


ISSN: 0973-7510
E-ISSN: 2581-690X
Multidrug-resistant Pseudomonas aeruginosa has emerged as an important opportunistic pathogen. This study elucidates the prevalence, antimicrobial resistance patterns, and biofilm formation of this pathogen in wound and urinary tract infections. Pseudomonas selective agar and MacConkey agar were used to analyze 432 clinical samples [wound swabs (210) and urine samples (222)] through selective culturing. Then, API 20E strips and the Vitek 2 Compact System were employed for the pathogen identification. Mueller-Hinton agar-based disk diffusion method revealed the antimicrobial susceptibility of Pseudomonas aeruginosa. The crystal violet staining method was adopted to investigate biofilm formation using microtiter plates. The virulence genes (toxA, algD, and phzS) were amplified via PCR. A low Pseudomonas aeruginosa prevalence (1.85%) was noted in analyzed samples. However, multidrug-resistance was noted in 65% of Pseudomonas aeruginosa isolates, whereas 25% isolates demonstrated extensive resistance. Despite the presence of the algD gene, biofilm formation was weak in most Pseudomonas aeruginosa isolates and did not correlate statistically with antimicrobial resistance. The virulence factors encoding genes (toxA, algD, phzS) were noted in most of the Pseudomonas aeruginosa isolates. The results highlighted the epidemiology of multidrug-resistant and virulent Pseudomonas aeruginosa in Saudi Arabia. The rising occurrence of Pseudomonas aeruginosa necessitates detailed elaboration on its virulence, prevalence, and antimicrobial susceptibility patterns in Saudi Arabia through molecular tools.
Pseudomonas aeruginosa, Wound Infections, Antibiotic Resistance, Virulence Factors, Biofilm
The Gram-negative Pseudomonas aeruginosa is a non-spore-forming and aerobic motile bacteria. It is widely prevalent in most environments even with minimal nutritional sources. Aquatic environment and soil are natural habitats of this bacterium but its growth has been reported in shampoo and distilled water as well.1 It rarely causes diseases in healthy individuals, however, this opportunistic pathogen is an established source of nosocomial infections in hospitalized and immunocompromised patients.2,3 Several chronic and acute infections are associated with this pathogen including pneumonia, hospital-acquired urinary tract infections, surgical wound infections, and septicemia. Moreover, various systemic infections are also linked to this pathogen in patients suffering from HIV, cystic fibrosis, hepatitis C, severe burns, and cancer.2,4
Cell-related virulence factors mainly contribute to Pseudomonas aeruginosa pathogenicity. Flagella is an important factor that facilitates its motility into the host tissues.3 Similarly, lipopolysaccharide (LPS) is another crucial factor regarding the pathogenesis of Pseudomonas aeruginosa.4 It also secretes various virulent toxins and enzymes such as pyocyanin, exoenzyme S, proteases, exotoxin A (ExoA), and hemolysins.4 ExoA catalyzes ADP-ribosylation of eukaryotic elongation factor-2 to affect host cells’ protein synthesis process leading to cellular mortality and pathogen invasion.5 Pyocyanin, a green-colored phenazine, is generally known to restrict pathogenic bacterial growth. Contrarily, this pigment helps in Pseudomonas aeruginosa pathogenesis by inhibiting wound recovery via cellular damage and neutrophil apoptosis acceleration.6 The algABCD operon of Pseudomonas aeruginosa secretes an extracellular water-soluble polysaccharide (alginate) that is critical for its biofilm-forming capability.7 Pseudomonas aeruginosa forms a polysaccharide-rich biofilm under stressed conditions to avoid antibiotics and environmental pressure. Biofilm helps the sustained growth of this pathogen, and is linked to various chronic infections, antibiotic resistance, and colonization of surgical implants, catheters, and endotracheal tubes.8,9
The lower antibiotic susceptibility of Pseudomonas aeruginosa is an alarming aspect, which is attributed to multidrug efflux pumps and antibiotic resistance encoding genes on chromosome and mobile genetic elements.3 The transfer of resistance genes in Pseudomonas aeruginosa could occur through mobile genetic elements’ horizontal transfer. Antibiotic resistance-linked genes and enzymes differ in various strains.2-4,10-13 Antibiotic-resistant Pseudomonas aeruginosa isolates cause treatment failures. This pathogen has even developed resistance against common first-line antibiotics.2-4,10-13
The incidence of antibiotic-resistant clinical Pseudomonas aeruginosa infections is rising in Saudi Arabia, which coincides with global trends. This pathogen is often detected in healthcare settings, particularly in samples from invasive devices and chronic and immunosuppressive patients.14-17 Studies in Saudi Arabian hospitals have demonstrated a high prevalence of cephalosporin, aminoglycoside, and carbapenem-resistant Pseudomonas aeruginosa strains. Pan-drug-resistant (PDR), multidrug-resistant (MDR), and extensively drug-resistant (XDR) strains of Pseudomonas aeruginosa have also been documented.15-20 This study investigated Pseudomonas aeruginosa prevalence, antimicrobial resistance patterns, and biofilm formation in urinary tract and wound infections in Western Saudi Arabia. The study involved the investigations of key virulence factors-related genes (toxA, algD, and phzS), assessment of epidemiological trends, and elucidation of challenges related to multidrug- and extensively drug resistant Pseudomonas aeruginosa strains in healthcare settings.
Sample collection
The clinical samples (432) of hospitalized patients included wound swabs (210) and mid-stream urine samples (222). Samples from one hospital in Makkah, Saudi Arabia, were collected from October 2021 to January 2022. Sterile swabs in Amies Charcoal Transport Medium (Zhejiang Runlab Technology Co. Ltd., China) were used to collect wound swab samples, whereas sterile urine sample bottles were used to collect midstream urine samples. Samples were transported to the lab in an ice box without direct sunlight exposure, and microbiological investigations were initiated on the same day. No data of personal, clinical or epidemiological type was provided for the study, except for the gender of the sample provider.
Isolation and identification of Pseudomonas aeruginosa
The urine and wound samples were cultured (24 hrs at 37 °C) in CFC (cephalothin-sodium fusidate-cetrimide) supplemented Pseudomonas selective agar base (HiMedia, India) and MacConkey agar (Oxoid, UK).21 Pseudomonas aeruginosa was presumptively identified via Gram staining, oxidase test strips (Oxoid), and API 20E strips (bioMerieux, France). Vitek 2 Compact System (bioMerieux) was employed for the further confirmation of Pseudomonas aeruginosa isolates.17
Antimicrobial susceptibility profiles of Pseudomonas aeruginosa
Antimicrobial susceptibility tests of confirmed Pseudomonas aeruginosa isolates were performed by following the Kirby-Bauer disk diffusion method22 and guidelines of the Clinical and Laboratory Standards Institute (CLSI).23 The tested antimicrobial agents were selected based on the CLSI recommendations regarding Pseudomonas aeruginosa clinical breakpoints.23 Briefly, a standard suspension (McFarland 0.5) of each isolate was spread onto Mueller-Hinton agar plates (HiMedia) and incubated (37 °C, 18-24 hrs) along with antibiotic disks. Twelve antibiotics (Oxoid, UK) from 7 antimicrobial classes were used in antimicrobial susceptibility test,23 which included Piperacillin-Tazobactam (100/10 µg) [β-lactam combinations], Piperacillin (100 µg) [penicillins], Levofloxacin (5 µg), Ciprofloxacin (5 µg) [fluoroquinolones], Cefepime (30 µg), Ceftazidime (30 µg) [cephalosporins], Tobramycin (10 µg), Amikacin (30 µg), Gentamicin (10 µg) [aminoglycosides], Aztreonam (30 µg) [monobactams], and Meropenem (10 µg), Imipenem (10 µg) [carbapenems]. ESBL (extended-spectrum beta-lactamase) phenotypes of Pseudomonas aeruginosa were detected using CHROMagar ESBL (Saudi Prepared Laboratory Media – SPLM, Saudi Arabia) after the incubation of plates (37 °C, 24 h).24
Multiple antibiotic resistance index (MARI)
MARI was determined as the ratio of the number of antibiotics to which an isolate exhibited resistance to the total number of antibiotics tested.25 A MARI ³0.2 indicated that the isolates from high antibiotic-use environments, whereas a MARI value £0.2 suggests isolates from low exposure settings.26
Biofilm formation in Pseudomonas aeruginosa
Biofilm-forming ability of Pseudomonas aeruginosa was evaluated using a modified 96-well microtiter plate assay as previously described.26-28 Briefly, a fresh medium (108 CFU, or 0.5 McFarland standard) was used to dilute overnight nutrient broth (HiMedia) cultures. Inoculum aliquots (20 µl) and fresh broth (180 µL) were separately added to sterile 96-well microtiter plates (Corning Incorporated, Life Sciences, USA), and incubated (37 °C) for 24 hrs. Microtiter plates were washed thrice with distilled water and stained with 1% crystal violet solution (200 µL). The non-adherent cells were removed by rinsing the plates again with distilled water. The plates were then air-dried, after which bound dye was solubilized using 150 µL of 95% ethanol. Absorbance was measured at 590 nm using a microplate reader (BioTek Synergy 2 Multimode Plate Reader, USA). The result consistency was assured by performing the assay in triplicate. Biofilm forming capability of Pseudomonas aeruginosa was calculated and ranked as strong (S), moderate (M), and weak (W) according to the formula mentioned in Table 1.26-28
Table (1):
Biofilm formation in Pseudomonas aeruginosa
Biofilm formation |
Strong |
Moderate |
Weak |
|---|---|---|---|
BF= AB – CW |
0.200-0.299 |
0.100-0.199 |
|
BF= AB / CW |
4.00-5.99 |
2.00-3.99 |
BF: biofilm, AB: stained attached bacteria, CW: stained control wells
Molecular detection of virulence factors-encoding genes in Pseudomonas aeruginosa
Genomic DNA from Pseudomonas aeruginosa isolates was extracted using the AllPrep Bacterial DNA kit (Qiagen, USA). A spectrophotometer (Denovix DS-11, USA) was used to measure the quality and quantity of the DNA. PCR amplification of virulence-associated genes, namely toxA (352 bp),29 algD (1310 bp),30 and phzS (1752 bp),31 was performed using DreamTaq Green PCR Master Mix (2×) (Thermo Fisher Scientific, USA). Primer sequences, expected amplicon size, and the annealing temperatures of the genes are provided in Table 2. PCR reactions were carried out in a Veriti Thermal Cycler (Thermo Fisher Scientific), with a final reaction volume of 25 µL, comprising 12.5 µL of master mix (2×), 1.0 µL of template DNA (10 ng/µL), 2.0 µL each of forward and reverse primers (10 µM), and 9.5 µL of nuclease-free water. Thermal cycling conditions for each gene were optimized as follows: toxA– initial denaturation at 95 °C for 3 min, 35 cycles of denaturation at 95 °C for 30 sec, and annealing at 52 °C for 30 sec, and extension at 72 °C, for 1 min, and a final extension at 72 °C for 7 min); phzS-similar conditions with annealing at 63 °C for 30 sec; and algD– with annealing at 50 °C for 30 sec.
Table (2):
Virulence factors-encoding gene primers in Pseudomonas aeruginosa
Gene primer |
Sequence 5’→3’ |
Product size (bp) |
Annealing temp. |
Ref. |
|---|---|---|---|---|
toxA |
F: 5′-GGTAACCAGCTCAGCCACAT-3′ R: 5′-TGATGTCCAGGTCATGCTTC-3′ |
352 |
52 °C |
[29] |
phzS |
F: 5′-TCGCCATGACCGATACGCTC-3′ R: 5′-ACAACCTGAGCCAGCCTTCC-3′ |
1752 |
63 °C |
[31] |
algD |
F: 5′-ATGCGAATCAGCATCTTTGGT-3′ R: 5′-CTACCAGCAGATGCCCTCGGC-3′ |
1310 |
50 °C |
[30] |
PCR amplified products’ size and quality were confirmed through agarose gel (2%) electrophoresis. TBE buffer (1X), ultrapure agarose (Cleaver Scientific, UK), and DNA stain (SYBR-safe) (Invitrogen, USA) were used in gel preparation. PCR amplification products (4 µL) were loaded into the gel wells. PCR products’ size was estimated against a reference 100-3000 bp DNA ladder (1 kb plus DNA ladder, Thermo Fisher). Gel electrophoresis was carried out for 30 minutes at 100 V, and a gel documentation system (UV Trans-illuminator) was used to observe the DNA fragments.26
Statistical analysis
Data analysis was conducted using SPSS Statistics (version 21.0). The relationship between multidrug-resistance profiles and biofilm-forming ability of Pseudomonas aeruginosa was examined by calculating Pearson’s correlation coefficient to assess linear association. A Fisher’s exact test was conducted to test the association between resistance phenotypes, resistance markers and virulence factors in Pseudomonas aeruginosa.
A P-value < 0.05 was considered statistically significant.
Control strains
Escherichia coli ATCC® 25922, Pseudomonas aeruginosa ATCC® BAA-1744, and Pseudomonas aeruginosa ATCC® 27853 served as the controls.
Pseudomonas aeruginosa prevalence in UTI and wound samples
The examination of wound swabs (210) and urine samples (222) depicted Pseudomonas aeruginosa occurrence in only 18 (8.57%) wound swabs and two (0.90%) urine samples (Figure 1). Overall Pseudomonas aeruginosa prevalence remained 4.63% in a total of 432 clinical samples. The growth of lactose-fermenting bacteria (Escherichia coli and Klebsiella spp.) was mostly noted in MacConkey plates, however, a few non-lactose fermenting bacteria (Acinetobacter spp. and/or Pseudomonas spp.) were observed as well. CFC-supplemented Pseudomonas selective agar established the presence of Pseudomonas spp., and the identity of all the growing isolates was further confirmed through API 20E strips and the Vitek 2 Compact System.
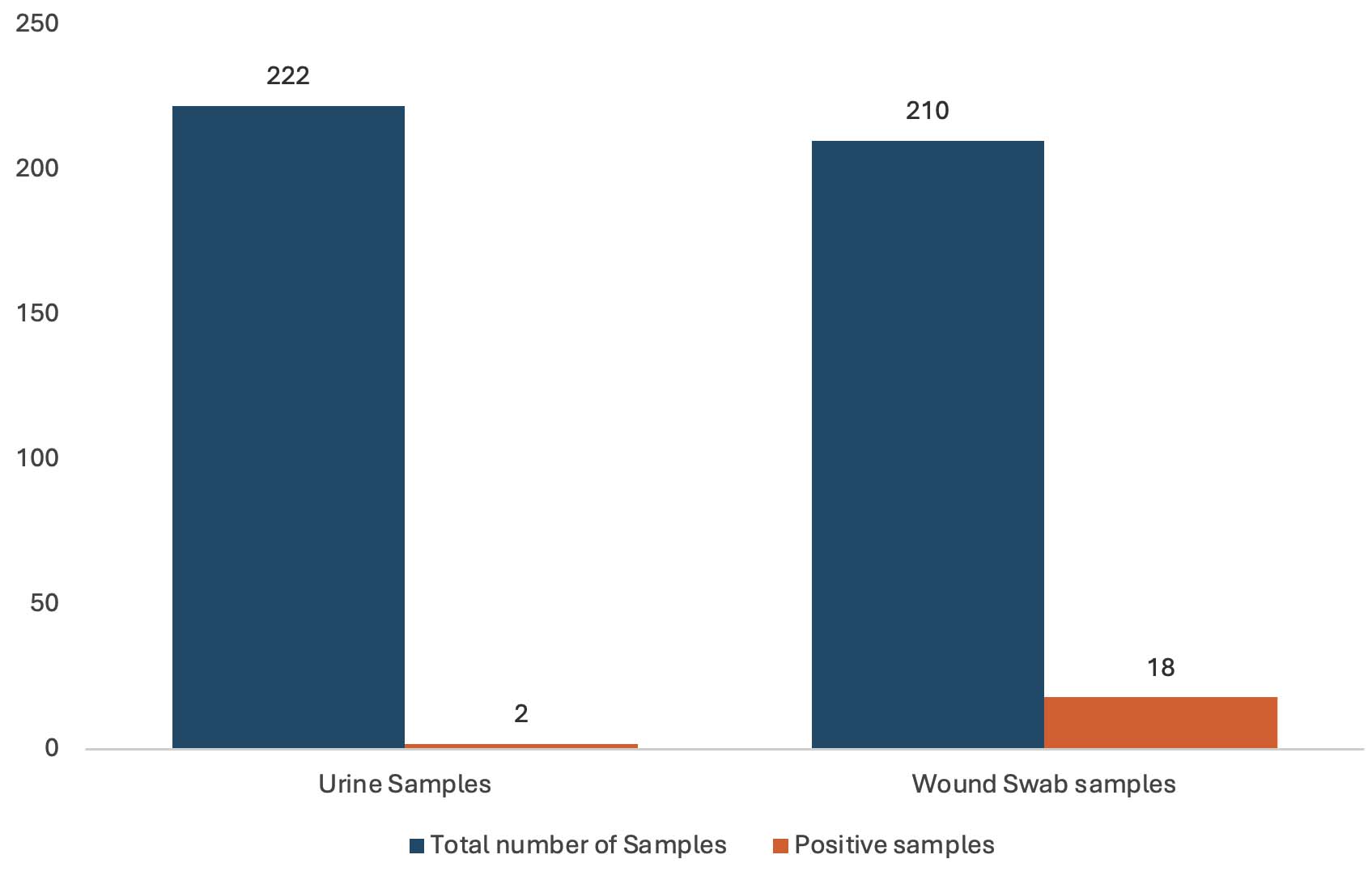
Figure 1. Prevalence of Pseudomonas aeruginosa in mid-stream urine and wound swab samples
Antimicrobial susceptibility testing of Pseudomonas aeruginosa
The resistance to two or more antimicrobial agents was detected in fifteen out of twenty (75%) wound swab Pseudomonas aeruginosa isolates (Table 3). Among the two urine isolates, one isolate was noted to be susceptible to all the tested antimicrobial agents except aztreonam. Overall, aztreonam faced the highest resistance rates (100%, n = 20) followed by levofloxacin, ceftazidime, and piperacillin (55%, n = 20) (Table 3). The resistance to imipenem and meropenem was observed in 30% of Pseudomonas aeruginosa isolates. Tobramycin and amikacin experienced the lowest resistance of 15% Pseudomonas aeruginosa isolates (Table 3).
Table (3):
Antimicrobial resistance profiles of Pseudomonas aeruginosa
| Sample | Isolates | Clear zone diameter (mm) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PRL | TZP | CAZ | FEP | IPM | MEM | ATM | CIP | LEV | TOB | CN | AK | ||
| Ps1 | WS | 28 (S) | 0 (R) | 25 (S) | 20 (S) | 30 (S) | 31 (S) | 0 (R) | 0 (R) | 0 (R) | 23 (S) | 15 (S) | 20 (S) |
| Ps2 | WS | 18 (I) | 22 (S) | 15 (I) | 27 (S) | 32 (S) | 30 (S) | 0 (R) | 35 (S) | 27 (S) | 22 (S) | 17 (S) | 21 (S) |
| Ps3 | WS | 29 (S) | 25 (S) | 28 (S) | 21 (S) | 30 (S) | 30 (S) | 0 (R) | 31 (S) | 26 (S) | 28 (S) | 19 (S) | 24 (S) |
| Ps4 | WS | 15 (I) | 11 (R) | 10 (R) | 14 (R) | 28 (S) | 15 (R) | 0 (R) | 8 (R) | 0 (R) | 20 (S) | 15 (S) | 19 (S) |
| Ps5 | WS | 32 (S) | 12 (R) | 29 (S) | 25 (S) | 31 (S) | 34 (S) | 0 (R) | 32 (S) | 28 (S) | 20 (S) | 15 (S) | 21 (S) |
| Ps6 | WS | 19 (I) | 19 (I) | 19 (S) | 21 (S) | 9 (R) | 3 (R) | 0 (R) | 25 (S) | 14 (R) | 21 (S) | 15 (S) | 18 (S) |
| Ps7 | UR | 16 (I) | 21 (S) | 14 (R) | 18 (S) | 31 (S) | 31 (S) | 0 (R) | 26 (S) | 15 (R) | 21 (S) | 16 (S) | 22 (S) |
| Ps8 | WS | 29 (S) | 27 (S) | 25 (S) | 25 (S) | 26 (S) | 33 (S) | 0 (R) | 32 (S) | 24 (S) | 20 (S) | 15 (S) | 19 (S) |
| Ps9 | WS | 15 (I) | 1 (R) | 14 (R) | 17 (I) | 26 (S) | 25 (S) | 0 (R) | 14 (R) | 8 (R) | 21 (S) | 15 (S) | 20 (S) |
| Ps10 | WS | 29 (S) | 24 (S) | 24 (S) | 25 (S) | 26 (S) | 33 (S) | 0 (R) | 0 (R) | 0 (R) | 20 (S) | 17 (S) | 22 (S) |
| Ps11 | WS | 25 (S) | 21 (S) | 13 (R) | 22 (S) | 29 (S) | 35 (S) | 20 (I) | 31 (S) | 25 (S) | 19 (S) | 15 (S) | 21 (S) |
| Ps12 | WS | 11 (R) | 0 (R) | 10 (R) | 12 (R) | 27 (S) | 20 (S) | 0 (R) | 6 (R) | 0 (R) | 19 (S) | 15 (S) | 19 (S) |
| Ps13 | WS | 11 (R) | 0 (R) | 0 (R) | 9 (R) | 11 (R) | 10 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) |
| Ps14 | WS | 11 (R) | 0 (R) | 0 (R) | 0 (R) | 11 (R) | 10 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) |
| Ps15 | WS | 11 (R) | 0 (R) | 0 (R) | 22 (S) | 0 (R) | 0 (R) | 0 (R) | 10 (R) | 0 (R) | 19 (S) | 15 (S) | 20 (S) |
| Ps16 | WS | 22 (S) | 22 (S) | 27 (S) | 29 (S) | 27 (S) | 32 (S) | 0 (R) | 30 (S) | 27 (S) | 28 (S) | 17 (S) | 21 (S) |
| Ps17 | WS | 24 (S) | 19 (I) | 24 (S) | 26 (S) | 28 (S) | 34 (S) | 0 (R) | 25 (S) | 15 (R) | 19 (S) | 14 (I) | 19 (S) |
| Ps18 | WS | 18 (I) | 14 (R) | 14 (I) | 15 (I) | 8 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 0 (R) | 7 (R) |
| Ps19 | WS | 16 (I) | 8 (R) | 11 (R) | 15 (I) | 31 (S) | 37 (S) | 0 (R) | 33 (S) | 25 (S) | 20 (S) | 16 (S) | 21 (S) |
| Ps20 | UR | 30 (S) | 21 (S) | 29 (S) | 25 (S) | 30 (S) | 35 (S) | 0 (R) | 30 (S) | 27 (S) | 31 (S) | 17 (S) | 24 (S) |
| Total resistance (%) | 11 (55) | 12 (60) | 11 (55) | 7 (35) | 5 (25) | 6 (30) | 20 (100) | 9 (45) | 11 (55) | 3 (15) | 4 (20) | 3 (15) | |
CAZ: Ceftazidime, FEP: Cefepime, IPM: Imipenem, MEM: Meropenem, TZP: Piperacillin-Tazobactam, PRL: Piperacillin, CN: Gentamicin, ATM: Aztreonam, AK: Amikacin, CIP: Ciprofloxacin, LEV: Levofloxacin, TOB: Tobramycin, S: susceptible, I: intermediate, R: resistant, WS: Wound swabs, UR: Urine samples, Interpretations of antimicrobial susceptibility testing were based on the CLSI clinical breakpoints for Pseudomonas aeruginosa23
MDR patterns were noted in thirteen (65%, n = 20) Pseudomonas aeruginosa isolates (Table 4). Notably, five Pseudomonas aeruginosa isolates (Ps4, Ps13, Ps14, Ps15, and Ps18) (25%) all from wound infection were established as XDR isolates (Table 4).
Table (4):
Antibiotic resistance patterns and correlation between antimicrobial resistance and biofilm formation in Pseudomonas aeruginosa
Isolate code |
Origin |
Resistance pattern |
Number of classes |
ESBL |
Type of resistance |
Biofilm formation |
MARI |
|---|---|---|---|---|---|---|---|
Ps1 |
WS |
TZP, ATM, CIP, LEV |
3 |
– |
MDR |
Weak |
0.34 |
Ps2 |
WS |
PRL (I)*, CAZ (I)*, ATM |
3 |
– |
MDR |
Weak |
0.25 |
Ps3 |
WS |
ATM |
1 |
– |
– |
Weak |
0.09 |
Ps4 |
WS |
PRL (I)*, TZP, CAZ, FEP, MEM, ATM, CIP, LEV |
6 |
+ |
XDR |
Weak |
0.67 |
Ps5 |
WS |
TZP, ATM |
2 |
– |
– |
Weak |
0.17 |
Ps6 |
WS |
PRL (I)*, TZP (I)*, IPM, MEM, ATM, LEV |
5 |
+ |
MDR |
Weak |
0.5 |
Ps7 |
UR |
PRL (I)*, CAZ, ATM, LEV |
4 |
– |
MDR |
Weak |
0.34 |
Ps8 |
WS |
ATM |
1 |
– |
– |
Weak |
0.09 |
Ps9 |
WS |
PRL (I)*, TZP (I)*, CAZ, FEP (I)*, ATM, CIP, LEV |
5 |
+ |
MDR |
Weak |
0.59 |
Ps10 |
WS |
ATM, CIP, LEV |
2 |
– |
– |
Weak |
0.25 |
Ps11 |
WS |
CAZ, ATM (I)* |
2 |
– |
– |
Weak |
0.17 |
Ps12 |
WS |
PRL, TZP, CAZ, FEP, ATM, CIP, LEV |
5 |
+ |
MDR |
Weak |
0.59 |
Ps13 |
WS |
PRL, TZP, CAZ, FEP, IPM, MEM, ATM, CIP, LEV, TOB, CN, AK |
7 |
+ |
XDR |
Weak |
1 |
Ps14 |
WS |
PRL, TZP, CAZ, FEP, IPM, MEM, ATM, CIP, LEV, TOB, CN, AK |
7 |
+ |
XDR |
Weak |
1 |
Ps15 |
WS |
PRL, TZP, CAZ, IPM, MEM, ATM, CIP, LEV |
6 |
+ |
XDR |
Weak |
0.67 |
Ps16 |
WS |
ATM |
1 |
– |
– |
Weak |
0.09 |
Ps17 |
WS |
TZP (I)*, ATM, LEV, CN (I)* |
4 |
+ |
MDR |
Weak |
0.34 |
Ps18 |
WS |
PRL (I)*, TZP, CAZ (I)*, FEP (I)*, IPM, MEM, ATM, CIP, LEV, TOB, CN, AK |
7 |
+ |
XDR |
Strong |
1 |
Ps19 |
WS |
PRL (I)*, TZP, CAZ, FEP (I)*, ATM |
5 |
+ |
MDR |
Weak |
0.42 |
Ps20 |
UR |
ATM |
1 |
– |
– |
Weak |
0.09 |
Pearson correlation |
r = – 0.1397 |
||||||
Total ESBL-positive N (%) |
10 (50) |
||||||
Total MDR N (%) |
13 (65)† |
||||||
Total XDR N (%) |
5 (25) |
CAZ: Ceftazidime, FEP: Cefepime, IPM: Imipenem, MEM: Meropenem, TZP: Piperacillin-Tazobactam, PRL: Piperacillin, CN: Gentamicin, ATM: Aztreonam, AK: Amikacin, CIP: Ciprofloxacin, LEV: Levofloxacin, TOB: Tobramycin, MARI: Multiple Antibiotic Resistance Index, MDR: multidrug resistance, XDR: extensive drug resistance, †MDR isolates counts also included XDR isolates.
ESBL: Extended-spectrum beta-lactamase. Each isolate was cultured on CHROMagar ESBL plates to determine ESBL. Pearson correlation analysis examined the association between biofilm formation and multidrug resistance in Pseudomonas aeruginosa.
CHROMagar ESBL facilitated the growth of ten (50%) ESBL-positive Pseudomonas aeruginosa isolates from wound swabs, whereas urine isolates remained unable to grow on CHROMagar ESBL (Table 4). MARI of 13 Pseudomonas aeruginosa isolates from wound swabs ranged between 0.25 and 1.0, which revealed their origin from frequent antibiotic administration areas. Contrarily, MARI of 0.09 and 0.17 in Pseudomonas aeruginosa isolates of wound infection and UTI demonstrated their origin from lesser antibiotic administration areas (Table 4).
Biofilm formation in Pseudomonas aeruginosa
Microtiter plates with crystal violet staining were used to examine the biofilm formation in all (20) of the detected Pseudomonas aeruginosa isolates. The data revealed a weak biofilm-forming efficiency in most Pseudomonas aeruginosa (95%, n = 20) isolates (Table 4).
Correlation between biofilm formation and multidrug-resistance in Pseudomonas aeruginosa isolates
Pearson correlation coefficients demonstrated a non-significant (r = -0.1397) relationship between biofilm formation and MDR (Table 4 and Figure 2). Notably, the particular drug resistance was not associated with biofilm formation. Biofilm-linked protein (algD)-encoding genes were detected in 16 (80%) Pseudomonas aeruginosa isolates (Table 5).
Table (5):
Prevalence of virulence factors-encoding genes in Pseudomonas aeruginosa
Isolate Code |
origin |
toxA |
phzS |
algD |
|---|---|---|---|---|
Ps |
WS |
– |
– |
+ |
Ps2 |
WS |
– |
– |
+ |
Ps3 |
WS |
+ |
+ |
+ |
Ps4 |
WS |
– |
– |
– |
Ps5 |
WS |
– |
+ |
+ |
Ps6 |
WS |
+ |
+ |
+ |
Ps7 |
UR |
+ |
+ |
+ |
Ps8 |
WS |
+ |
+ |
+ |
Ps9 |
WS |
+ |
+ |
+ |
Ps10 |
WS |
– |
+ |
+ |
Ps11 |
WS |
– |
– |
– |
Ps12 |
WS |
– |
– |
– |
Ps13 |
WS |
+ |
+ |
+ |
Ps14 |
WS |
+ |
+ |
+ |
Ps15 |
WS |
+ |
+ |
+ |
Ps16 |
WS |
– |
+ |
– |
Ps17 |
WS |
+ |
+ |
+ |
Ps18 |
WS |
+ |
+ |
+ |
Ps19 |
WS |
+ |
+ |
+ |
Ps20 |
UR |
+ |
+ |
+ |
BAA1774 |
Control |
+ |
+ |
+ |
Ps27853 |
Control |
+ |
+ |
+ |
Total N (%) |
12 (60) |
15 (75) |
16 (80) |
WS: wound swabs, UR: Urine samples, toxA: exotoxin A-encoding-gene, phzS: pyocyanin synthesis-encoding gene, algD: biofilm formation-encoding gene, BAA1774: Pseudomonas aeruginosa ATCC BAA1774, Ps27853: Pseudomonas aeruginosa ATCC 27853.
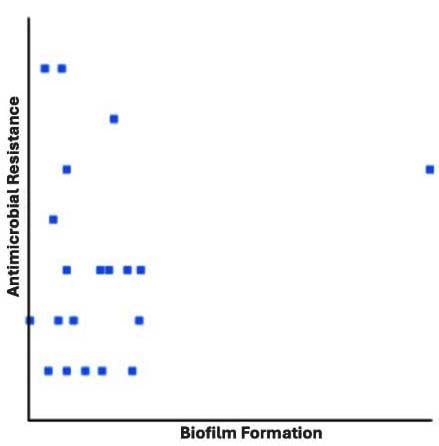
Figure 2. Pearson correlation between biofilm formation and antimicrobial resistance in Pseudomonas aeruginosa. No significant associated (r = – 0.1397) was observed
Virulence factors in Pseudomonas aeruginosa
Biofilm-associated protein (algD) gene was PCR amplified in 16 (80%) out of 20 Pseudomonas aeruginosa isolates (Table 5 and Figure 3). The presence of exotoxin A (toxA), and pyocyanin synthesis (phzS)-encoding genes was observed in 12 (60%) and 15 (75%) Pseudomonas aeruginosa isolates, respectively (Table 5, Figures 4 and 5). Twelve isolates harbored all three genes (toxA, algD, and phzS) where two isolates were of UTI origin and ten isolates came from wound infections (Table 5 and Figures 3, 4, and 5).
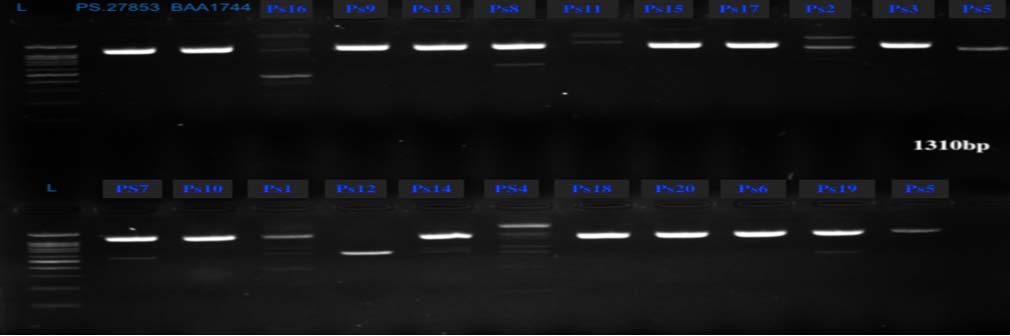
Figure 3. Agarose gel electrophoresis (1% agarose) depicting the algD gene (1310 bp) and DNA ladder (lane M, 1 kbp Plus [Thermo Fisher]). Lanes: Ps27853 and BAA1774, represent positive controls (BAA1774: Pseudomonas aeruginosa ATCC BAA1774, Ps27853: Pseudomonas aeruginosa ATCC 27853). Lanes: Ps1 to Ps20: represent Pseudomonas aeruginosa isolates

Figure 4. Agarose gel electrophoresis (1% agarose) depicting the toxA gene (352 bp) and DNA ladder (lane M, 1 kbp plus [Thermo Fisher]). Lanes: Ps27853 and BAA1774, represent positive controls (BAA1774: Pseudomonas aeruginosa ATCC BAA1774, Ps27853: Pseudomonas aeruginosa ATCC 27853). Lanes: Ps1 to Ps20: represent Pseudomonas aeruginosa isolates

Figure 5. Agarose gel electrophoresis (1% agarose) depicting the phzS gene (1752 bp) and DNA ladder (lane M, 1 kbp plus [Thermo Fisher]). Lanes: Ps27853 and BAA1774, represent positive controls (BAA1774: Pseudomonas aeruginosa ATCC BAA1774, Ps27853: Pseudomonas aeruginosa ATCC 27853). Lanes: Ps1-Ps20: represent Pseudomonas aeruginosa isolates
Association between resistance phenotypes, resistance markers and virulence factors Pseudomonas aeruginosa
The association between resistant phenotypes, resistance markers and virulence factors is depicted in Table 6. A significant association (P = 0.003) was found between ESBL production and MDR/XDR phenotypes, suggesting that ESBL-positive Pseudomonas aeruginosa are more likely to exhibit MDR/XDR pattern (Table 6). Similarly, MDR/XDR phenotypes were significantly associated (P = 0.001) with high MARI values (more than 0.2), indicating that isolates originate from environments with sustained antibiotic selective pressure are more likely to exhibit MDR/XDR phenotypes (Table 6). In contrast, no association was found between MDR/XDR phenotypes, biofilm formation and the presence of virulence-encoding genes (Table 6).
Table (6):
Association between resistant phenotypes, resistance markers and virulence factors in Pseudomonas aeruginosa
| Variable | Category | MDR/XDR (n = 13) | Non-MDR (n = 7) | P-value (Fisher’s exact)† |
|---|---|---|---|---|
| ESBL phenotypes | Positive | 10 | 0 | 0.003* |
| negative | 3 | 7 | ||
| MARI category | High risk > 0.2 | 13 | 1 | 0.001* |
| Low risk < 0.2 | 0 | 6 | ||
| Biofilm formation | Strong | 1 | 0 | 1.000 |
| Weak | 12 | 7 | ||
| toxA gene | Present | 9 | 3 | 0.356 |
| Absent | 4 | 4 | ||
| phzS gene | Present | 9 | 6 | 0.613 |
| Absent | 4 | 1 | ||
| algD gene | Present | 11 | 5 | 0.587 |
| Absent | 2 | 2 |
ESBL = Extended-spectrum beta-lactamase, MDR = Multidrug-resistance, XDR = Extensively drug-resistance, MARI = Multiple antibiotic resistance index, *significant association. †Fisher’s exact P-value < 0.005 is considered significant. Significant association suggests that phenotypic resistance markers such as ESBL production and MARI are strongly associated with MDR/XDR patterns rather than virulence determinants and biofilm-forming capacity.
Pseudomonas aeruginosa infections are more frequent in immunocompromised patients. This study elaborated on Pseudomonas aeruginosa incidence in hospitalized patient samples (wound swabs and urine) from multiple health centers in Makkah City, Saudi Arabia. The data revealed an overall low Pseudomonas aeruginosa prevalence (4.63%, n = 432) in both types of samples. The occurrence of Pseudomonas aeruginosa isolates (90%, n = 20) was significantly higher in wound swabs, whereas urine samples contained only two isolates. Thus, its clinical prevalence could vary among infection sites. For example, hospitalized pneumonia patients in the United States presented a higher Pseudomonas aeruginosa infection rate (16.2%) in a study. The same study revealed that, 5.7% of surgical site infections in the USA were associated with Pseudomonas aeruginosa from 2011-2014. Similarly, England’s data from 2000-2013 linked 3.4%-6.5% of surgical site infections to Pseudomonas spp. Pseudomonas aeruginosa is also an established source of 10% of UTI cases in the USA.11 Al-Shammari et al.18 reported that 32.4% of wound infections were caused by Pseudomonas aeruginosa in Riyadh during 2019-2021. Contrarily, they noted a considerably low occurrence (4.6%) of UTI-associated Pseudomonas aeruginosa infections. Another study investigated the incidence of Pseudomonas aeruginosa in seven hospitals in seven different Saudi Arabian administrative regions from 2022-2023. They aligned the Pseudomonas aeruginosa infections from higher to lower as respiratory infection (34.1%)>UTI (21.1%)>skin/soft tissue infections (17.8 %).20 Similarly, Momenah et al.17 reported lower Pseudomonas aeruginosa prevalence in clinical specimens from a tertiary hospital (2011-2021) in Makkah, Western Saudi Arabia. They noted a significant decline in Pseudomonas aeruginosa prevalence from 9.8% in 2011 to 5.2% in 2021. Overall Pseudomonas aeruginosa prevalence was recorded as 7.1%-7.3% among healthcare-related infections.11 Shrestha et al.32 have also reported a low Pseudomonas aeruginosa (5.1%) occurrence in clinical specimens from a teaching hospital in Kathmandu, Nepal. Similarly, a low Pseudomonas aeruginosa detection during this study could be associated with its opportunistic nature as only immunocompromised patients appeared as the primary target.32 Thus, the findings of this study coincide with the previous global11,32 and Saudi Arabian reports.17-20
Antibiotic overuse in medical, veterinary, and agricultural applications has globally raised the antimicrobial resistance (AMR) phenomenon.33 The prevalence of MDR, XDR, and PDR Pseudomonas aeruginosa isolates in nosocomial infections is increasingly reported worldwide.34 During this study, 13 out of 20 Pseudomonas aeruginosa isolates demonstrated MDR patterns. Five of the 20 Pseudomonas aeruginosa isolates emerged as XDR isolates. MDR Pseudomonas aeruginosa isolates have been reported in Saudi Arabia16,19,35 with a high frequency of XDR,16,19,35 PDR,19,20,35 and DTR (difficult to treat) isolates.20 The rising levels of clinical MDR, PDR, and XDR Pseudomonas aeruginosa isolates have been reported in Spain,36 Ethiopia,34 United States,12 Italy and Greece,37 North Africa, (Morocco and Egypt),15 and Middle East (Iraq, Bahrain, UAE, Qatar, Lebanon, Kuwait, and Oman).19
Hospital-acquired MDR, XDR, and PDR Pseudomonas aeruginosa infections are becoming DTR (difficult-to-treat) illnesses.2,20 It could be attributed to the intrinsic resistance of Pseudomonas aeruginosa against various antibiotic groups (cephalosporin and beta-lactam). Moreover, it could acquire resistance mechanisms against other antimicrobial agent classes including aminoglycoside, beta-lactam, and fluoroquinolones combinations through mutations, biofilms, adaptive resistance, and horizontal transfer of genes.10 This pathogen can survive in diverse environments by involving quorum sensing, enzyme promiscuity, and biofilm formation. The genomic plasticity of Pseudomonas aeruginosa facilitates its survivability and growth in hostile environments including the effects of antibiotics.2 The high MARI value during this study suggests that most Pseudomonas aeruginosa isolates originated from the areas of extensive antibiotic administrations.
Aminoglycosides (tobramycin and amikacin), fluoroquinolones (levofloxacin and ciprofloxacin), and carbapenems (meropenem) are effective drugs against Pseudomonas aeruginosa infections.36 Pseudomonas aeruginosa isolates (15%-20%) presented a low resistance profile to aminoglycosides during this study, which is in agreement with previous reports in Saudi Arabia. Thabit et al.20 have reported the highest Pseudomonas aeruginosa susceptibility (76.8% of isolates) to amikacin, whereas these isolates were moderately susceptible to tobramycin (57.8%) and gentamicin (57.3%). Hafiz et al.16 also noted amikacin susceptibility in 92.6% of Pseudomonas aeruginosa isolates (n = 379) in Riyadh. A study in the USA demonstrated lower Pseudomonas aeruginosa resistance to tobramycin (2.3%, n = 258) and amikacin (8.1%, n = 258).12 Similar amikacin susceptibility patterns of Pseudomonas aeruginosa isolates have been reported in Oman, Bahrain, and Kuwait.19 Contrarily, samples from Intensive Care Unit (ICU) patients from Middle Eastern and North African countries including Saudi Arabia have revealed higher Pseudomonas aeruginosa resistance against aminoglycosides. This could be attributed to frequent and less effective antibiotic prescriptions in ICUs, and the presence of highly resistant clones in these hospital sections.15
Pseudomonas aeruginosa isolates of wound swabs demonstrated a lower resistance to carbapenems [meropenem (30%) and imipenem (25%)] during this study. However, the prevalence of carbapenem-resistant Pseudomonas aeruginosa is increasingly reported in Saudi Arabia. Al-Tawfiq14 examined 2679 Pseudomonas aeruginosa isolates of various departments (outpatient and inpatient) from 1998-2003, in Dhahran, Saudi Arabia. They observed a rise in imipenem-resistant Pseudomonas aeruginosa isolates in community-acquired and nosocomial infections. Nosocomial infections of imipenem-resistant Pseudomonas aeruginosa increased from 1.4%-11%, whereas the rise in community-acquired infections was noted as 2%-3.2% during the study period.14 A recent study in Riyadh has revealed that carbapenem-resistant (imipenem) Pseudomonas aeruginosa contributed to 29.5% (n = 3579) infections in hospitalized patients from 2019-2021.16 Al-Shammari et al.18 investigated 1024 Pseudomonas aeruginosa isolates (2019-2021) in a tertiary hospital in Riyadh and reported a significant rise in meropenem-resistant (61%) and imipenem-resistant (77.8%) isolates. The increased prevalence of carbapenem-resistant Pseudomonas aeruginosa is well-documented in various countries. Zhao et al.38 studied 416 Pseudomonas aeruginosa isolates (2017-2020) from ten hospitals in Guangdong, China, and observed considerably high meropenem (79.4%) and imipenem (89.4%) resistance. The rising trends of carbapenem-resistant Pseudomonas aeruginosa isolates have been reported in Iran,39 Brazil,40 and Ethiopia.41 Multiple mechanisms contribute to carbapenem-resistance in Pseudomonas aeruginosa, such as (i) intrinsic alleviation of carbapenem uptake-related porine protein; (ii) efflux pump overexpression in resistance nodulation division; (iii) overproduction of chromosomal AmpC cephalosporinase; and (iv) chromosomal, integron- and transposon-mediated, and plasmid-encoded carbapenemase production.42 High mortality rates in carbapenem-resistant Pseudomonas aeruginosa-infected patients make this pathogen a global health concern.40,43 Moreover, it causes longer hospital stays, which results in high hospitalization costs and a burden on the healthcare systems.40
Fluoroquinolones are often prescribed to treat Pseudomonas aeruginosa infections. During this study, high resistance of Pseudomonas aeruginosa was noted against fluoroquinolones [levofloxacin (55%) and ciprofloxacin (45%)], which is in line with previous Saudi Arabian and global reports.20,44 Contrarily, several local and global investigations have reported lower Pseudomonas aeruginosa resistance to fluoroquinolones.15,18,45 The resistance to fluoroquinolones is most likely mediated by chromosomes rather than transferable mobile genetic elements.13,44 Diaz et al.36 stated that patients with a previous record of anti-Pseudomonas aeruginosa antibiotic administration could experience cross-resistance to other antibiotics. For example, ceftazidime-treated patients developed a cross-resistance against cefepime-aztreonam and piperacillin-tazobactam.36 Yang et al.12 have also reported its correlated resistance to aztreonam, cefepime, and piperacillin-tazobactam. For instance, 90% of cefepime-resistant Pseudomonas aeruginosa isolates also demonstrated resistance to aztreonam, whereas 75% of isolates exhibited piperacillin-tazobactam resistance. This phenomenon explains the detection of high resistance rates against cefepime (35%), ceftazidime (55%), piperacillin-tazobactam (60%), and aztreonam (100%) in this study. Therefore, XDR and MDR Pseudomonas aeruginosa infection treatments are a complicated task, which requires novel therapeutic options and strict infection control strategies.
Multiple investigations have elaborated on Pseudomonas aeruginosa virulence, but still further elucidation is mandatory. Pseudomonas aeruginosa pathogenicity is mainly based on the secretion of exoenzyme S and exotoxin A, pyocyanin production, and biofilm synthesis.3 This study analyzed key Pseudomonas aeruginosa virulence determinants such as biofilm formation and pyocyanin and exotoxin A-encoding genes (phzS, toxA). All the Pseudomonas aeruginosa isolates displayed biofilm-forming capability. However, biofilm formation was weak in most isolates except the Ps18 isolate from wound infections, which demonstrated strong biofilm synthesis efficiency. Khan et al.46 followed the crystal violet staining approach and reported weaker biofilm formation among Pseudomonas aeruginosa isolates (48.1%, n = 81) from hospital wastewater. Similarly, approximately 50% (n = 80) of clinical Pseudomonas aeruginosa isolates presented weak or non-biofilm formation properties during a study in Iran.47 Contrarily, other crystal violet technique-based studies have reported clinical Pseudomonas aeruginosa isolates’ strong biofilm-forming capabilities.48 These variations could be attributed to the modifications in the crystal violet assay. Different broth media for overnight isolates’ growth (Luria Bertani broth, nutrient broth, brain heart infusion broth, and Mueller-Hinton broth) might also impact the results. Ozyaman and Yilmaz49 evaluated the effects of various broth media (Mueller-Hinton broth, tryptic soy broth, brain heart infusion broth, and Luria Bertani broth) on Pseudomonas aeruginosa biofilm formation. They deduced that biofilm formation varied with overnight culturing of Pseudomonas aeruginosa in different growth media.
Development of biofilms has frequently been suggested as an important contributor to the increased prevalence of MDR Pseudomonas aeruginosa isolates.7 Nevertheless, in the present study, the Pearson correlation analysis (r = -0.1397) did not establish any significant association between MDR Pseudomonas aeruginosa and their biofilm-forming capability. Gajdacs et al.50 have reported similar findings based on the Pearson correlation analysis of the data. Kunwar et al.51 also did not observe any correlation between antibiotic resistance and Pseudomonas aeruginosa isolates from burn patients. Contrarily, Abdulhaq et al.52 have strongly correlated the biofilm formation and MDR in clinical Pseudomonas aeruginosa. They reported significantly higher resistance levels among biofilm-producing strains compared with others. Similarly, another study has also correlated MDR profiles with biofilm formation in Pseudomonas aeruginosa isolates from wound infection.53 Therefore, drawing definitive conclusions about the role of biofilm formation in MDR profiles of clinical Pseudomonas aeruginosa is complicated. Biofilm formation was weak in most Pseudomonas aeruginosa isolates. However, these isolates carried an alginate production-related algD gene that contributes to the formation and maturation of biofilms.54 However, the algD gene was not detected in all identified Pseudomonas aeruginosa isolates depicting that biofilm formation does not solely depend on this gene.55 Other exopolysaccharide production-related genes (psl and pel) in Pseudomonas aeruginosa also contribute to biofilm formation in clinical Pseudomonas aeruginosa.56 Furthermore, multiple factors could contribute to Pseudomonas aeruginosa biofilm production including biofilm-related genes and their expression rates, oxygen levels, nutrients, temperature, and pH levels.54
Pyocyanin and exotoxin-A are established virulence factors of Pseudomonas aeruginosa.57 The toxA gene-encoded exotoxin-A restricts protein synthesis, and damages Pseudomonas aeruginosa-associated soft skin, wound, and burn infections.58,59 The prevalence of the toxA gene varies among clinical Pseudomonas aeruginosa isolates of different geographic locations. During a study in Egypt, Edward et al.60 detected the toxA gene in 46.2% of clinical Pseudomonas aeruginosa isolates. Other studies have speculated the occurrence of the toxA gene in >80% of nosocomial Pseudomonas aeruginosa isolates,58,59 which is in line with the findings of this study. The phzS gene encodes the production of a blue pigment (pyocyanin) that causes tissue injury through ROS (reactive oxygen species) generation, and neutrophil apoptosis. This virulence factor contributes to Pseudomonas aeruginosa nosocomial infections’ severity including immunocompromised patients’ respiratory infections.57,61 The data of this study revealed the presence of both genes (toxA and phzS) in detected Pseudomonas aeruginosa isolates, which confirmed their high virulence. Multiple studies have reported toxA and phzS genes in clinical Pseudomonas aeruginosa isolates. These genes add to the infection’s severity in MDR isolates.60,62-64
Study limitations
The prevalence of Pseudomonas aeruginosa was low (20 isolates) despite the examination of 432 wound swabs and urine samples; this low prevalence may lower the power of the statistical analysis. Generalizability would have been improved by a large number of isolates from a variety of clinical samples. Given the known inter-hospital diversity in resistance profiles and infection control practices, the single center study (one hospital) conducted over a brief period may not accurately reflect regional or national epidemiological patterns. The genetic basis of antimicrobial resistance, such as carbapenemase genes (e.g. NDM, VIM, IMP, OXA), extended-spectrum beta-lactamase-associated genes, porin mutations of mobile genetic elements, was not investigated in this work; instead, resistance to antibiotics was determined phenotypically, this may hinder a more thorough comprehension of the molecular mechanisms underlying MDR and XDR patterns. Considering that the authors funded the entire research project, these study limitations were primarily caused by a lack of financing.
The study highlights the rapid rise in MDR, and XDR Pseudomonas aeruginosa isolates in the urinary tract and wound infections in Makkah, Western Saudi Arabia. Overall, a low Pseudomonas aeruginosa prevalence was noted. However, high antibiotic resistance, particularly against fluoroquinolones, aztreonam, and carbapenems could complicate the treatment regimen. The presence of virulence factors encoding genes (toxA, phzS, and algD) further accentuates the pathogenicity of these isolates. Despite the carriage of the algD gene in most isolates, weak biofilm formation was noticed, which demands for detailed exploration of biofilm-influencing factors. The absence of a significant correlation between MDR and biofilm formation during this study also requires in-depth elaboration on their interplay. The results necessitate robust infection control strategies, antibiotic stewardship programs, and continuous molecular investigations for effective restriction of resistant virulent strains. The remedial measures are crucial for the public health and availability of efficient Pseudomonas aeruginosa treatment options.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
HHA and LAN conceptualized the study. HHA, HAM, NAO and KE applied methodology. KE collected resources. HSA, HHA, LAN, NAO, MHA and MMSA performed Investigation. HSA, HHA, KE and HAM performed data curation. HSA, HAM, LAN and KE performed formal analysis. HSG, HHA and IA wrote the original draft. HHA, LAN and IA wrote, reviewed and revised the manuscript. HAM, LAN and HHA supervised the study. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study was approved by the Research Ethics Committee, Department of Biology, Faculty of Science (Approval number 342180214442/89647) on 20 October 2021.
- Crone S, Vives-Florez M, Kvich L, et al. The environmental occurrence of Pseudomonas aeruginosa. APMIS. 2020;128(3):220-231.
Crossref - Pachori P, Gothalwal R, Gandhi, P. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit: a critical review. Gene Dis. 2019;6(2):109-119.
Crossref - Sendra E, Fernandez-Munoz A, Zamorano L, et al. Impact of multidrug-resistance on the virulence and fitness of Pseudomonas aeruginosa: a microbiological and clinical perspective. Infection. 2024;52(4):1235-1268.
Crossref - Qin S, Xiao W, Zhou C, et al. Pseudomonas aeruginosa: pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct Target Ther. 2022;7(1):199.
Crossref - Ramachandran G. Gram-positive and Gram-negative bacterial toxins in sepsis. Virulence. 2013;5(1):213-218.
Crossref - Jabtonska J, Augustyniak A, Dubrowska K, Rakoczy R. The two face of pyocyanin – why and how to steer its production? World J Microbiol Biotechnol. 2023;39(4):103.
Crossref - Haidar A, Muazzam A, Nadeem A, et al. Biofilm formation and antibiotic resistance in Pseudomonas aeruginosa. Microbe. 2024;3:100078.
Crossref - Moradali MF, Ghods S, Rehm BHA. Pseudomonas aeruginosa lifestyle:A paradigm for adaptation, survival, and persistence. Front Cell Infect Microbiol. 2017;7:39.
Crossref - Rocha AJ, de Oliveira Barsottini MR, Rocha RR, Laurindo MV, de Moraes FLL, da Rocha SL. Pseudomonas aeruginosa: virulence factors and antibiotic resistance genes. Braz Arch Biol Technol. 2019;62:e19180503.
Crossref - Pong Z, Raudonis R, Glick BR, Lin TJ, Cheng Z. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol Adv. 2019;37(1):177-192.
Crossref - Reynolds D, Kollef M. The epidemiology and pathogenesis and treatment of Pseudomonas aeruginosa infections:an update. Drugs. 2021;81(18):2117-2131.
Crossref - Yang AF, Huang V, Samaroo-Campbell J, Augenbraun M. Multi-drug resistant Pseudomonas aeruginosa: a 2019-2020 single center retrospective case control study. Infect Prevent Pract. 2023;5(3):100296.
Crossref - Zhao L, Wang S, Li X, He X, Jian L. Development of in vitro resistance to fluoroquinolones in Pseudomonas aeruginosa. Antimicrob Resist Infect Control. 2020;9:124.
Crossref - Al-Tawfiq JA. Occurrence and antimicrobial resistance patterns of inpatient and outpatient isolates of Pseudomonas aeruginosa in Saudi Arabia hospital:1998-2003. Int J Infect Dis. 2007;11(2):109-114.
- Al-Orphaly M, Hadi HA, Eltayed FK, et al. Epidemiology of Multidrug-resistant Pseudomonas aeruginosa in the Middle East and North Africa Region. mSphere. 2021;6(3):10.1128.
Crossref - Hafiz TA, Bin Essa EA, Alharbi SR, et al. Epidemiological, microbiological, and clinical characteristics of multi-resistant Pseudomonas aeruginosa isolates in King Fahad Medical City, Riyadh, Saudi Arabia. Trop Med Infect Dis. 2023;8(4):205.
Crossref - Momenah AM, Bakri RA, Jalal NA, et al. Antimicrobial resistance pattern of Pseudomonas aeruginosa:an 11-year experience in a tertiary care hospital in Makkah, Saudi Arabia. Infect Drug Resist. 2023;16:4113-4122.
Crossref - Al-Shammari HO, Somily A, Qattan MY, Alsubki RA, Moussa IM. Susceptibility pattern of multi-drug resistance Pseudomonas aeruginosa isolates from tertiary care hospital in Riyadh, KSA. J King Saud Univ Sci. 2023;35:102702.
Crossref - Alatoom A, Alattas M, Alraddadi B, et al. Antimicrobial resistance profiles of Pseudomonas aeruginosa in the Arabia Gulf region over a 12-year period (2010-2021). J Epidemiol Glob Health. 2024;14:529-548.
Crossref - Thabit AK, Algh00amdi AM, Miaji MY, et al. Antibiotic susceptibility of Pseudomonas aeruginosa in Saudi Arabia:a national antimicrobial resistance surveillance study. Front Public Health. 2024;12:1436648.
Crossref - Tryfinopouloi P, Drosinos EH, Nychas GJ. Performance of Pseudomonas CFC-selective medium in the fish storage ecosystems. J Microbiol Methods. 2001;47(2):243-247.
Crossref - Bauer AW, Kirby WM, Sherris JC, Turck M. Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol. 1966;45(4):493-496
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 34th ed. CLSI supplement M100, March 2024. Clinical and Laboratory Standards Institute, USA.
- Peyroux J, Almahmoudh I, Prebe-Coquerel E, Girard T, Maurin M, Caspar Y. Rapid and automated screening of carbapenemase-and ESBL-producing Gram-negative bacteria from rectal swabs using chromogenic agar media and the ScanStation device. Microbiol Spectr. 2023;11(5):e02723-23.
Crossref - Khan HA, Neyaz LA, Malak HA, et al. Diversity and antimicrobial susceptibility patterns of clinical and environmental Salmonella enterica serovars in Western Saudi Arabia. Folia Microbiol. 2024;69(6):1305-1317.
Crossref - Al-Ghamdi HS, Malak HA, Neyaz LA, et al. Prevalence of extensively and pan-drug-resistant Acinetobacter spp. In nosocomial infections in Western Saudi Arabia. J Pure Appl Microbiol. 2024;18(2):886-998.
Crossref - Naves P, del Prado G, Huelves L, et al. Measurements of biofilm formation by clinical isolates of Escherichia coli is a method-dependent. J Appl Microbiol. 2008;105(2):585-590.
Crossref - Arafa SH, Alshehri WA, Organji SR, et al. Antimicrobial resistance, virulence factors-encoding genes, and biofilm-forming ability of community associated uropathogenic Escherichia coli in western Saudi Arabia. Polish J Microbiol. 2022;71(3):325-339.
Crossref - Adenipekun EO, Akinleye EF, Tewogbade OA, Iwalokun BA. Detection of virulence genes and multidrug resistance in Pseudomonas aeruginosa clinical isolates from a public hospital in Lagos, Nigeria. Sci Afr. 2023;22:e01950.
Crossref - Qader GM, Jaje0000000000es KK, Jarjees RK. Molecular detection of metallo-beta-lactamase and alginate in multidrug resistance Pseudomonas aeruginosa isolated from clinical specimen. J Med Life. 2022;15(9):1105-1109.
Crossref - Fazeli N, Momtaz H. Virulence gene profiles of multidrug-resistant Pseudomonas aeruginosa isolated from Iranian hospital infections. Iran Red Crescent Med J. 2014;16(10):e15722.
Crossref - Shrestha S, Amatya R, Adhikari RP. Prevalence and antibioiogram of Pseudomonas aeruginosa isolated from clinical specimens in a teaching hospital, Kathmandu. Int J Infect Dis 2016;45(Suppl 1):115-116
Crossref - Samreen, Ahmad I, Malak HA, Abulreesh HH. Environmental antimicrobial resistance and its drivers:a potential threat to public health. J Glob Antimicrob Resist. 2021;27:101-111.
Crossref - Addis T, Araya S, Desta K. Occurrence of multiple, extensive and pan drug-resistant Pseudomonas aeruginosa and carbapenemase production from presumptive isolates stored in Biobank at Ethiopian public health. Infect Drug Resist. 2021;14:3609-3618.
Crossref - Almakrami M, Salmen M, Aldashel YA, et al. Prevalence of multidrug-, extensively drug-, and pandrug-resistant bacteria in clinical isolates from King Khaled Hospital, Najran, Saudi Arabia. Discov Med. 2024;1:108.
Crossref - Diaz Santos E, Mora Jimenez C, del Rio-Carbajo L, Vidal-Cortes P. Treatment of severe multi-drug resistant Pseudomonas aeruginosa infections. Med Intensiva. 2022;46(9):508-520.
Crossref - Perez A, Gato E, Perez-Llarena J, et al. High incidence of MDR and XDR Pseudomonas aeruginosa isolates obtained from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J Antimicrob Chemother. 2019;47(5):1244-1252.
Crossref - Zhao Y, Chen D, Chen K, et al. Epidemiological and genetic characteristics of clinical carbapenem-resistant Pseudomonas aeruginosa strains in Guangdong Province, China. Microbiol Spectr. 2023;11(3):e04261-22.
Crossref - Rostami S, Shiekh AF, Shoja S, et al. Investigating of four main carbapenem-resistance mechanisms in high-level carbapenem resistant Pseudomonas aeruginosa isolated from burn patients. J Chin Med Assoc. 2018;81(2):127-132.
Crossref - de Souza GHdA, Rossato L, Brito GT, Bet GMdS, Simionatto S. Carbapenem-resistant Pseudomonas aeruginosa strains:a worrying health problem in intensive care units. Rev Inst Med Trop Sao Paulo. 2021;63:e71.
Crossref - Abdeta A, Negeri AA, Beyene D, et al. Prevalence and trends of carbapenem-resistant Pseudomonas aeruginosa and Acinetobacter species isolated from clinical specimens at the Ethiopian Public Health Institute, Addis Ababa, Ethiopia:a retrospective analysis. Infect Drug Resist. 2023;16:1381-1390.
Crossref - Halat DH, Moubareck CA. The intriguing carbapenemases of Pseudomonas aeruginosa:current status, genetic profile, and global epidemiology. Yale J Biol Med. 2022;95(4):507-515.
- Reyes J, Komarow L, Chen L, et al. Global epidemiology and clinical outcomes of carbapenem-resistant Pseudomonas aeruginosa and associated carbapenemase (POP):a prospective cohort study. Lancet Microbe. 2023;4(3):e159-e170.
Crossref - Yang, X, Xing, B, Liang, C, Ye Z, Zhang, Y. Prevalence and fluoroquinolone resistance of Pseudomonas aeruginosa in a hospital of South China. Int J Clin Exp Med. 2015;8(1):1386-1390.
- Khater ES, Abdo KHM. Detection of carbapenem-resistant Pseudomonas aeruginosa in a tertiary care hospital in Saudi Arabia. Microbe Infect Dis. 2022;3(3):693-702.
Crossref - Khan MAS, Islam Z, Shah SMT, Rahman SR. Characterization of biofilm-formation and multi-drug resistance among Pseudomonas aeruginosa isolated from hospital wastewater in Dhaka, Bangladesh. J Water Health. 2024;22(5):825-834.
Crossref - Kamali E, Jamali A, Ardebilli A, Ezadi E, Mohebbi A. Evaluation of antimicrobial resistance, biofilm forming potential, and the presence of biofilm-related genes among clinical Pseudomonas aeruginosa. BMC Res Notes. 2020;13(1):27.
Crossref - Abdelraheem WM, Abdelkader AE, Mohamed ES, Mohammed MS. Detection of biofilm formation and assessment of biofilm genes expression in different Pseudomonas aeruginosa clinical isolates. Meta Gene. 2020;23:100646.
Crossref - Ozyaman, F, Yilmaz, O. Effect of different culture media on Pseudomonas aeruginosa biofilm formation. Turk J Health Sci Life. 2023;6(3):133-140.
Crossref - Gajdacs M, Barath Z, Karpati K, et al. No correlation between biofilm formation, virulence factors, and antibiotic resistance in Pseudomonas aeruginosa:results from a laboratory-based in vitro study. Antibiotics. 2021;10(9):1134.
Crossref - Kunwar A, Shrestha P, Shrestha S, Thapa S, Shrestha S, Amatya NM. Detection of biofilm formation among Pseudomonas aeruginosa isolated from burn patients. Burns Open. 2021;5(3):125-129.
Crossref - Abdulhaq N, Nawaz Z, Zahoor MA, Siddique AB. Association of biofilm formation and multi-drug resistance in clinical isolates of Pseudomonas aeruginosa. EXCLI Journal. 2020;19:201-208.
Crossref - Chimi LY, Noubom M, Bisso BN, Njateng GSS, Dzoyem JP. Biofilm formation, pyocyanin production, and antibiotic resistance profile of Pseudomonas aeruginosa isolates from wounds. Int J Microbiol. 2024;2024:1207536.
Crossref - El-sayed HA, Fahmy YA. Correlation between biofilm formation and multidrug resistance in clinical isolates of Pseudomonas aeruginosa. Microbe Infect Dis. 2021;2:541-549.
Crossref - Thi MTT, Wibowo D, Rehm BHA. Pseudomonas aeruginosa biofilms. Int J Molec Sci. 2020;21(22):8671.
Crossref - Tuon FF, Dantas LR, Suss PH, Ribeiro VST. Pathogenesis of Pseudomonas aeruginosa biofilm:a review. Pathogens. 2022;11(3):300.
Crossref - Jurado-Martin I, Sainz-Mejias M, McClean S. Pseudomonas aeruginosa:a audacious pathogen with an adaptable arsenal of virulence factors. Int J Molec Sci. 2021;22(6):3128.
Crossref - El-Din AB, Elnagdy M, Badr R, El-Sabagh A. Pseudomonas aeruginosa exotoxin A: its role in burn wound infection and wound healing. Egypt J Plast Reconstr Surg. 2008;32:59-65.
- Javanmardi F, Emami A, Pirbonyeh N, Keshvarzi A, Rajaee M. A systematic review and meta-analysis on exo-toxins prevalence in hospital acquired Pseudomonas aeruginosa isolates. Infect Genet Evol. 2019;75:104037.
Crossref - Edward EA, El Shehawy MR, Abouelfetouh A, Aboulmagd E. Prevalence of different virulence factors and their association with antimicrobial resistance among Pseudomonas aeruginosa clinical isolates from Egypt. BMC Microbiol. 2023;23(1):161.
Crossref - Mudaliar SB, Prasad ASB. A biomedical perspective of pyocyanin from Pseudomonas aeruginosa:its applications and challenges. World J Microbiol Biotechnol. 2024;40(3):90.
Crossref - Alabdali YAJ. Detection and association of toxA gene with antibiotic resistance in Pseudomonas aeruginosa strains isolated from sources in Al-Muthanna City. Gene Rep. 2021;25:101358.
Crossref - Naik P, Pandey S, Gagan S, Biswas S, Joseph J. Virulence factors in multidrug (MDR) and pan-drug resistant (XDR) Pseudomonas aeruginosa:a cross sectional study of isolates recovered from ocular infections in high-incidence settings in southern India. J Ophthalmic Inflamm Infect. 2021;11(1):36.
Crossref - Panahi Z, Owrang M, Goli HR. Significant role of pyocyanin and exotoxin A in the pathogenesis of Pseudomonas aeruginosa isolated from hospitalized patients. Folia Med. 2024;66(1):88-96.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.