


ISSN: 0973-7510
E-ISSN: 2581-690X
Selenium (Se) is a necessary essential component that has antioxidant, immune-modulating, and anticancer activities. Emerging research highlights the role of gut microbiota in colorectal cancer, with probiotics like Lactobacillus and Bifidobacterium potentially enhancing treatment efficacy, reducing inflammation, and supporting gut homeostasis. Probiotics, particularly Bacillus subtilis subsp. stercoris, play a vital role in gut microbiota regulation, improving gastrointestinal health and nutrient bioavailability. In this study, Bacillus subtilis subsp. stercoris was isolated from fermented sorghum (Koozh), a traditional Tamilian fermented food, and enriched with sodium selenite to enhance its functional properties. The selenium enrichment process resulted in a distinct orange pigmentation, indicating the biotransformation of inorganic selenium into bioavailable organic selenium compounds. The enriched strain was characterized for probiotic potential, including gastric fluid and bile salt tolerance, carbohydrate fermentation, and hydrophobicity. To evaluate its anticancer effects, an MTT assay was performed on Caco-2 colorectal cancer cells, revealing dose-dependent cytotoxicity with an IC50 of 81.517 µg/mL. Further AO/ETBR staining confirmed apoptosis induction in treated cancer cells, while maintaining good viability in non-cancerous cells. Selenium-enriched probiotics, particularly Bacillus subtilis subsp. stercoris, show promise as functional food supplements with potential anticancer and gut health benefits, highlighting their therapeutic potential as nutraceuticals for probiotic formulations, functional foods, and colorectal cancer prevention.
Selenium-enriched Probiotics, Bacillus subtilis, Functional Foods, Koozh, Anticancer Activity, Gut Microbiota, Colorectal Cancer Therapy
Colorectal cancer (CRC) is one of the most prevalent cancers globally, impacted by hereditary, lifestyle, and environmental factors. Emerging research underlines the critical involvement of gut microbiota in CRC formation and progression, with an imbalance in microbial composition linked to inflammation, immunological dysregulation, and carcinogenesis.1 Probiotic strains like Lactobacillus and Bifidobacterium help to maintain gut homeostasis by suppressing pathogenic bacteria, generating beneficial metabolites, and lowering oxidative stress and irritation.2,3 Early clinical findings indicate that probiotics may improve chemotherapy tolerance, increase treatment efficacy, and reduce the risk of CRC by affecting cell proliferation, apoptosis, and immunological modulation.4
Selenium (Se) is a necessary component mineral that plays a crucial role in various biological functions, antioxidant defense, immune system modulation, and cancer prevention.5 Selenoproteins, including GPx and TrxR, play a crucial role in protecting cells from oxidative stress and lowering the risk of chronic illness.6 Selenium deficiency is associated with weakened immunity, neurodegenerative disorders, cardiovascular diseases, and increased susceptibility to cancer.7 The human body cannot synthesize selenium, so it must be obtained from dietary sources such as selenium-rich foods, supplements, or biofortified functional foods.8
Although selenium is essential for human health, its bioavailability and absorption depend on its chemical form.9 Most dietary sources contain inorganic selenium, which has limited absorption efficiency and potential toxicity.10 However, probiotic bacteria like Bacillus subtilis can naturally transform inorganic selenium into organic forms, such as selenocysteine, selenomethionine, and selenium nanoparticles, through enzymatic biotransformation.11 Research has highlighted selenium’s role in colorectal cancer prevention, as it enhances DNA repair mechanisms, modulates apoptosis pathways, and suppresses tumor progression.12 Probiotic bacteria, particularly Bacillus subtilis, have been identified as efficient microbial systems for selenium biotransformation, allowing the production of functional probiotics enriched with organic selenium for human health applications.13
Traditional fermented foods are known to harbor diverse probiotic microorganisms, which confer numerous health benefits when consumed regularly. Koozh, a fermented millet-based porridge, is a staple traditional food in Tamil Nadu, India, and has been consumed for centuries due to its digestive, nutritional, and probiotic properties.14 Sorghum (Sorghum bicolor) is a highly nutritious gluten-free cereal rich in dietary fiber, polyphenols, and essential minerals. The natural fermentation of Koozh enhances nutrient bioavailability and promotes the growth of beneficial probiotic bacteria such as Lactic Acid Bacteria (LAB) and Bacillus species.15
Fermentation transforms complex carbohydrates into easily digestible bioactive compounds, enhancing gut health and promoting beneficial microbiota balance.16 Probiotic strains isolated from fermented foods demonstrate high resistance to acidic pH, bile salts, and enzymatic digestion, making them ideal candidates for gut health enhancement and functional food applications.17 The presence of probiotics in koozh makes it an excellent source of spore-forming probiotics, known for their heat stability, long shelf-life, and ability to colonize the human gut efficiently.18
This microbial biotransformation enhances selenium bioavailability, reduces toxicity, and improves its therapeutic properties.19 Selenium-enriched probiotics demonstrate stronger antioxidant activity, improved gastrointestinal survival, and enhanced functional properties, making them highly beneficial for gut health, immune modulation, and cancer prevention.20 Moreover, selenium-enriched probiotics have been shown to inhibit cancerous and pre-cancerous lesions, and enhance the effectiveness of conventional cancer treatments.21 This study aims to isolate and identify probiotic strains from fermented sorghum koozh, enrich the identified strain with selenium and bioaccumulation analysis, evaluate probiotic properties and assess anticancer activity on Caco-2 Cells.
Collection of fermented sorghum (Koozh)
Sample preparation
White Sorghum grains were procured from the local market in Guduvanchery, Tamil Nadu, India. The grains were carefully selected to ensure quality and uniformity. Before processing, they were thoroughly cleaned by washing them under running water to remove any dirt, dust, or impurities. This washing step is crucial to eliminate potential contaminants and ensure the grains are hygienic for consumption. After washing, the sorghum grains were soaked in distilled water for a duration of 12 hrs. Once the soaking period was completed, the water was drained, and the softened grains were transferred to a grinding unit. The grains were ground into a fine, smooth paste using a high-speed mixer grinder. The grinding process ensured uniformity in texture, making the paste suitable for further cooking (Figure 1).

Figure 1. (a) Raw Sorghum Grains Soaked in distilled water and (b) Fermented Sorghum koozh after 24 hrs of fermentation
The prepared sorghum paste was then subjected to heat treatment using a pressure cooker. The paste was carefully placed in the cooker, and a measured quantity of water was added to achieve the desired consistency. The pressure cooker was sealed, and the mixture was cooked under controlled conditions for a duration of 1-3 whistles. The number of whistles was monitored to ensure proper cooking, preventing undercooking or overcooking, which could affect the final texture and taste. After cooking, the pressure was allowed to release naturally before opening the cooker. The cooked sorghum paste was then ready for further use in food formulations, dietary applications, or sensory evaluation studies.
Fermentation process
The cooked paste was left to ferment at the temperature (30-35 °C) for 24 hrs. Natural fermentation encouraged probiotic bacterial growth, enhancing nutrient bioavailability and flavor. Freshly fermented sorghum porridge was kept in sterile containers at 4 °C and brought to the laboratory within 24 hrs for microbiological investigation. After fermentation, pH and microbial load were measured before proceeding with bacterial isolation.
Isolation of Bacillus subtilis
Bacterial isolation and culture conditions
MRS agar (de Man, Rogosa, and Sharpe agar) serves as a selective medium for lactic acid bacteria and Bacillus species. In the serial dilution procedure, 1 mL of the fermented Koozh sample was diluted in sterile saline (0.85% NaCl) and plated onto MRS agar plates. The plates were then incubated at 37 °C for 24-48 hrs under aerobic conditions.
Morphological and biochemical characterization
Gram staining confirmed the presence of Gram-positive rod-shaped bacteria. Catalase and oxidase tests were performed to differentiate Bacillus subtilis from lactic acid bacteria. Additionally, carbohydrate fermentation tests using glucose, lactose, sucrose, and maltose were conducted to confirm the fermentation capability of the isolates.
16S rRNA gene sequencing
The 16S rRNA gene was amplified and sequenced to confirm the identified strain. Genomic DNA was extracted using the Qiagen DNeasy Blood and Tissue Kit with modifications to enhance bacterial DNA yield, and its quality and concentration were assessed by NanoDrop spectrophotometer. The V3-V4 region of the 16S rRNA gene, commonly used for identifying lactic acid bacteria, was amplified using primer pairs 341F and 805R. PCR was performed in a 25 µL reaction mixture containing 2x KAPA HiFi HotStart ReadyMix, 0.2 µM of each primer, and 10 ng of DNA template, following a thermal cycling protocol that included an initial denaturation at 95 °C for 3 mins, 25 cycles of denaturation at 95 °C for 30 seconds, annealing at 55 °C for 30 seconds, and extension at 72 °C for 30 seconds, with a final extension at 72 °C for 5 mins. The PCR products were purified using AMPure XP beads and verified by gel electrophoresis and a Qubit fluorometer. Sequencing libraries were prepared using the Nextera XT DNA Library Preparation Kit and sequenced on an Illumina MiSeq platform with 2×300 bp paired-end reads. Raw sequencing reads were quality-controlled using Trimmomatic, while the DADA2 pipeline in QIIME2 was used for denoising, chimera removal, and amplicon sequence variant (ASV) identification. The final sequences were compared with the NCBI GenBank database for accurate strain identification.
Selenium enrichment procedure
The selenium content in bacterial biomasses was determined using UV-Vis molecular absorption spectroscopy following the method described by Pant et al.22 To establish a calibration curve, aliquots of selenium (Se) in concentrations ranging from 0.025-0.250 mg/L were carefully drawn from a stock solution. These aliquots were then transferred into a series of 10 mL volumetric flasks to ensure consistent and accurate preparation of standard solutions.
To facilitate selenium detection, each flask received 1 mL of 2% potassium iodide (KI) solution (prepared as a mass/volume solution) and 1 mL of 2 M hydrochloric acid (HCl). The contents were gently swirled, allowing the reaction to proceed until a distinct yellow coloration appeared, which signified the release of iodine. This color change is an essential step, indicating the successful oxidation-reduction reaction that prepares selenium for spectroscopic analysis.
Next, 200 µL of 1% starch solution prepared as a (mass/volume solution) was added to each volumetric flask. Starch acts as an indicator by forming a blue-black complex with free iodine, enhancing the sensitivity and visibility of the reaction. Following this stage, each flask was filled with ultrapure water to bring the total volume up to 10 mL, guaranteeing consistent sample preparation.23
The prepared solutions were analysed using a UV-Vis molecular absorption spectrophotometer, with the absorbance measured at 589 nm. This specific wavelength was selected to maximize the detection of the selenium-iodine complex, ensuring precise quantification. To enhance reliability, the same protocol was applied to all bacterial biomass samples, and each measurement was performed in triplicate to account for any experimental variability.
The presence and enrichment of selenium in bacterial cultures were confirmed through a visible color change from white to orange. This transformation is a characteristic indicator of selenium uptake and accumulation by bacteria, providing a straightforward visual cue to supplement the spectrophotometric analysis.
Probiotic characterization
Assessment of Survival and Adaptability of probiotics in Simulated Gastrointestinal Conditions and Determination of Bile Salt Tolerance.
Gastrointestinal tolerance
The bacterial strain (1 mL suspension) was incubated with 9 mL of artificial gastric juice (0.2 g NaCl and 1 g of pepsin in 100 mL HCl, pH 2.0) at 37 °C for 120 mins. Samples were taken at 0, 30, 60, 90, and 120 mins to assess viable counts (CFU/mL). The bacteria were then transferred to 9 mL of artificial intestinal fluid (0.68 g KH2PO4 and 1 g of trypsin in 100 mL distilled water, pH 7.0) and incubated at 37 °C for up to 240 mins, with samples taken at 150, 180, 210, and 240 mins.
To assess bile salt tolerance, bacterial suspensions were prepared, and initial CFU/mL was determined through serial dilution and plating. The suspensions were then inoculated into media containing 0% (control), 0.3%, and 0.5% (w/v) oxgall and incubated at 37 °C for 24 hrs. Final bacterial counts were determined, and the survival rate (%) was calculated as:
Survival rate = [(Initial CFU/mL) / (Final CFU/mL)] × 100 … 1
To evaluate survival in simulated gastrointestinal conditions, selenium-enriched Bacillus subtilis was exposed to simulated gastric juice (pH 2.0, pepsin 3 mg/mL) for 120 mins, followed by exposure to simulated intestinal juice containing bile salts (0.3%-0.5%) for 4 hrs. Viability was assessed by plating bacterial suspensions on MRS agar after exposure.
Bile salt tolerance
Bacterial suspensions were prepared and their initial CFU/mL assessed. The strain was inoculated into media containing 0% (control), 0.3%, and 0.5% (w/v) oxgall and incubated at 37 °C for 24 hrs. Post-incubation, viable counts were determined, and survival rates were calculated using the same formula. All tests were conducted in triplicate, and statistical analysis was applied to assess the consistency and significance of bacterial survival across different conditions. This provides a reliable evaluation of probiotic strains ability to endure harsh gastrointestinal conditions, which is essential for determining their viability as probiotic supplements.
Carbohydrate fermentation profiles
Carbohydrate broths containing phenol red and a specific carbohydrate (glucose, lactose, sucrose, or mannitol) were prepared, each with a Durham tube to capture gas. Pure probiotic cultures have been infused and incubated for 24-48 hrs at 37 °C. Fermentation produced organic acids and/or gases, lowering the pH and turning the phenol red indicator yellow. Gas production was detected by bubble formation in the Durham tube. Results were analysed based on color change and gas presence.
Hydrophobicity and adhesion properties
The key characteristic of successful probiotics is their capacity to bind to the intestinal epithelium, which allows gut colonisation, competitive exclusion of pathogens, and immune system modulation. The initial step in probiotic adhesion is surface hydrophobicity, which influences bacterial interactions with intestinal mucus and epithelial cells. In this study, the hydrophobicity of selenium-enriched Bacillus subtilis was assessed using the microbial adhesion to hydrocarbons (MATH) assay. The bacterial culture was suspended in phosphate-buffered saline (PBS) and mixed with xylene, followed by optical density (OD) measurements at 600 nm before and after phase separation.
Lyophilization process for probiotic preservation
The bacterial culture was first centrifuged to separate the cells from the medium, forming a dense pellet while the supernatant was discarded. To protect the cells during freezing and drying, the pellet was resuspended in a cryoprotectant solution containing 10% skim milk and 5% sucrose. The mixture was then flash-frozen using liquid nitrogen. Next, the frozen suspension underwent freeze-drying at -50 °C to -80 °C under a vacuum of 0.01 mbar for 12 hours. This process removed moisture through sublimation without damaging the cells. The resulting dried probiotic powder was collected and stored in sterile vials at 4 °C to preserve bacterial viability and stability for further research.
In vitro analysis in Caco-2 cell line
Cytotoxicity study
In 1 mg of lyophilised sample was mixed with sterile complete media and diluted to the desired concentration. Caco-2 cancer cells (5 × 104 cells per well) were seeded into 96 well plates and incubated for 24 hrs for adhesion. The cells were then treated with varying concentrations (0-512 mg/mL) of the selenium-enriched lyophilized strain for 24 hrs. After treatment, the strain was removed, and the wells were rinsed with phosphate-buffered saline (PBS) to eliminate any residual components.
Wells were treated with 5 mg/mL MTT solution and incubated for 4 hrs to allow purple formazan crystal formation. The MTT solution removed, and 100 µL of dimethyl sulfoxide (DMSO) was added to dissolve the crystals and were evaluated using ELISA plate reader at 570 µm (BIORAD). The percentage of cell viability was calculated using the formula:
Cell viability (%) = [Absorbance of control / Absorbance of Test] × 100 …2
AO/EtBr staining for apoptosis analysis
To visualize nuclear changes and apoptotic body formation, 3 × 104 Caco-2 cells were seeded in 6-well plates, incubated in a CO2 chamber, and treated with the sample at its IC50 concentration for 24-48 hrs. After incubation, the media was removed, and cells were washed twice with DPBS. A 20 µL acridine orange and ethidium bromide stain (100 µg/mL, 1:1) was added, followed by 30 mins of dark incubation. Excess stain was washed off with PBS, and apoptosis was analyzed under a fluorescent microscope. Non-treated cells served as the control.
Apoptotic cell percentage was calculated using:
Apoptic cells (%) = [Total number of apoptotic cells / Total number of Live and Dead cells] × 100 … 3
Statistical analysis
Statistical analysis and graphs were generated using GraphPad Prism 8.4. The MTT assay and other in vitro experiments were repeated at least three times for accuracy. Data were presented as mean ± standard deviation (SD). Statistical tests were used to determine significance, with a p-value ≤0.05 considered statistically significant.
Isolation and identification of Bacillus subtilis
Accurate identification of probiotic bacterial strains is crucial for their effective use in functional foods, gut health improvement, and disease prevention. This study successfully isolated Bacillus subtilis subsp. stercoris from fermented sorghum (Koozh), a traditional Tamil beverage known for its probiotic-rich composition. The strain was identified using morphological, biochemical, and molecular techniques (Figures 2 and 3).
Figure 2. Initial Isolation of probiotics from Koozh
Figure 3. Growth of Bacillus subtilis subsp. stercoris
Morphological analysis confirmed Bacillus characteristics, including rod-shaped, Gram-positive, endospore-forming cells. Biochemical tests validated its identity, showing catalase positivity, starch hydrolysis, and carbohydrate fermentation patterns consistent with Bacillus subtilis subsp. stercoris. Molecular identification through 16S rRNA sequencing confirmed a high sequence similarity with reference strains (Figure 4). The combination of these methods ensured accurate strain identification, highlighting its potential as a probiotic for functional food applications and gut health benefits.

Figure 4. 16S rRNA sequencing result of Bacillus subtilis subsp. stercoris
Morphological and biochemical characterization
Bacterial strains were isolated on MRS agar and analyzed for morphology using a light microscope. The colonies were circular, smooth, cream-colored, and opaque, characteristic of Bacillus species. Gram staining confirmed a Gram-positive, rod-shaped bacterium with endospore-forming ability, a key trait of Bacillus subtilis.24 The cells were arranged in chains or clusters, enhancing resistance to harsh conditions.
Biochemical tests confirmed a positive catalase reaction, indicating the strain’s ability to break down hydrogen peroxide, a common trait in aerobic bacteria like Bacillus subtilis.25 The oxidase test was negative, distinguishing it from other Bacillus species. Carbohydrate fermentation tests showed efficient glucose and maltose fermentation, producing acid without gas, demonstrating metabolic adaptability.26 The strain also exhibited strong bile salt (0.3%-0.5%) and gastric tolerance, key indicators of probiotic potential.27
Molecular identification through 16S rRNA sequencing
The 16S rRNA sequencing analysis of selenium-enriched strains isolated from fermented samples revealed the taxonomic composition of the microbial community. Colonies subjected to 16S rRNA sequencing were identified as representatives of the Bacillus genera (Figure 4).
Bacillus species have been frequently isolated from fermented sorghum, known for their robustness and enzymatic production capabilities that facilitate the fermentation process. Research by Poornachandra et al identified Bacillus subtilis strains from fermented sorghum, demonstrating their probiotic potential through notable enzyme production and antimicrobial activities.28 Furthermore, Ilango and Antony reported the isolation of numerous probiotic strains from fermented sorghum, with Bacillus subtilis being prominently identified in their findings.14 The results of this study revealed the presence of Bacillus subtilis subsp. stercoris, consistent with previous research that has also identified Bacillus species, including Bacillus subtilis, in similar fermentation processes. This discovery further underscores the consistent occurrence of beneficial Bacillus strains in fermented sorghum koozh and highlights their potential probiotic applications.
Selenium enrichment and bioaccumulation
The enrichment of Bacillus subtilis subsp. stercoris with selenium was visually confirmed by a distinct color change from white to orange, a well-documented indicator of organic selenium compound formation. This transformation signifies selenium uptake and metabolic conversion, wherein bacterial cells reduce inorganic selenium (sodium selenite, Na2SeO3) into bioavailable organic selenium species such as selenomethionine, selenocysteine, or elemental selenium.29
Initially, Bacillus subtilis subsp. stercoris, colonies grown on MRS agar appeared creamy or white due to their natural pigmentation. However, when cultivated in MRS broth supplemented with 0.5 mg/L sodium selenite, the bacterial culture gradually changed from light yellow to an intense orange-red color within 32 hrs of incubation at 37 °C. This colour transition serves as a visual marker of selenium biotransformation and accumulation.
Further quantitative analysis of selenium uptake in bacteria isolated from fermented samples confirmed selenium metabolism and bioaccumulation. Among the isolates, Bacillus subtilis subsp. stercoris demonstrated a selenium concentration of 150 µg/g dry weight, with an absorbance of 0.450 AU, indicating moderate selenium accumulation. These findings suggest that Bacillus subtilis subsp. stercoris effectively assimilates and metabolizes selenium, potentially contributing to its role in probiotics, or bioremediation applications (Figure 5).
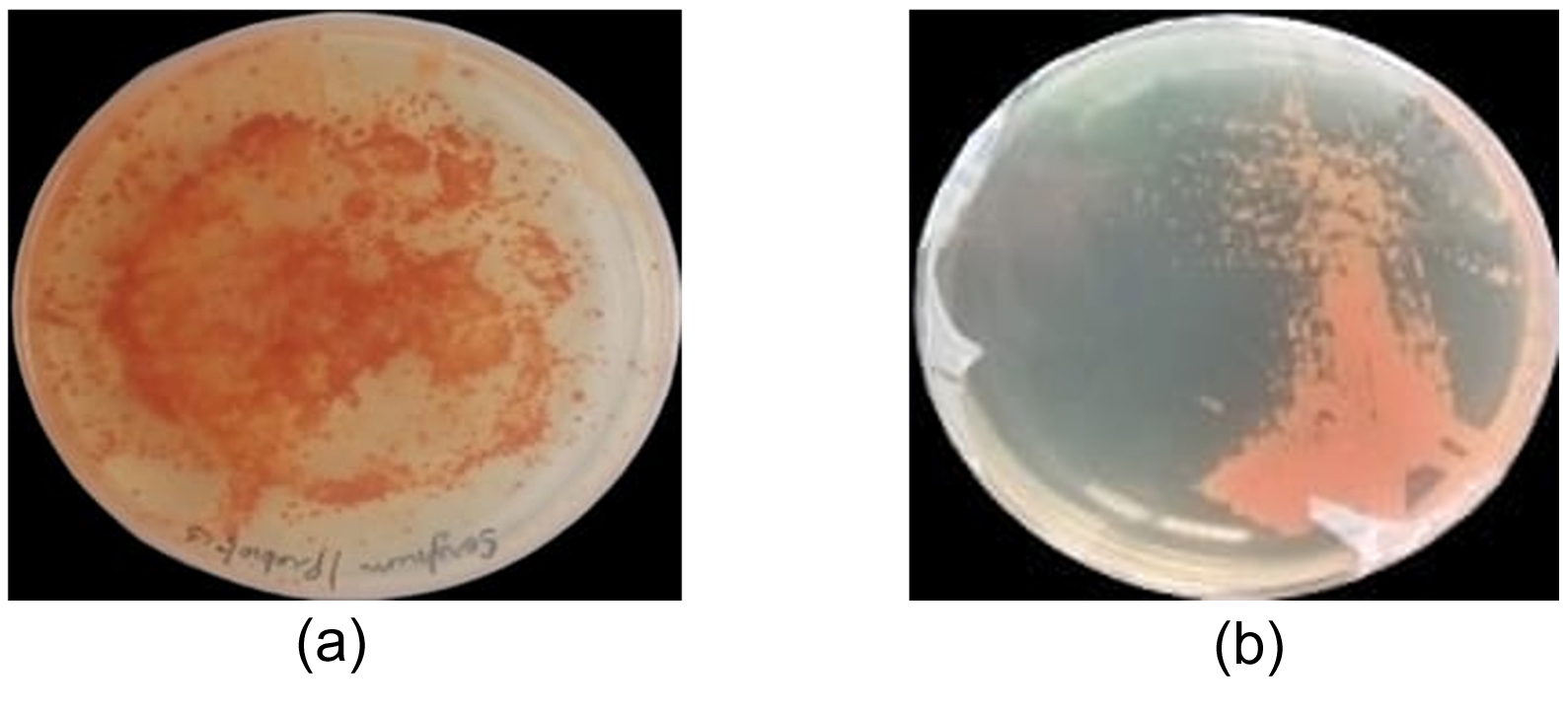
Figure 5. (a) & (b) Selenium-Enriched Bacillus subtilis showing orange pigmentation and colonies
UV-vis spectroscopy analysis
To quantitatively confirm selenium uptake UV-Vis spectroscopy was performed on selenium-enriched Bacillus subtilis subsp. stercoris cultures. This technique is widely used to detect characteristic absorbance peaks associated with selenium typically observed in the 260-300 nm range. After 24 hrs of incubation in selenium-supplemented media, analysis of the culture supernatant revealed a strong absorbance peak at approximately 270 nm, indicating the intracellular biosynthesis of selenium. The peak at ~270 nm is a chemical fingerprint, indicating selenium enrichment. UV-Vis spectral analysis also demonstrated that the selenium enriched Bacillus subtilis subsp. stercoris remained stable, with no significant peak shifts over extended incubation periods.
Carbohydrate fermentation profiles
Fermentation of dietary carbohydrates produces organic acids like acetic and lactic acid, which reduce intestinal pH, suppress pathogenic microorganisms, and improve nutrient absorption. In this study, the selenium-enriched Bacillus subtilis subsp. stercoris strain was evaluated for its ability to ferment various carbohydrates, including glucose (monosaccharide), lactose and maltose (disaccharides), sucrose (common dietary sugar), and mannitol (polyol sugar with prebiotic potential). The results showed that Bacillus subtilis subsp. stercoris efficiently fermented glucose, sucrose, lactose and maltose, producing significant amounts of acid but no gas, a desirable trait for probiotic strains. However, no fermentation was observed for mannitol. Fermentation efficiency was further assessed by measuring the pH drop around 4.5 after 24 hrs of incubation in carbohydrate broth, where a significant decrease for glucose, sucrose, lactose and maltose, confirmed the production of lactic acid and other beneficial metabolites that contribute to gut health.
Survival in simulated gastric and intestinal conditions
A key criterion for probiotic selection is survival in the gastrointestinal tract, where acidic pH and bile salts pose major challenges to microbial colonization. The stomach’s acidic environment (pH 1.5-3.0) aids digestion but threatens bacterial viability, while bile salts (0.3%-0.5%) in the small intestine can disrupt cell membranes and inhibit colonization. In this study, the acid tolerance of selenium-enriched Bacillus subtilis subsp. stercoris was assessed by incubating it in simulated gastric juice (pH 2.0, 3 mg/mL pepsin) and measuring survival at 30, 60, and 120 mins. The strain maintained ~85% viability after 2 Hrs, demonstrating its ability to withstand gastric acidity. Bile salt tolerance was evaluated by exposing the strain to MRS broth with 0.3%-0.5% oxgall for 4 hrs, where it retained over 80% viability, indicating strong adaptation to intestinal conditions. These high survival rates confirm the strain’s resilience, supporting its potential as a robust probiotic capable of withstanding digestive stress and exerting beneficial effects in the intestine.
Hydrophobicity and adhesion properties
The results showed that the strain exhibited a hydrophobicity of approximately 65%-70%, which is classified as moderate to high, indicating strong cell surface properties that enhance its ability to adhere to the intestinal lining. These findings suggest that selenium-enriched Bacillus subtilis possesses favourable adhesion characteristics, further supporting its potential as a robust probiotic strain for gut health applications.
Cytotoxicity studies on Caco-2 cells (MTT Assay)
The antiproliferative effects of selenium enriched-probiotics on cell viability were examined in Caco-2 colorectal cancer cell lines by MTT assay. These results indicated that the exposure of probiotics at concentrations of 1-512 µg/mL for 48 hrs significantly reduced the cell viability of Caco-2 cells in a dose dependent manner. The IC50 values of different concentration of (1, 2, 4, 8, 16, 32, 64, 128, 256 and 512) selenium enriched-probiotics for Caco-2 cells at 48 h was 95.4 ± 0.120 µM, 91.81 ± 0.08 µM, 87.45 ± 0.118 µM, 78.31 ± 0.905 µM, 67.58 ± 0.255 µM, 56.261 ± 0.061 µM, 50. 63 ± 0.115 µM, 44.63 ± 0.292 µM, 38.69 ± 0.359 µM, and 30.78 ± 0.392 µM, respectively. Moreover, the results showed that selenium enriched-probiotics treatment reduced Caco-2 cell viability in a concentration-dependent manner. Furthermore, the results revealed that the probiotic strains significantly reduced cell viability in colorectal cancer cells. The results demonstrated a dose-dependent cytotoxic effect, where increasing concentrations of selenium-enriched Bacillus subtilis subsp. stercoris led to a progressive decline in Caco-2 cell viability. The half-maximal inhibitory concentration (IC50) was determined to be 81.517 µg/mL, suggesting significant cytotoxicity at lower concentrations (1-8 µg/mL), cell viability remained above 90%, indicating minimal cytotoxicity and suggesting that low-dose selenium-enriched probiotics do not significantly disrupt normal cellular metabolism (Figure 6). However, at higher concentrations (16-512 µg/mL), a sharp decline in cell viability was observed, highlighting a direct correlation between selenium exposure and cancer cell growth inhibition.
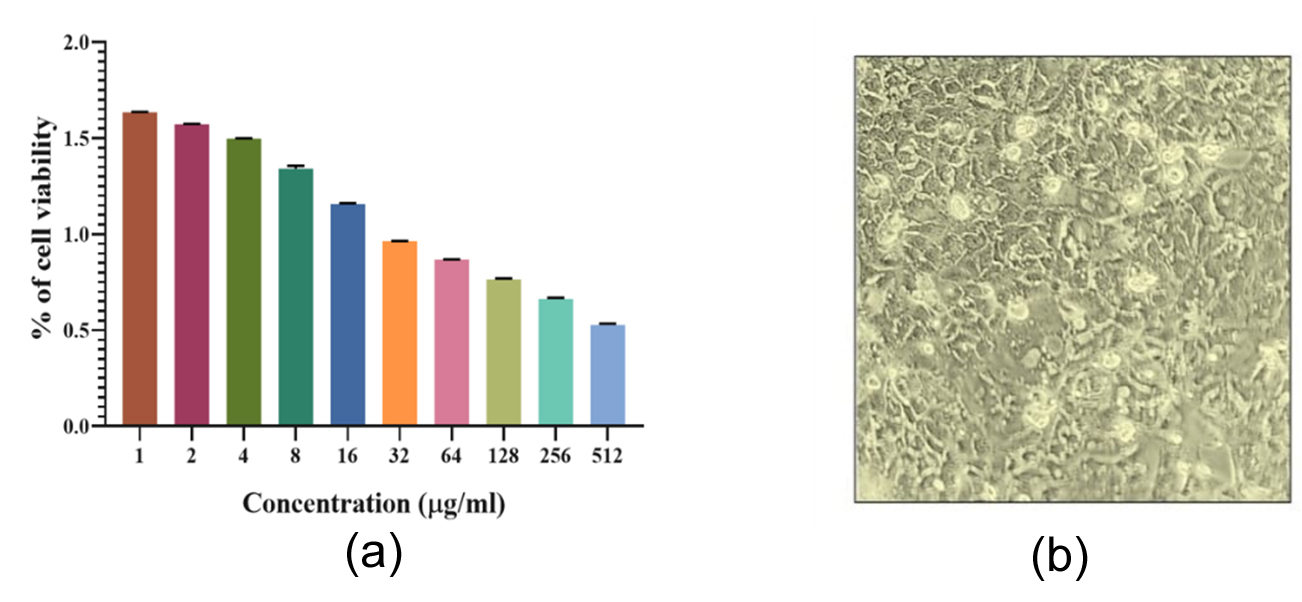
Figure 6. (a) The cell cytotoxicity assay image depicts Caco-2 cells treated with selenium-enriched probiotics at varying concentrations (1, 2, 4, … up to 512 µg/mL) and (b) 10X magnification of control Caco-2 cells with 90% confluency
This effect is likely mediated by selenium-induced oxidative stress and apoptosis activation, which disrupts mitochondrial function and triggers programmed cell death in cancerous cells while sparing normal cells. These findings suggest that selenium-enriched probiotics may serve as a selective anticancer agent, offering potential applications in colorectal cancer prevention and adjunct therapy.
Apoptotic cell analysis (AO/EtBr staining)
Caco-2 cells were treated with selenium-enriched probiotic extracts at IC50 value of selenium enriched probiotic for 24 and 48 Hrs, followed by AO/EtBr staining and fluorescence microscopy analysis. The control group (untreated cells) exhibited a higher proportion of apoptotic and necrotic cells, with orange-red nuclear staining, chromatin condensation, and membrane blebbing—hallmarks of apoptosis. Some control cells displayed intense red fluorescence, indicating necrotic cell death due to oxidative stress and DNA damage in the tumor microenvironment. In contrast, selenium-enriched probiotic-treated cells showed a lower Percentage of apoptotic cells, with the majority exhibiting bright green fluorescence, indicating high cell viability (Figure 7).
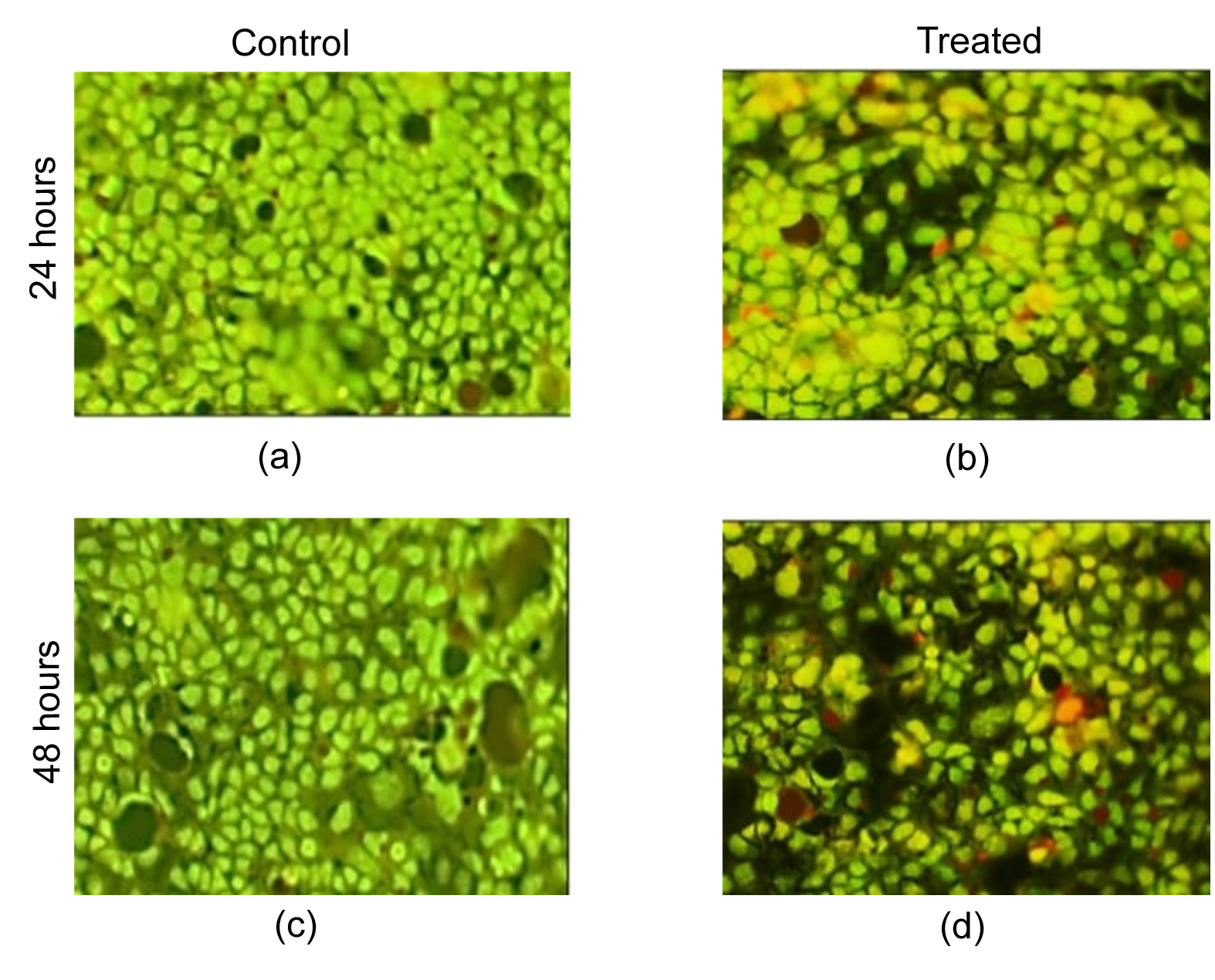
Figure 7. Fluorescent microscopic images of AO/EtBr staining at varying concentrations (1, 2, 3, 4, … up to 512 µg/mL). (a) & (c) represent the control groups at 24 and 48 Hrs, respectively. (b) & (d) show selenium-enriched probiotics-treated Caco-2 cells at 24 and 48 Hrs, respectively
The study began by isolating selenium-enriched strains of Bacillus subtilis subsp. stercoris from fermented sorghum (koozh), a traditional food consumed in Tamil Nadu, India. Using selective culture techniques and 16S rRNA gene sequencing, researchers accurately identified the bacterial species. The isolated strains demonstrated significant selenium uptake when cultured in selenium-supplemented media, confirming their ability to bioaccumulate selenium. This finding suggests potential applications in producing selenium-fortified fermented foods to address selenium deficiency and improve nutritional health.
The presence of Bacillus subtilis in fermented sorghum has been well-documented in previous studies. Research by Poornachandra et al. identified Bacillus subtilis strains in fermented sorghum and highlighted their probiotic potential due to enzyme production and antimicrobial activity.28 Similarly, Ilango and Antony reported multiple probiotic strains, including Bacillus subtilis, in fermented sorghum, confirming its consistent occurrence in such traditional foods.14 The identification of Bacillus subtilis subsp. stercoris in this study aligns with these findings, reinforcing its potential as a beneficial probiotic.
Quantification of selenium uptake among different bacterial species revealed variations in bioaccumulation efficiency. Previous research by Morschbacher et al. demonstrated that bacteria such as Enterococcus faecalis, Lactobacillus parabuchneri, L. paracasei, and L. plantarum convert selenite into elemental selenium, accumulating high intracellular selenium levels.30 This study supports similar conclusions, showing that Bacillus subtilis subsp. stercoris can efficiently transform selenium into a less toxic form while storing high concentrations within its cells.
Carbohydrate fermentation tests were conducted to assess the metabolic potential of the isolated Bacillus strains. Research by Simon et al. showed that Bacillus species could ferment 11 out of 49 tested carbohydrates, including D-ribose, L-arabinose, D-xylose, D-glucose, D-fructose, and D-mannitol.31 Similarly, Nithya & Halami found that Bacillus strains isolated from various food sources demonstrated diverse carbohydrate fermentation capabilities, which are critical for their probiotic functionality. The findings from this study confirm that Bacillus subtilis subsp. stercoris possesses significant carbohydrate fermentation abilities, supporting its role as a probiotic strain.32
To evaluate probiotic potential, the ability of Bacillus subtilis subsp. stercoris to survive gastrointestinal conditions was tested. Research by Lee et al and Khatri et al. demonstrated that probiotic strains possess mechanisms to withstand acidic stomach conditions and bile salts, often facilitated by spore formation.2,33 Additionally, studies by Chen et al. and Patel et al. confirmed that Bacillus species exhibit significant bile salt tolerance, ensuring survival in the digestive tract.8,34 This study aligns with these findings, confirming that the isolated probiotic strain can endure the harsh conditions of the gastrointestinal tract, reinforcing its suitability for probiotic applications.
The potential anticancer properties of selenium-enriched Bacillus subtilis subsp. stercoris were evaluated using the MTT assay, a method that assesses cell viability and metabolic function (Figure 6). Previous studies have reported similar findings: Cheng et al. demonstrated that Fengycin, a compound extracted from Bacillus subtilis, inhibited HT29 colon cancer cell growth by inducing apoptosis.35 Likewise, Sirpu et al. found that a crude extract of Bacillus subtilis exhibited strong antiproliferative effects against human breast cancer cells.36 In the present study, selenium-enriched Bacillus subtilis subsp. stercoris triggered the anti-proliferation of Caco-2 colon cancer cells, reducing cell viability in a dose-dependent manner. At higher selenium concentrations, cell viability declined sharply, followed by a stabilization phase, indicating a saturation threshold. These findings support the anticancer potential of selenium-enriched Bacillus subtilis subsp. stercoris.
To further confirm apoptotic effects, Acridine Orange/Ethidium Bromide (AO/EtBr) staining was performed. This fluorescence-based technique differentiates live, apoptotic, and necrotic cells based on membrane integrity and chromatin changes. AO stains live and early apoptotic cells green, while EtBr penetrates compromised membranes, staining late apoptotic and necrotic cells orange-red. The results revealed that selenium-treated cancer cells exhibited nuclear condensation, chromatin fragmentation, and increased EtBr uptake, indicating programmed cell death rather than necrotic damage. A dose-dependent increase in apoptotic (orange-stained) cells was observed at higher concentrations (>100 µg/mL), yet with significantly fewer necrotic (red-stained) cells compared to the untreated group. This suggests that selenium-enriched probiotics selectively induce apoptosis in cancer cells while preserving normal cell viability (~85%), supporting their potential as targeted anticancer agents.
There is currently no research has been conducted on the apoptotic effects of Bacillus subtilis subsp. stercoris using Acridine Orange/Ethidium Bromide (AO/EtBr) staining, making this study a pioneering effort in validating its apoptotic potential. This research provides the first documented evidence demonstrating the strain’s ability to induce apoptosis in colorectal cancer (Caco-2) cells. The use of AO/EtBr staining allowed for the visualization of characteristic apoptotic features, such as chromatin condensation, nuclear fragmentation, and membrane integrity loss, confirming selective cytotoxicity against cancerous cells while preserving the viability of non-cancerous cells. These findings establish a critical foundation for further investigation into the strain’s potential as a natural anticancer agent and highlight its relevance in functional food and nutraceutical applications aimed at colorectal cancer prevention and management.
This study provides compelling evidence that selenium-enriched Bacillus subtilis isolated from fermented sorghum (Koozh) possesses significant probiotic potential, selenium biotransformation capability, and anticancer properties. The strain demonstrated efficient conversion of inorganic sodium selenite (Na2SeO3) into bioavailable organic selenium confirmed by the characteristic color transition and UV-Vis spectroscopy analysis. Its strong probiotic characteristics, including high tolerance to gastric and bile conditions, carbohydrate fermentation ability, and adhesion to intestinal epithelial cells, highlight its potential for functional food applications. Notably, the selenium-enriched strain exhibited dose-dependent cytotoxicity against colorectal cancer (Caco-2) cells, with an IC50 value of 81.517 µg/mL, indicating moderate anticancer activity. AO/ETBR staining further validated the selective induction of apoptosis in cancerous cells while maintaining high viability in non-cancerous cells, suggesting a safe and targeted anticancer mechanism. The integration of selenium-enriched probiotics into functional foods and nutraceuticals offers a novel strategy for disease prevention, immune system modulation, and gut health enhancement.
In conclusion, the selenium-enriched Bacillus subtilis isolated from fermented sorghum (Koozh) exhibits promising probiotic potential, efficient selenium biotransformation, and moderate anticancer activity. Its ability to tolerate gastric conditions, adhere to intestinal cells, and selectively induce apoptosis in colorectal cancer cells underscores its potential as a functional food ingredient. The findings highlight the role of selenium-fortified probiotics in gut health, immune modulation, and disease prevention. Given the increasing demand for natural, health-promoting food ingredients, these findings support the potential of selenium-fortified probiotics as innovative dietary interventions for colorectal cancer management and overall well-being. Further in vivo studies and clinical trials are warranted to validate its therapeutic applications and enhance its integration into nutraceuticals for colorectal cancer management and overall well-being.
ACKNOWLEDGMENTS
The authors express their sincere gratitude to Dr. S. Periyar Selvam, Head of the Department of Food Process Engineering, School of Bioengineering, SRM Institute of Science and Technology (SRMIST), Kattankulathur, and to Dr. C. Meenakshisundaram, Associate Professor, Department of Microbiology, SRM Medical College Hospital, SRMIST, for their valuable support and guidance. The authors gratefully acknowledge the financial support provided by SRM Medical College Hospital and Research Centre, Faculty of Medicine and Health Sciences, SRMIST, Kattankulathur, for defraying the publication costs of this article.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
NG and SS conceptualized the study. NG, LSK and VS design the experiments. NG and LSK carried out the in vitro evaluation experiments. NG and VS performed isolation and characterization of the strains and data analysis. SK assisted in Biochemical test and data analysis. NG and LSK performed result interpretation. SS supervised the study. NG wrote the manuscript. SS and SK revised the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Pandey H, Tang DWT, Wong SH, Lal D. Gut microbiota in colorectal cancer: biological role and therapeutic opportunities. Cancers (Basel). 2023;15(3):866.
Crossref - Lee NK, Kim WS, Paik HD. Bacillus strains as human probiotics: characterization, safety, microbiome, and probiotic carrier. Food Sci Biotechnol. 2019;28(5):1297-1305.
Crossref - Nowak A, Paliwoda A, Blasiak J. Anti-proliferative, pro-apoptotic and anti-oxidative activity of Lactobacillus and Bifidobacterium strains: a review of mechanisms and therapeutic perspectives. Crit Rev Food Sci Nutr. 2019;59(21):3456-3467.
Crossref - Sivamaruthi BS, Kesika P, Chaiyasut C. The role of probiotics in colorectal cancer management. Evid Based Complement Alternat Med. 2020;2020:3535982.
Crossref - Rayman MP. The importance of selenium to human health. Lancet. 2000;356(9225):233-241.
Crossref - Pecoraro BM, Leal DF, Frias-De-Diego A, Browning M, Odle J, Crisci E. The health benefits of selenium in food animals: a review. J Anim Sci Biotechnol. 2022;13(1):58.
Crossref - Zhang F, Li X, Wei Y. Selenium and selenoproteins in health. Biomolecules. 2023;13(5):799.
Crossref - Chen Z, Lu Y, Dun X, Wang X, Wang H. Research progress of selenium-enriched foods. Nutrients. 2023;15(19):4189.
Crossref - Genchi G, Lauria G, Catalano A, Sinicropi MS, Carocci A. Biological activity of selenium and its impact on human health. Int J Mol Sci. 2023;24(3):2633.
Crossref - Gnanaprakash J, Jabastin J, Vijayasarathy M, Rajendran S, Devi PB. Functional traits of selenium accumulating lactic acid bacteria as a probiont from fermented foods of an Indian tribal community. J Agric Food Res. 2023;12:100601.
Crossref - Ataollahi F, Amirheidari B, Amirheidari Z, Ataollahi M. Clinical and mechanistic insights into biomedical application of Se-enriched probiotics and biogenic selenium nanoparticles. Biotechnol Lett. 2025;47(1):1-21.
Crossref - Zhao S, Meng Y, Cai W, et al. Docosahexaenoic acid coordinating with sodium selenite promotes paraptosis in colorectal cancer cells by disrupting the redox homeostasis and activating the MAPK pathway. Nutrients. 2024;16(11):1737.
Crossref - Hyrslova I, Kana A, Kantorova V, Krausova G, Mrvikova I, Doskocil I. Selenium accumulation and biotransformation in Streptococcus, Lactococcus, and Enterococcus strains. J Funct Foods. 2022;92:105056.
Crossref - Ilango S, Antony U. Assessment of the microbiological quality of koozh, a fermented millet beverage. Afr J Microbiol Res. 2014;8(3):308-312.
Crossref - Roopashri AN, Savitha J, Divyashree MS, Mamatha BS, Rani KAU, Kumar A. Indian traditional fermented foods: the role of lactic acid bacteria. In: Marta L, eds. Lactobacillus – A Multifunctional Genus. IntechOpen. 2023.
Crossref - Toma MM, Pokrotnieks J. Probiotics as functional food: microbiological and medical aspects. Acta Univ Latviensis. 2006;710:117-129.
- Ayyash MM, Abdalla AK, AlKalbani NS, et al. Characterization of new probiotics from dairy and nondairy products—Insights into acid tolerance, bile metabolism and tolerance, and adhesion capability. J Dairy Sci. 2021;104(8):8363-8379.
Crossref - Raungrusmee S, Kumar SR, Anal AK. Probiotic cereal-based food and beverages, their production and health benefits. In: Panesar PS, Anal AK, eds. Probiotics, Prebiotics and Synbiotics: Technological Advancements Towards Safety and Industrial Applications. Hoboken, NJ: John Wiley & Sons; 2022:163-184.
Crossref - Ferro C, Florindo HF, Santos HA. Selenium nanoparticles for biomedical applications: From development and characterization to therapeutics. Adv Healthc Mater. 2021;10(16):2100598.
Crossref - Li Z, Dong Y, Chen S, et al. Organic selenium increased gilts antioxidant capacity, immune function, and changed intestinal microbiota. Front Microbiol. 2021;12:723190.
Crossref - Abedi J, Saatloo MV, Nejati V, et al. Selenium-enriched Saccharomyces cerevisiae reduces the progression of colorectal cancer. Biol Trace Elem Res. 2018;185:424-432.
Crossref - Pant R, Sharma N, Kabeer SW, Sharma S, Tikoo K. Selenium-enriched probiotic alleviates western diet-induced non-alcoholic fatty liver disease in rats via modulation of autophagy through AMPK/SIRT-1 pathway. Biol Trace Elem Res. 2023;201(3):1344-1357.
Crossref - Kumari K, Kashyap P, Chakrabarti P. Germination and probiotic fermentation: a way to enhance nutritional and biochemical properties of cereals and millets. Food Sci Biotechnol. 2024;33(3):505-518.
Crossref - Kyprianos S. Identification and characterization of a novel bacterial species found in the Lebanese soil [dissertation]. Lebanese American University. 2022.
- Saeed SWZ, Naseer I, Zahir ZA, et al. Bacillus strains with catalase enzyme improve the physiology and growth of rice (Oryza sativa L.). Stresses. 2023;3(4):736-748.
Crossref - Nandha MC, Shukla RM. Exploration of probiotic attributes in lactic acid bacteria isolated from fermented Theobroma cacao L. fruit using in vitro techniques. Front Microbiol. 2023;14:1274636.
Crossref - Söylemez-Milli N, Ertürkmen P, Baltakesmez DA. The resistance abilities of some Bacillus species to gastrointestinal tract conditions: whole genome sequencing of the novel candidate probiotic strains Bacillus clausii BA8 and Bacillus subtilis BA11. Food Sci Nutr. 2025;13(2):e70018.
Crossref - Rao KP, Chennappa G, Suraj U, Nagaraja H, Raj APC, Sreenivasa MY. Probiotic potential of Lactobacillus strains isolated from sorghum-based traditional fermented food. Probiotics Antimicrob Proteins. 2015;7:146-156.
Crossref - Mota LA, Da Silva APM, Da Silva EA, Leite GMFL, Calegari RP, Baptista AS. Ability of the Saccharomyces cerevisiae Y904 to tolerate and adapt to high concentrations of selenium. Rev Bras Eng Biossistemas. 2022;16.
Crossref - Morschaacher AP, Dullius A, Dullius CH, et al. Assessment of selenium bioaccumulation in lactic acid bacteria. J Dairy Sci. 2018;101(12):10626-10635.
Crossref - Simon A, Colom J, Mazhar S, Khokhlova E, Deaton J, Rea K. Bacillus megaterium Renuspore® as a potential probiotic for gut health and detoxification of unwanted dietary contaminants. Front Microbiol. 2023;14:1125616.
Crossref - Nithya V, Halami PM. Evaluation of the probiotic characteristics of Bacillus species isolated from different food sources. Ann Microbiol. 2013;63(1):129-137.
Crossref - Khatri I, Sharma G, Subramanian S. Composite genome sequence of Bacillus clausii, a probiotic commercially available as Enterogermina®, and insights into its probiotic properties. BMC Microbiol. 2019;19(1):1-15.
Crossref - Patel AK, Singhania RR, Pandey A, Chincholkar SB. Probiotic bile salt hydrolase: current developments and perspectives. Appl Biochem Biotechnol. 2010;162:166-180.
Crossref - Cheng W, Feng YQ, Ren J, Jing D, Wang C. Anti-tumor role of Bacillus subtilis fmbJ-derived fengycin on human colon cancer HT29 cell line. Neoplasma. 2016;63(2):215-222.
Crossref - Sirpu Natesh N, Arumugam M, Karanam G. Apoptotic role of marine sponge symbiont Bacillus subtilis NMK17 through the activation of caspase-3 in human breast cancer cell line. Mol Biol Rep. 2018;45(6):2641-2651.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.