


ISSN: 0973-7510
E-ISSN: 2581-690X
In the realm of sericulture, ericulture stands as a beacon of hope for rural communities, fostering economic growth and livelihood prospects. However, the influence of disease, particularly flacherie, threatens to undermine this progress. To combat this menace, researchers embarked on a quest to unmask the causative agents behind bacterial flacherie in Samia ricini. For this, flacherie-infected larvae were collected, and hemolymph was isolated and streaked onto Petri plates having nutrient agar media. Yellowish colonies developed within 48-72 hours at 37 °C were subjected to identification through colony morphology study, biochemical tests, and 16S rRNA gene sequencing. The colonies were identified as Stenotrophomonas maltophilia, Stenotrophomonas rhizophila, and Kocuria rhizophila. The study reveals that the S. rhizophila, once deemed harmless, has the potential to cause bacterial flacherie in S. ricini and highlights their associated risks. This research documented the infestation of Stenotrophomonas maltophilia, Stenotrophomonas rhizophila, and Kocuria rhizophila in Samia ricini for the first time. This knowledge may further empower the sericulture industry to craft tailored solutions to mitigate flacherie, ensuring the resilience of rural livelihoods and the sustained growth of sericulture.
Pathogenicity, Coccus, Bacillus, NCBI BLAST, FESEM
Eri silkworm rearing is a vital economic activity in northeast India, contributing 21.10% to the nation’s silk production.1 However, the silkworms are susceptible to microbial contaminants like viruses, bacteria, fungi, and protozoa,2,3 which can significantly impact their growth, development, and productivity. Flacherie, a bacterial and viral disease, is a significant threat to eri silkworms, causing detrimental changes to their physiology and metabolism.4 This disease is characterized by symptoms such as sluggishness, vomiting of gut juices, loss of appetite, semi-solid, chain-type excreta, alteration of body color, and a foul smell.5-7
Various pathogenic bacteria, including Bacillus thuringiensis, Bacillus cereus, Bacillus bombysepticus, and Pseudomonas aeruginosa, have been identified as causative agents of flacherie in silkworms.2,8-11 The disease can lead to significant economic losses for sericulture farmers, emphasizing the need for effective management strategies. Factors such as overcrowding, malnutrition, insufficient cleaning, fluctuations in temperature and humidity, and contaminated or decayed leaves can encourage microbial growth and infection.
While research has primarily focused on Bombyx mori, the Indian mulberry silk moth, this study aims to identify infectious bacterial colonies isolated from Samia ricini, the eri silkworm. By understanding the potentially harmful impacts of bacteria on S. ricini’s health, we can develop targeted interventions to improve disease management and promote sustainable sericulture practices. This study will contribute to the growing body of research on eri silkworm health and provide valuable insights for the sericulture industry.
Collection of infected larvae and culture of bacteria
Under aseptic conditions, flaccid fifth instar larvae of eri silkworms (n = 30) were collected from three eri silkworm rearing sites located in Nayekgaon, Harinaguri, and Gendrabil village of Kokrajhar district, where sericulture is a prominent livelihood for the inhabitants. These infected silkworms, carried in sterilized collecting jars, were further rinsed twice with sterile distilled water and surface sterilized using 70% ethanol for 60 seconds under an aseptic environment. A final rinse with sterile distilled water followed. The hemolymph of infected silkworms was collected in Eppendorf tubes, and a loopful of hemolymph was streaked onto nutrient agar plates. These plates were incubated at a range of 30-37 °C for 48 hours.12,13 Dominant bacterial colonies grown on the plates were identified, subcultured for purity, and stored in sterile glycerol (15%) in screw-cap microfuge tubes (1:1 ratio) at -80 °C for further use.14
Identification of bacterial species
Colony morphology
Colony characteristics such as size, form, margin, elevation, and colour were studied following different methods.15-21
Gram staining method
Bacterial smears were prepared on slides, heat-fixed, and stained sequentially with crystal violet (1-2 minutes), Gram’s iodine (1-2 minutes), and ethanol for decolorization (15-20 seconds). Safranin (30 seconds-1 minute) was used as the counterstain. Slides were observed under a Leica DM750 microscope using immersion oil at 100 × magnification.22
Field Emission Scanning Electron Microscopy (FESEM)
For SEM, bacterial cultures in the nutrient broth were centrifuged at 7000 rpm for 10 minutes to collect pellets. The pellets were washed with 1 × PBS buffer, fixed with 2.5% glutaraldehyde overnight, and washed again with 1 × PBS buffer. Dehydration was carried out using graded ethanol concentrations (30%-100%), with centrifugation at 7000 rpm for 5 minutes at each step. Samples were spread on coverslips under a Stereo Zoom microscope (Stemi 508), gold-sputtered, and observed using a SIGMA VP FESEM (ZEISS) at 15.00 KX and 20.00 KX magnification.
Biochemical tests
IMViC tests (Indole, Methyl Red, Voges–Proskauer, Citrate) and carbohydrate utilization tests (glucose, adonitol, arabinose, lactose, sorbitol, mannitol, rhamnose, sucrose) were conducted using HiIMViCTM Biochemical Test Kits (HiMedia). Wells were aseptically inoculated with bacterial samples and incubated at 35-37 °C for 18-24 hours. Post-incubation, reagents such as Kovac’s, Methyl Red, and Barritt’s were added to respective wells for specific tests. Colour changes in wells were interpreted using the standard chart provided with the kits.
Molecular identification method
The 16S rRNA gene sequencing and the NCBI BLAST similarity search tool were used to identify the bacterial species. Genomic DNA was extracted using the Biobee Spin EXpure Microbial DNA Isolation Kit (Bogar Bio Bee Stores Pvt. Ltd., Tamil Nadu, India). PCR amplification of the 16S rRNA gene was performed using primers 27F (5′-AGAGTTTGATCTGGCTCAG-3′) and 1492R (5′-TACGGTACCTTGTTACGACTT-3′) in a reaction mixture containing Taq Master Mix, deionized water, and primers. Unincorporated primers and dNTPs were removed using a Montage PCR Clean-up Kit (Millipore). Sequencing was performed with an ABI PRISM® BigDye™ Terminator Cycle Sequencing Kit using an ABI 3730xl sequencer. Sequences were aligned using MUSCLE 3.7,23 and curated with Gblocks 0.91b24 to remove alignment noises. Phylogenetic analysis was conducted using PhyML 3.0 with the HKY85 substitution model. Sequences were submitted to the NCBI nucleotide database for GenBank accession numbers.
Further, to confirm the pathogenic effects of the isolated bacterial species, a pathogenicity test was performed in vitro on healthy fifth instar larvae. Bacterial culture (40 ml) was added to 5 sterile 50 ml Falcon tubes, centrifuged for 7 minutes at 7000 rpm, and the pellets were resuspended in 20 ml PBS buffer. Pathogenicity was tested by injecting 10 µl bacterial suspension into silkworms (n = 30) by U-40 insulin injection (0.25 mm (31G) × 6 mm needle) (BD Ultra-FineTM Needle Insulin Syringes) and dipping them in 100 µl suspension (n = 30) into petri plates (90 × 17 mm), with continuous supply of R. communis leaves, and observations were recorded.
The study found a total of eight colonies, of which three colonies (S1, S2, and S3) were selected for identification. These three colonies were dominant in the nutrient media and exhibited intriguing characteristics, such as a colour change corresponding to the incubation time, rapid growth within 24 hours, and their presence in all the petri plates. All the bacterial colonies showed luxuriant growth at 37 °C after 48 hours. Although many bacterial species prefer low temperatures to grow, the inherent phenotypic variability, adaptability, and genetic similarities enable a species to withstand rising temperatures and attain good growth. Additionally, the capacity of a species to grow at 37 °C is often considered a key trait in differentiating potential pathogens from harmless environmental strains within a particular genus.25,26
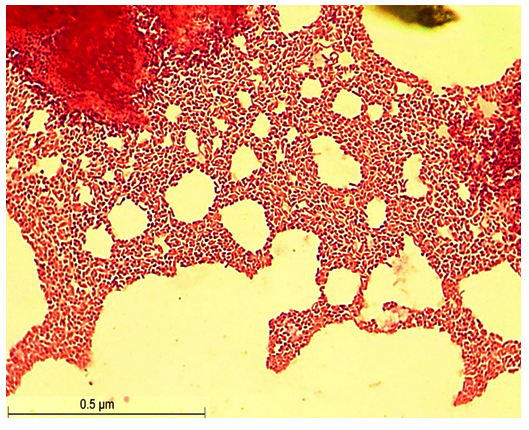
Figure 1. Gram Staining of S1 isolate
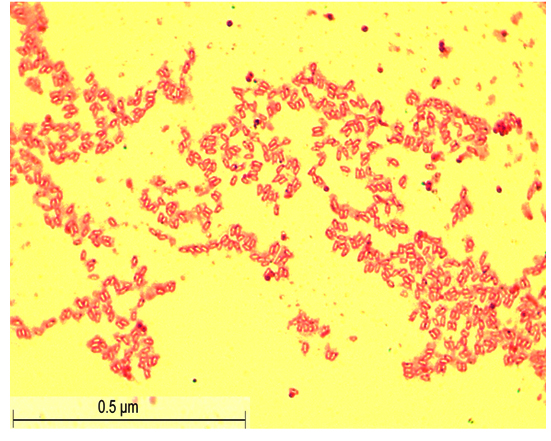
Figure 2. Gram Staining of S2 isolate
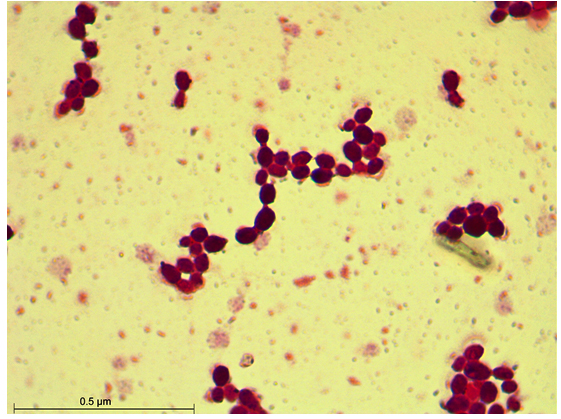
Figure 3. Gram Staining of S3 isolate
Colony morphology of the isolates
After 48 hours of incubation at 37 °C, the dominant colonies (S1, S2, S3) on nutrient agar appeared circular with entire margins and formed small (2-3 mm) yellowish colonies in the culture media. The S3 colony had a slight convex elevation, while S1 and S2 had total convex elevation. The S1 and S2 colonies were rod-shaped, pinkish-red Gram-negative bacilli, and the circular S3 colonies were stained purple and were arranged in various patterns, such as short chains, tetrads, grapelike packets, and irregular clusters, representing the Gram-positive coccus type of bacteria (Figures 1-3). Scanning electron microscopy of S1 and S2 revealed the presence of rod-shaped structures having lipopolysaccharide layers on their surfaces that facilitate cell-to-cell adhesion, confirming the bacillus type of bacteria and S3 as coccus (Figures 4-9) (Table 1).
Table (1):
Colony morphology of the isolates
Isolates |
Size |
Shape |
Type |
Colour |
Elevation |
Margin |
Gram-Staining |
|---|---|---|---|---|---|---|---|
S1 |
Small (2-3 mm) |
Circular |
Bacillus |
Yellow |
Convex |
Entire |
Negative |
S2 |
Small (2-3 mm) |
Circular |
Bacillus |
Yellow |
Convex |
Entire |
Negative |
S3 |
Small (2-3 mm) |
Circular |
Bacillus |
Yellow |
Slightly convex |
Entire |
Positive |

Figure 4. FESEM of S1 isolate (15.00 KX)
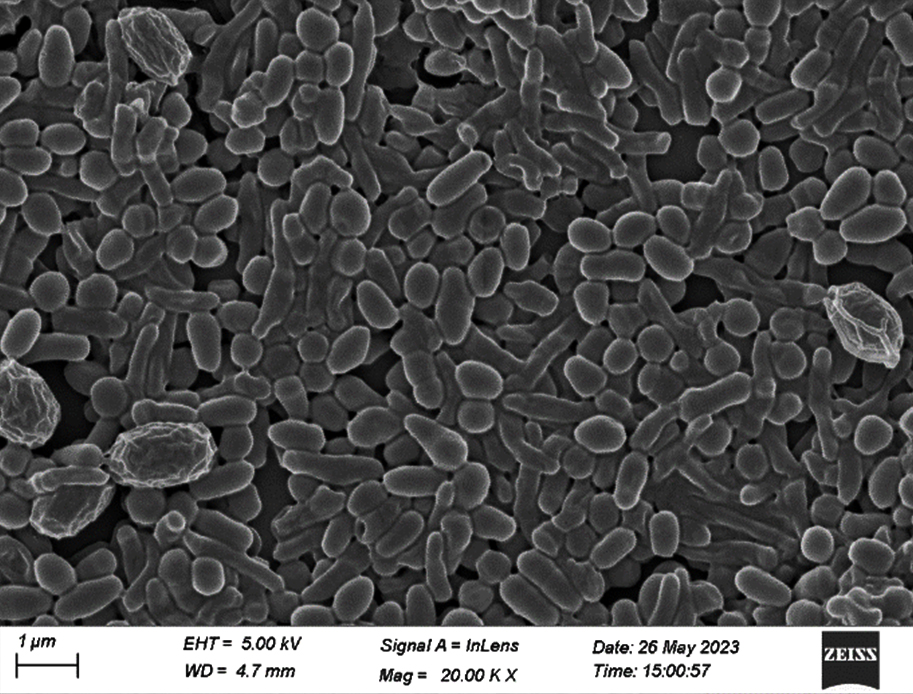
Figure 5. FESEM of S1 isolate (20. 00 KX)

Figure 6. FESEM of S2 isolate (15.00 KX)

Figure 7. FESEM of S2 isolate (20.00 KX)
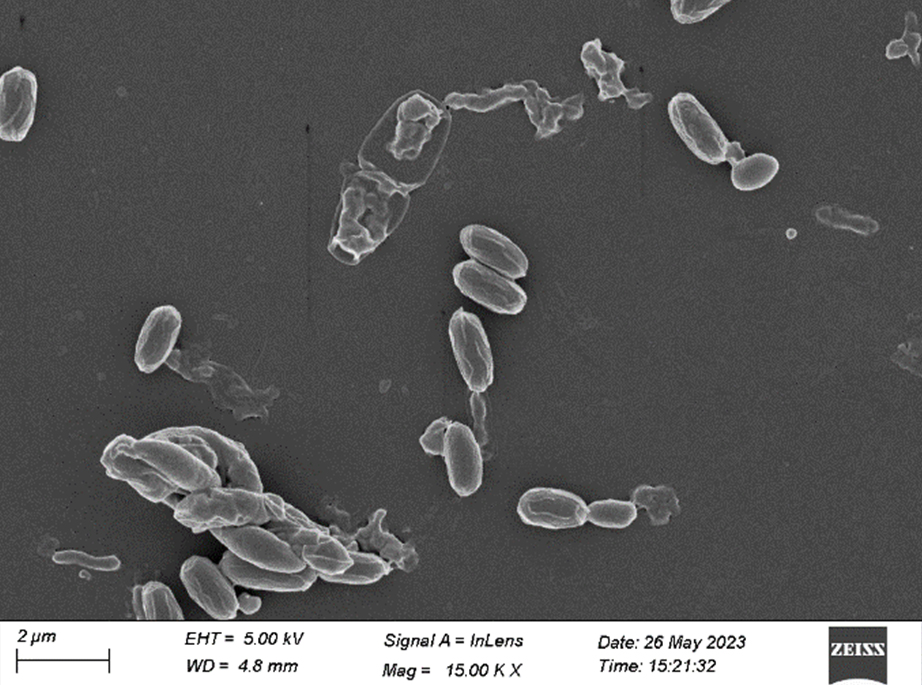
Figure 8. FESEM of S3 isolate (15.00 KX)
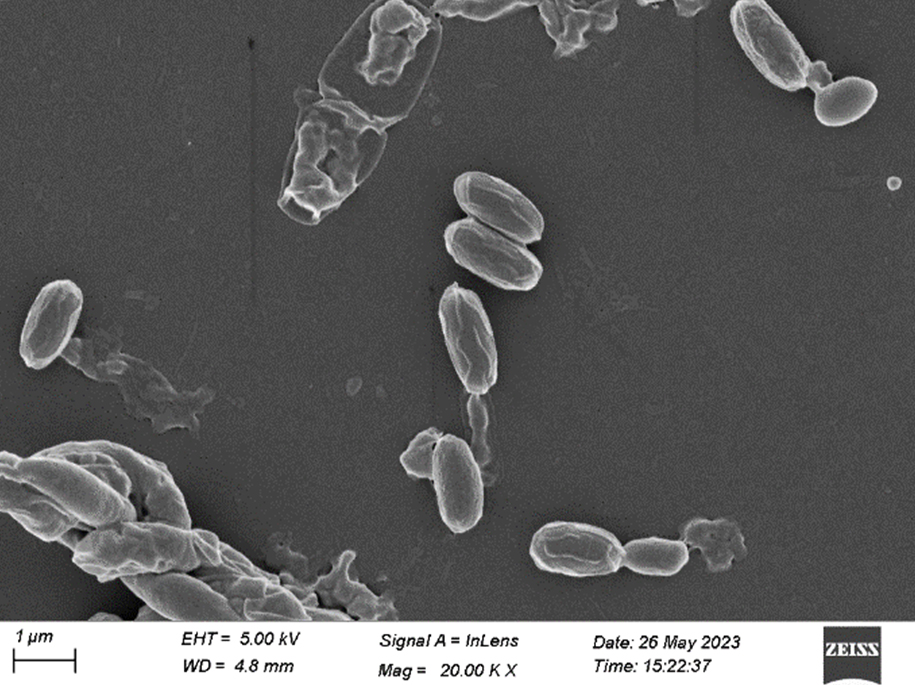
Figure 9. FESEM of S3 isolate (20.00 KX)
Biochemical tests
All the selected bacterial isolates (S1, S2, and S3) showed a negative result for the indole and methyl red tests, and a positive result for the citrate utilization test. Colony S1 and S2 showed negative results for Voges Proskauer’s test, while S3 showed positive results for the same (Table 2).
In the carbohydrate utilization test, the selected isolates demonstrated an inability to ferment lactose, sorbitol, mannitol, and rhamnose, resulting in negative results after 18-24 hours of incubation. However, the isolates were able to ferment glucose. Apart from this, S1 and S2 isolates were able to ferment sucrose and were unable to ferment adonitol and arabinose, while S3 was unable to ferment sucrose and able to ferment adonitol and arabinose (Table 2).
Table (2):
Biochemical tests of the isolates
Biochemical tests |
S1 |
S2 |
S3 |
|---|---|---|---|
Indole |
Negative |
Negative |
Negative |
Methyl Red |
Negative |
Negative |
Negative |
Voges Proskauer’s |
Negative |
Negative |
Positive |
Citrate |
Positive |
Positive |
Positive |
Sucrose |
Positive |
Positive |
Negative |
Glucose |
Positive |
Positive |
Positive |
Lactose |
Negative |
Negative |
Negative |
Mannitol |
Negative |
Negative |
Negative |
Sorbitol |
Negative |
Negative |
Negative |
Rhamnose |
Negative |
Negative |
Negative |
Adonitol |
Negative |
Negative |
Positive |
Arabinose |
Negative |
Negative |
Positive |
Molecular identification method
The 16S rRNA gene sequencing identified the bacterial isolates, S1, S2, and S3 as Stenotrophomonas maltophilia, Stenotrophomonas rhizophila, and Kocuria rhizophila. The identified sequences of both the isolates were submitted to the NCBI nucleotide database which were accepted and available under GenBank accession numbers OQ220315, OQ220316, and OQ220317 against Stenotrophomonas maltophilia, Stenotrophomonas rhizophila, and Kocuria rhizophila, respectively.
The results after the in vitro pathogenicity test confirmed the bacterial contamination in healthy larvae, thereby providing evidence for S. maltophilia, S. rhizophila, and K. rhizophila as flacherie-causing agents in S. ricini. After exposure to the bacterial suspension, silkworms showed reduced activity, appetite, clasping power, and movement within 72-100 hours. Notably, larvae injected with bacterial suspensions began to vomit and excrete semi-solid, chain-like faeces with a foul odour within 72 hours. The mortality rate varied among the different bacterial treatments, with silkworms treated with K. rhizophila succumbing to infection within 5-6 days, while those treated with S. maltophilia and S. rhizophila died within 6-7 days. As the infection progressed, the silkworms’ bodies underwent notable changes, including a foul odour, a brownish-black discoloration, and softening of the body. Post-mortem examination of the cadavers revealed liquefaction of internal organs, a hallmark of flacherie disease. Furthermore, the hemolymph collected from the treated larvae turned brownish-black, mirroring the symptoms observed in flacherie-infected silkworms collected from the field. These findings suggest that the bacterial isolates used in the study are capable of inducing flacherie-like symptoms in silkworms, consistent with the disease’s characteristic pathology.
The study identified bacterial colonies isolated from flacherie-infected eri silkworms, and all the colonies showed unique characteristics and gave analogous evidence of Stenotrophomonas maltophilia, Stenotrophomonas rhizophila, and Kocuria rhizophila based on colony characteristics, biochemical tests, and multiple sequence alignment. S. maltophilia and S. rhizophila hold substantial applications in agricultural biotechnology.27-30 These two species are always associated and persist as endosymbionts or endophytes31-33 with various plant species such as wheat plants, potatoes, tomatoes, cotton, and sweet peppers.28,30,34,35 Both play major roles such as promoting plant growth, helping in biocontrol, acting as a stress-protecting agent, protecting from draught and salinity, producing osmoprotective substances, degrading lignocellulose and xenobiotics, etc.30,36-39 Despite their agricultural benefits, S. maltophilia has been recognized as a nosocomial pathogen since the 1980s.40 Its close genetic relationship with S. rhizophila raises concerns about the potential pathogenic risks associated with agricultural use.30 However, S. rhizophila has not demonstrated pathogenicity in humans, being incapable of growth at human body temperatures.37 Nonetheless, its ability to form biofilms raises apprehensions about lateral gene transfer, potentially converting it into a pathogenic entity.41
Presently, S. maltophilia is deemed an emerging opportunistic pathogen due to its multidrug-resistance, high morbidity, and mortality rates among immunocompromised individuals due to its association with multidrug-resistance.18,42 The World Health Organization (WHO) has labelled it as an “underestimated important multidrug-resistant organism”.43,44 With an ecological presence in soil, water, hospital equipment, and soda lakes,40,45,46 S. maltophilia can colonize healthcare settings, causing outbreaks through contaminated water systems and appliances.18 In humans, S. maltophilia is associated with bacteremia, respiratory infections, and hematological malignancies.47-49 Its pathogenicity extends to plants, where it causes crippling white stripe disease in Oryza sativa,50,51 fruit soft rot in Lilium davidii,52 seedling soft rot in Zea mays,53 root rot in Panax notoginseng,54 and also responsible for the Persian oak (Quercus brantii) decline in Zagros forests, Iran.55 Such infections highlight its broad host range and ecological adaptability. Conversely, S. rhizophila is primarily recognized for its agricultural benefits, including plant growth promotion and phytopathogen inhibition.30,56-58 It colonizes plant roots as a non-pathogenic endophyte,33 presenting opportunities for sustainable agriculture. However, the mechanisms of its action remain largely unknown.
Similarly, the genus Kocuria, often overlooked as a common laboratory and specimen contaminant. K. rhizophila, another species within the genus, was previously reported as non-pathogenic to humans.59 Comprehensive studies on its interactions with other organisms are scarce. In Salmo trutta and Oncorhynchus mykiss, this species was responsible for developing exophthalmic symptoms, skin petechiae, increased skin melanization, focal lesions, intestinal inflammation, tail muscle hemorrhages, and liver congestion.14,60 K. rhizophila is a common commensal found in mammalian skin, mucous membranes, oropharynx, and in chicken meat treated with oxalic acid.14,61,62 It is also present in contaminated dust, fresh air, soil, marine sediments, freshwater, and various foods.14,62 Research into its occurrence as a contaminant in other organisms has been undervalued, as its accurate identification and risk assessment remain elusive.59 This species was frequently misidentified as coagulase-negative staphylococci (CoNS) because both share similar biochemical characteristics, such as a positive Gram test, a positive catalase reaction, and coagulase-negative properties.59,62 Furthermore, many clinical microbiology laboratories struggle to accurately identify the species due to diagnostic challenges and the lack of access to advanced molecular techniques.59
The interplay between the course of contamination and physiological changes due to pathogenic invasion in S. ricini is a field where no detailed studies have been conducted. The possibility that this type of contamination may increase the likelihood of colonization by specific pathogenic agents on the host’s body warrants examination due to its hazardous side effects. Bacterial flacherie, a common disease in the ericulture sites of India, especially in Assam, lacks comprehensive information regarding causative agents and pathogenic mechanisms. A study performed on ericulture in Papumpare district of Arunachal Pradesh, India only provides preliminary information on the bacterial types, coccus and bacillus, isolated from the hemolymph of contaminated S. ricini larvae.63 This aligns with the present findings, which elucidated the presence of two bacillus types (S. maltophilia and S. rhizophila) and one coccus type (K. rhizophila) of bacteria. No detailed reports of contamination by these three species have ever been recorded in S. ricini. Additionally, no data representing the sequence or strain of S. maltophilia, S. rhizophila, and K. rhizophila isolated from infected S. ricini is available in GenBank. This study, however, also addresses the gap in the molecular database for S. maltophilia, S. rhizophila, and K. rhizophila strains obtained from infected S. ricini larvae, thereby supplementing the knowledge concerning the pathogenic properties of these bacteria for S. ricini. The extensive host range of these species, from soil to skin, leaf to meat, and air to water, favours easy transmission to rearing beds while handling utensils, cleaning, leaf supply, and verbal communication. Moreover, being poikilothermic and immunologically sensitive, any changes in abiotic conditions such as temperature, relative humidity, dampness, and heavy rainfall create favorable conditions for the survival of various pathogenic microorganisms in the eri-rearing areas. Notably, sample collection for this study revealed that most eri silkworm rearers lacked awareness of modern tools and scientific rearing methods. Consequently, they often conducted ericulture in unhygienic and chaotic conditions. Furthermore, the seasonal scarcity of suitable host plants and limited plantation sites forced rearers to rely on decayed, waterlogged, and senescent foliage, and various host plants within a single generation. These factors significantly increase the risk of microbial infestation, creating an environment conducive to the proliferation of bacteria that can exert pathogenic effects on eri silkworm larvae. Interestingly, in the in vitro pathogenicity test, all three bacteria were found to exert pathogenic effects on the fifth instar larvae, and the symptoms were found consistent with the characteristic features of flacherie observed in the field study. Thus, the present study revealed S. maltophilia, S. rhizophila, and K. rhizophila as key contaminants of bacterial flacherie in S. ricini, filling the gap for detailed experimental work on causative agents of bacterial flacherie in S. ricini and highlighting the need for stringent rearing practices and regular microbial monitoring.
The present study unveils the role of S. maltophilia, S. rhizophila, and K. rhizophila in eri silkworm infestation and exposes the microbial menace threatening the sericulture industry. By tracing the pathogenic trajectory of these contaminants, the current investigation reveals a startling correlation between their presence and the degradation of S. ricini grainage parameters. This research fills a critical knowledge gap, providing a nuanced understanding of the complex interplay between microbial contaminants and their hosts. As the study established S. maltophilia, S. rhizophila, and K. rhizophila as formidable pathogens of S. ricini, findings of the present study could lay the groundwork for targeted interventions to safeguard the health and productivity of eri silkworms, ultimately fortifying the resilience of the sericulture sector.
ACKNOWLEDGMENTS
The authors remain indebted to Guwahati Biotech Park, Amingaon, Guwahati, Assam, for providing laboratory facilities during the experiment. Authors are grateful to YAAZH XENOMICS, TICEL BIOPARK Phase-III, Coimbatore, Tamil Nadu, India, for molecular analysis of the bacterial isolates. The authors are also thankful to the Sophisticated Analytical Instrument Centre (SAIF), Institute of Advanced Study in Science and Technology, Guwahati, for providing a field emission scanning electron microscopy facility. Lastly, the authors are grateful to the eri-rearers of Nayekgaon Pt-I village for providing valuable information regarding flacherie in eri silkworms and helping during the collection of samples.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
DB conceptualized the study and applied the methodology. DB and RRK visualized and investigated the study. RRK performed sample collection and experiments. DB performed data curation. RRK wrote the manuscript. DB reviewed the manuscript. Both authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in this manuscript.
ETHICS STATEMENT
Not applicable.
- Functioning of Central Silk Board and performance of Indian Silk Industry. Central Silk Board, Ministry of Textiles, Government of India, Bangalore. 2022. https://csb.gov.in/sites/default/files/what_s_new/files/NoteOnSeriCSB-2022-23-2nd-Qtr-English.pdf
- Sakthivel S, Angaleswari C, Mahalingam PU. Isolation and identification of bacteria responsible for flacherie in silkworms. Adv Appl Sci Res. 2012;3(6):4066-4068.
- Jiang L, Zhao P, Cheng T, et al. A transgenic animal with antiviral properties that might inhibit multiple stages of infection. Antiviral Res. 2013;98(2):171-173.
Crossref - Selvakumar T, Datta KR. Role of bacteria (Streptococcus faecalis and Staphylococcus aureus), viruses, and their combined infection in causation of flacherie under different environmental conditions. Sericologia. 2013;53(1):54-62
- Anantha Selvi GS, Chelladurai D, Pattu Kumar V, Navin Chandran M. Bacterial Flacherie in Bombyx mori (L.):Isolation and Identification of Pathogenic Organism. Uttar Pradesh Journal of Zoology. 2023;44(14):52-61.
Crossref - Sivaprasad V, Rahul K, Makwana P. Immunodiagnosis of silkworm diseases. Methods Microbiol. 2021;49:27-46.
Crossref - Zhang J, Shen ZY, Tang XD, Xu L, Zhu F. Isolation and identification of a pathogen, Providencia rettgeri, in Bombyx mori. Afr J Bacteriol Res. 2013;5(2):22-28.
Crossref - Chairman K, Singh AR, Amalarani G, Padmalatha C, Alagumuthu G. Effect of marine extracts on the microbial pathogens causing flacherie in the mulberry silkworm, Bombyx mori L. Asian Pac J Trop Biomed. 2012;2(3):1858-1861.
Crossref - Li GN, Xia XJ, Zhao HH, Sendegeya P, Zhu Y. Identification and Characterization of Bacillus cereus SW7-1 in Bombyx mori (Lepidoptera: Bombycidae). J Insect Sci. 2015;15(1):136.
Crossref - Haloi K, Kalita MK, Nath R, Devi D. Characterization and pathogenicity assessment of gut-associated microbes of silkworm Antheraea assamensis Helfer (Lepidoptera:Saturniidae). J Invertebr Pathol. 2016;138:73-85.
Crossref - Rahul K, Moamongba, Saikia K, Rabha M, Sivaprasad V. Identification and characterization of bacteria causing flacherie in mulberry silkworm, Bombyx mori L. J Crop Weed. 2019;15(3):178-181.
Crossref - Unban K, Klongklaew A, Kodchasee P, Pamueangmun P, Shetty K, Khanongnuch C. Enterococci as dominant xylose utilizing lactic acid bacteria in eri silkworm midgut and the potential use of Enterococcus hirae as probiotic for Ericulture. Insects. 2022;13(2):136.
Crossref - Gudimalla P, Akula S, Sujatha K. Isolation, identification, and characterization of pathogenic bacteria from the gut tissues of silkworm (Bombyx mori, L.) and its management using phytoessential oils. Int J Entomol Res. 2020;5(2):67-69.
- Pekala-Safinska A, Pazdzior E, Antychowicz J, et al. Kocuria rhizophila and Micrococcus luteus as emerging opportunist pathogens in brown trout (Salmo trutta Linnaeus, 1758) and rainbow trout (Oncorhynchus mykiss Walbaum, 1792). Aquaculture. 2018; 486:285-289.
Crossref - Juhnke ME, Jardin ED. Selective medium for isolation of Xanthomonas maltophilia from soil and rhizosphere environments. Appl Environ Microbiol. 1989;55(3):747-750.
Crossref - Brooke SJ. Stenotrophomonas maltophilia: an emerging global opportunistic pathogen. Clin Microbiol Rev. 2012;25(1):2-41.
Crossref - Cappuccino JG, Sherman N. Microbiology: A Laboratory Manual, 10th Ed. Pearson. 2014.
- Mahdi O, Eklund B, Fisher N. Laboratory Culture and Maintenance of Stenotrophomonas maltophilia. Curr Protoc Microbiol. 2014;32(1).
Crossref - Ghosh R, Chatterjee S, Mandal NC. Stenotrophomonas. In Book: Beneficial Microbes in Agro-Ecology. 2020; 427–442.
Crossref - Sherpa TM, Das S, Najar NI, Thakur N. Draft genome sequence of Stenotrophomonas maltophilia strain P13 gives insight into its protease production and assessment of sulfur and nitrogen metabolism. Curr Res Microb Sci. 2021;2:100012.
Crossref - Raio A, Brilli F, Neri L, et al. Stenotrophomonas rhizophila Ep 2.2 inhibits growth of Botrytis cinerea through the emission of volatile organic compounds, restricts leaf infection and primes defense genes. Front Plant Sci. 2023;14:1235669.
Crossref - Ranganathan K, Rathinam K, Danial M, Subramaniam S. Quick isolation and characterization of novel Bacillus thuringiensis strains from mosquito breeding sites in Malaysia. Emir J Food Agric. 2011;23(1):17-26.
Crossref - Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792-1797.
Crossref - Talavera G, Castresana J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst Biol. 2007;56(4):564-577.
Crossref - Alavi P, Starcher MR, Thallinger GG, Zachow C, Muller H, Berg G. Stenotrophomonas comparative genomics reveals genes and functions that differentiate beneficial and pathogenic bacteria. BMC Genomics. 2014;15(1):482.
Crossref - Sahin N, Ay H, Saygin H. Kocuria. Bergey’s Manual of Systematics of Archaea and Bacteria, Bergey’s Manual Trust. 2021.
Crossref - Kumar A, Rithesh L, Kumar V, et al. Stenotrophomonas in diversified cropping systems:friend or foe? Front Microbiol. 2023;3(14).
Crossref - Wolf A, Fritze A, Hagemann M, Berg G. Stenotrophomonas rhizophila sp. nov, a novel plant-associated bacterium with antifungal properties. Int J Syst Evol Microbiol. 2002;52(6):1937–1944.
Crossref - Kai M, Effmert U, Berg G, Piechulla B. Volatiles of bacterial antagonists inhibit mycelial growth of the plant pathogen Rhizoctonia solani. Arch Microbiol. 2007;187(5):351-360.
Crossref - Schmidt CS, Alavi M, Cardinale M, Muller H, Berg G. Stenotrophomonas rhizophila DSM14405T promotes plant growth probably by altering fungal communities in the rhizosphere. Biol Fertil Soils. 2012;48(8):947-960.
Crossref - Taghavi S, Garafola C, Monchy S, et al. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl Environ Microbiol. 2009;75(3):748-757.
Crossref - Zhu B, Liu H, Tian WX, et al. Genome sequence of Stenotrophomonas RR-10, isolated as an endophyte from rice root. J Bacteriol. 2012;194(5):1280-1281.
Crossref - Berg G, Martinez JL. Friends or foes: Can we make a distinction between beneficial and harmful strains of the Stenotrophomonas maltophilia complex? Front Microbiol. 2015;6:241.
Crossref - Singh RP, Jha PN. The PGPR Stenotrophomonas maltophilia SBP-9 Augments Resistance against Biotic and Abiotic Stress in Wheat Plants. Front Microbiol. 2017;8:1945.
Crossref - Messiha NAS, van Diepeningen AD, Farag NS, Abdallah SA, Janse JD, van Bruggen AHC. Stenotrophomonas maltophilia: a new potential biocontrol agent of Ralstonia solanacearum, causal agent of potato brown rot. Eur J Plant Pathol.2007;118(3):211–225.
Crossref - Ryan PR, Monchy S, Cardinale M, et al. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat Rev Microbiol. 2009;7:514–525.
Crossref - Berg G, Egamberdieva D, Lugtenberg B, Hagemann M. Symbiotic Plant–Microbe Interactions: Stress Protection, Plant Growth Promotion, and Biocontrol by Stenotrophomonas. In: Seckbach J, Grube M. (eds) Symbioses and Stress. Cellular Origin, Life in Extreme Habitats and Astrobiology, vol 17. Springer, Dordrecht.
Crossref - Alavi P, Starcher M, Zachow CS, Berg G, Muller H. Root-microbe systems: The effect and mode of interaction of Stress Protecting Agent (SPA) Stenotrophomonas rhizophila DSM14405T. Front Plant Sci. 2013;14(4):141.
Crossref - An SQ, Berg G. Stenotrophomonas maltophilia. Trends Microbiol. 2018;26(7):637-638.
Crossref - Pinski A, Zur J, Hasterok R, Hupert-Kocurek K. Comparative Genomics of Stenotrophomonas maltophilia and Stenotrophomonas rhizophila Revealed Characteristic Features of Both Species. Int J Mol Sci. 2020;21(14):4922.
Crossref - Ren D, Madsen JS, Sorensen SJ, Burmolle M. High Prevalence of Biofilm Synergy among Bacterial Soil Isolates in Cocultures Indicates Bacterial Interspecific Cooperation. ISME J. 2015;9(1):81-89.
Crossref - Said MS, Tirthani E, Lesho E. Stenotrophomonas maltophilia, StatPearls Publishing. 2023.
- Brooke JS. New strategies against Stenotrophomonas maltophilia: a serious worldwide intrinsically drug-resistant opportunistic pathogen. Expert Rev Anti Infect Ther. 2014;12(1):1-4.
Crossref - Willsey GG, Eckstrom K, LaBauve AE, et al. Stenotrophomonas maltophilia Differential Gene Expression in Synthetic Cystic Fibrosis Sputum Reveals Shared and Cystic Fibrosis Strain-Specific Responses to the Sputum Environment. J Bacteriol. 2019;201(15):e00074-19.
Crossref - Hauben L, Vauterin L, Moore ERB, Hoste B, Swings J. Genomic diversity of the genus Stenotrophomonas. Int J Syst Evol Microbiol. 1999;49(4):1749-1760.
Crossref - Wisplinghoff H, Seifert H. Pseudomonas spp., Acinetobacter spp. and miscellaneous Gram-negative bacilli. Infect Dis; 2010;2:1704-1727.
Crossref - Abbott IJ, Slavin MA, Turnidge JD, Thursky KA, Worth LJ. Stenotrophomonas maltophilia: emerging disease patterns and challenges for treatment. Expert Rev Anti Infect Ther. 2011;9(4):471-488.
Crossref - Kwa AL, Low JGH, Lim TP, Leow PC, Kurup A, Tam VH. Independent predictors for mortality in patients with positive Stenotrophomonas maltophilia cultures. Ann Acad Med Singap. 2008;37(10):826-830
- Paez JIG, Tengan FM, Barone AA, Levin AS, Costa SF. Factors associated with mortality in patients with bloodstream infection and pneumonia due to Stenotrophomonas maltophilia. Eur J Clin Microbiol Infect Dis. 2008;27(10):901-906.
Crossref - Singh IN, Swings IRJ, Devi TKR, Imotomba RK. White stripe, a new disease of rice caused by Stenotrophomonas maltophilia in India. Indian Phytopathol. 2001;54 (2):275-277
- Azmat S, Riaz K, Dilferoze A, et al. Morphogenetic characterization of Stenotrophomonas maltophilia infecting white stripe disease of rice (Oryza sativa L.). Not Bot Horti Agrobo. 2024;52(2):1347.
Crossref - LingL, Jiao Z, Wenxia M, et al. Preliminary report on the study of postharvest fruit rot bacteria and yeasts in Lanzhou Lily (Lilium davidii var. unicolor) in China, J Phytopathol. 2018;167(3):135-145.
Crossref - Heidari M, Aeini M. Stenotrophomonas maltophilia causing maize (Zea mays L.) seedling soft rot disease, an emerging threat to maize crop. J Phytopathol. 2024;172(2)e13299.
Crossref - Zheng KY, Su X, Zheng X, et al. First report of Stenotrophomonas maltophilia causing root soft rot of Sanqi (Panax notoginseng) in China. Plant Dis. 2022;106(2):755.
Crossref - Ahmadi E, Kowsari M, Azadfar D, Jouzani SG. Bacillus pumilus and Stenotrophomonas maltophilia as two potentially causative agents involved in Persian oak decline in Zagros forests (Iran). For Pathol. 2019;49(5):e12541.
Crossref - Maurer KA, Zachow C, Seefelder S, Berg G. Initial steps towards biocontrol in hops: Successful colonization and plant growth promotion by four bacterial biocontrol agents. Agronomy. 2013;3(4):583-594.
Crossref - Rivas-Garcia T, Murillo-Amador B, Nieto-Garibay A, Rincon-Enriquez G, Chiquito-Contreras RG, Hernandez-Montiel LG. Enhanced biocontrol of fruit rot on muskmelon by combination treatment with marine Debaryomyces hansenii and Stenotrophomonas rhizophila and their potential modes of action. Postharvest Biol Technol. 2019;151:61-67.
Crossref - Reyes-Perez JJ, Hernandez-Montiel LG, VeroS, Noa-Carrazana JC, Quinones-Aguilar EE, Rincon-Enriquez G. Postharvest biocontrol of Colletotrichum gloeosporioides on mango using the marine bacterium Stenotrophomonas rhizophila and its possible mechanisms of action. J Food Sci Technol. 2007;56(11):4992–4999.
Crossref - Kandi V, Palange P, Vaish R, et al. Emerging bacterial infection: identification and clinical significance of Kocuria Species. Cureus. 2016;8(8):1-6.
Crossref - Hajam YA, Kumar R, Rani R, Sharma P. Efficacy of different treatments available against bacterial pathogens in fish. In: Dar OI, ed. Bacterial Fish Diseases: Environmental and Economic Constraints. Amsterdam, Netherlands: Academic Press/Elsevier; 2022:379-398.
Crossref - Stackebrandt E, Koch C, Gvozdiak O, Schumann P. Taxonomic dissection of the genus Micrococcus: Kocuria gen. nov, Nesterenkonia gen. nov, Kytococcus gen. nov, Dermacoccus gen. nov, and Micrococcus Cohn 1872 gen. emend. Int J Syst Evol Microbiol. 1995;45(4):682-692.
Crossref - Savini V, Catavitello C, Masciarelli G, et al. Drug sensitivity and clinical impact of members of the genus Kocuria. J Med Microbiol. 2010;59(12):1395-1402.
Crossref - Meth T, Gogoi H. Rearing of Eri Silkworm (Samia Cynthia ricini Boisd.) (Lepidoptera: Saturniidae) In Arunachal Pradesh: A Study in Papumpare District.
J Bioresourc. 2016;3(1):46-52.
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.