


ISSN: 0973-7510
E-ISSN: 2581-690X
Phytases are enzyme capable of hydrolysing phytic acid to myo-inositol and inorganic phosphorus. The aim of the present study was the screening and isolation of efficient phytase producing bacteria from soil samples. 14 bacterial isolates were found to produce clear zone of hydrolysis on PSM containing 0.5% ca-phytate. Out of all phytase positive isolates, five bacterial isolates 100A, 9H, 14A, 4D, and 2B showed significant phytase production in submerged fermentation. The isolate 14A was found to be the best among all with the maximum production of 89.09 U/ml after 72 h of incubation at 30ºC. Colony morphology and microscopic examination (Gram’s staining) of maximal phytase producer suggested that the isolate 14A is gram negative rod. Further on molecular analysis the bacterial isolate showed closest similarity with Pseudomonas sp. 16S rDNA sequencing and sequence similarity search using BLAST showed that it matches 97 % with Pseudomonas pulluriniana, while 3% was found to be unique sequence and named as Pseudomonas sp. aazad. The crude phytase from the isolate showed optimum activity at pH 6 and temperature 50ºC.
Phytase, PSM, Pseudomonas sp., ca-phytate, fermentation.
Phytic acid is the major storage form of phosphorus and inostiol in plant seeds, comprising about 3-5% of the dry weight particularly in cereal grains and legumes (Zhang et al., 2010). Phytic acid can chelates divalent cations like calcium, magnesium, iron and zinc, decreasing their bioavailability. These minerals are essential in the diet of both humans and animals. Phytic acid has anti-nutritional effect since it reduce food intake and nutrient utilization in animals and humans (Urbano et al., 2000; Lei et al., 2003). Phytases are enzymes capable of hydrolyzing phytic acid to myo-inostiol and inorganic phosphorus. Phytases are mainly used as animal feed additive, as they enhance the nutritional quality of plant material in feed by increasing the bioavailability of minerals for monagastric animals. Moreover, animal feeds devoid of phytase causes immense release of undigested phytate into the environment. The phosphorus excreted in the environment enhances the growth of phosphorus assimilating microorganisms resulting in eutrophication (Schroder et al., 1996).
Phytases are found in plants, animals and microorganisms. However, researchers have shown that microbial phytases are most promising for biotechnological application (Angelis et al., 2003). Studies have shown that bacterial phytases offer several advantages over fungal phytases because of higher substrate specificity, greater resistance to proteolysis and better catalytic capability. There is an ongoing interest in microorganisms including bacteria for novel and efficient phytases as there is no single phytase that may be able to meet the diverse need for all commercial and environmental applications. The present study reports isolation, production and molecular identification of efficient phytate degrading bacteria, Pseudomonas sp.
Sample collection
Isolation of the phytase-producing bacteria was carried out by sampling 150 soil samples from different geographical regions. Samples were collected from 10 cm below soil surface aseptically with the help of sterile spatula in sterile polythene bags. Three serial dilutions of each sample were prepared (10-1, 10-3, and 10-5) in normal saline. 100 µl of each dilution were inoculated on freshly prepared nutrient agar plates having pH of (4.0, 7.0 and 10.0) and incubated at 10ºC (24-96 h), 30 ºC (24–72 h) and 60ºC (12–24 h). Isolated colonies of different morphological characteristics (variations in size, shape, color and regularity) were sub-cultured on fresh nutrient agar medium plates at neutral pH.
Isolation of phytase producing bacteria
The bacterial isolates to be screened for phytase production were point inoculated in phytase screening medium (15 g-1 Glucose, 5.0 gl-1 NH4 NO3, 0.5 gl- 1 KCl, 0.5 gl-1 MgSO4. 7H2O, 0.01 gl-1 FeSO4.7H2O, 0.01 gl-1 MnSO4.7H2 O, 0.5 % Ca-phytate,20.0 gl-1 Agar; pH adjusted to 5.5) and incubated at 30 ºC for 24-48 h. Bacterial colonies showing the zone of hydrolysis on the solid plates indicate phytase activity. The isolates with distinct zone of hydrolysis were considered positive and selected for further study on the basis of size of zone (mm).
Phytase enzyme assay
Phytase producing bacterial isolates were inoculated in 50ml PSM broth containing 0.5% calcium phytate and were cultured in a rotary shaker (200rpm) at 30ºC for 72hr. 2ml cultures were withdrawn aseptically from flasks periodically. The cell-free supernatant obtained after centrifugation at 10,000rpm for 10min, was used as the source of extracellular phytase enzyme. Phytase assay was done by measuring the amount of phosphate released using sodium phytate as a substrate. The reaction mixture comprised 0.5 ml of 0.1 M Mop’s buffer (pH 7.0), 0.5ml of sodium phytate and 1.0ml crude enzyme. The reaction was carried out at 40 ºC for 10min and stopped by adding 2.0ml 10% (w/v) TCA. The phosphate released was determined spectrophotometrically according to Fiske and Subbarow (1925). A reference standard curve of KH2PO4 was simultaneously assayed with the sample. ‘One unit of enzyme activity was defined as the amount of enzyme used to liberate one µmole inorganic phosphate/ml/minute under the assay conditions’.
pH and temperature optima of phytase enzyme
The pH optima of enzyme were determined by incubating reaction mixture on different pH ranging from pH 2.0-9.0 [0.2 M glycine/HCl buffer (pH 2.0 to 3.0), 0.2 M acetic acid/sodium acetate buffer (pH 4.0 to 6.0)], 0.2 M Tris/HCl (pH 7.0 to 8.0)]. The temperature optima of enzyme were determined by incubating the reaction mixture at different temperature 300C-700C and the phytase activity was assayed
Identification of phytase producing bacterial isolate
The phytase producing bacterial isolates were identified by colony morphology and microscopic examination (gram staining). Metabolic fingerprinting is also done for the identification. The culture was sent to BTK biosciences, New Delhi, for the biochemical identification of bacterial species.
Molecular identification
Genomic DNA was extracted by a modified method of Bazzicalupo and Fani (1994). PCR amplification of 16S rDNA of bacterial isolate was carried out using eubacterial universal primers Gm3f and Gm4r. Primer sequences were as follow: Gm3f (5′- AGA GTT TGA TCM TGG – 3′) and Gm4r (5′ – TAC CTT GTT ACG ACT T – 3′)(Eden et al., 1991). PCR amplification was done in 10 µl reaction mixture containing 5 µl taq master mix, 1.25 µl primers, 1.25 µl genomic DNA and 1.25 µl sterile deionised water. The reaction conditions were as follows: initial denaturation at 950C for 7 min, 35 amplification cycles of denaturation at 940C for 1 min, annealing at 510C for 1 min and primer extension at 720C for 1 min; followed by a final extension at 720C for 10 min. The PCR product was run on 0.8% agarose gel and visualized under UV transilluminator. The nucleotide sequence was determined by PCR-directed sequencing by Chromous biotech Pvt. Ltd., Bangalore, India.
Isolation of phytase producing bacteria
Phytase, much in demand, has potential application in food and feed industry for ameliorating digestibility and assimilation of nutrients by improving growth of poultry, pigs and fishes. On primary qualitative screening, 14 bacterial isolates were found to produce clear zone of hydrolysis. The formation of zone of hydrolysis around the bacterial colony was a positive indication of extracellular phytase production (Hill et al. 2009). Fig.1 show zone of clearance around colonies of some isolate.
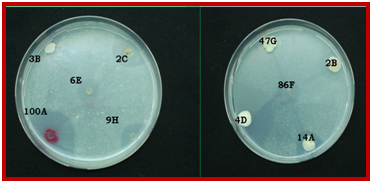
Fig. 1. Zone of hydrolysis shown by some isolates on phytase screening medium containing Ca phytate.
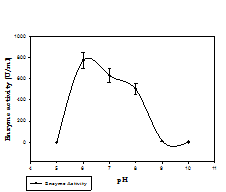
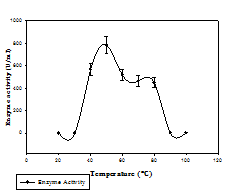
Fig. 2. Phytase activity of bacterial isolate at different pH (a) and temperature (b)
Phytase activity and optimization
Phytase activity was determined by measuring the amount of liberated inorganic phosphate and its reaction with colour reagent. Phytase production was done in PSM broth and five bacterial isolates 100A, 9H, 14A, 4D, and 2B showed significant phytase production in submerged fermentation. The isolate 14A was found to be the best among all with the maximum production of 89.09 U/ml at 72 h (Table.1), and was finally selected for further study. Most bacterial phytases studied show their optimum activity in acidic range. For instance, for Aerobacter aerogenes (Greaves et al.,1967), E. coli (Greiner et al., 1993), S. castellii (Segueilha et al.,1992), A. adeninivorans (Sano et al.,1999), Selenomonas ruminatium (Sreeramuluet al., 1996) and Pseudomonas sp. (Irving and Cosgrove, 1971), the optimum pH was between 4.0 and 5.5. In our study, the maximum enzyme activity (771 U/ml) was observed at pH 6, and comparable activity was observed at pH range (6-8). Activity of enzyme showed a decrement as the pH increases. The maximum activity (784 U/ml) was measured at 50 ºC, and the activity was also considerably good at temperature range 40-60 ºC. Activity decreases as the temperature increases above 50 ºC (Fig.2).Phytase production at elevated temperature is desirable for feed application.
Identification of phytase producing bacterial isolate
Colony morphology and microscopic examination (Gram’s staining) of maximal phytase producers suggested that the isolates 100A, and 14A are gram negative rod, and 9H, 4D, and 2B are gram positive rods and cocci (Fig.3). Using the Biolog system the metabolic pattern of 14A was analyzed. The culture was positive for pH 6, it can utilize D-Glactouronic acid, L-Aspartic acid, D-Glucuronic acid, D-Serine and was found sensitive towards Troleandomycin, Lincomycin, Vancomycin, Rifamycin, Tetrazolium violet, Tetrazolium blue, 1% Sodium lactate, Potassium tellurite and 1% NaCl. The result was recorded which identified it as a species of Pseudomonas florescence. Although, when strain 14A was treated under UV light (Brodsky and Nixon, 1974), it did not show any florescence
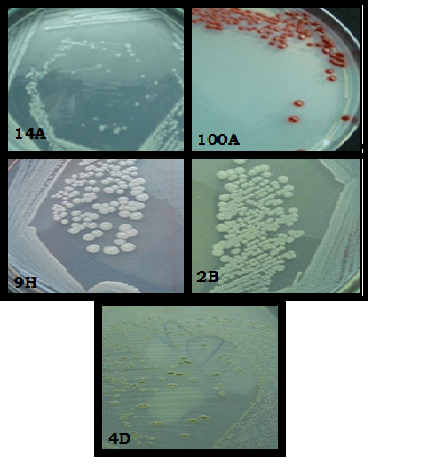
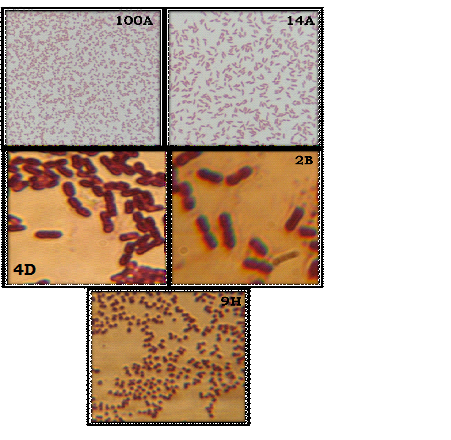
Fig. 3.Colony morphology of phytase producing bacterial isolates (a).Microscopic examination of selected bacterial isolates (100X)(b).
Molecular identification
The molecular identification of bacterial isolate (14A) was done using 16S rDNA (Fig.4). The 16S rDNA gene was amplified by PCR using eubacterial universal primers. The PCR product was estimated to be approximately 1.5 kb. By constructing the phylogenetic tree of the gene sequence the isolate showed maximum similarity with Pseudomonas sp. Phylogenetic relationship is shown in Fig.5. 16S rDNA sequencing and sequence similarity search using BLAST showed that it matches 97 % with Pseudomonas pulluriniana, while 3% was found to be unique sequence (submitted at NCBI Gene bank with the accession number 157554).On the basis of these results, we concluded that the strain 14A belongs to genus Pseudomonas and were named as Pseudomonas sp. strain aazad. Phytase from Pseudomonas spp. were also reported by Richardson and Hadobas2001 and Kim et al. 2002) B. amyloliquefaciens (Kerovuo et al. 1998), Bacillus spp. (Powar and Jagannathan 1982, Shimizu 1992, Kim et al. 1998), Klebsiella spp. (Greiner et al. 1997), E. coli (Greiner et al. 1993) and Mitsuokella spp. (Lan et al. 2002) are some of the phytase producing bacteria reported so far with each producing a positive response to the plate-clearing as well as an enzyme activity. The phytase from the isolated Pseudomonas sp. aazad showed a good enzyme titre and an optimum pH and temperature of 6 and 60ºC respectively that can be used for application as feed supplement for enhancing the utilization of phytate bound phosphorus and thus decreasing pollution caused by it.
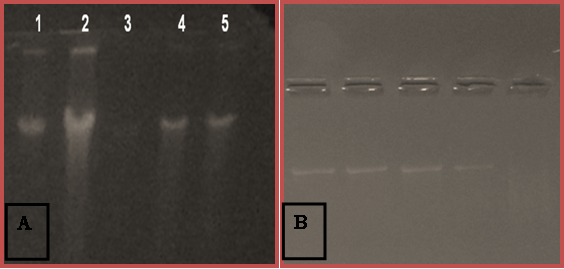
Fig. 4. Gel pictures showing genomic DNA and 16S rDNA amplicon of bacterial isolates.
A. Genomic DNA on agarose gel B. Amplified PCR product of 16S rDNA.
 Fig. 5.Phylogenetic relationship of 16S rDNA sequence of Pseudomonas sp. strain aazad (14A) with available sequences on NCBI nucleotide database.
Fig. 5.Phylogenetic relationship of 16S rDNA sequence of Pseudomonas sp. strain aazad (14A) with available sequences on NCBI nucleotide database. ACKNOWLEDGMENTS
The research is supported by Department of Microbiology, Chaudhary Charan Singh University, Meerut, India. The second author is thankful to Department of Science and Technology (DST), Ministry of Science & Technology, Government of India, New Delhi for financial assistance. All the authors of the research papers and books cited are highly acknowledged.
- Howson, S.J., Davis, R.P. Production of phytate hydrolyzing enzymes by some fungi. Enz. Microb. Technol., 1983; 5: 377-382.
- Urbano, G., Lopez-Jurado, M., Aranda, P., Vidal-Valverde, C., Tenorio, E., Porres, J. The role of phytic acid in legumes: Antinutrient or beneficial function? J Physiol Biochem., 2000; 56: 283–294.
- Lei, X.G., Porres, J.M. Phytase enzymology, application and biotechnology. Biotechnol Lett., 2003; 25: 1787-1794.
- Schroder, B., Breves, G., Rodrhutscord, M. Mechanisms of intestinal phosphorus absorption and bioavailability of dietry phosphorus in pigs. Dtsch Tierarztl Wochenschr.,1996; 103: 209-214.
- Angelis, M.D., Gallo, G., Corbo, M.R., Mc Sweeney, P.L.H., Faccia, M., Giovine, M., Gobbetti, M. Phytase activity in sourdough lactic acid bacteria: purification and characterization of a phytase from Lactobacillus sanfranciscensis CB1.Int. J. Food Microbiol., 2003; (87): 259-270.
- Fiske, C.H., Subbarow, Y. The colorimetric determination of phosphorus. The journl of biological chemistry. , 1925; 6 :375-400.
- Fani, R., P. Lio, I. Chiarelli, M. Bazzicalupo. The evolution of the histidine biosynthetic genes in prokaryotes: a common ancestor for the hisA and hisF genes. J.Mol.Evol., 1994; 38: 489-495.
- Eden, P.A., Schmidst, T.M., Blakemore, R.P., Pace, N.R.Phylogenetic analysis of aquaspirillum magnetotactium using PCR-amplified 16s-rDNA- specific DNA. International journal of systemic bacteriology., 1991; 41: 324-5.
- Hill, B.E., Sutton, A.L., Richert, B.T. Effects of low-phytic acid corn, low-phytic acid soybean meal, and phytase on nutrient digestibility and excretion in growing pigs. J. Anim. Sci., 2009; 87: 1518-1527.
- Greaves, M.P., Anderson, G., Webley, D.M. The hydrolysis of inositol phosphates by Aerobacter aerogenes. Biochim. Biophys. Acta. , 1967; 132: 412-418.
- Greiner, R., Konitzny, U., Jany, K.D. Purification and characterization of two phytases from Escherchia coli. Arch. Biochem. Biophys. , 1993; 303: 107-113.
- Segueilha, L., Lambrechts, C., Boze, H., Moulin, G., Galzy, P. Purification and properties of phytase from Schwanniomyces castellii. J. Ferment. Bioeng., 1992; 74: 7-11.
- Sano, L., Fukuhara, H., Nakamura, Y. Phytase of the yeast Arxula adeninivorans. Biotechnol. Lett., 1999; 21: 33-38.
- Sreeramulu, G., Srinivasa, D.S., Nand, K., Joseph, R. Lactobacillus amylovorus as a phytase producer in submerged culture. Lett Appl Microbiol., 1996; 23: 385-388.
- Irving, G.C.J., Cosgrove, D.J. Inositol phosphate phosphatases of microbiological origin. Some properties of a partially purified bacterial (Pseudomonas sp.) phytase. Aust. J. Biol. Sci., 1971; 24: 547-557.
- Richardson, A.E., Hadobas, P.A., Hayes, J.E. Extracellular secretion of Aspergillus phytase from Arabidopsis roots enables plants to obtain phosphorous from phytate. Plant J., 2001; 25: 641-649.
- Kim, Y.O., Kim, H.K., Bae, K.S., Yu, J.H., Oh, T.K. Purification and properties of thermostable phytase from Bacillus sp. DS11. Enz. Microb. Technol., 1998: 22: 2-7.
- Kerovuo, J., Lauraeus, M., Nurminen, P., Kalkkinen, N., Apajalahti, J. Isolation, characterization, molecular gene cloning and sequencing of a novel phytase from Bacillus subtilis. Appl. Environ. Microbiol., 1998; 64: 2079-2085.
- Powar, V.K., Jagannathan, V. Purification and properties of phytate-specific phosphatase from Bacillus subtilis. J. Bacteriol., 1982; 51: 1102-1108.
- Shimizu, M. Purification and characterization of phytase from Bacillus subtilis (natto) N-77. Biosci. Biotechnol. Biochem., 1992; 56: 1266-1269.
© The Author(s) 2017. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.