


Enzymes are biocatalysts, that facilitate chemical reactions by lowering their activation energy. Among these, cellulase emerges as a significant enzyme, consisting of a triad of components that work in synergy to degrade cellulosic biomass. Its significance is mostly pronounced in agricultural contexts, where there is an abundance of lignocellulosic biomass making it pivotal for utilization and conversion of biomass. Utilizing the biomass as a substrate for cellulase production offers dual advantages. Firstly, it simplifies the enzymatic synthesis process by the utilization of naturally occurring precursors. Secondly, it contributes to cost reduction by leveraging readily available resources thereby making it economically viable. Microbial cellulases, sourced from diverse microbes found globally, can aid in efficient enzymatic production. Advances in fermentation processes, coupled with the application of biotechnological tools, have significant impacts in production scalability and cost-effectiveness. Optimizing production strategies is crucial to meet the increasing demands of industrial applications while ensuring sustainability. Emphasizing the utilization of biomass substrates and harnessing the potential of emerging biotechnological advancements are key aspects of enzyme production. This review shall aim to provide an in-depth exploration of current cellulase production technologies and future prospects. By elucidating the underlying principles of cellulase catalysis and the intricacies of production methodologies.
Biomass, Cellulase, Enzyme, Fermentation, Lignocellulose
Enzymes are biological catalysts and are able to catalyse, i.e. speed up biological reactions by means of lowering the activation energy of the reactions that it part takes in. Enzymes have a role in all biological systems and are an imperative part constituting the living world. In the later parts of the 19th and earlier parts of the 20th centuries, notable progress was achieved in the isolation, characterization, and commercial application of various enzymes. However, it wasn’t until the 1920s that the crystallization of enzymes unveiled their association with protein molecules, establishing the proteinaceous nature of catalytic activity. A paradigm shift occurred during the late 20th century, in the 1980s when the discovery was made that certain ribonucleic acid (RNA) molecules, termed ribozymes, possessed catalytic capabilities. These ribozymes were identified as key contributors to gene expression processes. Concurrently, within the same epoch, biochemists developed methodologies for generating antibodies endowed with catalytic properties, colloquially referred to as ‘abzymes’. The emergence of abzymes presented substantial prospects for innovative applications both as catalysts in industrial processes and as therapeutic agents. This transformative period in enzymology expanded the conceptual framework beyond the exclusive protein-centric view, acknowledging the catalytic potential inherent in RNA molecules and engineered antibodies. Consequently, a diversification of enzymatic entities occurred, with ribozymes and abzymes contributing significantly to the understanding and application of catalytic processes.1 There has been a recent shift to use bio-catalysts in order to promote “green chemistry”. This approach has led to many research and development studies that have focused on cost-effective enzyme production. One such enzyme is Cellulase, that degrade cellulose. Cellulose, as studies have suggested is the most abundant biological polymer that is a homopolymer in nature comprising D-glucose units which are connected by β-1, 4 bonds, stands as the pre-eminent organic polymer. Functioning as a crucial structural constituent within the primary cell wall of plants which are green in colour comprising of 50% of its total biomass in dry weight.2 Cellulose holds promise as a renewable natural resource and has wide applications in various industries. Consequently, heightened economic interest surrounds the establishment of systems geared towards way to efficiently manage and utilize cellulose-containing industrial wastes, and presenting it as cost-effective carbon sources.3
Cellulose rich materials have been in use for the production of cellulases, especially using agro-residues such as tea and coffee waste,4 fruit waste, rice bran wastes, etc. as has been reported.5 The utilization of such economical and low-cost substrates poses to be a good alternative for enzyme production. Cellulases, have distinct and diverse activities. The degradation of cellulose rich lignocellulosic biomass. The general division of cellulase is done into exoglucanases (EC 3.2.1.74), endoglucanases (EC 3.2.1.4), cellobiohydrolases (EC 3.2.1.91), and b-glucosidases (EC 3.2.1.21). Exoglucanases exhibit enzymatic activity on either reducing or nonreducing ends of cellulose chains, liberating glucose units. Conversely, endoglucanases catalyze the random hydrolysis of b-1,4-glycosidic bonds within cellulose chains, yielding dextran chains of variable lengths. Cellobiohydrolase specifically cleave glycosidic bonds at nonreducing ends, releasing cellobiose units. These enzymes play a crucial role in the hydrolysis of crystalline cellulose, owing to their processivity. b-glucosidases contribute to the hydrolysis of cellobiose units, thereby converting them into monomeric glucose. The complete hydrolysis of cellulose involves a synergistic effect among these enzymes, manifesting as synergy between endoglucanases and exoglucanases (endo-exo synergy), exoglucanases acting on both reducing and nonreducing ends (exo-exo synergy), synergy between cellobiohydrolases and b-glucosidases, as well as between catalytic and carbohydrate binding domains. Various technologies and substrates have been in use for the production of cellulase. This review seeks to elucidate the microbes used, the current technologies that are being employed and the limitations and future prospects, all pertaining to cellulase production. Figure 1 demonstrates the action of cellulase on cellulose containing materials.
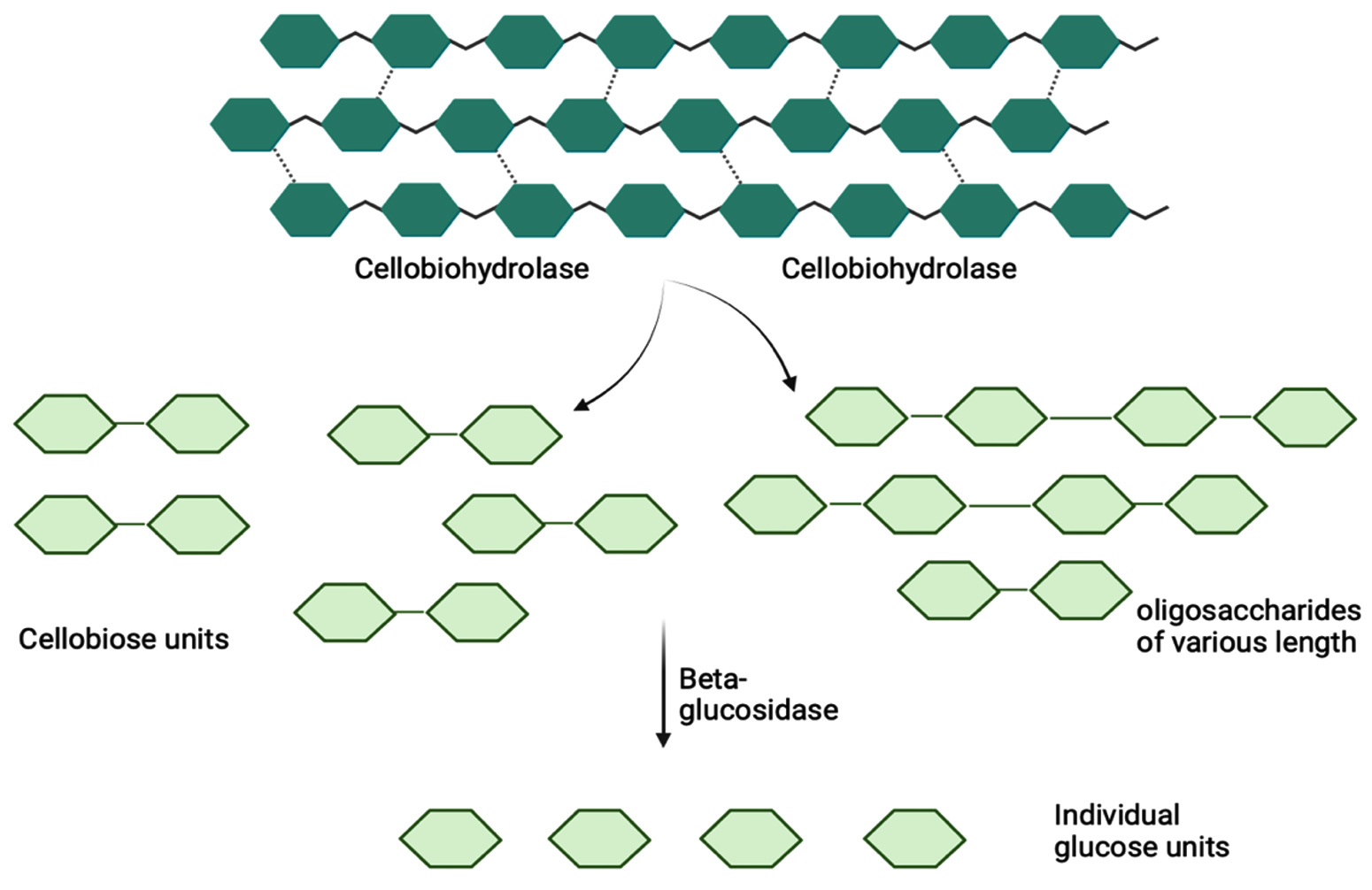
Figure 1. Action of cellulase complex for cellulose hydrolysis
A synergy of three enzymes constituting cellulase complex degrade the lignocellulosic biomass into its constituent monomeric units.
Microbial cellulases
Cellulolytic enzymes, characterized by significant enzymatic activity and stability variations, are predominantly derived from microbial sources. Notably, variations in physical parameters such as pH and temperature contribute to the observed diversity in enzymatic properties. A plethora of microorganisms, including bacteria, protozoans, fungi, etc. have been documented for their capacity to produce cellulases. There is a need for enumeration of such microbes, namely, bacteria, fungi, actinobacteria and yeasts for the potential in production of cellulolytic enzymes. Bacteria and fungi, exhibit proficiency as producers of cellulolytic enzymes, with fungi proving particularly suitable producers of enzymes owing to their extracellular properties. Therefore, the ongoing exploration for novel microorganisms, is imperative to align with industrial requirements. At an industrial scale, various fungal and bacterial strains have been harnessed for cellulase production. Notably, the use of commercial substrates such as cellulose or carboxymethyl cellulose (CMC) has been prevalent, albeit associated with high costs. Microbial cellulases have drawn significant academic interest because they are more economically viable than conventional methods. In contrast, alternative techniques like chemical hydrolysis, physical treatments, thermochemical processes, and solvent-based methods often involve higher costs due to their energy-intensive nature, the use of costly reagents, complex machinery, and negative environmental impacts. As a result, microbial cellulases are seen as a more sustainable and cost-effective option in various industrial applications. Consequently, scientific endeavours have been directed towards substituting these costly substrates with naturally abundant cellulose-rich biomass derived from agricultural and forest residues.6,7
Fungal cellulases
Fungi, formerly regarded as antagonistic microorganisms, have now become an indispensable industrial raw material across diverse applications. The dynamic cellulose decomposers inherent in fungi may be implicated in the global decomposition processes. Additionally, the preferential framework for cellulase synthesis through fungal cellulosomes has been identified as more resilient to environmental variabilities.8 In comparative analyses involving Aspergillus and Humicola species, Trichoderma species have been identified as particularly suitable for cellulase synthesis and industrial utilization.9 The most extensively studied cellulolytic microorganisms encompass white-rot fungi, such as Phanerochaete chrysosporium, and soft-rot fungi, including Fusarium solani, Penicillium funiculosum, Talaromyces emersonii, Trichoderma koningii, and Trichoderma reesei. Prominent among commercially employed microorganisms exhibiting significant cellulolytic capabilities are Aspergillus fumigatus, Aspergillus nidulans, Aspergillus aculeatus, Aspergillus niger, Aspergillus oryzae, and Aspergillus niger;10 Within the Indo-Myanamar Biodiversity Hotspot, three cellulolytic fungi, namely Talaromyces verruculosus SGMNPf3, Trichoderma gamsii SGSPf7, and Trichoderma atroviride SGBMf4, were meticulously identified.11
Bacterial cellulases
Bacterial cellulases have been a part of many industrial applications and there have been many species which have been used for the production of cellulase. Seven bacterial isolates originating from the genera Ochrobactrum, Acinetobacter, Pseudoxanthomonas, Paenibacillus, Stenotrophomonas, and Comamonas were identified in the composting leachate derived from wheat straw.12 The screening of the cellulolytic activity of 398 bacteria isolated from various ecosystems in Algeria (cave, ruins, chott, thermal station, and rhizosphere of arid and semi-arid region). Studies have established that five isolates that belonged to the genus Bacillus. viz, B. velezensis, B. subtilis, B. mojavensis and
B. cereus were able to flourish after being subjected to various stress conditions.13 Specific enzymes produced by microorganisms; however, cellulases of bacterial origin attract more interest due to their natural diversity and ability to inhabit a variety of niches, allowing the selection of cellulolytic strains resistant to environmental stresses. Several reported isolates also belonged to the aforementioned genus with the species being, B. flexus, B. licheniformis and B. paralicheniformis.14 The existence of cellulolytic microorganisms in a variety of habitats is highlighted by the isolation of cellulase-producing bacterial isolates from different settings. These results suggest that favourable environmental factors may aid in initiating the formation of cellulase. Pseudomonas, Paenibacillus, Acidothermus and Micromonospora represent distinctive taxa within cellulolytic species, that make them promising candidates for the exploration of novel enzymes involved in the degradation of polysaccharides, particularly cellulose. These microorganisms highlight the ecosystem as a favourable platform for investigating enzymatic mechanisms associated with cellulose degradation.15
Yeast cellulases
Essentially, only a small number of yeasts exhibit the capacity to break down cellulose. A few of them have been elucidated as of now, with the recent research even forging towards the untouched arena of marine yeast. The first ever report on cellulase producing yeast was by Dennis in 1972,16 where they isolated various strains from the Rhubarb plant out of which nine (out of 12 strains) of Trichosporon cutaneum and 12 out of 13 strains of T. pullulans showed cellulolytic activity. Since then, several new species have been identified that have the potential to degrade cellulose via the production of cellulase such as yeasts obtained from rotten dahlias for the degradation of coconut oil cake belonging to the genus Meyerozyma, named M. guillermondii CBS 2030 which has a relatively high cellulase activity of 102.96 U/mL.2 Naturally, relatively few yeasts have the capacity to break down cellulose. Rhodotorula glutinis, a psychrophile by nature, was reported that purported to be the first of its type. Aureobasidium sp. is the major cellulolytic yeast identified in the habitat of Forcipomyia taiwana, also called the biting midge, according to research on the subject. It has been observed that crude lignocellulosic substrates may be hydrolyzed using yeast strains such as Dendrobium spathilingue, Moesziomyces sp., Candida easanensis, and Saccharomyces species.17 51 species of yeast were recovered from rotting wood; these species were classified into the following genera: Pichia, Candida, Filobasidium, Kluyveromyces, and Aureobasidium. When cultivated on carboxymethyl cellulose or wood chips, only Aureobasidium microstictum and Trichosporon pullulans showed cellulase activity.18
Actinobacterial cellulases
Actinobacteria constitute a taxonomically diverse group of Gram-positive bacteria that predominantly inhabit the soil. They also exhibit a notable capacity for prolific mycelial growth and the formation of chains of aerial conidia similar to that of imperfect fungi, Actinobacteria are characterized by their saprophytic lifestyle. Their presence in various ecological niches makes them one of the most diverse bacterial groups in the natural environment. The physiological traits of Actinobacteria span a wide spectrum, encompassing anaerobic unicellular organisms as well as aerobic, filamentous, and spore-forming lineages. This inherent variability in metabolic and morphological features contributes to the adaptability and ecological success of Actinobacteria across a wide array of habitats. Within the wide sphere of cellulase-producing actinomycetes, considerable attention has been devoted to the investigation of Cellulomonas fimi, Microbispora bispora, and Thermobifida fusca. In particular, Thermobifida fusca emerged as a thermophilic, spore-forming actinomycete species. The genomic constitution of T. fusca comprises 3.6 billion base pairs organized in a singular circular chromosome, hosting 3117 coding sequences. Notably, the genomic composition manifests a G+C content of 67.5%, thereby imparting stability to DNA under conditions of extreme temperature.19 Several studies have proposed the isolation of actinomycetes from marine sediments as a promising strategy for obtaining novel actinomycetal strains capable of yielding potentially valuable new products. Concurrently, actinomycetes are recognized for their proficiency in producing diverse extracellular enzymes, cellulase being one. The proficiency of Streptomyces strains, namely Streptomyces sp. LCJ10A, Streptomyces sp. LCJ11A, Streptomyces sp. LCJ13A, Streptomyces sp. LCJ14A, and Streptomyces sp. LCJ16A, isolated from the Pichavaram mangroves has also been documented.20 These strains were found to exhibit high efficiency in the production of economically significant enzymes, including lipase, cellulase, and asparaginase. The identified enzymes hold substantial potential as valuable assets for innovative biotechnological processes. The overview of the same has been depicted in Table 1.
Table (1):
Bacterial, fungal, actinobacterial and yeast cellulases
Microorganism |
CMC-ase |
𝛃-glucosidase |
Ref. |
|---|---|---|---|
Aspergillus niger |
+ |
+ |
21,22 |
Aspergillus nidulans |
– |
+ |
23 |
Fusarium solani |
+ |
+ |
24 |
Humicola insolens |
+ |
+ |
25 |
Penicillium spp. |
+ |
+ |
26 |
Trichoderma reesei |
+ |
+ |
27,28 |
Neurospora crassa |
+ |
+ |
29 |
Acinetobacter kyonggiensis sp. nov |
– |
+ |
30 |
Acidothermus cellulolyticus |
+ |
+ |
31 |
Paenibacillus terrae |
+ |
+ |
32,33 |
Bacillus subtilis |
+ |
+ |
34 |
Fibrobacter succinogenes |
+ |
+ |
35 |
Clostridium acetobutylium |
+ |
+ |
36 |
Thermomonospora fusca |
+ |
+ |
37 |
Streptomyces thermodiastaticus |
+ |
+ |
37 |
Streptomyces globosus |
+ |
– |
38 |
Nocardiopsis aegyptia |
+ |
– |
38 |
Cellulomonas fimi |
+ |
– |
39 |
Ambrosiozyma sp. |
+ |
+ |
40 |
Aureobasidium pullulans |
+ |
+ |
41,42 |
Brettanomyces intermedius |
– |
+ |
43 |
Candida homilentoma |
+ |
+ |
40,40 |
Candida kashinagacola |
+ |
+ |
40 |
Candida parapsilosis |
+ |
43 |
|
Debaryomyces hansenii |
+ |
+ |
44,45 |
Hanseniaspora osmophila |
+ |
46 |
|
Hanseniaspora uvarum |
+ |
+ |
46,47 |
Lachancea thermotolerans |
+ |
+ |
46,48 |
Metschnikowia fructicola |
– |
+ |
46 |
Metschnikowia pulcherrima |
+ |
+ |
46,48 |
Metschnikowia viticola |
+ |
46 |
|
Meyerozyma guilliermondii |
+ |
+ |
40,43,44,46,49,50 |
Saccharomyces cerevisiae |
– |
+ |
43 |
+ : Production reported. , – : Not reported.
Current technologies in microbial cellulase production
Predominantly, two fermentation techniques are utilized for the production of cellulase, viz. submerged fermentation (SmF) and solid-state fermentation (SSF). In SmF, microorganisms, predominantly fungi are cultivated in a liquid medium containing cellulose or other cellulosic substrates as the primary carbon source. This method allows meticulous control over crucial parameters including pH, temperature, and nutrient availability, facilitates the obtainment of elevated yields of cellulase enzymes. In contrast, SSF involves the utilization of a solid substrate, typically agro-residues, in place of a liquid medium.
In SSF, solid substrates such as bagasse, bran, paper pulp, and rice straw are typically employed, facilitating the gradual metabolism of the substrate by microorganisms.51,52 This technique is particularly advantageous for fungi (with the exception of yeasts) and other microorganisms that necessitate diminished moisture conditions.53 Nonetheless, SSF encounters several challenges including the precise regulation of environmental parameters, scalability, heat dissipation, and mass transfer efficiency. Despite these obstacles, SSF presents benefits such as enhanced yield, cost-effectiveness, and a more streamlined operational process. Conversely, SmF entails the cultivation of microorganisms on either soluble or insoluble substrates, which are either dissolved or submerged within liquid media.54,55 Examples of liquid media comprise domestic wastewater and hydrolysate derived from grape pomace54. SmF facilitates uniform substrate metabolism and affords manageable control over external conditions. This approach is more apt for bacteria, yeasts, and other microorganisms that require elevated moisture levels. While SmF simplifies handling processes and improves heat transfer dynamics, it also poses challenges such as the viscosity of the culture broth, which can result in foaming and incomplete saccharification of lignocellulosic biomass. Ultimately, the enhanced ease of handling and the regulated external conditions render SmF a more efficient and manageable fermentation process.
Active research endeavours are directed towards the refinement and optimization of both fermentation methodologies with the aim of enhancing cellulase production efficiency and mitigating associated costs, thereby enhancing accessibility for diverse industrial application.55 A brief comparative overview of the fermentation technologies has been highlighted in Table 2.
Table (2):
Comparative overview of fermentation technologies employed for cellulase production
Feature(s) |
Solid state fermentation |
Submerged fermentation |
Ref. |
|---|---|---|---|
Substrate |
Solid substrates are generally used. Example: Bagasse, bran, paper pulp, rice straw, etc. |
Microbes are grown on soluble (xylan, pectin, mannan) or insoluble substrates (wheat bran, rice bran, wheat straw) which are dissolved or submerged in liquid media. Example of some liquid medium include domestic wastewater, Grape Pomace Hydrolysate, etc |
51,52,54 |
Metabolism of substrate |
Substrate is metabolized gradually |
Uniform metabolism of substrate |
64 |
Control of external factors |
Not feasible |
Feasible and in practice |
65 |
Suitable microbes |
Fungi (excluding yeasts) and other such low moisture requirement microbes. |
Bacteria, yeasts and other high moisture requirement microbes. |
53,66 |
Handling |
Cumbersome |
Simplified |
67,68 |
Challenges |
Effective control of environmental factors, Scale-up, heat dissipation, mass transfer. |
Viscosity of culture broth leading to foaming, Partial saccharification of lignocellulosic biomass |
69,70 |
Advantages |
Better yield, cost reduction, streamlined process. |
Ease of handling, Enhanced heat transfer. |
68 |
Solid state fermentation
Solid-state fermentation involves the utilization of solid substrates such as bagasse, bran, and paper pulp. A notable benefit of employing these substrates lies in the facile recycling of nutrient-rich waste materials. In this fermentation methodology, the substrates are metabolized gradually, enabling prolonged utilization of the same substrate throughout extended fermentation periods. Consequently, this technique facilitates the controlled release of specific nutrients. Solid-state fermentation is particularly well-suited for processes involving fungi and microorganisms that thrive under low moisture conditions. However, its applicability is constrained in fermentation processes that encompass a diverse range of organisms.56 Solid-state fermentation surpasses submerged fermentation in cellulase yield, and effective cost reduction is achievable through appropriate technology, enhanced bioreactor design, and a streamlined cellulase production process. The constituents of the medium play a pivotal role in influencing enzyme synthesis by various bacteria57 β–fructans and α-galactans, which are polymers of glucose, fructose and galactose, respectively. The synthesis of these compounds is catalyzed by glycosyltransferases (glycansucrases). The challenges associated with scaling up solid-state fermentation for industrial applications include difficulties in maintaining optimal temperature, pH, and moisture levels. Additional issues encompass the absence of homogeneous mass transfer, uneven fungal growth within the SSF media, and a lack of methodologies to accurately estimate fungal biomass concentrations.58 Once the production is complete, the recovery of the crude enzyme is done which are made to undergo certain downstream processes, viz., Solid-liquid extraction in which the enzyme is typically extracted by washing the solid substrate with a suitable buffer solution; Filtration or Pressing where the liquid (containing the enzyme) is filtered using filter presses, mesh filters or vacuum filters; Clarification is then done to remove fine particles and debris from the extracted liquid by processes such as centrifugation or by microfiltration; post this step, the enzyme is concentrated by ultrafiltration and precipitation with agents like ammonium sulfate to precipitate the enzyme. The ultimate step of obtaining the pure enzyme is purification in which the purity of cellulase is enhanced by removing other proteins and contaminants by means of processes like chromatography, dialysis, etc. The enzyme is then refined and stabilized using stabilizers such as glycerol for making it competent for storage.59 The overview of the process of SSF has been depicted in Figure 2.
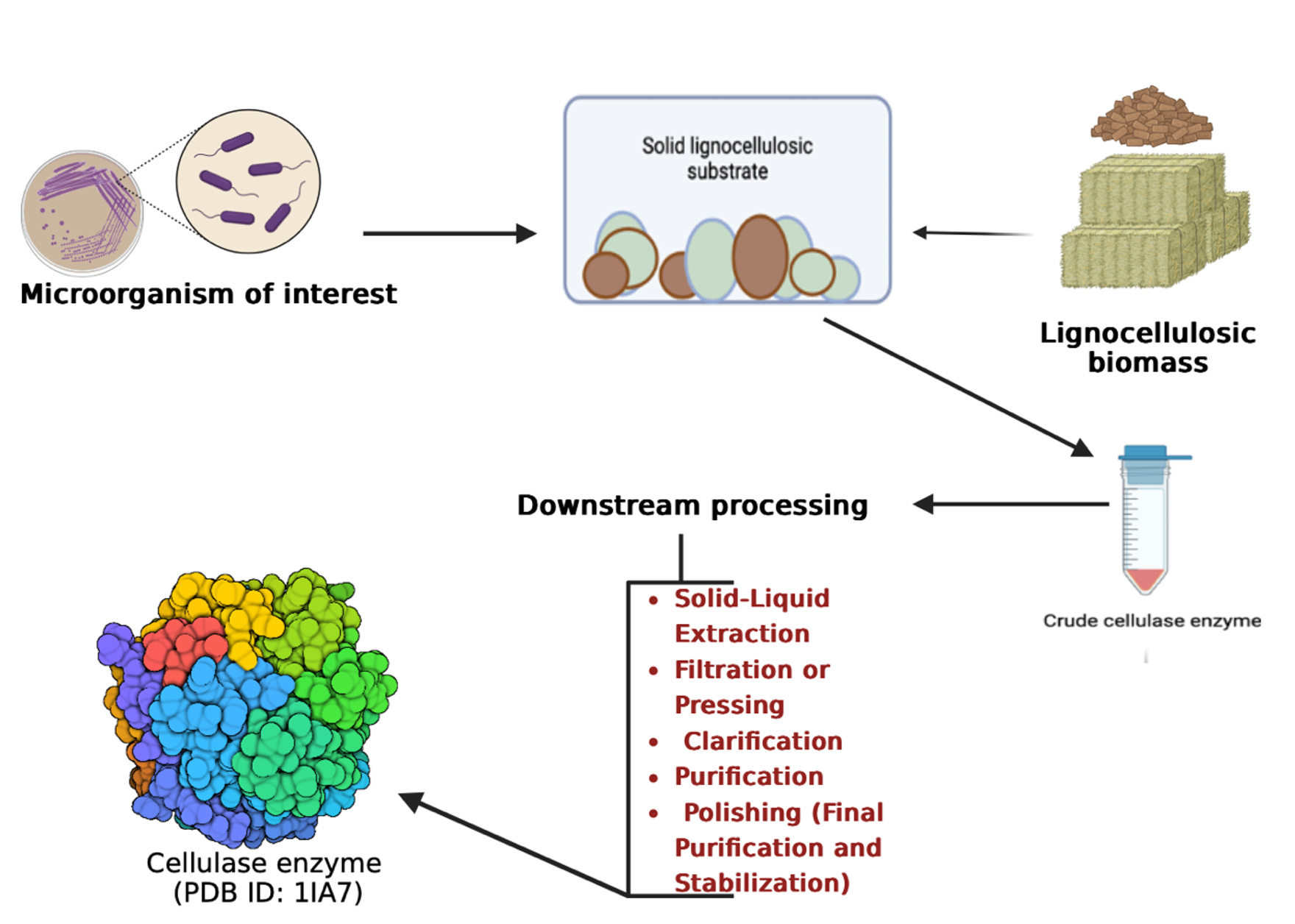
Figure 2. Solid state fermentation for cellulase production. Cellulase producing microorganism (s) employed to degrade lignocellulosic biomass substrate in solid state fermenter for the production of crude cellulase which is then purified by various downstream processes
Submerged fermentation
It is operationally characterized as the fermentation process conducted in an environment where water is present in excess up to 95%.60 The prevailing practice in numerous large-scale enzyme production facilities involves the utilization of SmF owing to its established technological efficacy, facilitating enhanced monitoring capabilities and simplified handling procedures.61 One of the most imperative reasons for the prevalence of submerged fermentation is due to the fact that the environmental factors such as pH and temperature can be controlled thereby reducing errors and cost.62 Enzymes for commercial applications are presently synthesized through submerged fermentation as a result of its cost-effectiveness and reduced enzyme consumption. Numerous potential applications of these enzymes have been successfully industrialized. Submerged fermentation exhibits advantages such as lower labor costs, reduced space requirements, enhanced control over environmental conditions, and less complexity in scaling up, when compared to solid-state fermentation.52
Within the context of submerged fermentation, extracellular endoglucanase activity was identified, and four strains exhibited similar patterns of enzyme secretion. The Klebsiella sp. (B2) strain displayed the lowest extracellular activity, although this disparity did not achieve statistical significance when contrasted with the activity of the other strains. The findings on petri dishes consistently mirrored CMC growth patterns and enzymatic hydrolysis profiles.58 It has been documented that during the implementation of submerged fermentation for cellulase production, several challenges are observed which generally reoccur.8 One of these challenges include an increase in the culture broth’s viscosity induced by cellulase production. Attempts to mitigate this viscosity increase through heightened agitation may result in unmanageable foaming. Additionally, the presence of secreted cellulase in the culture broth may lead to partial saccharification of lignocellulosic biomass and concentration of sugars (specifically glucose and xylose). Furthermore, a decline in dissolved oxygen percentage (DO%) in the broth is noted due to the heightened oxygen uptake by the proliferating fungus. Despite the multi-fold advantages presented by solid-state fermentation process-based bioreactors as compared to submerged fermentation bioreactors, the stirred-tank reactor with an automatic process control system prevails and emerges as the preferred choice for large-scale production of cellulase enzyme. This preference is attributed to its ease of handling, operation, and superior heat transfer characteristics.63 The overview of the process has been elucidated in Figure 3 which showcases the various steps of submerged fermentation.
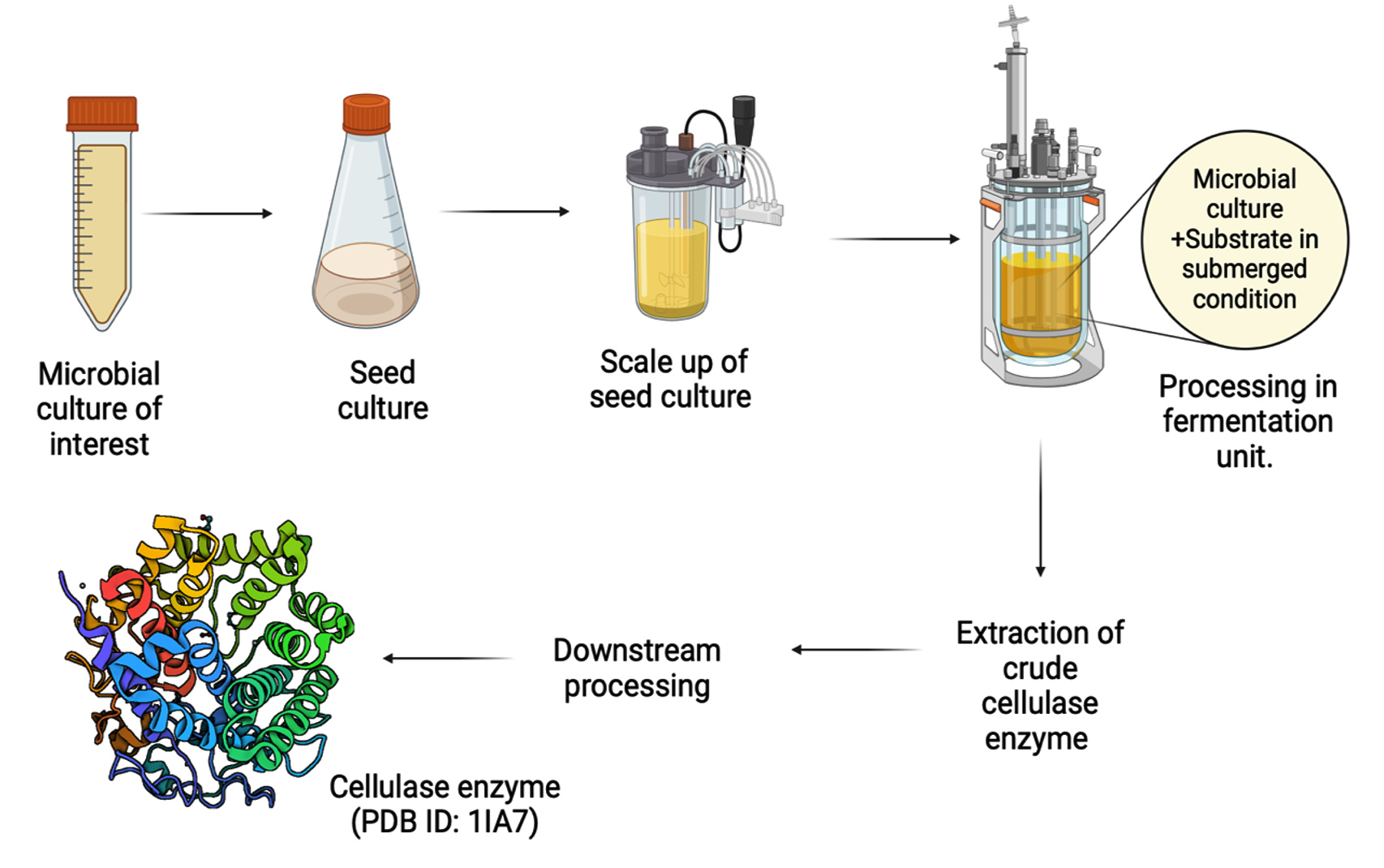
Figure 3. Submerged state fermentation for cellulase production. Novel microbial strains are used as seed culture which are then scaled up and processed in submerged state bioreactor for extraction of crude cellulase which is further subjected to downstream processes to obtain purified cellulase
Applications of cellulase enzyme
Cellulase exhibits significant potential across various industrial sectors. In the domain of industrial waste management, this enzyme is imperative for the degradation of cellulosic materials present in both domestic and industrial refuse, thereby enhancing the overall effectiveness of waste management protocols. Its contribution to bioethanol production is equally paramount, as it facilitates the conversion of cellulose into fermentable sugars, consequently improving the efficiency of biofuel production from cellulosic feedstocks. Moreover, in the paper and pulp industry, cellulase enzymes augment product quality by refining cellulose fibers and optimizing manufacturing processes.
Within the textile sector, cellulase enzymes are essential for environmentally sustainable processing by substituting conventional chemical techniques. They are extensively employed in denim washing to achieve a soft, aged appearance without compromising fabric integrity, as well as in fabric modification to enhance texture and quality. These enzymes also assume a critical function in dye removal and biofinishing applications, contributing to fabric softness, durability, and superior dye absorption. By supplanting traditional chemical methodologies in biodesizing and bioscouring, cellulase enzymes not only enhance operational efficiency but also diminish water usage and mitigate environmental repercussions, thereby endorsing sustainable practices.
In the agricultural sector, cellulase enzymes improve soil health and promote plant growth by facilitating the breakdown of crop residues and recycling vital nutrients back into the soil. They also assist in the decomposition of cellulosic substances within animal feed, enhancing nutrient assimilation and livestock productivity. In composting processes, these enzymes expedite the decomposition of organic matter, yielding nutrient-rich compost for soil enrichment. The applications extend further into bioremediation, where cellulase enzymes facilitate the degradation of cellulose-based pollutants, thereby aiding environmental remediation efforts. Bioethanol represents a sustainable energy source. Notably, second-generation bioethanol production is gaining traction due to the availability of economical raw materials. Lignocellulosic biomass, the primary feedstock, encompasses agricultural residues, herbaceous and woody crops, forestry waste, and various municipal and industrial by-products. Lignocellulose constitutes the most prevalent renewable biomass. Global lignocellulose yield is estimated at around 200 billion metric tons annually. Utilizing these feedstocks for bioethanol production offers numerous benefits. It effectively facilitates the disposal of non-edible plant materials. Its lower pollutant emissions render it environmentally advantageous. Crucially, this process does not compromise food security as it avoids the use of food crops pre-harvest. Moreover, it provides a year-round supply of raw materials.
Conversely, a significant limitation of this production method lies in the structural composition of lignocellulosic biomass. This biomass is predominantly comprised of lignin, cellulose, and hemicellulose, forming a highly stable matrix. To liberate fermentable sugars from this substrate, a pretreatment is necessary to disrupt the stable structure, wherein cellulase enzymes play a crucial role.71
Additionally, they find extensive application in the food and beverage industry for juice extraction, flavor enhancement in wine production, and product clarification, while also playing an indispensable role in biofuel generation and pharmaceutical applications.6
Strain improvement
The enhancement of cellulase production can be achieved through strain improvement techniques such as involving the genetic modification of microbial strains, employing advanced methodologies, that can be used to attain significant results. Genetic modifications and random mutagenesis of parental strains play a pivotal role in amplifying the commercial production of cellulases. Additionally, the molecular cloning of cellulase genes and the augmentation of gene copy numbers exert a substantial influence on the degradation of cellulosic materials and the production of high-quality value-added products.
Strain improvement emerges as a promising strategy for elevating cellulase production Wild-type or normal strains may possess distinct characteristics suitable for industrial applications. To fully exploit the potential of wild-type microorganisms, strain improvement is imperative, allowing for the modification or elimination of specific traits. Recombinant DNA technology, site-directed mutagenesis, recombination protoplast fusion, and random mutagenesis, contribute significantly to enhancing commercial production. Immobilizing cellulase on a solid support represents another method to improve the catalytic efficiency of enzymes for industrial applications.72 Introducing a novel strategy for enhancing catalytic activity involves the application of substrate-induced gene-expression screening (SIGEX) coupled with fluorescence-activated cell sorting (FACS). Additionally, bacterial mutagenesis has been employed to elevate cellulase activity in thermophilic cellulase-producing organisms. For an extended duration, the limited efficacy in gene targeting has posed a significant challenge in achieving a substantial yield of transformants through homologous integration or expression cassette deletion. These obstacles stem from the difficulties in executing such processes. Strains capable of thriving on more cost-effective substrates have emerged as promising reservoirs for metabolites of industrial interest. Extensive research endeavours and practical applications over the years have contributed to a comprehensive understanding of fungal genetics.73 Using fluorescence-activated cell sorting (FACS) in conjunction with substrate-induced gene-expression screening (SIGEX) is one way to improve catalytic activity. In thermophiles that produce cellulases, there was an increase in cellulase activity by bacterial mutagenesis. For instance, changed clones were obtained when NTG (N-methyl-N′-nitro- N-nitrosoguanidine) was applied as a mutagen to Bacillus sp. strain C1.74 Analysis of the properties of bacterial cellulose generated by Komagataeibacter sp. nov. CGMCC 17276 under static and agitated conditions revealed the presence of strongly cross-linked cellulose nanomaterial.69,75 Studies76 based on phylogenetic position and phenotypic characteristics, the high-yield bacteria cellulase-producing strain Komagataeibacter sp. nov. CGMCC 17,276 was assigned as a novel species in the Komagataeibacter genus with good properties of rapid cell growth and high bacterial cellulase production. Sequential mutagenesis employing three agents-ultraviolet irradiation (UV), N-methyl-N′-nitro-N-nitrosoguanidine (NTG), and ethyl methanesulfonate-resulted in enhanced activity of Streptomyces durhamensis EMS.77-79 Following mutagenesis, the cellulase activity of mutant Streptomyces durhamensis GC23 exhibited a 1.86-fold improvement compared to the wild-type strain (vs15). Subsequent optimization of the cellulase activity in mutant Streptomyces durhamensis GC23 led to a twofold increase relative to the wild type, as documented by Ega et al.78
The various strategies to enhance the production of cellulases include molecular docking, by employing computational methodologies and strategies to forecast enzyme-substrate interactions, facilitating enhanced enzyme engineering. Molecular docking insights on the cellulase production of bacterium Bacillus amyloliquefaciens indicate that the cellulase enzyme derived from the Bacillus amyloliquefaciens strain elh1 exhibits a robust and specific affinity A critical factor for its cellulolytic activity.80 These findings facilitate subsequent optimization and augmentation of cellulase performance; Inclusion of multifunctional cellulase genes along with the combination of various cellulase genes can help provide a means to lower enzyme costs by reducing the complexity of enzyme cocktails while aiding to increase the efficiency of enzyme hydrolysis. Use of effective promoters can upregulate the transcription of the enzyme leading to heightened enzyme production. This was elucidated in a study,81 where overexpression of the transcription factors Xyr1 and Ace3 of Trichoderma orientalis EU7-22 was done which led to the increased production of cellulase and hemicellulase in the recombinant strain dxyA-8 illustrating the efficacy of the chosen promoters in attaining elevated enzyme activity. The overview of the strategies is depicted in Figure 4.
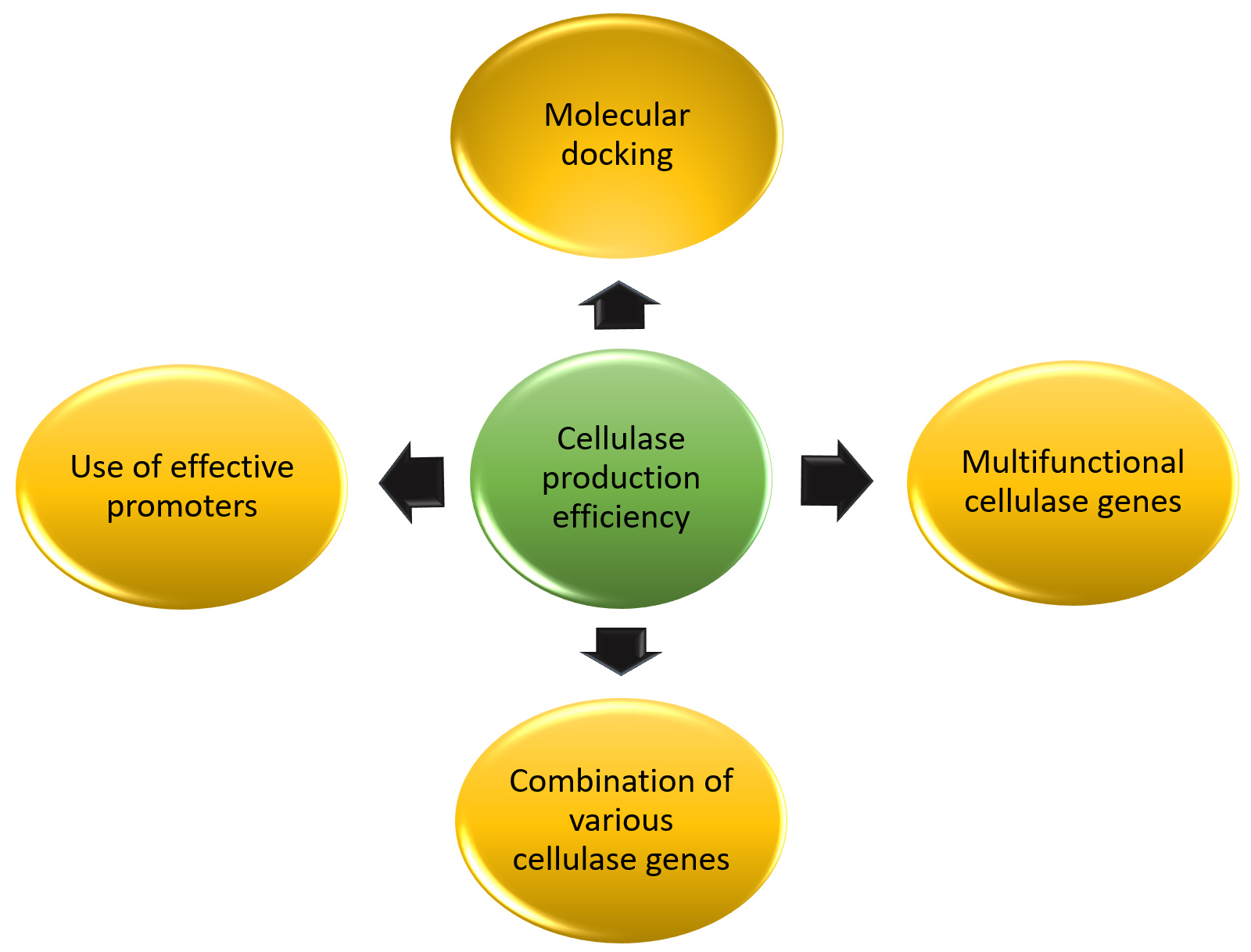
Figure 4. Strategies to enhance cellulase production. Techniques such as molecular docking, identification of multifunctional cellulase producing genes, combination of various cellulase genes, use of effective promoters can be considered as effective strategies for the same
Challenges in cellulase production
Lignocellulosic biomass represents a promising reservoir for the generation of biofuels, biofertilizers, animal feed, and chemicals, in addition to serving as a primary constituent for the paper industry. The harnessing of this renewable resource necessitates either chemical or biological treatment methods, with cellulases emerging as prominently favoured agents in the latter approach over the preceding decades. Investigative endeavours have elucidated the intricacies underlying microbial cellulase production mechanisms, leading to the evolution of technologies dedicated to the production and application of cellulose-degrading enzymes; namely cellulase. Despite these advancements, a challenge persists in the absence of a singular, cost-effective, and efficient process for converting natural lignocellulosic materials into valuable metabolites or biofuel. The utilization of existing commercial cellulase preparations for the bioconversion of lignocellulosic waste is presently deemed economically impractical as there is still a debate as to which method is deemed suitable for the production of cellulase.74 Some practical drawbacks include enzyme synergy, improperly treated lignocellulosic biomass, excess solid concentrations, sub-optimal ratio of critical factors, etc.82
The following points delve into the challenges faced in cellulase production
- Fermentation Techniques: Various fermentation methodologies, including submerged and solid-state fermentation, present distinct challenges in the optimization of cellulase yield. Each methodology possesses its own array of operational difficulties that can significantly influence the efficiency of enzyme production.83
- Scale-Up Issues: The transition from laboratory-scale to pilot-scale production frequently encounters bottlenecks, complicating the maintenance of efficiency and cost-effectiveness.74
Cost and Economic Feasibility
- Production Costs: The substantial expenses associated with cellulase production continue to pose a significant barrier, prompting the necessity for the investigation of more economical substrates and enhanced microbial strains.84
- Monitoring and Control: The effective monitoring of the fermentation process is paramount for cost reduction; however, the intricate nature of the bioprocess presents challenges in the formulation of precise models.85
Optimization of Microbial Strains
- Genetic Modification: Although the genetic modification of microbial strains has the potential to augment cellulase production, the initial yield of isolated strains is frequently suboptimal, thereby necessitating further refinement.84
Challenges faced during isolation, screening and growth on various substrates
- Cultivation Recalcitrance: Numerous cellulolytic microbes exhibit recalcitrance to laboratory cultivation, restricting access to a plethora of potential cellulase-producing organisms found in natural ecosystems.
- Diversity of Microbial Communities: The extensive diversity within microbial communities that synthesize cellulases complicates the isolation of specific strains, thereby hindering the identification and characterization of the most effective cellulolytic microbes.
- Variability in Fermentation Conditions: Variations in fermentation parameters, including media composition and raw materials, influence cellulase production across different microbial strains, thereby complicating comparative analyses of cellulase yields and activities.
- Assay Method Discrepancies: The variability in methodologies employed to assay cellulase activity across studies creates challenges in result comparison and in determining the most effective cellulolytic strains.
- Technological Limitations: Conventional methods for isolating cellulolytic microbes may fail to encompass the complete spectrum of potential cellulase producers, particularly those that are unculturable, necessitating advanced methodologies such as metagenomics to access these elusive microbial resources.
- Environmental Factors: The extreme conditions prevalent in the natural habitats of cellulolytic microbes (e.g., high temperature, salinity) complicate isolation efforts, making it imperative to comprehend their specific environmental requirements for successful cultivation.86
In spite of these obstacles, ongoing research endeavours strive to enhance cellulase production through innovative biotechnological strategies and a more profound comprehension of enzymatic mechanisms, which could ultimately facilitate the development of more sustainable biofuel production methodologies.
Future prospects in cellulase production
Cellulases are imperative enzymes as they are employed in degradation of lignocellulosic biomass for their subsequent conversion to biofuels. Use of micro-organisms not only makes the process eco-friendly and sustainable but also adds to the cost-effectiveness of the process.
The following points elucidate the future prospects that can aid in the effective and enhanced production of cellulase enzyme. The methods by which effective cellulase production can be achieved by,
- Bacteria mediated transformation; chemical and/or natural.75
- Genetic engineering for multifunctional cellulase genes: Creating genetic modification to cellulase genes to enhanced production capabilities and efficiency,
- Combining different cellulase genes: Strategically combining genes from various sources,
- Identification of genes from cellulose-degrading insects: Studying insects like termites to discover novel and potentially powerful cellulase genes,
- Use of effective promoters: Use of genetic promoters to improve the expression and production of cellulase enzymes,
- Molecular docking: Utilizing computational tools and techniques to predict enzyme-substrate interactions, enabling improved enzyme design.
- Consolidated Bioprocessing (CBP): Creating one-pot procedures that integrate the conversion of biomass and the synthesis of cellulase into a single stage. This simplifies the process and lowers manufacturing costs by eliminating the requirement for distinct enzyme purification processes.
Cellulases, a class of hydrolytic enzymes, occupy a prominent position within the enzyme market. Their significance extends beyond commercial value, which plays a crucial role in the pretreatment of lignocellulosic biomass. This pretreatment step is essential for the production of biofuels from renewable sources. Furthermore, cellulases find diverse applications in various industries, including textiles and pulp and paper production. Microbial cellulases have garnered significant research interest due to their economic viability compared to alternative methods. The alternative methodologies—chemical hydrolysis, physical treatments, thermochemical processes, and solvent-based strategies—frequently incur higher expenses attributable to factors such as substantial energy requirements, costly reagents, intricate apparatus, and ecological implications. In contrast, microbial cellulases are synthesized biologically under benign conditions utilizing cost-effective substrates. This results in reduced production expenses, diminished environmental concerns, and enhanced scalability, rendering them a more economically feasible option for cellulose degradation and conversion within industrial contexts. Microbial cellulases have attracted considerable scholarly attention owing to their economic advantages relative to alternative methodologies. Researchers are actively exploring both traditional and novel technologies for cellulase production. These advancements hold immense promise for the efficient and cost-effective production of this critical enzyme.
ACKNOWLEDGMENTS
The authors are thankful to Tamil Nadu Agricultural University, Coimbatore, India, for their support.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Robinson PK. Enzymes: principles and biotechnological applications. Essays Biochem. 2015;59:1-41.
Crossref - Fu ZH, Liu J, Zhong LB, et al. Screening of cellulose-degrading yeast and evaluation of its potential for degradation of coconut oil cake. Front Microbiol. 2022;13:996930.
Crossref - Islam F, Roy N. Screening, purification and characterization of cellulase from cellulase producing bacteria in molasses. BMC Res Notes. 2018;11(1):445.
Crossref - Saldana-Mendoza SA, Ascacio-Valdes JA, Palacios-Ponce AS, et al. Use of wastes from the tea and coffee industries for the production of cellulases using fungi isolated from the Western Ghats of India. Syst Microbiol Biomanuf. 2021;1(1):33-41.
Crossref - Marin M, Sanchez A, Artola A. Production and recovery of cellulases through solid-state fermentation of selected lignocellulosic wastes. J Clean Prod. 2019;209:937-946.
Crossref - de Almeida MN, Falkoski DL, Guimaraes VM, de Rezende ST. Study of gamba grass as carbon source for cellulase production by Fusarium verticillioides and its application on sugarcane bagasse saccharification. Ind Crops Prod. 2019;133:33-43.
Crossref - Crognale S, Liuzzi F, D’Annibale A, de Bari I, Petruccioli M. Cynara cardunculus a novel substrate for solid-state production of Aspergillus tubingensis cellulases and sugar hydrolysates. Biomass Bioenergy. 2019;127:105276.
Crossref - Mattam AJ, Chaudhari YB, Velankar HR. Factors regulating cellulolytic gene expression in filamentous fungi: an overview. Microb Cell Fact. 2022;21(1):44.
Crossref - Christopher M, Sreeja-Raju A, Kooloth-Valappil P, Gokhale DV, Sukumaran RK. Cellulase Hyper-Producing Fungus Penicillium janthinellum NCIM 1366 Elaborates a Wider Array of Proteins Involved in Transport and Secretion, Potentially Enabling a Diverse Substrate Range. Bioenerg Res. 2023;16(1):61-73.
Crossref - Isola D, Bartoli F, Meloni P, Caneva G, Zucconi L. Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides. Appl Sci. 2022;12(4):2038.
Crossref - Goyari S, Devi SS, Kalita MC, Talukdar NC. Population, diversity and characteristics of cellulolytic microorganisms from the Indo-Burma Biodiversity hotspot. SpringerPlus. 2014;3(1):700.
Crossref - Mohammadipour Z, Enayatizamir N, Ghezelbash G, Moezzi A. Bacterial Diversity and Chemical Properties of Wheat Straw-Based Compost Leachate and Screening of Cellulase Producing Bacteria. Waste Biomass Valor. 2021;12(3):1293-1302.
Crossref - Balla A, Silini A, Cherif-Silini H, et al. Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions. Catalysts. 2022;12(7):769.
Crossref - Bhagat SA, Kokitkar SS. Isolation and identification of bacteria with cellulose-degrading potential from soil and optimization of cellulase production. J App Biol Biotech. 2021;9(6).
Crossref - Larson JA, Bagley DM. Sessile and Planktonic Microbial Taxonomy of a Methanogenic Cellulolytic Enrichment Reactor Sourced from the Organic Fraction of Municipal Solid Waste. J Environ Eng. 2022;148(4):04022004.
Crossref - Dennis C. Breakdown of Cellulose by Yeast Species. J Gen Microbiol. 1972;71(2):409-411.
Crossref - Sohail M, Barzkar N, Michaud P, et al. Cellulolytic and Xylanolytic Enzymes from Yeasts: Properties and Industrial Applications. Molecules. 2022;27(12):3783.
Crossref - Jimenez M, Gonzalez AE, Martinez MJ, Martinez AT, Dale BE. Screening of yeasts isolated from decayed wood for lignocellulose-degrading enzyme activities. Mycological Research. 1991;95(11):1299-1302.
Crossref - Saini A, Aggarwal NK, Sharma A, Yadav A. Actinomycetes: A Source of Lignocellulolytic Enzymes. Enzyme Res. 2015;2015:1-15.
Crossref - Swarna D, Gnanadoss JJ. Screening and Molecular Characterization of Actinomycetes from Mangrove Soil Producing Industrially Important Enzymes. J Sci Res. 2020;64(02):87-95.
Crossref - Baraldo Junior A, Borges DG, Tardioli PW, Farinas CS. Characterization of β-Glucosidase Produced by Aspergillus niger under Solid-State Fermentation and Partially Purified Using MANAE-Agarose. Biotechnol Res Int. 2014;2014:1-8.
Crossref - Santos GB, de Sousa FFA, da Silva RJR, Rodrigues deSR. Cellulase production by Aspergillus niger using urban lignocellulosic waste as substrate: Evaluation of different cultivation strategies. J Environ Manag. 2022;305:114431.
Crossref - Kumar A. Aspergillus nidulans: A Potential Resource of the Production of the Native and Heterologous Enzymes for Industrial Applications. Int J Microbiol. 2020;2020:1-11.
Crossref - Dutta SD, Tarafder M, Islam R, Datta B. Characterization of cellulolytic enzymes of Fusarium soil Isolates. Biocatal Agric Biotechnol. 2018;14:279-285.
Crossref - Riaz H, Abdul J, Hamid RM, Riaz S, Latif F. Endoglucanase Production by Humicola insolens: Effect of Physiochemical Factors on Growth Kinetics and Thermodynamics. Int J Agric Biol. 2014;16(6):1141-1146.
- Santa-Rosa PS, Souza AL, Roque RA, et al. Production of thermostable β-glucosidase and CMCase by Penicillium sp. LMI01 isolated from the Amazon region. Electron J Biotechnol. 2018;31:84-92.
Crossref - Tiwari P, Misra BN, Sangwan NS. β-Glucosidases from the Fungus Trichoderma: An Efficient Cellulase Machinery in Biotechnological Applications. BioMed Res Int. 2013;2013:1-10.
Crossref - Zhao CH, Liu X, Zhan T, He J. Production of cellulase by Trichoderma reesei from pretreated straw and furfural residues. RSC Adv. 2018;8(63):36233-36238.
Crossref - Macris BJ, Ketos D, Evangelidou X, Galiotou-Panayotou M, Sodis P. Solid state fermentation of straw with Neurospora crassa for CMCase and β-glucosidase production. Biotechnol Lett. 1987:661-664.
Crossref - Lee CK, Darah I, Ibrahim CO. Production and Optimization of Cellulase Enzyme Using Aspergillus niger USM AI 1 and Comparison with Trichoderma reesei via Solid State Fermentation System. Biotechnol Res Int. 2011;2011:1-6.
Crossref - Liu J, Wang X, Xu D. QM/MM Study on the Catalytic Mechanism of Cellulose Hydrolysis Catalyzed by Cellulase Cel5A from Acidothermus cellulolyticus. J Phys Chem B. 2010;114(3):1462-1470.
Crossref - Dong W, Xue M, Zhang Y, et al. Characterization of a β-glucosidase from Paenibacillus species and its application for succinic acid production from sugarcane bagasse hydrolysate. Bioresour Technol. 2017;241:309-316.
Crossref - Liang YL, Zhang Z, Wu M, Wu Y, Feng JX. Isolation, Screening, and Identification of Cellulolytic Bacteria from Natural Reserves in the Subtropical Region of China and Optimization of Cellulase Production by Paenibacillus terrae ME27-1. BioMed Res Int. 2014;2014(5):512497.
Crossref - Deka D, Das SP, Sahoo N, et al. Enhanced Cellulase Production from Bacillus subtilis by Optimizing Physical Parameters for Bioethanol Production. ISRN Biotechnol. 2013;2013:965310.
Crossref - Raut MP, Couto N, Karunakaran E, Biggs CA, Wright PC. Deciphering the unique cellulose degradation mechanism of the ruminal bacterium Fibrobacter succinogenes S85. Sci Rep. 2019;9(1):16542.
Crossref - Maki ML, Broere M, Leung KT, Qin W. Characterization of some efficient cellulase producing bacteria isolated from paper mill sludges and organic fertilizers. Int J Biochem Mol Biol. 2011;2(2):146-154.
- Crawford DL, McCoy E. Cellulases of Thermomonospora fusca and Streptomyces thermodiastaticus. Appl Microbiol. 1972;24(1):150-152.
Crossref - El-Sersy NA, Abd-Elnaby H, Abou-Elela GM, Ibrahim HAH, El-Toukhy NMK. Optimization, economization and characterization of cellulase produced by marine Streptomyces ruber. Afr J Biotechnol. 2010;9(38):6355-6364.
- Langsford ML, Gilkes NR, Wakarchuk WW, Jr. Miller RC, R.A.J Warren. The cellulase system of Cellulomonas fimi. J Gen Microbiol. 1984;130(6):1367-1376.
Crossref - Yun YH, Suh DY, Yoo HD, Oh MH, Kim SH. Yeast Associated with the Ambrosia Beetle, Platypus koryoensis, the Pest of Oak Trees in Korea. Mycobiology. 2015;43(4):458-466.
Crossref - Korhola M, Hakonen R, Juuti K, et al. Production of folate in oat bran fermentation by yeasts isolated from barley and diverse foods. J Appl Microbiol. 2014;117(3):679-689.
Crossref - Morais CG, Sena LMF, Lopes MR, et al. Production of ethanol and xylanolytic enzymes by yeasts inhabiting rotting wood isolated in sugarcane bagasse hydrolysate. Fungal Biol. 2020;124(7):639-647.
Crossref - McMahon H, Zoecklein BW, Fugelsang K, Jasinski Y. Quantification of glycosidase activities in selected yeasts and lactic acid bacteria. J Ind Microbiol Biotechnol. 1999;23(3):198-203.
Crossref - Ali SS, Wu J, Xie R, Zhou F, Sun J, Huang M. Screening and characterizing of xylanolytic and xylose-fermenting yeasts isolated from the wood-feeding termite, Reticulitermes chinensis. Yang S, ed. PLoS ONE. 2017;12(7):e0181141.
Crossref - Giese EC, Dussan KJ, Pierozzi M, Chandel AK, Pagnocca FC, Da Silva SS. Cellulase Production by Trichosporon laibachii. Orbital: Electron J Chem. 2017;9(4):271-278.
Crossref - Belda I, Ruiz J, Alastruey-Izquierdo A, Navascues E, Marquina D, Santos A. Unraveling the Enzymatic Basis of Wine “Flavorome”: A Phylo-Functional Study of Wine Related Yeast Species. Front Microbiol. 2016;7:12.
Crossref - Elhalis H, Cox J, Frank D, Zhao J. Microbiological and biochemical performances of six yeast species as potential starter cultures for wet fermentation of coffee beans. LWT. 2021;137:110430.
Crossref - Escribano-Viana R, Gonzalez-Arenzana L, Garijo P, Lopez R, Santamaria P, Gutierrez AR. Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja. Fermentation. 2021;7(3):148.
Crossref - Gomes J, Gomes I, Steiner W. Thermolabile xylanase of the Antarctic yeast Cryptococcus adeliae: production and properties. Extremophiles. 2000;4(4):227-235.
Crossref - Lara CA, Santos RO, Cadete RM, et al. Identification and characterisation of xylanolytic yeasts isolated from decaying wood and sugarcane bagasse in Brazil. Antonie van Leeuwenhoek. 2014;105(6):1107-1119.
Crossref - Singh A, Bajar S, Devi A, Bishnoi NR. Evaluation of cellulase production from Aspergillus niger and Aspergillus heteromorphus under submerged and solid-state fermentation. Environ Sustain. 2021;4(2):437-442.
Crossref - Libardi N, Soccol CR, de Carvalho JC, de Souza VLP. Simultaneous cellulase production using domestic wastewater and bioprocess effluent treatment – A biorefinery approach. Bioresour Technol. 2019;276:42-50.
Crossref - Behera SS, Ray RC. Chapter 12 – Forest Bioresources for Bioethanol and Biodiesel Production With Emphasis on Mohua (Madhuca latifolia L.) Flowers and Seeds. Bioethanol Prod Food Crops. 2019:233-247.
Crossref - Jin Q, O’Hair J, Stewart AC, et al. Compositional Characterization of Different Industrial White and Red Grape Pomaces in Virginia and the Potential Valorization of the Major Components. Foods. 2019;8(12):667.
Crossref - Ilic N, Milic M, Beluhan S, Dimitrijevic-Brankovic S. Cellulases: From Lignocellulosic Biomass to Improved Production. Energies. 2023;16(8):3598.
Crossref - Dasari PR, Ramteke PW, Kesri S, Kongala PR. Comparative Study of Cellulase Production Using Submerged and Solid-State Fermentation. Fungal Biol. 2019:37-52.
Crossref - Nabot M, Guerin M, Sivakumar D, Remize F, Garcia C. Variability of Bacterial Homopolysaccharide Production and Properties during Food Processing. Biology. 2022;11(2):171.
Crossref - Kumar Ramamoorthy N, Sambavi TR, Renganathan S. A study on cellulase production from a mixture of lignocellulosic wastes. Process Biochem. 2019;83:148-158.
Crossref - Adhyaru DN, Bhatt NS, Modi HA. Optimization of upstream and downstream process parameters for cellulase-poor-thermo-solvent-stable xylanase production and extraction by Aspergillus tubingensis FDHN1. Bioresour Bioprocess. 2015;2(1):3.
Crossref - Martinez-Medina GA, Barragan AP, Ruiz HA, et al. Fungal Proteases and Production of Bioactive Peptides for the Food Industry. Enzymes Food Biotechnol. 2019:221-246.
Crossref - Patel AK, Pandey A, Singhania RR. Production of Celluloytic Enzymes for Lignocellulosic Biomass Hydrolysis. Biofuels: Alternative Feedstocks and Conversion Processes for the Production of Liquid and Gaseous Biofuels. 2019:401-426.
Crossref - Oh EJ, Jin YS. Engineering of Saccharomyces cerevisiae for efficient fermentation of cellulose. FEMS Yeast Research. 2020;20(1):foz089.
Crossref - Ramesh D, Muniraj IK, Thangavelu K, Karthikeyan S. Knowledge Update on Bioreactor Technology for cellulase Production. In: Elsevier eBooks.; 2019:181-193.
Crossref - Sharif M, Zafar MH, Aqib AI, Saeed M, Farag MR, Alagawany M. Single cell protein: Sources, mechanism of production, nutritional value and its uses in aquaculture nutrition. Aquaculture. 2021;531:735885.
Crossref - Sentis-More P, Romero-Fabregat MP, Rodriguez-Marca C, Guerra-Sanchez AJ, Ortega-Olive N. Design Optimization of a Tray Bioreactor for Solid-State Fermentation: Study of Process Parameters through Protein Modification of By-Products. Fermentation. 2023;9(10):921.
Crossref - Basak A, Murmu N, Gavande PV, Ghosh S. Valorization of Rice Straw for Optimization of Production of Thermotolerant Cellulase by Parageobacillus Thermoglucosidasius NBCB1 Under Submerged. Fermentation. 2022.
Crossref - Arora S, Rani R, Ghosh S. Bioreactors in solid state fermentation technology: Design, applications and engineering aspects. J Biotechnol. 2018;269:16-34.
Crossref - Behl M, Thakar S, Ghai H, Sakhuja D, Bhatt AK. Fundamentals of fermentation technology. Basic Biotechniques for Bioprocess and Bioentrepreneurship. 2023:313-328.
Crossref - Kumar V, Ahluwalia V, Saran S, Kumar J, Patel AK, Singhania RR. Recent developments on solid-state fermentation for production of microbial secondary metabolites: Challenges and solutions. Bioresour Technol. 2021;323:124566.
Crossref - Liu X, Kokare C. Microbial Enzymes of Use in Industry. Biotechnol Microb Enzymes. 2017:267-298.
Crossref - AA Abdulsalam, S Aliyu, B.L Bashar, et al. Bioethanol: A Sustainable Liquid Fuel as Substitute to Gasoline. IJRASET. 2024;12(7):360-371.
Crossref - H Sher, N Zeb, S Zeb, et al. Microbial Cellulases: A Review on Strain Development, Purification, Characterization and their Industrial Applications. Journal of Bacteriology and Mycology. 2021;8(5).
Crossref - U-thai P, Poonsrisawat A, Arnthong J, et al. Enhanced viscosity reduction efficacy of cassava root mash by Aspergillus aculeatinus mutant enzyme cocktail. Biomass Conv Bioref. 2023;13(13):11803-11812.
Crossref - Singhania RR, Ruiz HA, Awasthi MK, Dong CD, Chen CW, Patel AK. Challenges in cellulase bioprocess for biofuel applications. Renew Sustain Energy Rev. 2021;151:111622.
Crossref - Betlej I, Zakaria S, Krajewski KJ, Boruszewski P. Bacterial Cellulose – Properties and Its Potential Application. Sains Malaysiana. 2021;50(2):493-505.
Crossref - Lu T, Gao H, Liao B, et al. Characterization and optimization of production of bacterial cellulose from strain CGMCC 17276 based on whole-genome analysis. Carbohydr Polym. 2020;232:115788.
Crossref - Azouz R. Bottom-Up Approach Through Microbial Green Biosynthesis of Nanoparticles from Waste. Waste Recycling Technologies for Nanomaterials Manufacturing. Topics in Mining, Metallurgy and Materials Engineering. 2021:671-697.
Crossref - Ega SL, Rao MRN, Sudhamani M. Response surface methodology-artificial neural network based optimization and strain improvement of cellulase production by Streptomyces sp. Biosci J. 2020;36(4).
Crossref - Golinska P, Montero-Calasanz MDC, Swiecimska M, et al. Modestobacter excelsi sp. nov., a novel actinobacterium isolated from a high altitude Atacama Desert soil. Syst Appl Microbiol. 2020;43(1):126051.
Crossref - El-Khamisi EF, Soliman EAM, El-Sayed GM, Nour SA, Abdel-Monem MO, Hassan MG. Optimization, gene cloning, expression, and molecular docking insights for enhanced cellulase enzyme production by Bacillus amyloliquefaciens strain elh1. Microb Cell Fact. 2024;23(1):191.
Crossref - Xue Y, Han J, Li Y, Liu J, Gan L, Long M. Promoting cellulase and hemicellulase production from Trichoderma orientalis EU7-22 by overexpression of transcription factors Xyr1 and Ace3. Bioresour Technol. 2020;296:122355.
Crossref - Bhardwaj N, Kumar B, Agrawal K, Verma P. Current perspective on production and applications of microbial cellulases: a review. Bioresour Bioprocess. 2021;8(1):95.
Crossref - Bajpai P. Current production status of cellulases and challenges. Cellulases in the Biofuel Industry. 2023:91-108.
Crossref - Rabby MRI, Ahmed ZB, Paul GK, et al. A Combined Study on Optimization, In Silico Modeling, and Genetic Modification of Large Scale Microbial Cellulase Production. Mohamed SA, ed. Biochem Res Int. 2022;2022:1-14.
Crossref - Murugan C. Soft Sensors for Biomass Monitoring during Low Cost Cellulase Production. Biotechnological Applications of Biomass. IntechOpen. 2021.
Crossref - Kuhad RC, Deswal D, Sharma S, et al. Revisiting cellulase production and redefining current strategies based on major challenges. Renew Sustain Energy Rev. 2016;55:249-272.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.