


In the recent past, multidrug-resistant Staphylococcus aureus (MDR S. aureus) and especially methicillin-resistant Staphylococcus aureus (MRSA) has been a major health problem affecting population worldwide. The MDR S. aureus burden has continued to be reported in healthcare and community environments between 2015 and 2025 and is becoming more resistant to a variety of antibiotic groups that complicate treatment and control. This review will summarize the epidemiological patterns of the world, local inequalities, molecular pathways of resistance, clinical and population health outcomes, and new treatment options regarding MDR S. aureus. Surveillance data indicates that MRSA has been observed to show a high prevalence of hospital-associated infections with a continued increase in community transmission, as well as a high heterogeneity between low- and middle-income and high-income countries. The appearance and spread of dominant clonal lineages, which are propelled by mobile genetic elements and horizontal gene transfer, have led to resistance beyond β-lactams and also the macrolide, fluoroquinolones, aminoglycosides, and tetracyclines. MDR S. aureus infections are related to treatment failure, extended hospitalization, morbidity, and mortality, as well as significant economic impact. The existing treatment strategies are limited by the resistance of antibiotics, a limited antimicrobial development flow, and the problems of diagnosis and the use of antibiotics in an abused manner. The promising alternatives are anti-virulence therapies, bacteriophage therapy, CRISPR-based antimicrobials, antimicrobial peptides, microbiome-based therapy, and artificial intelligence-based drug discovery. In order to curb the increasing threat of MDR-S. aureus, strengthening antimicrobial surveillance, infection control measures and stewardship programs and coordinated global research and policy efforts is necessary.
Staphylococcus aureus, MRSA, Multidrug-resistance, Antimicrobial Resistance, Molecular Mechanisms, Global Epidemiology, Novel Therapeutics
Staphylococcus aureus (S. aureus) is a Gram-positive bacterium that is a widespread part of the human microbiota. It is commonly colonized by many healthy individuals on their skin, in their nose, or elsewhere on their body, but it still has the capacity to be a highly dangerous opportunistic pathogen.1 It may lead to an entire continuum of diseases, such as superficial skin and soft tissue infections (SSTIs) and invasive and life-threatening diseases such as bacteremia, endocarditis, pneumonia, and sepsis.1,2 Historically, antibiotics had a tremendous impact on clinical outcome of staphylococcal infections as penicillin was initially discovered in early 1940s, it was remarkably effective against S. aureus, but within a short period it became penicillin-resistant, because of β-lactamase production that is encoded by blaZ gene.3,4
By overcoming penicillin-resistant Staphylococcus aureus, the semisynthetic and penicillinase stable β-lactam antibiotics like methicillin were developed and introduced into clinical practice in 1959.5 Methicillin-resistance in Staphylococcus aureus was first reported in 1961, within one year of methicillin introduction in 1960, highlighting the rapid adaptability of the organism. MRSA contains a mecA gene on a mobile genetic element staphylococcal cassette chromosome mec (SCCmec).5,6 This gene carries an alternative penicillin-binding protein, PBP2a, with an extremely lower affinity towards β-lactam antibiotics, and it provides resistance to almost all the agents in the β-lactam family.2,6
MRSA which used to be regarded as a hospital acquired (nosocomial) pathogen disseminated worldwide and diversified. What was formerly mostly restricted to clinical settings has now extended into community areas, altering the epidemiology of S. aureus infections.2,7 Concurrently, the epidemiology of S. aureus infections has been shifted with increased resistance to other classes of antibiotics, increasing the potential risk of multidrug-resistant (MDR) strains, complicating treatment, and reducing the therapeutic options. The high level of plasticity of S. aureus through horizontal gene transfer, mobile genetic elements as well as mutation highlights the current problem of controlling its distribution and treatment of the infection.2,8
Considering such a dynamic development of antimicrobial resistance in S. aureus, it is especially timely to conduct a systematic review of studies published from 2015-2025. In the last ten years, the resistance landscape has changed due to improvements in molecular surveillance (e.g., whole genome sequencing), increased global data, even low- and middle-income countries, and changes in usage patterns of antibiotics. It is our hope that this review helps to synthesize the trends in epidemiology around the world, dissect the molecular processes of resistance, assess the clinical and public health outcomes, and comment on the new therapeutic approaches and directions in developing countries in fighting MDR S. aureus.
Global Epidemiology and Trends (2015-2025)
Prevalence and incidence: hospital vs community settings
One of the most widespread causes of skin and soft tissue infection (SSTI), bloodstream infection (BSI), pneumonia, and device-associated infection (DAI) globally is Staphylococcus aureus, which has a prevalence rate of 14.7% according to a global meta-analysis of 119 studies involving 164,717 patients in 29 countries.9,10 Selective antibiotic pressure, length of hospitalization, invasive procedures, and cross-transmission between patients and healthcare workers continue to contribute to the prevalence of hospital-acquired MRSA.11
Among community members, the burden of MRSA is moderate but considerable. Carriage of MRSA across the globe among children is 3.4%, and in adults with HIV, prevalence is nearly 7%.12,13 The clones of community-acquired MRSA (CA-MRSA), which is frequently linked to the presence of virulence factors, including Panton Valentine leukocidin (PVL), still leads to the SSTIs and outbreaks in households, sports teams, and residential communities.14 The emergence of CA-MRSA obscures the historic view of hospital and community strains, and with several studies reporting both directions of transmission.15
Regional differences: high-income vs low- and middle-income countries
Global MRSA epidemiology is characterized by marked geographic heterogeneity. Over the last ten years High-income countries (HICs) have reported a decline or stabilization in invasive MRSA infections with better antimicrobial stewardship, enhanced screening of patients and more effective infection prevention and control measures.10,16 A recent report from the European Center for Disease Prevention and Control (ECDC) reports, decreased rates of MRSA bloodstream infections in Western Europe.16
On the other hand, low- and middle-income countries (LMICs) have an inordinately heavy burden of disease. MRSA pooled prevalence for community residents in Africa was reported at about 4.1% compared with hospital patients and healthcare workers prevalence of about 13.6%.17 India has also shown a relatively consistent high prevalence of MRSA (30%-45%) among clinical isolates from tertiary-care hospitals due to factors such as overcrowding, inadequate isolation areas and poor compliance with infection control practices and empiric antibiotic therapy.18
The growing incidence of multidrug- resistant (MDR) Staphylococcus aureus strains has been reported across South-East Asia and China, however, MDR S. aureus resistance now extends well beyond β-lactam antibiotics to include macrolide antibiotics, fluoroquinolone antibiotics, tetracycline antibiotics and occasionally linezolid antibiotics.19,20 Additionally, the high prevalence of livestock-associated methicillin-resistant S. aureus (LA-MRSA) is found in countries or regions that are involved in large scale commercial pork and poultry farming.21
Temporal trends in MDR S. aureus and MRSA (2015-2025)
In terms of a global decrease in MRSA during the period of 2015-2025 there is no overall decrease in MRSA according to global surveillance data, however, there is an alarming increase in MDR strains. According to the 2024 Global Research on Antimicrobial Resistance (GRAM) Report on antimicrobial resistance research, deaths attributed to MRSA (i.e., directly caused by the infection) have almost doubled since 1990, when they were 57,000, to approximately 130,000 in 2021. In contrast, deaths associated with MRSA (where MRSA is a contributing factor but not the sole cause) have increased from approximately 261,000-550,000.15,19 Despite improvements in infection control in certain regions, these rising figures indicate a growing global clinical burden of MRSA.
Rates of invasive MRSA are stable or declining in some European nations, while they continue to rise or remain elevated across areas in Asia, Africa, and Latin America.16 The majority of low- and middle-income Countries (LMIC) do not have standardized infection surveillance systems. This leads to underreporting of disease incidence as well as limited ability to track longitudinal trends.17,18 Studies conducted in China from 2018 through 2024 demonstrate a significant increase in antibiotic resistance of MRSA isolates to clindamycin, erythromycin, and fluoroquinolones. A number of reports have demonstrated an increase in multidrug-resistant S. aureus infections in patients with SSTIs, BSIs and ventilator-associated pneumonia in tertiary hospitals in India and Southeast Asia.18,20
The rise of CA-MRSA clones, with their increased carriage of macrolide and fluoroquinolone resistance determinants, has made it difficult for empirically treated infections to be effectively treated. In addition, the emergence of these resistant strains has complicated the treatment of both SSTI’s and acute emergencies.20,21
Clonal Lineages and Their Global Spread
The historically prevalent ST22 (EMRSA-15) continues to be the most prevalent clone in many European countries; however, in a few areas, the incidence is reportedly declining as a result of the implementation of robust infection control measures.16 The predominant clones identified in China are ST59-t437 (typically community-associated, PVL positive), and there is an increasing number of hospital-based isolates of ST22-t223.19 In Thailand and Southeast Asia, the ST22-MRSA-IV-t032 lineage has emerged as the predominant clone in tertiary care facilities.20 Whole-genome sequencing (WGS) studies have dramatically changed our understanding of how MRSA clonal populations evolve. Notable global shifts in MRSA clonal evolution during the years 2015-2025 include the emergence of new or expanding lineages such as ST22, ST59, ST8 and ST5.19-22
The livestock-associated strain of MRSA (LA-MRSA) has continued to expand internationally, and zoonotic transmission is increasing in regions such as Europe, China, and South America.21 WGS data indicates there is an ongoing exchange of LA-MRSA among humans and animals. Therefore, One Health surveillance is critical.
Concerns for emerging highly virulent strains that possess both resistance and virulence factors are present. For example, recent WGS studies have shown a growing trend of dual-resistant, PVL-positive ST59 and ST8 strains that contain resistance to macrolides and fluoroquinolones.22
Synthesis: key patterns, gaps, and challenges
There is a persistent global presence of MRSA in hospitals, long-term care facilities, communities, and livestock.9,10,22 The fact that there are people carrying MRSA in the elderly care facilities demonstrates the need to implement control measures at the level of the community.11 In many low-to-middle-income countries (LMICs), the prevalence of MRSA is much higher than in high-income countries (HICs), and in addition, they have more severe MDR phenotypes, primarily due to overuse of antibiotics, lack of surveillance systems, and limited resources available.17,18
Between 2015 and 2025, all the temporal trends show an increase in the number of MDR phenotypes; therefore, there is no decline in the burden of MRSA on a global basis.15,19 The clonal evolution of MRSA continues to evolve with the development of new strains of MDR MRSA such as ST22 and ST59, as well as the emergence of hypervirulent MDR clones and an increase in the zoonotic transmission of LA-MRSA.19-22 There is a significant amount of fragmentation in global surveillance of MRSA, especially in rural areas and in LMICs, which limits the ability to determine the true burden of MRSA and slow down international coordination. Therefore, it is important to enhance genomic surveillance of MRSA and to standardize the reporting of antimicrobial resistance (AMR).
Molecular Mechanisms of Resistance (2015-2025)
Understanding the molecular mechanism of antimicrobial resistance in Staphylococcus aureus is a crucial component of the interpretation of the epidemiology of this organism as well as prediction of outcomes of therapy for infections caused by it. Since 2015 through 2025, whole-genome sequencing (WGS), and functional genomics have provided considerable insight into the Staphylococcus aureus resistome, illustrating that there are numerous interactions between chromosomal mutation, mobile genetic elements, and the selective pressure for evolution that lead to the continued proliferation of multidrug-resistant (MDR) clones within all three environments of healthcare, community, and livestock.23
β-Lactam Resistance (mecA, mecC, PBP2a)
Methicillin-resistance in Staphylococcus aureus (MRSA) is caused largely by an organism acquiring a copy of the mecA gene. The mecA gene produces a low-affinity penicillin-binding protein called PBP2a.24 This permits peptidoglycan to be synthesized regardless of whether the organism is exposed to beta-lactam antibiotics; as such, the organism fails to respond to antibiotic treatment and persists within host tissue. Expression of mecA is controlled by mecI-mecR1 system that can induce mecA expression on exposure to beta-lactams. Some strains have genetic mutations or deletions of either mecI or mecR1, causing constitutive expression of mecA and therefore greater levels of resistance to methicillin.24,25
A small number of MRSA isolates contain mecC, a mecA equivalent that was identified in 2011 and has since been observed in various parts of Europe and Asia, as well as in livestock reservoirs. The mecC-MRSA strain contains a variant of PBP2a with decreased affinity for cefoxitin; this makes it difficult to detect through standard phenotyping methods.26 In addition, whole genome sequencing studies have shown an increased level of mecC carriage in wildlife-livestock interfaces, further emphasizing the need for One Health surveillance.27
The ability of β-lactams to be resistant is contained within the staphylococcal cassette chromosome mec (SCCmec), a mobile genetic element that integrates at the orfX site.28 SCCmec types I-III, which are larger and more complex than other SCCmec types, are associated with hospital strains that frequently harbor additional resistance genes.29 The degree of variation among the different SCCmec types (I-XIII) will influence resistance patterns, fitness, and the spread of the bacteria. An example would be SCCmec type IV, which is smaller and thus more mobile than other SCCmecs and is responsible for the dominance of community-associated MRSA (CA-MRSA).29,30
Resistance to other antibiotic classes
Macrolides, Lincosamides, and Streptogramins (MLSB)
The most common mechanism for macrolide resistance is through erm genes (ermA, ermB, ermC) expression which encodes ribosomal methyltransferase enzymes which modify the 23S rRNA target site for macrolides, lincosamides and streptogramin antibiotics.30 The erm gene products can either be constitutively expressed or expressable upon induction (iMLSB) which would require a positive result with the D test to detect. Additionally, efflux pumps such as msrA have been shown to contribute to macrolide resistance, particularly in CA-MRSA lineages.31
Aminoglycosides
Aminoglycoside-modifying enzyme (AME) mediated resistance to aminoglycosides is caused by three types of enzymes: acetyltransferases (aac), nucleotidyltransferases (ant) and phosphotransferases (aph). The most clinically relevant aac(6′)-Ie-aph(2″)-Ia bimodular gene results in high level resista tetM nce to gentamicin and is typically found alongside multiple antibiotic resistance genes (ARGs) in plasmids.32
Ribosomal protection proteins (tetM, tetO) and efflux pumps (tetK) are the two main mechanisms that cause tetracycline resistance. tetK is commonly linked to plasmids in CA-MRSA strains, whereas tetM is the more prevalent mechanism of resistance in hospital-acquired (HA-MRSA) strains.33 As the use of tetracyclines as antibiotics in livestock increases, there will be increased transmission of MRSA strains (both tetM and tetK positive) from agricultural settings to humans.34
Fluoroquinolones
Quinolone resistance occurs when quinolones are unable to bind to the quinolone resistance-determining regions (QRDRs) of gyrA and/or grlA in bacteria because of mutations within those genes. gyrA encodes for DNA gyrase, whereas grIA encodes for topoisomerase IV. The most common mutations that occur due to exposure to a fluoroquinolone are serine-84-leucine in (gyrA), and serine-80-tyrosine or serine-80-phenylalanine in (grIA). These mutations reduce the affinity of the drug for its target.35 In addition to these mechanisms, horizontal gene transfer may also provide an additional mechanism of resistance through the acquisition of a plasmid-borne efflux pump, e.g., qacA/qacB. It is notable that co-selection of fluoroquinolone-resistant strains with β-lactamase -producing strains is likely to be enhanced in a hospital setting.36
Mobile Genetic Elements (MGEs): Plasmids, Transposons, SCCmec, and Phages
Staphylococcus aureus harbors many different types of mobile genetic elements (MGEs), which allow it to quickly adapt and share genes horizontally in order to survive on other bacteria, including plasmids, transposons, SCCmec elements, prophages, pathogenicity islands and insertional sequences. Many plasmids contain numerous antibiotic resistance genes (ARGs), including blaZ, ermC, qacA/B and amingolycoside modifying enzyme genes. Due to their ability to be transferred from one strain or species to another via conjugation, large conjugative plasmids have allowed for the widespread distribution of these genes among Staphylococcus aureus.28,32 With whole-genome sequencing studies increasingly express plasmid co-carriage of ARGs and virulence determinants, there are now hybrid virulence-resistance phenotypes emerging.34 Transposons (including Tn554, Tn916) enable the spread of resistance genes (such as ermA and tetM) in response to antimicrobial selection pressures.32-34
The MGE most clinically relevant to SCCmec continues to evolve through ccr-mediated recombination, new ccr-mec configurations have been identified between 2018 and 2025 indicating continued genetic diversity.29 Bacteriophages are generally known to contribute to the dissemination of virulence factors, however they can also serve as genetic reservoirs that enhance the bacterial’s ability to adapt to changing environments.37
Interaction between resistance and virulence
The interplay of virulence and antimicrobial resistance is now considered as a significant determinant of the pathogenesis of methicillin-resistant Staphylococcus aureus (MRSA). Historically, it has been believed that the process by which bacteria become resistant to antibiotics decreases their ability to cause disease. Contemporary MRSA strains such as ST22, ST59, and USA300 (ST8) have demonstrated both an increase in resistance to β-lactam and an increased ability to cause disease.34 Research utilizing whole genome sequencing has shown that different SCCmec types IV and V do not result in significant reductions in fitness (Section 2.1).29
Some plasmids and transposons harbor resistance genes (e.g., ermC, tetK) and virulence factors simultaneously, this enables these plasmids/transposons to be selected for at the same time when bacteria are exposed to antimicrobials. Additionally, global regulatory systems such as agr influence toxin production, biofilm formation, and tolerance to antibiotics, furthermore, dysfunction of agr can contribute to persistence phenotypes of MRSA (Table 1).37 Overall, the simultaneous development of resistance and virulence in successful clones of MRSA demonstrates the necessity of using genomic-based surveillance methods to identify emerging, high-risk lineages of MRSA.
Table (1):
Major Antimicrobial Resistance Mechanisms in Staphylococcus aureus
Antibiotic Class / Mechanism |
Key Resistance Genes |
Mechanistic Basis |
Associated MGEs |
Ref. |
|---|---|---|---|---|
β-lactams (methicillin, penicillins) |
mecA, mecC, blaZ |
PBP2a-mediated β-lactam insensitivity; β-lactamase production |
SCCmec I–XIII; plasmids carrying blaZ |
24,26-28 |
MLSB (macrolides, lincosamides, streptogramins) |
ermA, ermB, ermC, msrA |
Ribosomal methylation; efflux pump activity |
Plasmids; transposon Tn554 |
30,31 |
Aminoglycosides |
aac(6′)-Ie-aph(2″)-Ia, ant, aph |
Enzymatic drug modification (acetylation, adenylation, phosphorylation) |
Plasmids; AME-carrying transposons |
10 |
Tetracyclines |
tetK, tetM, tetO |
Efflux pump; ribosomal protection proteins |
Plasmids; Tn916 |
11 |
Fluoroquinolones |
gyrA, grlA mutations; qacA/B |
Target-site mutations; efflux-mediated tolerance |
Plasmids encoding efflux pumps |
35,36 |
Biocide / disinfectant resistance |
qacA, qacB |
Efflux of quaternary ammonium compounds |
Plasmids co-carrying ARGs |
36 |
Multidrug resistance via MGEs |
Mixed ARG clusters |
Co-location of resistance and virulence determinants |
Plasmids, prophages, SaPIs, SCCmec, IS elements |
32,34,37 |
Virulence–resistance interaction |
agr mutations; SCCmec IV/V |
Low-fitness-cost SCCmec; coupled virulence–resistance selection |
SCCmec; plasmids carrying toxins + ARGs |
29,34,37 |
Summary of major antimicrobial resistance mechanisms, associated genes, mobile genetic elements (MGEs), and key references
Clinical and Public Health Implications (2015-2025)
The clinical and public health burden of multidrug-resistant Staphylococcus aureus (MDR S. aureus) worldwide is due to a significant increase in treatment failures, morbidity, mortality, prolonged hospitalization, and healthcare costs associated with empirical antibiotic therapy for MDR strains.38 As a result of this, treatment options are often limited to use of last-line antibiotics or antibiotic combinations that have a greater toxicity profile or limited availability.39,40 Furthermore, severe infections such as bacteremia and pneumonia have shown to have a poor prognosis, a recent 2024 meta-analysis showed that patients suffering from MRSA bacteremia had a higher mortality rate than those suffering from MSSA bacteremia. Additionally, biofilm development, decreased susceptibility to vancomycin, and resistance-virulence co-selection will continue to prolong hospital stays.40,41
There is increasing complexity in the infection control of hospitals because of the co-circulating hospital-associated and community-associated MRSA lineages. Poor hand hygiene compliance, overcrowded wards, lack of trained infection-control workers, and the use of more invasive procedures become the facilitating factors of the continued spread in healthcare facilities, where the colonization rates are above 14% globally and create a continuous transfer of the MRSA-colonized individuals between the hospitals, nursing homes, and the community.42
There is also a rise in community-level transmission, especially by means of skin and soft tissue infections (SSTIs), household clusters and crowded environments.43 Selection pressures associated with the emergence of MDR-resistant strains such as ST8, ST22 and ST59 have increased the epidemiology of MRSA beyond the traditional health care setting due to poor antibiotic stewardship, including over-the-counter availability of drugs, inappropriate prescribing and self-treatment.44
The cost to the economy is high. The WHO AMR report of 2024 states that MRSA infections impose disproportionate costs in terms of AMR and estimated per-patient costs are 1.5-3 times more than those of MSSA infections because of the prolonged illness, disability, or death, leading to a loss of productivity in society, particularly in low- and middle-income countries (LMICs).44 Weaknesses in surveillance are a significant obstacle to contain. The LMICs do not have a well-organized reporting system, standard laboratory capacity, or a genomic surveillance system. In India, MRSA in sentinel sites has been found to be between 33-43 percent over 2015-23, according to the Indian Council of Medical Research (ICMR) AMR Surveillance Network, but the region is highly heterogeneous, making it difficult to estimate the prevalence of this bacterium.45 Surveillance in the region is widely heterogeneous, with few sentinel sites and most studies being small, making it challenging to estimate the prevalence of this bacterium (Table 2).44,45
Table (2):
Clinical and Public Health Implications of MDR-Staphylococcus aureus (2015-2025)
Domain |
Key Findings / Impact |
Ref. |
|---|---|---|
Clinical Outcomes |
Higher treatment failure; severe disease in bacteremia, pneumonia, device infections; MRSA bacteremia mortality OR ≈ 1.79 vs MSSA |
40,41 |
Therapeutic Challenges |
Need for last-line agents (linezolid, daptomycin, ceftaroline); higher toxicity and cost |
38,39 |
Hospital Burden |
Excess hospital stay 6–12 days; increased per-patient cost; ICU pressure |
38,40 |
Biofilm-Driven Persistence |
VISA/hVISA, biofilm formation, reduced vancomycin susceptibility cause persistent bacteremia |
41 |
Healthcare Transmission |
Co-circulation of HA- and CA-MRSA; poor hand hygiene; overcrowding; staffing deficits |
42 |
Long-Term Care Facilities |
High colonization (>14% globally); continuous movement between hospitals and LTCFs |
43 |
Community Transmission |
CA-MRSA spread via SSTIs, households, crowded areas; rise of ST8, ST22, ST59 |
43,44 |
Antibiotic Stewardship Gaps |
OTC antibiotic access, inappropriate prescriptions, self-medication |
44 |
Economic Burden |
MRSA cost 1.5–3× higher than MSSA; greater productivity loss in LMICs |
38,44 |
Surveillance Limitations |
India MRSA 33–43% (ICMR 2015–2023); limited WGS; fragmented LMIC data |
38,45 |
The under-resourced stewardship programs, slow diagnosis, inability to regulate the sale of antibiotics, and ineffective integration of hospital and community surveillance systems continue to pose a challenge to the efforts of public health to respond. Any improvements in the infrastructure of the diagnostic system, the increase in genomic surveillance, the self-protecting character of infection control requirements, and the obligatory observance of the requirements of stewardship are essential to decrease the burden of MDR-S. aureus.
Clinical, therapeutic, and public health impact of multidrug-resistant Staphylococcus aureus (MRSA) infections, emphasizing increased treatment failure, mortality, healthcare burden, transmission, and gaps in stewardship and surveillance.
Therapeutic challenges and limitations of current antibiotics
Treatment of Staphylococcus aureus (specifically MRSA) continues to be one of the major problems facing global health today because of the decline in antibiotic effectiveness, slow pace of new drug development, the gap in diagnostics, and improper use of antimicrobials.46 Over the last decade there has been an increase in the number of resistant strains to β-lactams antibiotics, macrolides, aminoglycoside antibiotics, fluoroquinolone antibiotics, and polymyxin antibiotics, as a result the number of viable therapeutic options available to treat these infections has dramatically decreased.47
Declining efficacy of standard antibiotics
The front-line antibiotic treatments are no longer effective for the majority of people with bacterial infections. β-lactams are ineffective against MRSA due to mecA or mecC-mediated alterations in penicillin-binding proteins and beta-lactamase production.48,49 Carbapenem-resistance rates have exceeded 40% in low-middle-income countries (LMICs), and MRSA has remained prevalent at greater than 30% in these regions since the 2015-2025 surveillance.50 Resistance to other antibiotics (clindamycin, fluoroquinolone, tetracycline, etc.) has become so widespread that even the last line of treatment options (vancomycin, linezolid) is being compromised by resistant bacteria.51,52
Limited antibiotic development pipeline
The number of cases of Multidrug-resistant (MDR) infections has increased but there has been no new class of antibiotics for many years. Only a few drug approval in the past decade are simply modifications to existing antibiotics.53,54 There are economic barriers that discourage investment in new antibiotics and resistance emerges rapidly after a new agent is approved.55 The cost of some new antibiotics, such as ceftaroline, tedizolid, dalbavancin, and cefiderocol, along with the development of resistance to these agents, will greatly reduce their effectiveness, especially in low- and middle-income countries.56
Phenotypic Vs genotypic discrepancies in resistance detection
The main obstacle in managing antimicrobials is the poor correlation between the phenotypic susceptibility testing and genomic resistance prediction.57 Although phenotypic AST remains the gold standard, that may be missed in the presence of an inducible or low-level resistance mechanism that is active in the case of treatment failure, genomic methods can identify the resistance mechanism overestimated.58
These inconsistencies complicate treatment decisions, particularly in multidrug-resistance-capable pathogens, including P. aeruginosa, which tightly control efflux pumps and porins dynamically to vary the response to therapy.59 Without integrated phenotypic-genotypic analysis, an incorrect therapeutic decision and an increased delay in effective treatment is more likely.
Misuse/overuse of antibiotics and stewardship challenges
Antibiotic misuse (such as the unnecessary prescribing of antibiotics, incomplete courses, and over-the-counter access) is one of the primary contributors to resistance, being a significant problem on a global scale and most notably in LMICs with minimal regulation and diagnostics (Table 3).60-62,67 The role of antibiotic stewardship programs is clear, but unless integrated with stewardship, surveillance and infection-control, resistance will keep outpacing the creation of new antibiotics.
Table (3):
Therapeutic Challenges and Limitations of Current Antibiotics
Therapeutic Challenge |
Summary of Evidence |
Ref. |
|---|---|---|
Declining efficacy of standard antibiotics |
MRSA shows high β-lactam resistance via mecA/mecC and demonstrates rising carbapenem resistance (>40% in many LMICs). Resistance increasing across clindamycin, fluoroquinolones, tetracyclines; reduced susceptibility even to vancomycin and linezolid. |
46-52 |
Limited antibiotic development pipeline |
Few new antibiotic classes in 30 years; reliance on modified old classes. The development of new antibiotics incurs high costs and offers limited incentives. Newer agents (ceftaroline, tedizolid, cefiderocol, and ceftolozane–tazobactam) exist, but access and early resistance remain major issues. |
53-56 |
Phenotypic–genotypic discrepancies in resistance detection |
Phenotypic AST may miss inducible or low-level resistance (e.g., iMLSB, VISA). Genomics may detect ARGs not phenotypically expressed, complicating treatment decisions. Dynamic resistance modulation in pathogens like P. aeruginosa worsens mismatch. |
57-59 |
Misuse/overuse of antibiotics and stewardship challenges |
Inappropriate prescriptions, incomplete courses, OTC access, and inadequate regulation drive resistance. Globally, stewardship programs remain uneven, particularly in areas with limited diagnostics and trained personnel. |
60-62 |
Table emphasizes the impact of antibiotic misuse and gaps in antimicrobial stewardship on resistance emergence.
Future directions and novel therapies
The increasing number of multidrug-resistant Staphylococcus aureus worldwide will require alternative treatments or prevention methods for S. aureus infections than conventional antibiotic treatments. These new methods include a number of emerging approaches to treat or prevent S. aureus infections, including the attenuation of virulence, the modulation of the host immune system (immunomodulation), bacteriophages and CRISPR-based therapy, precision antimicrobial treatment, manipulation of the microbiota, and artificial intelligence based drug development. Representative examples of the key innovations, the biological basis of these innovative technologies and the evidence from research studies that support each strategy is shown in Table 4 and Figure.
Table (4):
Emerging and innovative strategies for the prevention and treatment of methicillin-resistant Staphylococcus aureus (MRSA) infections
Section |
Novel approach |
Key strategy and mechanism |
Representative references |
|---|---|---|---|
5.1 |
Novel antimicrobials & synthetic antibiotics |
Fifth-generation cephalosporins (ceftaroline, ceftobiprole) target PBP2a; synthetic scaffolds designed to evade enzymatic degradation and efflux mechanisms |
63–65 |
5.2 |
Anti-virulence and host-directed therapies |
Inhibition of key virulence factors (α-hemolysin, PVL, Agr quorum sensing) to reduce pathogenicity and selective pressure; immune-enhancing monoclonal antibodies |
66 |
5.3 |
Bacteriophage therapy |
Phages selectively lyse MRSA, penetrate biofilms, and act synergistically with antibiotics; engineered phages enhance lytic activity or deliver CRISPR payloads |
68,69 |
5.4 |
CRISPR-based antimicrobials |
Targeted silencing of resistance genes (mecA, erm, tet) or induction of lethal chromosomal damage; species- and gene-specific eradication |
70 |
5.5 |
Antimicrobial peptides & nanotechnology |
AMPs exhibit potent anti-MRSA activity; nanoparticle systems improve stability, penetration, and biofilm-targeted delivery |
71 |
5.6 |
Vaccines and immunotherapies |
Multi-antigen vaccine strategies targeting conserved virulence determinants; passive immunotherapy using monoclonal antibodies under evaluation |
72 |
5.7 |
Microbiome Modulation |
Modulation of skin and nasal microbiota using S. epidermidis–derived bacteriocins or FMT-derived components to prevent MRSA colonization |
73 |
5.8 |
AI-driven drug discovery & global priorities |
AI and deep learning accelerate identification of novel anti-MRSA compounds, with emphasis on global AMR surveillance, genomic monitoring, and stewardship |
74,75 |
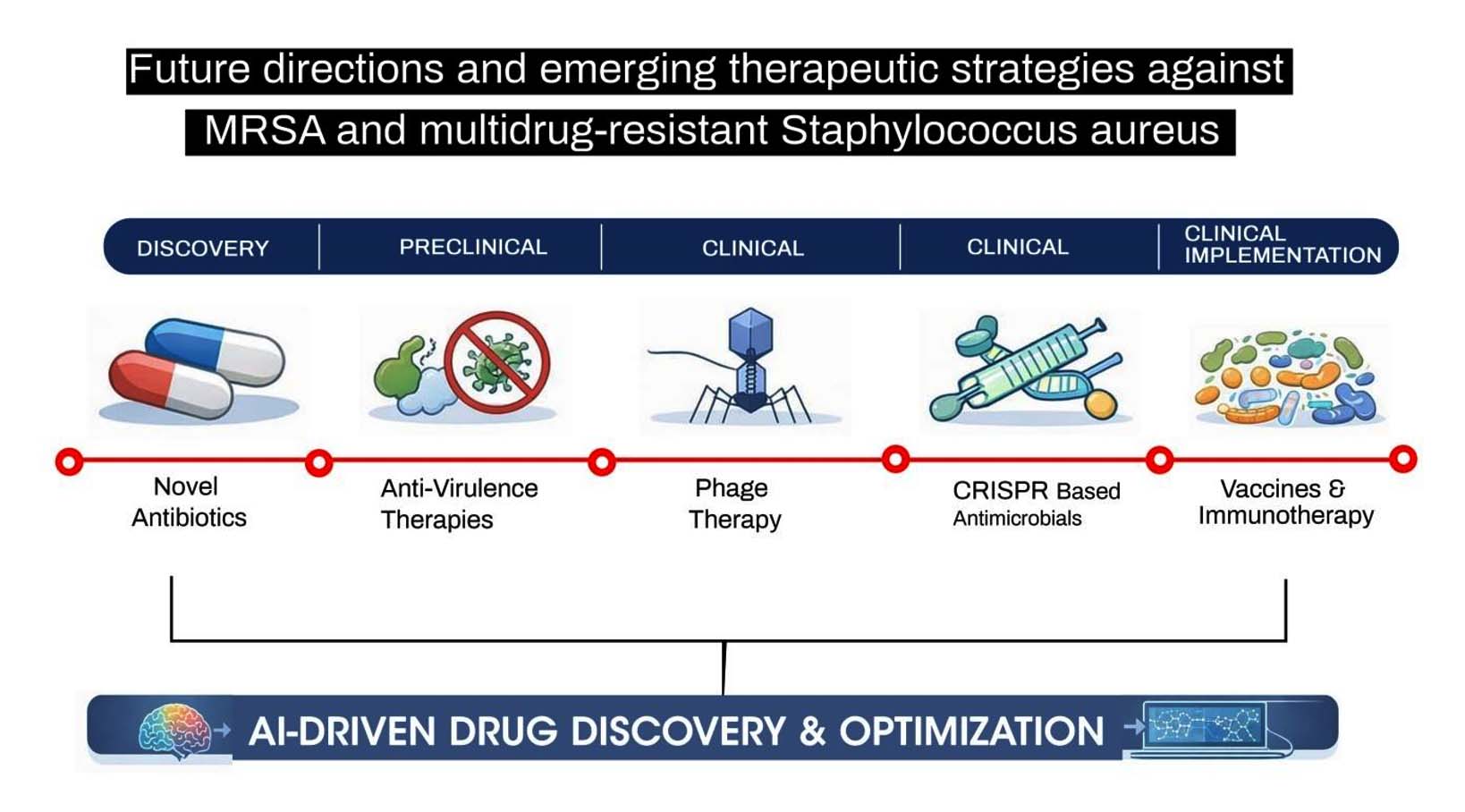
Figure. Future directions and emerging therapeutic strategies against methicillin-resistant Staphylococcus aureus (MRSA) and multidrug-resistant S. aureus
The multidrug-resistant (MDR) form of Staphylococcus aureus continues to be one of the most significant threats to public health around the world with continued trends of resistance from 2015-2025 especially in lower- and middle-income countries. Additionally, the emergence of new strains of MDR S. aureus as a result of the spread of high-risk clones due to the movement of mobile genetic elements, highlights the importance of improved molecular and genomic surveillance. To effectively prevent the further spread of MDR S. aureus will require an integration of appropriate antimicrobial stewardship programs, prudent antibiotic use, effective and consistent infection control practices and ongoing funding to develop new therapeutic options. Global collaboration among the clinical, public health and research communities is critical to reducing the potential burden of MDR S. aureus that will continue into the future.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Touaitia R, Mairi A, Ibrahim NA, Basher NS, Idres T, Touati A. Staphylococcus aureus: a review of the pathogenesis and virulence mechanisms. Antibiotics. 2025;14(5):470.
Crossref - Michalik M, Podbielska-Kubera A, Dmowska-Koroblewska A. Antibiotic resistance of Staphylococcus aureus strains—searching for new antimicrobial agents—review. Pharmaceuticals. 2025;18(1):81.
Crossref - Livermore DM. Antibiotic resistance in staphylococci. Int J Antimicrob Agents. 2000;16(Suppl 1):S3-S10.
Crossref - Lade H, Kim JS. Molecular determinants of β-lactam resistance in methicillin-resistant Staphylococcus aureus (MRSA): an updated review. Antibiotics. 2023;12(9):1362.
Crossref - Lakhundi S, Zhang K. Methicillin-resistant Staphylococcus aureus: molecular characterization, evolution, and epidemiology. Clin Microbiol Rev. 2018;31(4):e00020-18.
Crossref - DeLeo FR, Chambers HF. Reemergence of antibiotic-resistant Staphylococcus aureus in the genomics era. J Clin Invest. 2009;119(9):2464-2474.
Crossref - Chambers HF, DeLeo FR. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol. 2009;7(9):629-641.
Crossref - Lindsay JA. Hospital-associated MRSA and antibiotic resistance—what have we learned from genomics? Int J Med Microbiol. 2013;303(6-7):318-323.
Crossref - Otto M. Community-associated MRSA: what makes them special? Int J Med Microbiol. 2013;303(6-7):324-330.
Crossref - Laxminarayan R, Duse A, Wattal C, et al. Antibiotic resistance—the need for global solutions. Lancet Infect Dis. 2013;13(12):1057-1098.
Crossref - GBD 2021 Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. Lancet. 2024;404(10459):1199-1226.
Crossref - European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2021–2022. EFSA J. 2024;22(2):8583.
Crossref - Haindongo EH, Ndakolo D, Hedimbi M, Vainio O, Hakanen A, Vuopio J. Antimicrobial resistance prevalence of Escherichia coli and Staphylococcus aureus among bacteremic patients in Africa: a systematic review. J Glob Antimicrob Resist. 2023;32:35-43.
Crossref - Shyam H, Bhuvaneshwari G, Aruna D, Neelusree P. Retrospective Study on the Prevelance of Antimicrobial susceptibility pattern of staphylococcus species among patients attending Tertiary Care Hospital. Research Journal of Pharmacy and Technology. 2023; 16(11):5354-7.
Crossref - Hao M, Wang J. Molecular epidemiology, antimicrobial resistance, and virulence characteristics of predominant methicillin-resistant Staphylococcus aureus clones with strong biofilm-producing capability from a tertiary teaching hospital in China. BMC Microbiol. 2025;25(1):510.
Crossref - Guo W, Zhao Y, Yang Y, et al. YZ462 exhibits potent antibacterial activity against methicillin-resistant Staphylococcus aureus through bacterial membrane disruption. BMC Microbiol. 2025;25:728.
Crossref - Lindsay JA, Holden MTG. Understanding the rise of the superbug: investigation of the evolution and genomic variation of Staphylococcus aureus. Funct Integr Genomics. 2006;6(3):186-201.
Crossref - Turner NA, Sharma-Kuinkel BK, Maskarinec SA, et al. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nat Rev Microbiol. 2019;17(4):203-218.
Crossref - Foster TJ. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol Rev. 2017;41(3):430-449.
Crossref - Shopsin0 B, Kreiswirth BN. Molecular epidemiology of methicillin-resistant Staphylococcus aureus. Emerg Infect Dis. 2001;7(2):323-326.
Crossref - Elbehiry A, Marzouk E, Abalkhail A. Antimicrobial resistance at a turning point: microbial drivers, One Health, and global futures. Front Microbiol. 2025;16:1698809.
Crossref - Ho CS, Wong CTH, Wong C, et al. Antimicrobial resistance: a concise update. Lancet Microbe. 2025;6(1):100947.
Crossref - Foster TJ. Immune evasion by staphylococci. Nat Rev Microbiol. 2005;3(12):948-958.
Crossref - Kourtis AP, Hatfield K, Baggs J, et al. Vital signs: epidemiology and recent trends in methicillin resistant Staphylococcus aureus infections in the United States. MMWR Morbidity and Mortality Weekly Report. 2019;68(9):214–219.
Crossref - Fuda C, Suvorov M, Vakulenko SB, Mobashery S. The basis for resistance to β-lactam antibiotics by penicillin-binding protein 2a of methicillin-resistant Staphylococcus aureus. J Biol Chem. 2004;279(39):40802–40806.
Crossref - Garcia-Alvarez L, Holden MTG, Lindsay H, et al. Methicillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: a descriptive study. Lancet Infect Dis. 2011;11(8):595–603.
Crossref - Paterson GK, Harrison EM, Holmes MA. The emergence of mecC methicillin-resistant Staphylococcus aureus. MBio. 2014;22(1):42-47.
Crossref - Hiramatsu K, Ito T, Tsubakishita S, et al. Genomic basis for methicillin resistance in Staphylococcus aureus. Infect Chemother. 2013;45(2):117–136.
Crossref - Jibu RM, Geetha RV, Lakshmi T. Isolation, detection and molecularcharacterization of Staphylococcus aureus from postoperative infections. Journal of Pharmaceutical Research International. 2020;32(15):63-67.
Crossref - Leclercq R. Mechanisms of resistance to macrolides and lincosamides: nature of the resistance elements and their clinical implications. Clin Infect Dis. 2002;34(4):482-492.
Crossref - Truong-Bolduc QC, Strahilevitz J, Hooper DC. NorC, a new efflux pump regulated by MgrA of Staphylococcus aureus. Antimicrob Agents Chemother. 2006;50(3):1104–1107.
Crossref - D’Souza N, Rodrigues C, Mehta A. Molecular characterization of methicillin-resistant Staphylococcus aureus with emergence of epidemic clones of sequence type (ST) 22 and ST 772 in Mumbai, India. J Clin Microbiol. 2010;48(5):1806-1811.
Crossref - Roberts MC. Update on acquired tetracycline resistance genes. FEMS Microbiol Rev. 2005;245(2):195-203.
Crossref - Murray CJL, Ikuta KS, Sharara F, et al. Global burden of bacterial antimicrobial resistance in 2019. Lancet. 2022;399(10325):629–655.
Crossref - Holmes AH, Moore LSP, Sundsfjord A, et al. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet. 2016;387(10014):176-187.
Crossref - Tong SYC, Davis JS, Eichenberger E, Holland TL, Fowler VG Jr. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28(3):603-661.
Crossref - Novick RP, Geisinger E. Quorum sensing in staphylococci. Annu Rev Genet. 2008;42:541-564.
Crossref - World Health Organization. Global antimicrobial resistance and use surveillance report 2024. Geneva, Switzerland: World Health Organization; 2025. Accessed December 11, 2025. https://www.who.int/publications/i/item/9789240116337
- Liu C, Bayer A, Cosgrove SE, et al. Clinical practice guidelines by the Infectious Diseases Society of America for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children. Clin Infect Dis. 2011;52(3):e18-e55.
Crossref - Laxminarayan R, Matsoso P, Pant S, et al. Access to effective antimicrobials: a worldwide challenge. Lancet. 2016;387(10014):168-175.
Crossref - Howden BP, Davies JK, Johnson PD, Stinear TP, Grayson ML. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: resistance mechanisms, laboratory detection, and clinical implications. Clin Microbiol Rev. 2010;23(1):99-139.
Crossref - Otter JA, French GL. Community-associated methicillin-resistant Staphylococcus aureus strains as a cause of healthcare-associated infection. J Hosp Infect. 2011;79(3):189-193.
Crossref - Munita JM, Arias CA. Mechanisms of antibiotic resistance. Microbiol Spectr. 2016;4(2):VMBF-0016- 2015.
Crossref - O’Neill J. Tackling drug-resistant infections globally: final report and recommendations (Review on Antimicrobial Resistance). London: HM Government and Wellcome Trust. 2016. https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf. Accessed December 13, 2025.
- Indian Council of Medical Research (ICMR). Annual Report: Antimicrobial Resistance Surveillance and Research Network (Jan 2023–Dec 2023). New Delhi: ICMR. 2023. https://www.icmr.gov.in/icmrobject/uploads/Documents/1725536060_annual_report_2023.pdf. Accessed December 13, 2025.
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629-655.
Crossref - Prestinaci F, Pezzotti P, and Pantosti A. Antimicrobial resistance is a complex global issue. Pathogens and Global Health. 2015;109(7):309-318.
Crossref - Ahmad M, Aduru SV, Smith RP, Zhao Z, Lopatkin AJ. The role of bacterial metabolism in antimicrobial resistance. Nat Rev Microbiol. 2025;23(7):439-454.
Crossref - Christaki E, Marcou M, Tofarides A. Antimicrobial resistance in bacteria: mechanisms, evolution, and persistence. J Mol Evol. 2020;88(1):26-40.
Crossref - World Health Organization. Global Antibiotic Resistance and Use Surveillance Report: 2024. Geneva: World Health Organization. 2024. Accessed December 13, 2025. https://www.who.int/publications/i/item/9789240116337
- Xie Y, Lu H, Liu Y, et al. Unveiling the mechanisms of bacterial resistance and countermeasures. Pathogens. 2025;14(11):1085.
Crossref - Zhanel GG, Voth D, Nichol K, Karlowsky JA, Noreddin AM, Hoban DJ. Pharmacodynamic activity of ceftobiprole compared with vancomycin versus methicillinresistant Staphylococcus aureus (MRSA), vancomycin-intermediate Staphylococcus aureus (VISA), and vancomycin-resistant Staphylococcus aureus (VRSA) using an in vitro model. J Antimicrob Chemother. 2009;64(2):364-369.
Crossref - Theuretzbacher U, Bush K, Harbarth S, Paul M, Rex JH, Tacconelli E, Thwaites GE. Critical analysis of antibacterial agents in clinical development. Nat Rev Microbiol. 2020;18(5):286-298.
Crossref - Renwick MJ, Mossialos E. What are the economic barriers of antibiotic R&D and how can we overcome them? Expert Opin Drug Discov. 2018;13(10):889-892.
Crossref - Jones RN, Sader HS, Beach ML. Contemporary in vitro spectrum of activity summary for antimicrobial agents tested against 18569 strains non-fermentative Gram-negative bacilli isolated in the SENTRY Antimicrobial Surveillance Program (1997-2001). Int J Antimicrob Agents. 2003;22(6):551-6.
Crossref - Boucher HW, Wilcox M, Talbot GH, et al. Once-weekly dalbavancin versus daily conventional therapy for skin infection. N Engl J Med. 2014;370(23):2169–2179.
Crossref - Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing. 34th ed. CLSI supplement M100. Wayne, PA: Clinical and Laboratory Standards Institute; 2024. Accessed December 13, 2025
- Boolchandani M, D’Souza AW, Dantas G. Sequencing-based methods and resources to study antimicrobial resistance. Nat Rev Genet. 2019;20(6):356-370.
Crossref - Moradali MF, Ghods S, Rehm BHA. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front Cell Infect Microbiol. 2017;7:39.
Crossref - Interagency Coordination Group on Antimicrobial Resistance. No Time to Wait: Securing the Future from Drug-Resistant Infections: Report to the Secretary-General of the United Nations. World Health Organization; 2019. Accessed December 13, 2025. https://www.who.int/publications/i/item/no-time-to-wait-securing-the-future-from-drug-resistant-infections
- Centers for Disease Control and Prevention. Antibiotic Resistance; 2019 Antibiotic Resistance Threats Report. Accessed December 13, 2025. https://www.cdc.gov/antimicrobial-resistance/data-research/threats/index.html
- Indian Council of Medical Research. Antimicrobial Resistance Surveillance Network Annual Report 2023. New Delhi: ICMR. 2023. https://www.icmr.gov.in/icmrobject/uploads/Documents/1725536060_annual_report_2023.pdf. Accessed December 13, 2025.
- Nandhini P, Kumar P, Mickymaray S, Alothaim AS, Somasundaram J, Rajan M. Recent developments in methicillin-resistant Staphylococcus aureus (MRSA) treatment: a review. Antibiotics. 2022;11(5):606.
Crossref - Koulenti D, Xu E, Mok IYS, et al. Novel antibiotics for multidrug-resistant gram-positive microorganisms. Microorganisms. 2019;7(8):270.
Crossref - Ling LL, Schneider T, Peoples AJ, et al. A new antibiotic kills pathogens without detectable resistance. Nature. 2015;517(7535):455-459.
Crossref - Sully EK, Malachowa N, Elmore BO, et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoSPathog. 2014;10(6):e1004174.
Crossref - Fowler VG Jr, Das AF, Lipka-Diamond J, et al. Exebacase (CF-301) for patients with Staphylococcus aureus bloodstream infection and endocarditis: results of a randomized, double-blind, placebo-controlled trial. J Clin Invest. 2020;130(7):3750-3760.
Crossref - Plumet L, Ahmad-Mansour N, Dunyach-Remy C, et al. Bacteriophage Therapy for Staphylococcus Aureus Infections: A Review of Animal Models, Treatments, and Clinical Trials. Front Cell Infect Microbiol. 2022;12:907314.
Crossref - Dedrick RM, Guerrero-Bustamante CA, Garlena RA, et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus infection. Nat Med. 2019;25(5):730-733.
Crossref - Bikard D, Euler CW, Jiang W, et al. Exploiting CRISPR-Cas systems for targeted antimicrobial therapy against Staphylococcus aureus. Nat Biotechnol. 2014;32(11):1141-1145.
Crossref - Mahlapuu M, Hakansson J, Ringstad L, Bjorn C. Antimicrobial peptides: an emerging category of therapeutic agents. Front Microbiol. 2020;11:566.
Crossref - Proctor RA. Challenges for a universal Staphylococcus aureus vaccine. Clin Infect Dis. 2012;54(8):1179-1186.
Crossref - Nakatsuji T, Chen TH, Narala S, et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci Transl Med. 2017;9(378):eaah4680.
Crossref - Stokes JM, Yang K, Swanson K, et al. A deep learning approach to antibiotic discovery. Cell. 2020;180(4):688-702.
Crossref - World Health Organization. Global antimicrobial resistance and use surveillance system (GLASS) reports. Geneva: World Health Organization; 2022-2024. https://www.who.int/initiatives/glass. Accessed December 13, 2025.
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.