternative to conventional chemical interventions. RNA interference may be employed to attenuate the expression of genes associated with mycotoxin biosynthesis, consequently lessening their ramifications for food safety. Certain species of Fusarium have exhibited resistance to chemical fungicides. The advent of RNA interference technology presents a novel mechanism of action that has the potential to circumvent this resistance and bolster crop protection efforts. Post-transcriptional gene silencing (PTGS), referred to as RNA silencing or RNA interference (RNAi), is a conserved regulatory mechanism found in a wide range of eukaryotic organisms. This phenomenon plays a crucial role in the regulation of gene expression and serves as a defense mechanism against the invasion of external genetic elements, such as viruses and transposons. The essential steps in the process of RNA silencing involve the transfer of double-stranded RNA (dsRNA) molecules into the cellular environment. Various pathways can lead to this delivery, including viral infections or the introduction of artificial dsRNA. The dsRNA introduced undergoes a modification into small RNA molecules, typically 21-25 nucleotides long, recognized as small interfering RNAs (siRNAs) or microRNAs (miRNAs). This transformation process is facilitated by enzymes such as Dicer. The siRNAs or miRNAs are assimilated into a protein assembly called the RNA-induced silencing complex (RISC). The RISC complex uses the siRNA or miRNA as a guide to recognize and bind to complementary messenger RNA (mRNA) molecules. Once bound to the target mRNA, the RISC complex can induce cleavage of the mRNA or inhibit its translation. This resulted in the degradation of the target mRNA, preventing the synthesis of the corresponding protein.73 The RNAi technology is illustrated in Figure 3.
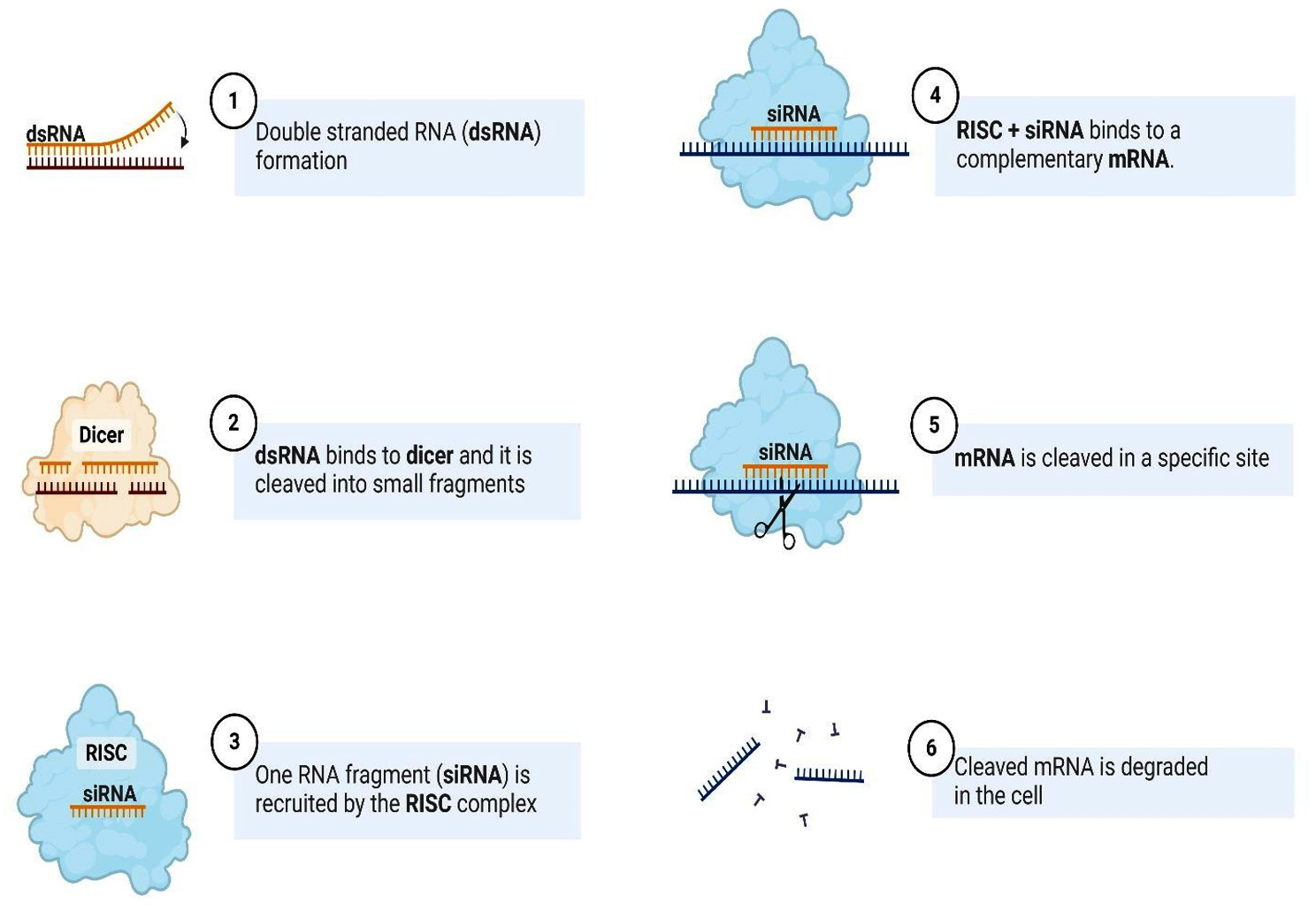
Figure 3. RNA interference technology for sugarcane diseases management
Host-induced Gene silencing in the Fusarium sp.
Transgenic plants generate siRNAs that can penetrate fungal infections and thereby mute fungal genes.74 It is commonly referred to as host-induced gene silencing (HIGS), where the target was the GUS (b-glucuronidase) originating from F. verticillioides. A hairpin-shaped 627-bp dsRNA fragment that targets the GUS gene is generated. The plant receives this hairpin dsRNA, most likely by genetic transformation techniques. And it generates siRNAs from the hairpin dsRNA structure and it is released into the plant tissues. Then it will be uptake by the fungal pathogen. The complementary GUS mRNA is targeted by the siRNAs. As a result, the fungi expression of the GUS gene decreases and the GUS mRNA is degraded.75 Furthermore, it aims to regulate Fusarium wilt by targeting the genes velvet and Fusarium transcription factor 1.76
RNA-controlled reduction of the PKS1 gene in C. falcatum
It is most likely a plasmid vector intended for RNA interference applications, called pSilent-1. The creation of hairpin RNA structures is made possible by this vector. Gene silencing is mediated by small interfering RNAs (siRNAs), which are produced through processing hairpin RNA structures. Hairpin constructs are made to specifically target PKS1 gene sequences.
These constructions are probably driven by a potent promoter that allows the PKS1 gene’s double-stranded RNA (dsRNA) to be expressed. The pSilent-1 vector containing the hairpin constructions is introduced into the fungal cells of C. falcatum using Agrobacterium-mediated transformation (ATMT) and leads to the synthesis of dsRNA and this dsRNA is converted into siRNAs that specifically target and degrade the PKS1 gene’s mRNA transcripts.77
Transcriptomics analysis
Transcription factors (TF) are recognized for their pivotal regulatory function within signaling pathways associated with both developmental processes and disease resistance mechanisms in Saccharum. TFs initiate various pathways through direct or indirect interactions by modulating critical genes implicated in plant defense responses, which are identified as the primary targets by pathogens.78 All of the transcripts present in a cell under particular circumstances are collectively referred to as the transcriptome. A wide array of RNA molecules, transcribed by specific cells under certain conditions, can be examined through transcriptomic sequencing. Stress affects cells with secondary metabolism and important meristem cells-stem cells and rapidly proliferating cells-differently. Therefore, different cell types or phases require distinct transcriptome analyses, and the foundation for resolving biological issues is a precise sampling method.79
Comparative transcriptome profiling of sugarcane phenotype against F. verticilliode
The etiological agent responsible for the development of Pokkah Boeng disease in sugarcane is the fungal pathogen F. verticillioides. Transcriptome analyses were conducted on the sugarcane varieties “YT94/128” and “GT37”, which exhibit resistance and susceptibility to F. verticillioides, respectively, both pre-and post-inoculation. The remarkable association between the key Differentially Expressed Genes (DEGs) linked to resistance and various metabolic pathways such as phenylpropanoid biosynthesis, cutin, suberine, and wax biosynthesis, nitrogenous metabolism, and plant-pathogen interactions in these two cultivars. The DEGs were mostly tied to a variety of categories including catalytic enzyme activity, cellular protease actions, hydrolytic enzymes, peptide enzymes, the inhibition of protein metabolic processes, phenylpropanoid metabolism, the extracellular region, aldehyde dehydrogenases, endopeptidases, REDOX enzymes, protein kinases, and phosphoric acid transferases, which act as facilitators in the biosynthesis of secondary metabolites, as well as in plant-pathogen interactions.80
Comparative transcriptome profiling of sugarcane phenotype against C. falcatum
The interactions between the host organism and the pathogen, specifically involving sugarcane and C. falcatum (the causative agent of red rot), have facilitated the discovery of numerous pathogenesis-related (PR) proteins. Noteworthy contributions to the plant’s defense mechanisms come from chitinase, b-1,3 glucanase, and thaumatin-like proteins among the identified proteins. Moreover, the detection of 3-deoxyanthocyanidin phytoalexins at the loci of infection has been confirmed as a crucial marker of resistance against red rot.81 The exploration of transcription factors (TFs) within the framework of sugarcane’s defense mechanisms against fungal pathogens has clarified the expression of numerous TFs, significantly featuring the WRKY family, recognized for its contribution to mediating pathogen-associated molecular patterns (PAMP) and systemic acquired resistance (SAR).82 Specifically, the expression of WRKY transcription factors was identified during the interaction with C. falcatum, representing the inaugural extensive documentation in the context of sugarcane. The transcription factors are crucial in complex signaling networks that regulate reactions to both biological and environmental stressors.83 The NAC family represents another vital set of transcription factors (TFs) that is instrumental in modulating both innate immune responses and effector-triggered immunity (ETI). A comprehensive transcriptome analysis demonstrated that 21 NAC transcription factors were expressed during the interaction between sugarcane and C. falcatum, with 11 NACs being activated during the priming of systemic acquired resistance (SAR).84
PGPRs function as biological control agents by engaging in direct antagonistic interactions with pathogens, employing strategies like antibiosis, competition for resources, and mycoparasitism. They serve to safeguard sugarcane cultivation from afflictions such as red rot, wilt, and Pokkah Boeng. Additionally, they trigger the plant’s innate defense mechanisms, eliciting a systemic resistance reaction that offers protection against a diverse array of diseases. It contributes to nutrient cycling, solubilizing essential minerals, and promoting their uptake by plants, which enhances overall plant health and resilience. Integrating into comprehensive IPM strategies, complementing other control methods, and promoting a holistic and environmentally friendly approach to disease management and most commonly Bacillus and Pseudomonas control the disease incidence. PGPRs not only protect sugarcane against diseases but also positively influence plant growth, leading to improved yields and overall crop performance and they increase sugarcane’s ability to withstand both biotic and abiotic stresses, making them valuable components in crop resilience strategies. In both research and therapeutic applications, RNA interference has proven to be an adaptable and innovative tool. Future RNAi-based methods are expected to overcome obstacles and expand their coverage through ongoing research and technology developments. Also, new emerging technology transcriptomics has emerged as a fundamental field in biological and medical sciences, offering an abundance of knowledge about regulatory processes and patterns of gene expression. Our understanding of biological processes will grow as a result of the continuous integration of transcriptomics with other omics technologies and the creation of sophisticated analytical tools, which will also benefit biotechnology and medicine.
ACKNOWLEDGMENTS
The authors express gratitude to Kothari Sugars, Trichy, India, for providing a student fellowship to support postgraduate research.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Comstock JC; Saumtally AS; Rott P; Bailey RA; Croft BJ; A guide to sugarcane diseases. Quae. 2000
- Satyavir S. Prof. MS Pavgi Award Lecture-Red rot of sugarcane-Current Scenario-satyavir. Indian Phytopathol. 2012:56(3):245-254.
- Viswanathan R, Rao GP. Disease scenario and management of major sugarcane diseases in India. Sugar Tech. 2011;13(4):336-353.
Crossref - Viswanathan R. Fusarium diseases affecting sugarcane production in India. Indian Phytopathol. 2020;73(3):415-424.
Crossref - Wang ZP, Sun HJ, Guo Q, et al. Artificial inoculation method of Pokkah Boeng disease of sugarcane and screening of resistant germplasm resources in subtropical China. Sugar Tech. 2017;19(3):283-92.
Crossref - Tiwari A, Bharti Y, Tripathi S, et al. Biotechnological approaches to improve sugarcane crop with special reference to disease resistance. Acta Phytopathol Entomol Hung. 2010;45(2):235-249.
Crossref - Pandey C, Shahi NV, Pawar BH. Synergistic effect of chitosan and Trichoderma viride against C. paradoxa, the causal agent of pineapple disease in sugarcane. J Rur Agric Res. 2014;14(2):70-74.
- Pal KK, Gardener BM. Biological control of plant pathogens. The Plant Health Instructor. 2006;6(5):1117-1142.
Crossref - O’Brien PA. Biological control of plant diseases. Australas Plant Pathol. 2017a;46(4):293-304.
Crossref - Patil AS, Singh H, Sharma SH, Rao GP. Morphology and pathogenicity of isolates of Fusarium moniliforme causing Pokkah Boeng disease of sugarcane in Maharashtra. Microbial Diversity: Modern Trends. 2007:234-263.
- Ashwin NMR, Barnabas L, Sundar AR, et al. Comparative secretome analysis of Colletotrichum falcatum identifies a cerato-platanin protein (EPL1) as a potential pathogen-associated molecular pattern (PAMP) inducing systemic resistance in sugarcane. J Proteom. 2017;169:2-20.
Crossref - Viswanathan R, Samiyappan R. Bio-formulation of fluorescent Pseudomonas spp. induces systemic resistance against red rot disease and enhances commercial sugar yield in sugarcane. Arch Phytopathol Pflanzenschutz. 2008;41(5):377-88.
Crossref - Hassan MN, Afghan S, Hafeez FY. Erratum to: Suppression of red rot caused by Colletotrichum falcatum on sugarcane plants using plant growth-promoting rhizobacteria. Biocontrol. 2010;55(4):531-542.
Crossref - Viswanathan R. Pathogen virulence in sugarcane red rot pathogen versus varieties in cultivation: Classical case of loss in virulence in the pathotype CF06 (Cf671). Sugar Tech. 2017;19(3):293-299.
Crossref - Viswanathan R. Plant disease: Red rot of sugarcane. Anmol Publications. 2010.
- Silva TC, Moreira SI, Felizardo LM, et al. Dry rot caused by the complex Colletotrichum falcatum and Thielaviopsis paradoxa emerges as a key stalk disorder in newly expanded sugarcane plantations from Northwestern Sao Paulo, Brazil. Agronomy. 2023;13(11):2729.
Crossref - Harish S, Manikandan R, Durgadevi D, Raguchander T. Population Diversity of Fusarium spp. and its Interaction with Plant Growth Promoting Rhizobacteria. The Phytopathogen. 2017:417-435.
Crossref - Lin Z, Xu S, Que Y, et al. Species-specific detection and identification of Fusarium species complex, the causal agent of sugarcane Pokkah Boeng in China. PloS one. 2014;9(8):e104195.
Crossref - Govender P, McFarlane SA, Rutherford RS. Fusarium species causing pokkah boeng and their effect on Eldana saccharina Walker (Lepidoptera: Pyralidae). In Proc. S. Afr. Sug. Technol. Ass. 2010: 267-270.
- Nordahliawate MS, Izzati MZ, Azmi AR, Salleh B. Distribution, Morphological Characterization and Pathogenicity of Fusarium sacchari Associated with Pokkah Boeng Disease of Sugarcane in Peninsular Malaysia. Pertanika J Trop Agric Sci. 2008;31(2): 279-286.
- Vishwakarma SK, Kumar P, Nigam A, Singh A, Kumar A. Pokkah Boeng: an emerging disease of sugarcane. J Plant Pathol Microbiol. 2013;4(170):2.
Crossref - Viswanathan R, Balaji CG, Selvakumar R, et al. Epidemiology of Fusarium diseases in sugarcane: a new discovery of same Fusarium sacchari causing two distinct diseases, wilt and Pokkah Boeng. Sugar Tech. 2017;19:638-46.
Crossref - Raghvendra T, Shukla SK, Jaiswal VP, et al. Isolation and Molecular Identification of Fusarium spp., associated with Pokkah Boeng disease of sugarcane. J Biotech. 2021;16:4.
- Viswanathan R, Malathi P, Anna A, Naveen CP, Scindiya M. Sudden occurrence of wilt and Pokkah Boeng in sugarcane and status of resistance in the parental clones in national hybridization garden to these diseases. J Sugarcane Res. 2014;4(1):62-81.
- Viswanathan R, Malathi P, Sundar AR, Poongothai M, Singh N. Current status of sugarcane wilt in India. Sug. internat. 2006:4(1);3-7.
- Costa MM, Melo MP, Guimarדes EA, et al. Identification and pathogenicity of Fusarium species associated with Pokkah Boeng of sugarcane in Brazil. Plant Pathol. 2019;68(7):1350-1360.
Crossref - Vijaya HK, Srikant SK, Hegde YR. Chemical control of sett rot of sugarcane caused by Ceratocystis paradoxa. Karnataka j. Agri. Sci. 2007;62-64.
- Kuo TT, Chien MM, Li HW. Ethyl acetate produced by Ceratocystis paradoxa and C. adiposum and its role in the inhibition of the germination of sugarcane buds. Can J Bot. 1969;47(9):1459-1463.
Crossref - Bertaccini A, Lee IM. Phytoplasmas: an update. Phytoplasmas: Plant pathogenic bacteria-I: Characterisation and epidemiology of phytoplasma-associated diseases. 2018:1-29.
Crossref - Venkataravanappa V, Suryanarayana V, Upadhya V, Pai V, Rao GP, Reddy CNL. First record of ‘Candidatus phytoplasma asteris’ (16SrI group) associated with Annona reticulata L. in India. Indian Phytopathol. 2023;76(3):897-901.
Crossref - Viswanathan R, Chinnaraja C, Karuppaiah R, Kumar VG, Rooba JJ, Malathi P. Genetic diversity of sugarcane grassy shoot (SCGS)-phytoplasmas causing grassy shoot disease in India. Sugar Tech. 2011;13(3):220-228.
Crossref - Nejat N, Vadamalai G. Diagnostic techniques for detection of phytoplasma diseases: past and present. J Plant Dis Prot. 2013;120(1):16-25.
Crossref - Lo Iacono G, van den Bosch F, Gilligan CA. Durable resistance to crop pathogens: an epidemiological framework to predict risk under uncertainty. PLoS Comput Biol. 2013;9(1):e1002870.
Crossref - Hayasaka T, Ishiguro K, Shibutani K, Namai T. Seed disinfection using hot water immersion to control several seed-borne diseases of rice plants. Japanese J Phytopathol. 2001;67(1):26-32.
Crossref - Anuradha KL, Kumar R, Singh P. Sugarcane Grassy Shoot (SCGS) disease-an overview. Ind J Pure App Biosci. 2019;7(4):371-378.
Crossref - Sunder S, Satyavir. Vegetative compatibility, biosynthesis of GA3 and virulence of Fusarium moniliforme isolates from bakanae disease of rice. Plant Pathology. 1998;47(6):767-772.
- Manandhar T, Yami KD. Biological control of foot rot disease of rice using fermented products of compost and vermicompost. Scientific World. 2008;6(6):52-57.
Crossref - Agnihotri VP. Current sugarcane disease scenario and management strategies. Ind Phytopathl. 1996;49(2):109-126.
- Sharma R., Tamta S. A Review on red rot: The “cancer” of sugarcane. J Plant Pathol Microbiol. 2015;S1:1-8.
Crossref - Gupta RN, Sah SB, Kumar S, Kumar A, Kishore C, Chand G. Impact of red rot disease on nutrient status of sugarcane. Int J Curr Microbiol App Sci. 2018;Speical Issue7:3533-3538.
- Bhuiyan SA, Croft BJ, Tucker GR. Efficacy of the fungicide flutriafol for the control of pineapple sett rot of sugarcane in Australia. Australas Plant Pathol. 2014;43(4):413-419.
Crossref - Malathi P, Padmanaban P, Viswanathan R, Mohanraj D, Sundar AR. Efficacy of thiophanate methyl against red rot of sugarcane. Acta Phytopathol Entomol Hung. 2004;39(1-3):39-41.
Crossref - Ora N, Faruq AN, Islam MT, Akhtar N, Rahman MM. Detection and identification of seed borne pathogens from some cultivated hybrid rice varieties in Bangladesh. Middle-East J Sci Res. 2011;10(4):482-488.
- Latif MA, Uddin MB, Rashid MM, et al. Rice bakanae disease: yield loss and management issues in Bangladesh. Food Sci Technol. 2021;9(1):7-16.
Crossref - Bharti YP, Singh BK, Kumar A, Singh SP, Shukla DN. Efficacy of fungicides and antibiotics against spore germination and sporulation of Colletotrichum falcatum Went:causing red rot disease of sugarcane in vitro and in vivo condition. Agriways. 2014;2(2):100-105.
- Mendes R, Pizzirani-Kleiner AA, Araujo WL; Raaijmakers JM. Diversity of cultivated endophytic bacteria from sugarcane: genetic and biochemical characterization of Burkholderia cepacia complex isolates. App. Environ. Micro. 2007:73(22):7259-7267
- Arya A, Sharma R, Sharma G, Kabdwal BC, NEgi A, Mishra B. Evaluation of fungal and bacterial antagonists for managing phytopathogen Fusarium moniliforme var. subglutinans Sheldon, causing Pokkah Boeng disease of sugarcane. Journal of Biological Control. 2017;31(4):217-222.
- Joshi D, Singh P, Holkar SK, Kumar S. Trichoderma-mediated suppression of red rot of sugarcane under field conditions in subtropical India. Sugar Tech. 2019;21(3):496-504.
Crossref - Vinale F, Sivasithamparam K, Ghisalberti EL, Marra R, Woo SL, Larito M. Trichoderma-plant-pathogen interactions. Soil Biol Biochem. 2008;40(1):1-10.
Crossref - Harish S, Parthasarathy S, Prabhukarthikeyan SR, et al. Omics: A potential tool to delineate the mechanism of biocontrol agents against plant pathogens. Plant-Microbe Interactions. 2022;11:79-100.
Crossref - Shaterian J, Waterer D, De Jong H, Tanino KK. Differential stress responses to NaCl salt application in early-and late-maturing diploid potato (Solanum sp.) clones. Environ Exp Bot. 2005;54(3):202-212.
Crossref - Karlidag H, Esitken A, Turan M, Sahin F. Effects of root inoculation of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient element contents of leaves of apple. Sci Hortic. 2007;114(1):16-20.
Crossref - Belimov AA, Dodd IC, Safronova VI, et al. Abscisic acid metabolizing rhizobacteria decrease ABA concentrations in planta and alter plant growth. Plant Physiol Biochem. 2014;74:84-91.
Crossref - Bhattacharyya PN, Jha DK. Plant growth-promoting rhizobacteria (PGPR):emergence in agriculture. World J Microbiol Biotechnol. 2012;28(4):1327-1350.
Crossref - Mendes R, Pizzirani-Kleiner AA, Araujo WL, Raaijmakers JM. Diversity of cultivated endophytic bacteria from sugarcane:genetic and biochemical characterization of Burkholderia cepacia complex isolates. Appl Environ Microbiol. 2007;73(22):7259-7267.
Crossref - Hassan MN, Afghan S, Hafeez FY. Biological suppression of sugarcane red rot by Bacillus spp. under field conditions. Plant Pathol J. 2012;94(2):325-329. http://www.jstor.org/stable/45156041
- Singh PC, Nautiyal CS. A novel method to prepare concentrated conidial biomass formulation of Trichoderma harzianum for seed application. J Appl Microbiol. 2012;113(6):1442-1450.
Crossref - Viswanathan R, Samiyappan R. Pseudomonas spp. colonization in sugarcane rhizosphere reduces titre of Colletotrichum falcatum Went-causing red rot disease of sugarcane. Arch Phytopathol Pflanzenschutz. 2006;39(1):39-44.
Crossref - Bhardwaj G, Shah R, Joshi B, Patel P. Klebsiella pneumoniae VRE36 as a PGPR isolated from Saccharum officinarum cultivar Co99004. J Appl Biol Biotechnol. 2017;5(1):47-52.
Crossref - Hassan MN, Namood-e-Sahar, Zia-Ul-Husnain SS, Afghan S, Hafeez FY. Suppression of red rot disease by Bacillus sp. based biopesticide formulated in non-sterilized sugarcane filter cake. BioControl. 2015;60:(5):691-702.
Crossref - Manjula PAK, Kishore VP, Manoj KV, Sandhya RC. Efficacy of Bacillus spp., against Fusarium sacchari causing Pokkah boeng disease of sugarcane under in vitro condition. Management. 2017;SP-7(2):468-473.
- Patel P, Shah R, Joshi B, Ramar K, Natarajan A. Molecular identification and biocontrol activity of sugarcane rhizosphere bacteria against red rot pathogen Colletotrichum falcatum. Biotechnol Rep. 2019;21:e00317.
Crossref - Tiwari R, Shukla SK, Jaiswal VP, et al. Bio-control potential of Trichoderma spp., against Fusarium spp., the incitants of Pokkah Boeng disease of sugarcane under in-vitro conditions. Indian Phytopathol. 2021;74(3):691-701.
Crossref - Joshi D, Misra SC. Characterization of Trichoderma isolates from sugarcane agro-ecosystem and their efficacy against Colletotrichum falcatum causing red rot of sugarcane. Sugar Tech. 2013;15:192-196.
Crossref - Sharma P. Complexity of ‘Trichoderma-fusarium’ interaction and manifestation of biological control. Aust J Crop Sci. 2011;5(8):1027-1038.
- Bhattacharjee R, Dey U. An overview of fungal and bacterial biopesticides to control plant pathogens/diseases. Afr J Microbiol Res. 2014;8(17):1749-1762.
Crossref - Yadav RL, Vijai Singh VS, Srivastav SN, et al. Use of Trichoderma harzianum for the control of red rot disease of sugarcane. Sugar Cane International. 2008;26-31.
- Singh V, Joshi BB, Awasthi SK, Srivastava SN. Eco-friendly management of red rot disease of sugarcane with Trichoderma strains. Sugar Tech. 2008;10(2):158-161.
Crossref - Elamathi E, Malathi P, Viswanathan R, Sundar AR. Potential of Paenibacillus and Trichoderma isolates against Colletotrichum falcatum causing red rot disease in sugarcane. 2016;6(2):1-12.
- Hassan MN, Afghan S, Hafeez FY. Biological control of red rot in sugarcane by native pyoluteorin producing Pseudomonas putida strain NH 50 under field conditions and its potential modes of action. Pest Manag Sci. 2011;67(9):1147-1154.
Crossref - Yamunarani K, Sangeetha S, Chitra K, Dhanalakshmi K, Dhanushkodi V. Developing Biocontrol Strategies for the Management of Set rot Disease of Sugarcane under Wider Row Planting. J Sci Res Rep. 2023;29(12):67-76.
Crossref - Singh SB, Rao GP, Easwaramoorthy S. Sugarcane crop management. Houston:Science and Technology Publishing Llc. 2002.
- Pyott DE, Molnar A. Going mobile: non cell autonomous small RNAs shape the genetic landscape of plants. Plant Biotechnol J. 2015;13(3):306-318.
Crossref - Machado AK, Brown NA, Urban M, Kanyuka K, Hammond-Kosack KE. RNAi as an emerging approach to control Fusarium head blight disease and mycotoxin contamination in cereals. Pest Manag Sci. 2018;74(4):790-799.
Crossref - Tinoco MLP, Dias BB, Dall’Astta RC, Pamphile JA, Aragao FJL. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biol. 2010;8:1-11.
Crossref - Ghag SB, Shekhawat UKS, Ganapathi TR. Host induced post transcriptional hairpin RNA mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium wilt in banana. Plant Biotechnol J. 2014;12(5):541-553.
Crossref - Scindiya M, Malathi P, Kaverinathan K, Sundar AR, Viswanathan R. RNA-mediated silencing of PKS1 gene in Colletotrichum falcatum causing red rot in sugarcane. Eur J Plant Pathol. 2019;153:371-84.
Crossref - Javed T, Shabbir R, Ali A, et al. Transcription Factors in Plant Stress Responses: Challenges and Potential for Sugarcane Improvement. Plants. 2020;9:491.
Crossref - Marguerat S, Bהhler J. RNA-seq:from technology to biology. Cell Mol Life Sci. 2010;67(4):569-579.
Crossref - Wang Z, Li Y, Li C, Song X, Lei J, Gao Y, Liang Q. Comparative transcriptome profiling of resistant and susceptible sugarcane genotypes in response to the airborne pathogen Fusarium verticillioides. Mol Biol Rep. 2019;46(2):3777-3789.
Crossref - Viswanathan R, Nandakumar R, Samiyappan R. Involvement of pathogenesis-related proteinsin Pseudomonas spp. induced systemic resistance against Colletotrichum falcatum in sugarcane. JPlant Dis Protect. 2003;110:524-537.
- Che F. Hu Y, Vannozzi A, et al. The WRKY transcription factor family in model plants and crops. Crit Rev Plant Sci. 2017;36(5-6):311-335.
Crossref - Higuera JJ, Garrido-Gala J, Lekhbou A, et al. The strawberry FaWRKY1 transcription factor negatively regulates resistance to Colletotrichum acutatum in fruit upon infection. Front Plant Sci. 2019;10:480.
Crossref - Muthumeena M, Ashwin R, Sundar AR, Malathi P, Viswanathan R. Expression profiling of transcription factors (TFs) in sugarcane X Colletotrichum falcatum interaction. J Plant Biochem Phsiol. 2013;22(3):286-294.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.