


ISSN: 0973-7510
E-ISSN: 2581-690X
Traditionally, diseases have been managed with synthetic chemicals, but concerns over their environmental and health impacts have led to increased interest in biological control methods as sustainable alternatives. This study explores the potential of the seed-borne endophytes for eco-friendly biological control agent against brown spot Bipolaris oryzae (Breda de Haan) and sheath blight disease (Rhizoctonia solani Kuhn) in rice. The findings revealed that B. subtilis RE-1 effectively reduced the mycelial growth of brown spot and sheath blight pathogen up to 46.27% and 40.37%, respectively under in vitro conditions. Endophytes produced extracellular hydrolytic enzymes viz., amylase, protease, cellulase and lipase that degrade fungal mycelia. Additionally, B. subtilis RE-1 synthesized antimicrobial peptides (AMPs) including Iturin (C, D), Bacilysin, Fengycin D, Mycosubtilin, Bacillomycin (A,D) and hydrolytic enzymes viz., β-glucanase, which collectively demonstrated potent antimicrobial and antifungal activities. The identification of six antimicrobial lipopeptides genes and one hydrolytic enzyme gene in seed-borne endophytes is a novel finding of this study, highlighting its comprehensive disease-fighting potential. Furthermore, B. subtilis enhances seed germination rates to 96% and increases the vigor index to 1574.784, compared to the control seeds with 92% germination and a vigor index of 1393.984. The augmentation of seedling growth can be attributed to the release of IAA, siderophore and ammonia utilization. These findings underscore B. subtilis as a promising endophyte that could enhance rice production sustainably.
Endophytes, Bacillus subtilis, Extracellular Hydrolytic Enzymes, Antimicrobial Peptides, Plant Growth Promotion
Rice is the primary agricultural crop for millions of people worldwide. It is a semiaquatic, free-tillering, water-loving, and Angiosperm plant. Globally paddy ranks second behind the wheat crop in area production and productivity.1 India produced 135.76 MT of milled rice in 2023 next to China 145.95 MT. Around 520.4 million metric tons (MMT) of rice were used globally in the 2022/23 season, compared to 437.18 million metric tons in the 2008/2009 season (Statista 2024). The consumption of rice worldwide has experienced a rise in recent years due to the geometrically growing population. The average production and productivity of the crops were comparatively low due to the influence of many external factors. Hence the major external factors are pests and diseases. Diseases are major constraints in limiting grain production. Among them, rice sheath blight incited by Rhizoctonia solani and brown spot incited by Bipolaris oryzae are the most problematic and economically important diseases in all the paddy growing areas. The sheath blight is mostly affected during the tillering stage, with an average incidence of 7.2 to 38.9% causing a loss of 14.3 to 39.7% in Uttar Pradesh.2 Brown spot causes significant grain yield losses (50%-90% ,3 and the mean incidence and intensity of 58.70 and 36.93% in 2018.4 Since 18th century onwards, people have started to use synthetic biocide to control plant disease. The fungicides are potentially used to control the pathogen, but their excess application is detrimental to plant and pollute environment. The concern of managing plant disease using biological control agents (BCAs) as a safe approach to the environment.5
In recent decades, BCAs are potentially used to control plant pathogens. In 1866 de Barry identified the non-pathogenic microorganism residing within the plant tissue named it an endophyte and gave the first definition as “Any organism that lives within plant tissue”.6 Endophytes either bacteria or fungi sustained inside a plant with or without harm to the plant.7,8 Endophytes are identified in various plant species such as tomato, sugarcane, cowpea, groundnut, tea, etc. All plant species identified are associated with endophytes.9 Plants have complex microbes that contribute significantly for growth.10 Endophytes are isolated in various sections of plants viz., leaf, stem, roots, seeds, ovules, germ-free callus tissue, and seed endosphere.11,7 Seeds have a bus vehicle for different microbial communities. The spermatophytes were dormant in seeds for a long period, where they got appropriate conditions, and germinated into new plants. Seeds can benefit from microorganisms that are associated with them during preservation and germination. As the seeds start germinating and absorb water, they release substances to attract bacteria. The bacteria then colonize the root, rhizosphere, and seedling, using nutrients to grow and multiply, ultimately enhancing plant growth and development.12 Seed-borne endophytes vertically transmitted from F1 to F2 generation. Endophytes help to protect the seed from seed and soil-borne pathogens. It influences the physiology and metabolic activity of the host plant. It offers great potential to host plants through various activities viz., antagonistic activity, nutrition competition to pathogens, and Induced systemic resistance. Further, it enhances hydrolytic enzymes to solubilize the phosphate and improve nitrogen fixation and phytohormone production thereby facilitating the growth of plants.13
Bacillus are dominant groups of bacteria that reside inside a plant as an endophyte and in rhizosphere. It has a highly competitive ability and has efficient biocontrol potential against plant pathogens. Bacillus spp. has a rich diverse group of antimicrobial peptides like pyoluteorin, phenazines, bacillomycins, fengycin, lipopeptides, pyrrolnitrin, surfactin, iturin A, iturin C, bacilysin, bafilomycin, mersacidin, ericin, subtilin, subtilisin, and mycosubtilin. The metabolic compound has high antibiotic and antibacterial activities against various plant diseases. Bacillus subtilis var. amyloliquefaciens produces surfactin, iturin, bacillomycin, and azalomycin F which are effective against R. solani,14 Bacillus safensis B21 produces iturin compounds effective against Pyricularia oryzae.15 Antimicrobial peptide genes bacilysin and iturin have substantially decreased the prevalence Fusarium wilt disease in cucumber and Phytophthora blight in pepper.16
Traditional rice varieties have different microbial communities with high immune activities against numerous pathogenic microorganisms. The exploration of endophytes from the plant to control plant pathogens effectively reduces the use of synthetic chemicals, increases production, and productivity, and reduces the use of greenhouse gases.17 This study emphasizes the characterization of endophytes from the traditional rice varieties and explores their antagonistic activity against phytopathogens and their growth-promoting activities.
Isolation of bacterial endophytes
The seed samples of traditional rice varieties were collected from different rice-growing areas viz., Attur Kichili Samba, Karupukavuni, and Thuya Malli from Erode district, Kottara Samba and Arupathankurai were collected from Kanyakumari district. Twenty-five seeds from each variety were selected for endophyte isolation. Seed samples were cleaned for three to four minutes in 2% sodium hypochlorite followed by, rinsing in 70% ethanol for two minutes. The disinfectant was removed by washing three times in sterile water and dried on sterile filter paper. Surface disinfected seed sample was transferred into a sterilized pestle and mortar and crushed using a phosphate buffer solution. The samples were serially diluted and 1 ml of 10-1 and 10-2 dilution was spread onto a Tryptic soya agar medium (TSA) in triplicates and 1 ml of final water wash was poured onto a TSA medium maintained as a control. The dishes were incubated for two to three days at 28 ± 2°C and the colonies were selected colonies with varied morphology (color, size, shape) and purified by single colony method and preserved in glycerol stock at -20°C.18
Isolation of Pathogen
The diseased samples of rice sheath blight were collected from Bhavanisagar, Coimbatore, and Madurai, whereas rice brown spot samples were collected from Coimbatore and Aduthurai. The infected portion was cut into 0.5 to 1 cm in size and disinfected in 70% ethanol followed by washing in sterile water and dried in sterile filter paper. The infected bits were transferred in Potato Dextrose Agar (PDA) and incubated for two to three days. The fungal pathogen was purified by the hyphal tip method and stored at -20°C.19
Pathogenicity test for Rhizoctonia solani and Bipolaris oryzae
The rice seeds of variety ADT 57 were collected from Tamil Nadu Rice Research Institute, Aduthurai, and grown in the glass house till the plant reaches the active tillering stage.20 The white to brown irregular coiled sclerotia of R. solani were placed between the paddy leaf sheath for symptom expression. For the brown spot disease, the plants were already grown glass house. The chaffy grains were autoclaved and inoculated with the mycelial disc of B. oryzae under controlled conditions and kept at room temperature for 15 days for complete sporulation. The spore suspensions were collected by adding sterile water and sprayed by adding two drops of Tween-20 on 30 day plants.19
In vitro screening of endophytes
Biocontrol efficacy of isolated bacterial endophytes was screened in vitro against R. solani, B. oryzae, by Dual culture technique. The isolated endophytic bacterial culture was streaked on the petriplate amended with PDA medium on one side and 5 mm mycelial disc was kept perpendicularly on opposite side and kept at 28 ± 2°C until the control plate reached full growth. The efficient bacterial endophytes were selected based on mycelial growth inhibition. The percent inhibition zone was calculated.21
The percent inhibition zone was measured by using
Percent inhibition = C – T/ C x 100
Where C is the test pathogen growth (mm) in the control plate and T is the growth of test pathogen with endophyte (mm).
Molecular characterization of fungal pathogens
Molecular conformation of the fungal pathogen by using ITS-1 (5’-TCCGTAGGTGAACCTGCGG-3’) and ITS 4 (5’-TCCTCCGCTTA TTGATATGC-3’) with following condition viz., 95°C for 2 min, followed by 35 cycles of 94°C for 30 sec, 56°C for 1 min, 72°C for 2 min and final extension 72°C for 8 min was conducted in Bio radar PCR. The PCR reaction contains 5 µl master mix, 1 µl forward primer, 1 µl reverse primer, 1 µl DNA and 2 µl water. The amplified product was resolved in 1.2% (w/v) agarose and visualized under a UV detector and documented. The amplified PCR fragments were purified, sequenced and compared in the NCBI database and sequences were submitted in NCBI database.
Characterization of endophytes
Gram staining was used to examine the morphological characteristics of endophytes and biochemical tests like urease, citrate utilization, gelatinase, indole methyl red, Voges Proskauer and glucose utilization tests were made in 24 to 48 hrs old bacterial culture.13
Molecular characterization of bacterial endophytes
Molecular characterization of bacterial endophyte isolates was done by extraction of DNA and amplification of 16S rDNA region using universal primers 27F (5-GAGTTTGATCCTGGCTCA-3) and 1492 (R-TACGGYTACCTTGTTACGACTT) under following condition viz., 94°C for 4 min. followed by 35 cycles of 94°C for 1 min, 55°C for 1 min and 72°C for 2 min and final extension 72°C for 10 min in a Bio rader thermocycler. The PCR product was separated in 1.2% (w/v) agarose gel, visualized under UV detector and documented. The amplified PCR fragment was purified, sequenced and compared in the NCBI database and sequences were submitted in NCBI database.
Assessment of the characteristics of Bacillus subtilis for plant growth under in vitro
Traits that promote plant growth both directly and indirectly were investigated for the effective endophyte.
Amylase activity
Endophyte isolate RE-1 was streaked in a nutrient medium amended with 1% soluble starch with adjusted pH 6 and incubated at 28°C for 3 to 4 days. When iodine solution was added to the plates, the starch hydrolysis test came back positive, indicating the presence of a distinct halo zone surrounding the bacterial colony.22
Protease Production
Endophyte isolate RE-1 was streaked on skim milk agar (1%) and kept at 30°C for 5-7 days. The clear halo region surrounding cultured spot was considered to be indicative of protease production.12
Cellulase Production
By spreading on carboxy methyl cellulose agar medium, the cellulase activity of bacterial isolate RE-1 was assessed by incubation at 30°C for 4-5 days. Then it was submerged in a 1% w/v Congo red solution for 20 minutes.12 After removing any excess Congo red solution, the plate was filled with iodine solution and left for a minute. The appearance of a transparent area surrounding the bacteria demonstrated cellulose decomposition.
Lipase activity
The 24 hrs old bacteria were streaked in a Tween 20 agar plate media containing 10 g of peptone, 0.1 g of CaCl2.2H20, 5 g of NaCl, agar 20 g, 10 ml of Tween 20 in 1 litre distilled water and incubated for 5 days. Observance of milky precipitation around the bacterial colony indicated positive results.23
Amplification of antimicrobial peptide genes using PCR
From endophytic bacteria isolate RE-1, the genomic DNA was isolated. The PCR reaction was carried out 10 µl mixture containing 5 µl master mix, 1 µl of each forward and reverse primer, 1 µl DNA, and 2 µl water. The specific primers were used to detect antimicrobial lipopeptide genes from the endophyte isolate RE-1 (Table 1). The amplified products were run in 1.2% agarose and documented.
Table (1):
Gene-specific primers of lipopeptide genes
Antibiotic Primer |
Gene |
Primer Name |
Sequence (5’ -3’) |
Ref. |
|---|---|---|---|---|
ITURIN D |
ItuD |
ITUP4-F ITUP5-R |
CCCCTGTTCTAGATGATCGGA GGAATCTCTGCATCGATTCTGTCCATCTAACCGGCATC |
24 |
BACILLOMYCIN D |
BamD |
ITUD-F1 ITUD-R1 |
TTGAAYGTCAGYGCSCCTTT TGCGMAAATAATGGSGTCGT |
16 |
BACILYSIN |
BacD |
BACAB-F1 BACAB-R1 |
CTTCTCCAAGGGGTGAACAG TGTAGGTTTCACCGGCTTTC |
16 |
ITURIN C |
ItuC |
ITUC-F ITUC-R |
TTCACTTTTGATCTGGCGAT CGTCCGGTACATTTTCAC |
25 |
BACILLOMYCIN A |
Bmy A |
BACA-F BACA-R |
TGAAACAAAGGCATATGCTC AAAAAATGCATCTGCCGTTCC |
25 |
FENGYCIN D |
FenD |
FEN D-F FEN D-R |
CCTGCAGAAGGAGGAGAA GAAGTGCTCATCGTCTTCCGTTTC |
25 |
MYCOSUBTILIN |
FenF |
ITUD-F1 ITUD-R1 |
TTGAAYGTCAGYGCSCCTTT TGCGMAAATAATGGSGTCGT |
16 |
β-GLUCANASE |
Encoding PR2 protein |
b- GLU- F b-GLU- R |
AATGGCGGTGTATTCCTTGACC GCGCGTAGTCACAGTCAAAGTT |
25 |
Plant Growth Promoting Activity of Endophyte
Ammonia utilization
The utilization ability of ammonia was performed by inoculation of 24 hour old culture in 10 ml of peptone broth and incubated at 120 rpm for 96 hrs at 30°C.26 Nessler’s reagent was added and allowed for 5 min observed the colour changed from yellow to brown colour as positive result.
IAA production test
The IAA production was examined qualitatively by using Salowski’s reagent. The bacteria was spot inoculated in NA medium amended with L-Tryptophan (1 mg/ml) and cultured at 37°C in orbital shaker for 72 hours. The culture was centrifuged at 10000 rpm for 20 min and 2 ml of Salowski’s reagent was added to 1 ml of supernatant. The appearance of a pink color indicated positive result.12
Siderophore production activity
Twenty-four hours old endophyte was streaked in a chromo azurol S (CAS) agar medium and kept at at 37°C for two to four days for chelating iron. Observation of orange to yellow indicates that the bacteria can chelate the iron as a positive result.12
Assessment of growth promoting activities of endophyte in rice by paper towel method
Bacteria’s ability to enhance growth was evaluated using paper towel method. The endophytic isolate RE-1 was cultured in nutrient agar broth and maintained at 27°C for 24 hours. The cell suspension was centrifuged at 10000 rpm for 20 min and bacterial cells were dissolved in phosphate buffer containing 0.5% carboxymethyl cellulose. The rice seeds of variety ADT 57 disinfected in 5% NaClO solution and rinsed in sterile water. Disinfected seeds were soaked in endophytic culture for 2 hrs in a shaker and seeds soaked in water served as control. Endophytes treated seeds and control seeds were arranged in a germination paper at 25 seeds per sheet. On the seventh day, the germination percentage, root length, and shoot length of the seedlings were recorded in endophytes treated seeds and control seed and the seedling vigor index was calculated27
VI = X × Y
Where, VI= Vigour index, X= Mean root length+ mean shoot length, Y= Germination percentage %.
Statistical analysis
All the in vitro experiments were carried out in Completely Randomized Design (CRD) and analyzed independently. The treatment means were compared and data’s were analyzed through Duncan’s Multiple Range-Test (DMRT) test in SPSS version 2022 software.
Isolation, pathogenicity and molecular confirmation of pathogens
The pathogen was isolated on a PDA medium based on symptom-identified on leaf tissue i.e., oval or ellipsoid lesion just above the soil with greenish to grey. For sheath blight disease, the actively coiling white color mycelial sclerotia were placed between the leaf sheath of the actively growing paddy variety (ADT 57). The symptom of sheath blight was initially expressed as a grey-colored oval-shaped lesion on the 3rd day, followed by a brown elongated necrotic spot vertically on the leaf sheath. For brown spot disease, expression of a sesame-shaped spot with a dark brown margin with yellow hallow around the necrotic spot on the 4th day. The pathogens were reisolated from the infected portion and confirmed pathogenicity based on morphology and pathogenicity was proved.
Molecular characterization of fungal pathogens
The pathogens viz., B. oryzae and R. solani were molecularly confirmed by using ITS1 and ITS4 primers for fungi. The amplified product of 163 bp of B. oryzae was sequenced and blasted in the NCBI database, and submitted in NCBI database with accession number B. oryzae (PP785999). Comparison of the sequence in NCBI database showed 96.84% identity with OP425396 (B. oryzae) followed by 96.20% identity with MT874332 (B. oryzae) and 95.57% identity with PQ475956 (B. oryzae). The amplified product of 669 bp of R. solani was sequenced and blasted in the NCBI database, and submitted in NCBI database with accession number R. solani (PP785992). Comparison of the sequence in NCBI database showed 98.80% identity with R. solani accessions viz., ON974838 , ON974836 and ON974740.
Isolation and in vitro screening of endophytes
From seed samples, twenty endophytes were isolated named RE-1 to RE-20 and further the endophytes were screened against R. solani, and B. oryzae under controlled conditions. Among the twelve endophytes screened, the isolate RE- 1 showed maximum inhibition of 40.37% resulting in the lowest mycelial growth of 53.67 mm (Figure 1), followed by R.E-8 (37.41%) and R.E-11 (37.41%). The mycelial growth of the control plate was 90 mm (Table 2). Eleven endophytes were screened against B. oryzae under in vitro and among them, the isolate RE-1 showed the highest inhibition percent (46.27%) with 46 mm radial mycelial growth (Figure 2) followed by R.E-4 39.61 percent inhibition and R.E-6 with 38.82 percent inhibition of mycelial growth under in vitro. The mycelial growth of the control plate was 85 mm (Table 3).
Table (2):
In vitro screening of bacterial endophytes against R. solani
Endophytes |
Mycelial Growth (mm) |
Percent inhibition (%) |
Inhibition Zone (mm) |
|---|---|---|---|
R.E – 1 |
53.67a |
40.37a |
13.00a |
R.E – 20 |
56.33b |
37.41b |
0.00f |
R.E – 11 |
56.33b |
37.41b |
9.33b |
R.E – 16 |
58.00c |
35.56c |
7.67c |
R.E – 18 |
59.67d |
33.70d |
4.33e |
R.E – 8 |
60.00d |
33.33d |
0.00f |
R.E – 10 |
62.67e |
30.37e |
4.00e |
R.E – 7 |
63.00e |
30.00e |
5.67d |
R.E – 17 |
64.67f |
28.15f |
0.00f |
R.E – 15 |
68.00g |
24.44g |
0.00f |
R.E – 12 |
68.67g |
23.70h |
0.00f |
R.E – 9 |
70.00h |
22.22h |
0.00f |
Control |
90.00i |
0.00i |
0.00f |
SD error |
0.068 |
0.075 |
0.095 |
SCD |
0.19 |
0.22 |
0.20 |
CD (0.05%) |
0.32414 |
0.37532 |
0.44 |
CV % |
0.66 |
1.6 |
6.9 |
Table (3):
In vitro screening of bacterial endophytes against B. oryzae
Endophytes |
Mycelial Growth (mm) |
Percent inhibition (%) |
Inhibition Zone (mm) |
|---|---|---|---|
R.E-1 |
45.67a |
46.27a |
16.67a |
R.E-4 |
51.33b |
39.61b |
5.00c |
R.E- 6 |
52.00b |
38.82b |
6.33b |
R.E-11 |
53.00bc |
37.65bc |
1.00g |
R.E-15 |
53.33bc |
37.25bc |
6.67b |
R.E-17 |
53.33bc |
37.25bc |
4.67cd |
R.E -12 |
53.33bc |
37.25bc |
4.00e |
R.E-19 |
56.00cd |
34.12cd |
4.33cd |
R.E-18 |
57.67d |
32.16d |
3.00f |
R.E- 10 |
60.00e |
29.41e |
0.00h |
R.E-13 |
64.33f |
24.31f |
7.00b |
control |
85.00g |
0.00g |
0.00h |
SD error |
0.252 |
0.296 |
0.065 |
SCD |
0.711493 |
0.837058 |
0.179505 |
CD |
1.21736 |
1.196748 |
0.307133 |
CV% |
2.651764 |
5.374463 |
7.788682 |
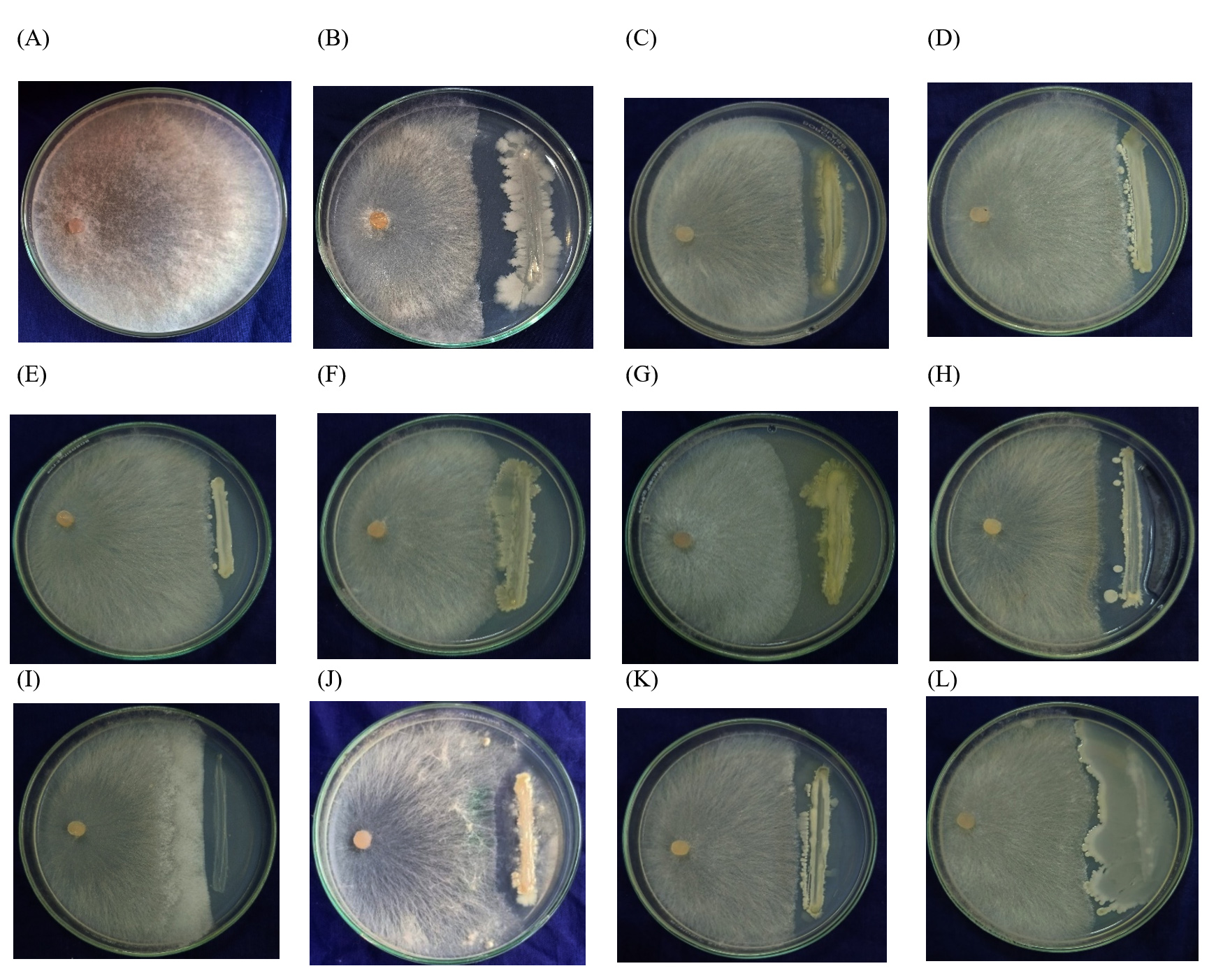
Figure 1. Antagonistic effect isolated endophytes against Rice Sheath blight (R. solani) A – Control, B – RE-1, C – RE-7, D – RE-9, E – RE-12, F – RE-12, G – RE-8, H – RE-11, I – RE-17, J – RE-18, K – RE-16, L – RE-20
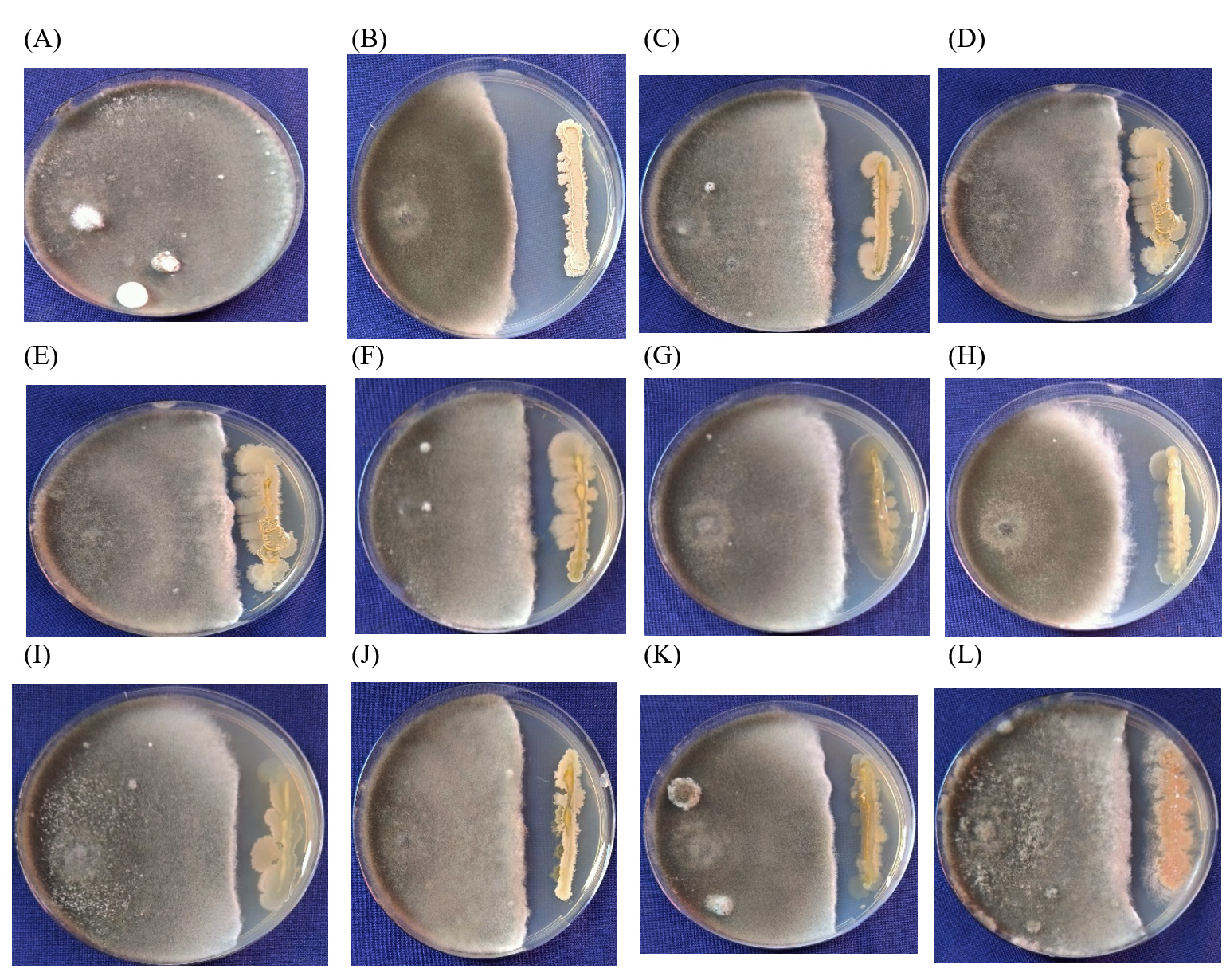
Figure 2. Antagonistic effect isolated endophytes against Rice brown spot (B. oryzae) A – Control, B – RE-1, C – RE-15, D – RE-10, E – RE-12, F – RE-13, G – RE-11, H – RE-17, I – RE-13, J – RE-14, K – RE-6, L – RE-18
Characterization of bacterial endophytes
In vitro research findings demonstrated that isolate RE-1 performed noticeably better than the other isolates. Hence the isolate RE-1 was studied in detail for all morphological, biochemical, and molecular characterization. In the gram-staining experiment, the bacteria colony showed a smooth and creamy white color, indicating a gram-positive reaction. The biochemical test showed a positive result for citrate utilization, urease test, methyl red test, gelatinase test, and glucose utilization test, and negative result on the indole test, VP test, and hydrogen sulfide production test.
Isolate R.E1 showed positive results for protease, amylase, and cellulase test with a clear holo zone around the bacterial colony. Lipase activity also exhibited positive results with white color precipitation around the bacterial colony (Figure 3).
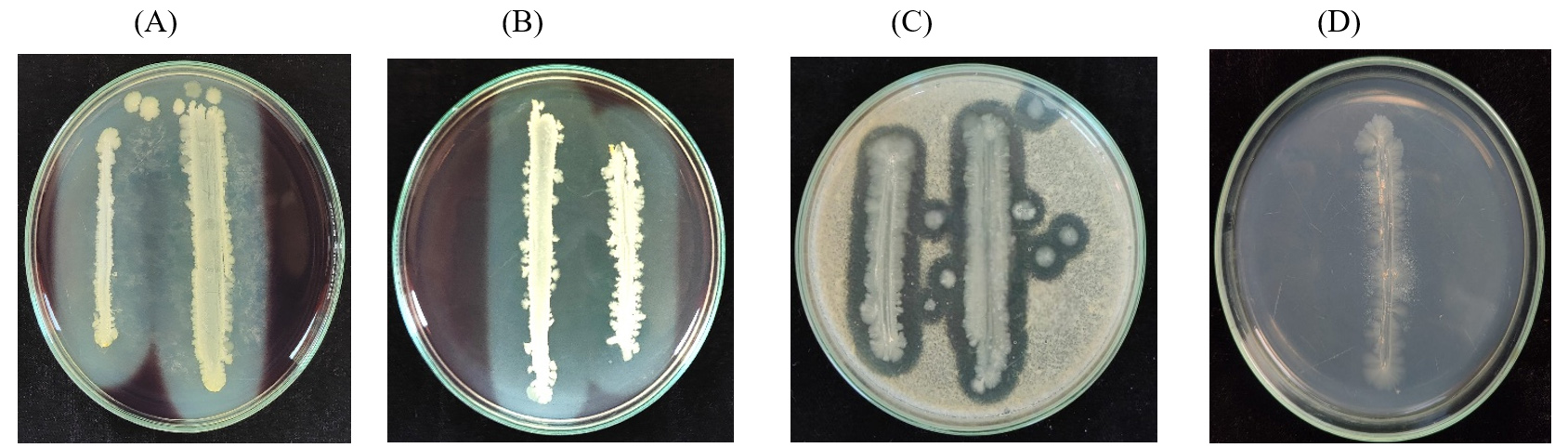
Figure 3. Extracellular hydrolytic enzyme of RE-1 isolate. A) Cellulase; B) Amylase; C) Protease; D) Lipase
Based on gram staining and biochemical tests revealed that the isolate further goes for R.E-1 molecular characterization. RE-1 was molecularly confirmed using primer 16S rRNA using universal primer 27F and 1492R primers. The 904 bp amplified product was sequenced and submitted in the gene bank with Accession number (PQ056582). Comparison of the sequence in NCBI database showed 95.44% identity with Bacillus subtilis strain PF216S ribosomal RNA gene (PP408666) followed by 95.10% identity with Bacillus amyloliquefaciens strain WNB1 16S ribosomal RNA gene (OR447622.1) and 95.11% identity with Bacillus sp. M63 (2010) strain M63 (GQ340501) The results revealed that the isolate matched with Bacillus subtilis. The Morphological, Biochemical, and Molecular characterization results reveal that the isolate RE-1 as B. subtilis
Qualitative analysis of extracellular hydrolytic activity of B. subtilis
The B. subtilis isolate RE-1 was verified for its indirect growth promotion and defense activity. The protease, amylase and cellulase test shows a clear halo zone around the bacterial colony positively. Lipase activity also exhibited positive results with white color precipitation around the bacterial colony.
Characterization of secondary antibiotic metabolites by B. subtilis isolate RE-1
The B. subtilis isolate RE-1 was further evaluated using different antibiotic biosynthetic genes (lipopeptides). Gene-specific primers for Iturin D, Iturin C, Bacillomycin D, Bacilysin, Bacillomycin A, Fengycin D, Mycosubtilin, and β-glucanase were used to amplify the lipopeptide genes by PCR in the Table 1. The six antibiotic lipopeptides were amplified with desired amplicon sizes viz., Iturin D (800 bp), Bacillomycin D (482 bp), Bacilysin (815 bp), Bacillomycin A (344 bp), Fengycin D (220 bp), Mycosubtilin (582 bp) except Iturin C (506 bp) and one hydrolytic enzyme β-Glucanase (400 bp) (Figure 4) were amplifed. These results expressed that the seed core endophytes B. subtilis isolate RE-1 sustain inside the seed material and protect the seeds and seedling from plant pathogens viz., seed-borne (B. oryzae), soil-borne (R. solani). The lipopeptides genes group has high antibacterial and antifungal activity.

Figure 4. Documentation of Antimicrobial peptide gene of B. subtilis RE-1 isolate, (A). Bacillomycin D, Bacilysin, and Mycosubtilin (B). Itu D (C). Bacillomycin A (D). β-Glucanase (E). Fengycin D
Growth enhancement activity of endophyte
The isolate RE-1 showed a positive reaction for IAA production, Siderophore production and Ammonia utilization test. The bacteria produced a dark pink color after adding the Salowski’s reagent. B. subtilis creates a yellow hollow zone surrounding the colony in the CAS agar plate, indicating positive result. Addition of Nessler’s reagent changed from yellow to brown indicated a positive result for ammonia utilization.
Growth promoting activity of B. subtilis isolate RE-1 using roll towel paper method
The plant growth-promoting activity of the B. subtilis isolate RE-1 isolate was evaluated using the roll towel paper method. Twenty five seeds were primed with B. subtilis isolate RE-1 and untreated control as a check. Out of 25 seeds, 24 were germinated in the B. subtilis RE-1 treated seeds, which shows 96% germination percentage with significant increases in root length of 11.9 cm, shoot length of 4.5 cm, and seedling vigor of 1574.40 whereas only 23 seeds germinated in the untreated control that exhibited a 92% germination percentage with root growth of 11.33 cm, shoot growth of 3.82 cm, and seedling vigor of 1393.80 when compared with control. Compared to the control, the B. subtilis RE- 1 treated seeds showed significant increases in the secondary root growth
The seeds of Attur Kichili Samba, Karupukavuni, Thuya Malli Kottara Samba, and Arupathankurai traditional rice varieties were chosen for isolating rice endophytes, the inner spermosphere has crucial taxa ability to support important microorganisms that aid in germination and have the potential for promoting plant growth.28,29 In this research, rice seeds had increased total microbial numbers in comparison.30,31 Rice endophytes were screened for their biocontrol potential under in vitro condition before molecular identification yielded relatively promising endophytes.
Bacillus spp are the dominant genera in the soil and living plants.8,12-14 The bacterial endophytes effectively curb the plant’s pathogenic activity. The biocontrol agents potentially reduce disease incidence through various mechanisms involving antibiosis, parasitism, competition, quorum sensing, induced resistance, and plant growth promotion activities.13,32 B. subtilis showed potential biocontrol against brown spot, bacterial blight, sheath blight, and blast disease in rice.13,14,33-35 The in vitro dual plate assay revealed that isolate R.E-1 exhibited the highest percent inhibition for both R. solani and B. oryzae.
Based on biochemical analysis and 16s rRNA gene sequencing, the bacterial isolate R.E-1 was identified as B. subtilis.12,13,36,37 The biochemical test is used to distinguish between groups and categories of bacteria and their potential activity in the plant health system. The positive result of the methyl red test indicates that bacteria produce large amounts of acids by fermenting glucose, resulting in the end product of acetoin as a neutral product. Some bacteria produce high levels of acids towards the end of the fermentation process to maintain the buffering system.38 Bacteria release protease enzymes outside the cell to break down gelatin into amino acids for their metabolism. B. subtilis ferments glucose, converting the end product of glycolysis into gaseous by-products like formic acid, which produces hydrogen and carbon dioxide gas as end products. The microorganism helps to increase soil fertility by either degradation or hydrolysis of the synthetic urea.39 The urease test determines the bacteria’s capacity to convert urea into a different form of nitrogen that plants can easily absorb. The bacteria also exhibited unfavorable outcomes in the citrate and VP test. Molecular identification of bacterial isolate R.E-1 with 16S rRNA gene amplified 3000 base pairs which is similar to B. subtilis.
The bacterial extracellular hydrolytic enzymes viz., cellulase, amylase, protease, and lipase limit the activity of fungal pathogens and increased the growth. B. subtilis used cellulose which is a major component of cell walls of fungi and cell wall lysis results in hyphal leakage.
In fungi, Pythium aphanidermatum and Rhizoctonia solani have been reported high inhibition due to cellulose.40,41 The extracellular enzyme amylase produced by B. subtilis inhibits plant pathogens viz., Alternaria, Bipolaris and hydrolyzing the starch into simple sugars like glucose, which improves plant growth promotion.42 B. subtilis produces extracellular protease that hydrolysis the protein peptide bond in pathogens and absorbs the hydrolysis cells reported in Ralstonia solanacaerum which also involved in the breakdown of biofilm formation and enhances disease resistance.43 The Bacillus subtilis isolate R.E-1 has potentially produced the hydrolytic enzymes viz., cellulase, amylase, protease, and lipase activity, which are important lysis activities for effectivity controlling the pathogen.
The AMP genes have a high frequency of antifungal and antibacterial activity against plant pathogenic microorganisms. Bacillus spp. are the predominant gene groups of antimicrobial peptides like lipopeptides, pyrrolnitrin, surfactin, iturin A, iturin C, bacilysin, bafilomycin, pyoluteorin, phenazines, bacillomycins, fengycin, subtilin, subtilisin, and mycosubtilin. Bacilius has the highest antibiotic activity.44 Iturin, Bacillomycin D, Surfactin, iturin, and Bacillomycin have high antifungal activity against R. solani,13,44 C15 surfactin produced by Bacillus spp has antibacterial activity against Xanthomonas oryzae pv oryzae.33 The AMPs genes such as iturin, fengycin and surfactin produced by B. subtilis has antifungal activity against B. cinerea causing grey mold disease in apples.45 Surfactin, bacillaene, difficidin, macrolactins and fengycin produced by Bacillus spp are having antifungal action action against Saccharicola, Cochliobolus, Alternaria and Fusarium. 46
To detect the presence of lipopeptide genes based on PCR-based detection.47 The following eight antibiotic genes were amplified viz., fengycin D, α-glucanase, mycosubtilin, bacillomycin D, bacilysin, bacillomycin A, Iturin C and Iturin D by the specific primers which showed that the amplification of seven AMP genes for the first time in seed borne endophyte B. subtilis which has significant influences in controlling plant pathogens. As concluded seed-borne endophytes have rich hydrolytic and antimicrobial lipopeptide activity against plant diseases in rice. The extracellular enzymatic genes (β-glucanase) have not only a crucial role in antibiotic activity but also increase growth promotion activity.
The endophytic B. subtilis has potentially increased plant growth and development.8,12 The effective isolate shows significant production of IAA, Siderophore, and Ammonia utilization. The phytohormone activity of bacteria increases the growth activity and competition to nutrition to plant pathogens. IAA production activity helps in increasing cell differentiation. Siderophore production of B. subtilis uses bacillibactin to chelate the iron in the surrounding environment and make competition to harmful microorganisms and increasing iron (Fe³) nutrient availability to the plant.48,35 Ammonia-utilized bacteria help convert ammonia into available form nitrogen, which effectively boosts up to increasing chlorophyll content. Based on the phytohormone production activity of bacteria, the roll towel method of seed germination activity experiment were conducted.27 There was significantly increasing root growth, shoot growth, and germination percentage. The biopriming rice seeds with B. subtilis show 96% of germination percentage and seedling vigor.
B. subtilis has potential multifunctional activity viz., Disease management, industrial application, and plant nutrition improvement which concluded that host manipulation using beneficial microorganisms helps to mitigate plant stress.
In summary, the exploration of rice endophytes, particularly Bacillus subtilis RE-1, highlights their significant role in combating sheath blight caused by R. oryzae and brown spot caused by B. oryzae. This research demonstrates that B. subtilis RE-1 not only exhibit strong antifungal activity but also synthesized antimicrobial peptides genes (AMPs) including Iturin (C, D), Bacilysin, Fengycin D, Mycosubtilin, Bacillomycin (A,D) and hydrolytic enzymes viz., α-glucanase, which collectively demonstrated potent antimicrobial and antifungal activities. It enhances plant growth through various mechanisms, including nutrient uptake, production of growth-promoting hormones like indole acetic acid, and secretion of siderophores. The diverse biochemical activities underscore their potential and sustainable role in crop protection. As the agricultural landscape shifts towards more eco-friendly practices, harnessing the capabilities of endophytic bacteria may provide an effective strategy for managing plant diseases and promoting healthy crop growth. Further studies are needed to optimize their application and fully understand their interactions within the plant microbiome.
ACKNOWLEDGMENTS
The authors are thankful to the Tamil Nadu Agricultural University, Coimbatore, India, for funding support to the Research Scholar as Scholarship from Dean (SPGS).
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Chaudhari PR, Tamrakar N, Singh L, Tandon A, Sharma D. Rice nutritional and medicinal properties: A review article. Journal of Pharmacognosy and Phytochemistry . 2018; 7(2): 150-156. (https://www.phytojournal.com/archives/2018/vol7issue2/PartC/7-1-256-103.pdf)
- Ahamad F, Khan MR. Incidence of Sheath Blight in Irrigated Rice and Associated Yield Losses in Northern India. Plant Dis. 2023;107(10):2907-2915.
Crossref - Monisha S, Praveen N, Ramanathan A. Isolation, characterization and management of brown spot disease of rice. 2019;8(3):4539-4545. (https://www.phytojournal.com/archives/2019/vol8issue3/PartBP/8-3-519-227.pdf)
- Abrol S, Singh SK, Singh VB, Singh AK, Singh R, Change C. Morpho-Cultural Variability Characterization of Bipolaris oryzae Causing Brown Leaf Spot of Rice in Jammu Sub-Tropics. Int J Environ Clim Change. 2022;12(11):3378-3384.
Crossref - Nicolaisen MH, Cuong ND, Herschend J, et al. Biological control of rice sheath blight using hyphae-associated bacteria: development of an in planta screening assay to predict biological control agent performance under field conditions. Bio Control. 2018;63:843-53.
Crossref - Bary A. Morphologie und physiologie der pilze, flechten und myxomyceten: W. Engelmann; 1866.
Crossref - Hardoim PR, Hardoim CC, van Overbeek LS, van Elsas JD. Dynamics of seed-borne rice endophytes on early plant growth stages. Plos One. 2012;7(2):e30438.
Crossref - Shahzad R, Waqas M, Khan AL, et al. Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol Biochem. 2016;106:236-243.
Crossref - Smith SA, Tank DC, Boulanger L-A, et al. Bioactive endophytes warrant intensified exploration and conservation. PLoS One. 2008;3(8):e3052.
Crossref - Okubo T, Ikeda S, Sasaki K, et al. Phylogeny and functions of bacterial communities associated with field-grown rice shoots. Microbes Environ. 2014;29(3):329-332.
Crossref - Wang W, Zhai Y, Cao L, Tan H, Zhang R. Endophytic bacterial and fungal microbiota in sprouts, roots and stems of rice (Oryza sativa L.). Microbiol Res. 2016;188-189:1-8.
Crossref - Jamali H, Sharma A, Roohi, Srivastava AK. Biocontrol potential of Bacillus subtilis RH5 against sheath blight of rice caused by Rhizoctonia solani. J Basic Microbiol. 2020;60(3):268-80.
Crossref - Kumar V, Jain L, Jain SK, Chaturvedi S, Kaushal P. Bacterial endophytes of rice (Oryza sativa L.) and their potential for plant growth promotion and antagonistic activities. S Afr J Bot. 2020;134:50-63.
Crossref - Nagendran K, Karthikeyan G, Mohammed Faisal P, et al. Exploiting endophytic bacteria for the management of sheath blight disease in rice. Biological Agriculture & Horticulture. 2014;30(1):8-23.
Crossref - Rong S, Xu H, Li L, Chen R, Gao X, Xu Z. Antifungal activity of endophytic Bacillus safensis B21 and its potential application as a biopesticide to control rice blast. Pestic Biochem Physiol. 2020;162:69-77.
Crossref - Chung S, Kong H, Buyer JS, et al. Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soilborne pathogens of cucumber and pepper. Appl Microbiol Biotechnol. 2008;80(1):115-23.
Crossref - Turner TR, James EK, Poole PS. The plant microbiome. Genome Biol. 2013;14(6):209.
Crossref - Elbeltagy A, Nishioka K, Suzuki H, et al. Isolation and characterization of endophytic bacteria from wild and traditionally cultivated rice varieties. Soil Sci Plant Nutr. 2000;46(3):617-629.
Crossref - Nazari S, Javan-Nikkhah M, Fotouhifar K-B, Khosravi V, Alizadeh A. Bipolaris species associated with rice plant: pathogenicity and genetic diversity of Bipolaris oryzae using rep-PCR in Mazandaran province of Iran. J Crop Prot. 2015;4(4):497-508.
- Durgadevi D, Harish S, Manikandan R, Prabhukarthikeyan SR, Alice D, Raguchander T. Proteomic profiling of defense/resistant genes induced during the tripartite interaction of Oryza sativa, Rhizoctonia solani AG1-1A, and Bacillus subtilis against rice sheath blight. Physiol Mol Plant Pathol. 2021;115:101669.
Crossref - Dennis C, Webster J. Antagonistic properties of species-groups of Trichoderma. II. Production of volatile antibiotics. Trans Br Mycol Soc. 1971;57(1): 41-48
Crossref - Alexander D, Zuberer DJB, soils Fo. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biology and Fertility of Soils. 1991;12:39-45.
Crossref - Hankin L, Anagnostakis SL. The use of solid media for detection of enzyme production by fungi. Mycologia. 1975;67(3):597-607.
Crossref - Tsuge K, Akiyama T, Shoda M. Cloning, sequencing, and characterization of the iturin A operon. J Bacteriol. 2001;183(21):6265-6273.
Crossref - Baysal O, Calykan M, Yesilova O, Pathology M. An inhibitory effect of a new Bacillus subtilis strain (EU07) against Fusarium oxysporum f. sp. radicis-lycopersici. Physiol Mol Plant Pathol. 2008;73(1-3):25-32.
Crossref - Mbai FN, Magiri EN, Matiru VN, Nganga JN, Nyambati VCS. Isolation and characterization of bacterial root endophytes with potential to enhance plant growth from Kenyan Basmati rice. Am Int J Contemp Res. 2013;3(4):25-40. https://repository.dkut.ac.ke:8080/xmlui/handle/123456789/7779
- Siva M, Sreeja S, Thara SS, Heera G, Anith KN. Screening and evaluation of bacterial endophytes of cowpea [Vigna unguiculata (L.) Walp.] for plant growth promotion and biocontrol potential. Plant Science Today. 2024;11(2).
Crossref - Shahzad R, Khan AL, Bilal S, Asaf S, Lee I-J. What is there in seeds? Vertically transmitted endophytic resources for sustainable improvement in plant growth. Front Plant Sci. 2018;9:24.
Crossref - Verma SK, Kingsley K, Irizarry I, Bergen M, Kharwar RN, White Jr JF. Seed vectored endophytic bacteria modulate development of rice seedlings. J Appl Microbiol. 2017;122(6):1680-1691.
Crossref - Hameed A, Yeh M-W, Hsieh Y-T, Chung WC, Lo CT, Young LS. Diversity and functional characterization of bacterial endophytes dwelling in various rice (Oryza sativa L.) tissues, and their seed-borne dissemination into rhizosphere under gnotobiotic P-stress. Plant and Soil. 2015;394:177-97.
Crossref - Loaces I, Ferrando L, Fernandez Scavino A. Dynamics, diversity and function of endophytic siderophore-producing bacteria in rice. Microb Ecol. 2011;61:606-618.
Crossref - Danish M, Shahid M, Altaf M, Tyagi A, Ali S. Pathology MP. Plant growth-promoting rhizobacteria and biocontrol agents triggered plant defence responses against phytopathogenic fungi and improved rice growth. Physiol Mol Plant Pathol. 2024;133:102337.
Crossref - Jin L, Yang L, Li W, et al. Diversity and biocontrol potential of culturable endophytic fungi in cotton. Front Microbiol. 2021;12:698930.
Crossref - Sirivella N, Gopalakrishnan C, Kannan R, et al. Analysis of Bioactive Secondary Metabolites Produced by Endophytic Bacillus amyloliquefaciens against Rice Sheath Blight Pathogen Rhizoctonia solani. Agricultural Science Digest – A Research Journal.2024.
- Tu C-K, Huang W-D, Wang P-H, et al. The rice endophytic bacterium Bacillus velezensis LS123N provides protection against multiple pathogens and enhances rice resistance to wind with increase in yield. Biological Control. 2024;192:105507.
Crossref - Karthikeyan G, Rajendran L, Sendhilvel V, Prabakar K, Raguchander T. Diversity and functions of secondary metabolites secreted by epi-endophytic microbes and their interaction with phytopathogens. Biocontrol Agents and Secondary Metabolites. 2021:495-517.
Crossref - Vinayarani G, Prakash HS, Biotechnology. Fungal endophytes of turmeric (Curcuma longa L.) and their biocontrol potential against pathogens Pythium aphanidermatum and Rhizoctonia solani. World J Microbiol Biotechnol. 2018;34(3):1-17.
Crossref - Beuvink JMW, Spoelstra SF. Interactions between substrate, fermentation end-products, buffering systems and gas production upon fermentation of different carbohydrates by mixed rumen microorganisms in vitro. Environmental Biotechnology. 1992;37:505-509.
Crossref - Hasan HAH. Ureolytic microorganisms and soil fertility: A review. Commun Soil Sci Plant Anal. 2000;31(15-16):2565-89.
Crossref - Sindhu SS, Dadarwal KR. Chitinolytic and cellulolytic Pseudomonas sp. antagonistic to fungal pathogens enhances nodulation by Mesorhizobium sp. Cicer in chickpea. Microbiol Res. 2001;156(4):353-358.
Crossref - Reetha S, Selvakumar G, Thamizhiniyan P, Ravimycin T, Bhuvaneswari G. Screening of cellulase and pectinase by using Pseudomonas fluorescens and Bacillus subtilis. Int Lett Nat Sci. 2014;31(2):75-80.
Crossref - Rafanomezantsoa P, Gharbi S, Karkachi N, Kihal M. Optimization of amylase production by the biological control agent Bacillus halotolerans RFP74 using response surface methodology. J Genet Eng Biotechnol. 2023;21(1):63.
Crossref - Prihatiningsih N, Asnani A, Djatmiko HA. Extracellular protease from Bacillus subtilis B315 with antagonistic activity against bacterial wilt pathogen (Ralstonia solanacearum) of chili. Biodiversitas Journal of Biological Diversity. 2021;22(3):1291-1295.
Crossref - Gond SK, Bergen MS, Torres MS, White Jr JF. Endophytic Bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol Res. 2015;172:79-87.
Crossref - Toure Y, Ongena M, Jacques P, Guiro A, Thonart P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J Appl Microbiol. 2004;96(5):1151-60.
Crossref - Hazarika DJ, Goswami G, Gautom T, et al. Lipopeptide mediated biocontrol activity of endophytic Bacillus subtilis against fungal phytopathogens. BMC Microbiol. 2019;19(1):1-13.
Crossref - Ramyabharathi SA, Raguchander T. Characterization of antifungal antibiotic synthesis genes from different strains of Bacillus subtilis. J Pure Appl Microbiol. 2014;8(3):2337-2344.
- Pradhan S, Choudhury A, Dey S, Hossain MF, Saha A, Saha D. Siderophore-producing Bacillus amyloliquefaciens BM3 mitigate arsenic contamination and suppress Fusarium wilt in brinjal plants. J Appl Microbiol2023;134(10):lxad217.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.