


ISSN: 0973-7510
E-ISSN: 2581-690X
The goal of this study was to isolate and characterize phages that might be used in transport and treatment of potential E.coli pathogen in water sources. E. coli KP005067 was especially targeted because this is the dominant strain obtained from the sewage canal. The abundance of phage and its host directly from the sewage water was analysed using Epifluorescence Microscopy. The morphological characterization of the bacteriophages was studied using transmission electronic microscopy (TEM) and the genomic studies using Agarose Gel Electrophoresis. Studies on phage growth curve and its survival rate with respect to UV irradiation was also carried out. Almost 64% of the phages produced large, clear plaques indicating lytic activity. Phages had long noncontractile tail and icosahedral capsid head and therefore belonged to the Siphoviridae. Their estimated genome sizes was about 31 kb. One-step growth kinetics of the phage showed the latent period 15 min, rise period 20 min, and the average burst size 35 phage particles/infected cell. Also a clear downward trend in lytic activity was observed in phage groups exposed to longer than 30 minutes of UV. The characterization of the phages will be helpful in its future application as microbial tracers and bio-disinfectants in water sources.
Bacteriophage, Sewage, Characterization, Lytic, Siphoviridae.
Phages are predators of host bacteria which although are able to persist in the environment independently of their hosts, have the absolute requirement to infect host cells in order to multiply and survive as dynamic populations1. They are highly specific in their host-cell recognition, often only infecting specific members of a single genus and as such offer an alternative natural method to reduce bacterial pathogens2. This high level of specificity leads to little disturbance of commensal bacteria through the oral consumption of phages targeted at pathogens, and while bacteria develop specialized phage-defence mechanisms, phages also continuosly adapt to these changed host systems3.
Escherichia coli is one of the predominant species present in the gut of warm blooded animals and, is used as an indictor of the possible contamination of water, food or surfaces with human or animal faeces. It is also one of the most important bacteria that might be transported to the groundwater sources, through on-site human waste disposal. This pathogenic bacteria responsible for water-borne diseases are spread by the fecal-oral route, in which untreated water may play an immediate role. Unfortunately these pathogenic bacteria do not respond to the traditional water purification methods due to its high antibiotic resistance. Bacteriophages have evolved to be the most important and useful probes for pathogen transport and survival since they are closest in size, shape, and surface properties to these pathogens. The relative simplicity by which phages can be isolated from the environment can be attributed to the fact that, the natural environment of the bacteria of interest is likely to contain the specific phages capable of infecting and lysing the microorganism4.
The advantages in the use of phages in tracer and treatment studies over chemical agents is numerous. First of all, phages are highly host specific. They only infect specific bacterial hosts and cause rapid bacterial lysis. They do not infect humans and other eukaryotes. Phages specific for pathogenic bacteria do not disrupt normal microflora in humans or in animals5. Secondly, phages are not toxic to humans. Although certain cell lysis may release endotoxins, phages themselves do not generate any toxic products during their multiplication6. Thirdly, phages are stable7 and they do not replicate unless their bacterial hosts are present8. Moreover, phages are the most abundant biological entities and naturally present in the environment and a wide variety of foods9. It is relatively easy to isolate phages from the environment and propagate them in laboratories.
The goal of this research was therefore to isolate phages that were active against E.coli KP005067 and to characterize them with respect to morphology, genome size and UV irradiation activity before, exploring its possibility as microbial tracers and bio-disinfectants.
Bacterial strain and growth conditions
In this study, Escherichia coli KP005067 obtained from Canoli canal, Kozhikode district, Kerala, India was selected as the host bacteria for phage isolation. For experiments, the strain was cultured on EMB agar, Macconkey agar and Nutrient agar. Incubation was done at 37°C for 24 h and the working cultures were taken from EMB plates. For enrichment of the strain, a colony from nutrient agar was emulsified in a glass tube with alkaline peptone water (5-10ml) and incubated for 6-8h at 37°C.
Sample collection
Sewage samples were collected from stagnant water at a depth of 3 m, in sterile containers from 11 sites of Canoli canal, Kozhikode district, Kerala, India. Precautions were taken while collecting the samples by wearing sterile gloves and masks. Approximately 100 ml of sewage samples were collected and transported immediately to the laboratory and stored at 4°C, until processing.
SYBR staining
The viral and bacterial abundance in the sewage sample was assessed using Epifluorescence Microscopy. Bacteria like particles and virus like particles were enumerated from sewage samples according to10 using 0.8 µm nitrocellulose membrane support filters (Millipore) and 0.02 µm filters (Whatman).
Bacteriophage isolation
Bacteriophages were isolated from the collected sewage samples by enrichment technique.11 Under sterile conditions, 4.5 ml of sewage sample was emulsified with 0.5 ml of tryptone broth and 0.5 ml of log-phase broth culture (optical density measured at 600 nm) of Escherichia coli KP005067. Suspension was incubated overnight at 37°C, with shaking (120 rev/minutes) and was centrifuged at 10,000 rpm for 10 minutes at 4°C. The supernatant was filtered through a 0.22 µm syringe filter (Nupore, India) to obtain a bacterial free filtrate and analyzed for the presence of phages through spot test method12. Double layer plaque technique was performed for bacteriophage isolation13, 14. Three millilitre of 0.6% of trypticase soy agar (TSA) was inoculated with 100 µl of 3-5 hrs old bacterial culture and 100 µl of the bacteria free filtrate. The suspension was plated on fresh 1.5% TSA plate and incubated in an upright position at 37°C overnight. Formation of plaques or zones of clearance indicated the presence of phages15.
One-step Growth Kinetics
One step growth experiments were carried out based on the method described by16, 17 with some modifications. Briefly, the experiment started at a multiplicity of infection (MOI) 0.1 in a 15 ml tube containing phage (approximately 4 X 106 PFU/ml) and its host E.coli strain KP005067 (approximately 2X 108 CFU/ml) in 10 ml TSB. After incubation in water bath at 37°C for 10 min (to allow phage adsorption), the tube was centrifuged at 13000 rpm for 15 min. The supernatant was removed and subjected to plaque assay to determine the titre of the un-absorbed phage. The pellet containing (partially) infected cells was immediately re-suspended in 10 ml of pre-warmed TSB. After taking the first sample, the tube was returned to the water bath (37°C). A sample (100 µL) was collected every 5 min (up to 60 min). Each sample was immediately diluted and subjected to plaque assay. Latent period was defined as the time interval between the end of adsorption and the beginning of the first burst, as indicated by the initial rise in phage titer15, 18. Burst size was calculated as the ratio of the final number of liberated phage particles to the initial number of infected bacterial cells during the latent period15.
Electron Microscopy
Bacteriophage morphology examination was performed as described by19 with some modifications. Phage particles were sedimented by centrifugation at 25,000 rpm for 60 min. Phages were washed twice in 0.1 M ammonium acetate pH 7.0 (Vetec, Rio de Janeiro, Brazil), filtered through a 0.22-µm-pore-size membrane filter, deposited on copper grids provided with carbon-coated formvar films and examined in a Zeiss EM109 electron microscope.
Extraction of phage DNA
Phage DNA was extracted from the purified phages by using sodium dodecyl sulfate (SDS) and proteinase K and the DNA was precipitated by adding 3 M sodium acetate and absolute ethanol.21 The pellet was dried and resuspended in an appropriate volume of sterile high-purity water and stored at “20°C. The DNA samples were simultaneously checked on 0.8% agarose gel along with DNA ladder (ë DNA digested with EcoRI/HindIII).
UV Irradiation on phage replication
Purified phages were diluted using SM buffer to concentrations of 10-3, 10-4, 10-5 and 10-6. Sample trials suggested that most effective results would be seen at 10-3, so all future trials were run at that concentration. Four 100 µL drops of virus were placed close together on an empty Petri dish and placed under the UV lamp, at every 15 min, one drop was removed and placed into a micro centrifuge tube along with 400 µL of E.coli KP005067. The tubes were mixed well and kept at 37°C for 10 min. After incubation, 250 µL of the virus and bacteria mixture was added to 11 ml of warm soft agar and poured onto a Petri dish. The dishes were incubated overnight, and the number of clear zones on the plate were counted.
SYBR staining and viral count
SYBR green was made to stain double stranded DNA, so the probability that particles observed were eventually phage, was higher. By using 0.015µ pore size nuclepore filter, numerous small DNA-associated particles together with large DNA- associated particles were recognized from the sewage samples (Fig.1). Our criteria for the distinction between larger DNA-associated particles (possibly bacteria) and other smaller particles (phages) were to check the abundance of phage and its host bacteria in the water samples. The results showed a very high concentration of phages in the collected water samples. Enumeration of Escherichia coli and somatic coliphages from polluted sewage pipe, by filtering the water samples through 0.02 µm pore-size filters, stained with SYBR Green was done by.22 Also, SYBR green is reported to stain both RNA and DNA particles. Even though RNA viruses are likely to make up only a minor fraction of the total pool of viruses based on information from surveys,23 studies have proved its application in determining the abundance of total virus particles in the water body.
 Fig. 1. SYBR staining of bacteria and virus like particles
Fig. 1. SYBR staining of bacteria and virus like particlesPhage assays and titre value
Sewage in general contains a large diversity of coliforms due to fecal contamination. Therefore, sewage water is a reservoir of enteric pathogens. The results of plaque assay and spot assay are shown in table 1. The development of clear zones of lysis against the host bacterium using specific phage lysate indicated that 64% of the phages isolated were lytic phages. It was found that the virulent phages were able to lyse the host bacterium by developing clear circular zones with average plaque diameter of 5-7mm. Studies on bacetriophages have revealed that the process is initiated when the virion interacts with host cell surface receptor molecules.24 Out of the eleven sewage samples, only 45% were positive for plaque assays. S4 showed highest plaque assay with 86 PFU/ml, while S7 had the lowest: 21 PFU/ml. The presence of these zones of clearing indicated that amplifying bacteriophage from a raw sewage sample and inoculating the phage into the host was an effective way of isolating (and visualizing) phage.25 Amplification of phages in the sewage samples allow them to infect and reproduce within fresh E. coli. The infection was further spread as the virus reproduced resulting in the cell lysis and eventually forming visible plaques. Many bacteriophages are known to be highly specific for their receptors and show little or no interaction with receptors with an even slightly different structure. This specificity forms the basis of numerous phage typing methods for the identification of bacterial species or subspecies. The results obtained thus clearly indicated that, the bacteriophages for E. coli KP005067 were highly specific against its respective host. The results of our study suggest that, host specific bacteriophages against human pathogens are prevalent and can be readily isolated from sewage ecosystems. Present findings are in agreement with the work done by23 where, they had isolated host range lytic bacteriophages from sewage.
Table (1):
Plaque and spot formation in different water samples.
Samples # |
Plaque assay in PFU/ml |
Spot assay |
|---|---|---|
S1 |
48 |
+ |
S2 |
ND |
– |
S3 |
42 |
+ |
S4 |
86 |
+ |
S5 |
ND |
+ |
S6 |
39 |
+ |
S7 |
21 |
+ |
S8 |
ND |
+ |
S9 |
ND |
– |
S10 |
ND |
– |
S11 |
ND |
– |
ND- Not detected
One step growth kinetics
The latent period, rise period and the burst size of the phage were determined by countable number of plaques and phage titre (titer: plaque-forming unit (p.f.u)) during 10, 20, 30, 40, 50, and 60 minutes. Figure 2 shows the one step growth curve of bacteriophage. The latent period was only 15 min (excluding 10 min for adsorption), which is shorter than the typical latent periods (21-120 min). The rise period began after 20 minutes, with host cell lysis completed after 30 minutes releasing 35 progeny virions per infected cell. Similar results were obtained in the phage growth curve, where there was a steady low level of viruses during the latent period, which was followed by a sudden increase in the number of viruses or a rise period.26 Important characteristics of the viral reproductive process such as, the number of bacteria released per infected cell and the time it takes to produce new viruses can readily be determined by plotting a single step growth curve. These important characteristics vary between viruses but are often similar for related ones. Thus, they can serve as taxonomic criteria, mostly for high-level taxonomy. the number of released phages or burst size, and the time it takes to make them or latent period, are mainly determined by the complexity and size of the virion so it is expected that these criteria are conserved between phages with similar morphology.27
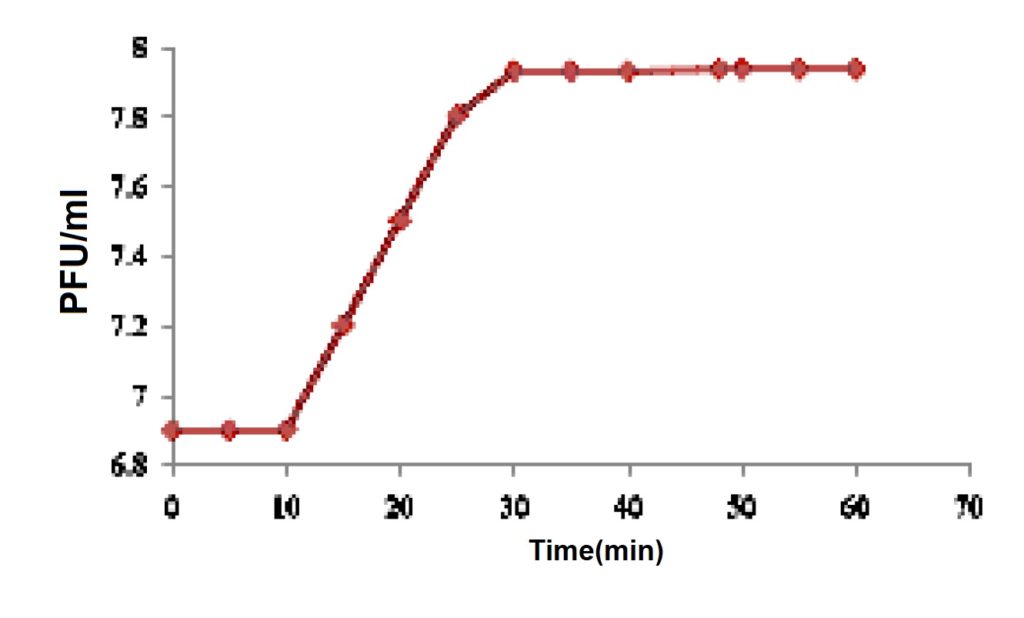 Fig. 2. One step growth curve of Escherichia coli phage
Fig. 2. One step growth curve of Escherichia coli phageMorphological analysis by TEM
Analysis of phages using electron microscopy allowed the morphological classification of phage into viral family and order. Isolated phage is classified into order Caudovirales and since it possess a long noncontractile tail and icosahedral capsid head, thus is a member of the family Siphoviridae.28 Heads were measured between opposite apices with an approximate 49.7±5.37 nm in size and the length of tails was 143 ± 3.25 nm in size (Fig 3). Only DNA coliphages, mostly tailed phages, were isolated during the sampling period. The detection of tailed phages agrees with previous reports that suggest that somatic DNA coliphages are easier and faster28 and that tailed phages account for over 90% of all known phages26. The phage morphology observed in this study agrees with the somatic coliphage morphological characters reported in a river system.
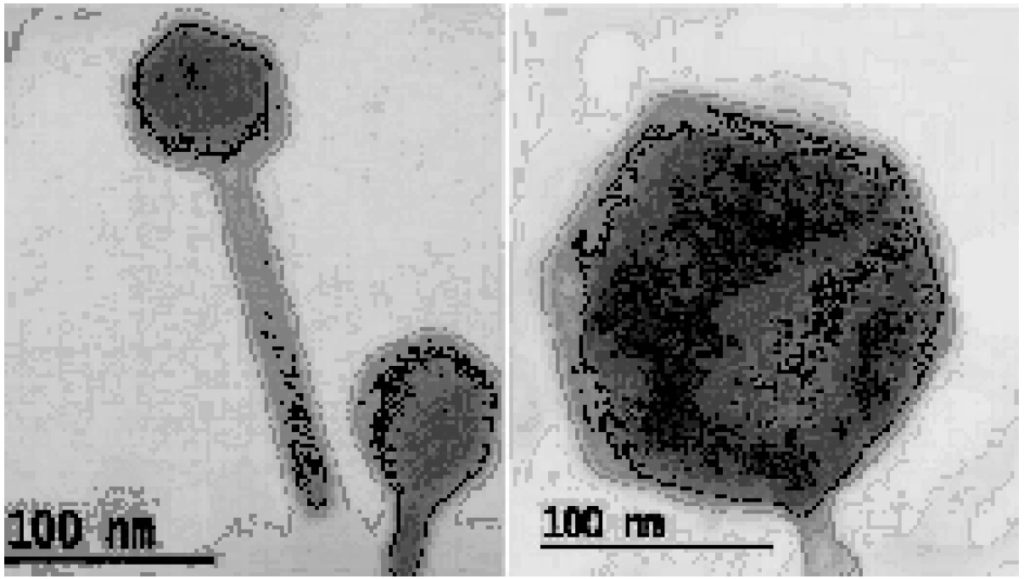 Fig. 3. Electron microscope image of Escherichia coli phage
Fig. 3. Electron microscope image of Escherichia coli phagePhage genome quantification
The DNA of the phage was isolated and electrophoresed on 0.8% agarose gel with a ë Hind/III marker. Molecular weight of the phage DNA was found to be 31 kb. For molecular characterization of phages, the isolated phage DNA samples were subjected to digestion with type II restriction enzyme Hindf I and BsuRI. The result revealed that the phages were sensitive to Hindf I and BsuRI, and exhibited different banding patterns confirming that the phages harboured double stranded DNA as genetic material (fig 4). Bacteriophages, with larger genome size of 16 kb or greater was found to be in the virus order of Caudovirales.29 Also, phages with larger genome size have higher gene number, greater infectivity and virion sophistication than phages with smaller genome size. The genome size of an E.coli phage isolated from sewage canal of Haryana also revealed a double-stranded DNA with approx. 33.5 kb.30 This DNA size was found to be smaller than the standard lambda DNA.
 Fig. 4. Extraction of phage DNA using Agarose Gel Electrophoresis
Fig. 4. Extraction of phage DNA using Agarose Gel ElectrophoresisUV Irradiation studies
The number of plaques were counted for each length of time and compared to the control group corresponding to that trial. The averages were taken and plotted as percentages in Fig 5. As seen in the plot, the control group had a significantly higher survival rate than any of the groups that were exposed to UV. Comparing plaque count of irradiated phage to non-irradiated phage showed that, the lytic activity decreased with increased exposure to UV. There was a small difference in lytic activity between the 15 and 30 minute trials, but after that, a clear downward trend is seen in groups exposed for longer than 30 minutes. Table 2 shows the calculated standard deviations of each time segment. The UV radiation can cause 5% loss in viable phage per hour for surface water due to the formation of thymine dimer.31 Thus it is evident from the results that, over time lytic activity decreased falling in a steady decline for exposure times greater than 30 minutes.
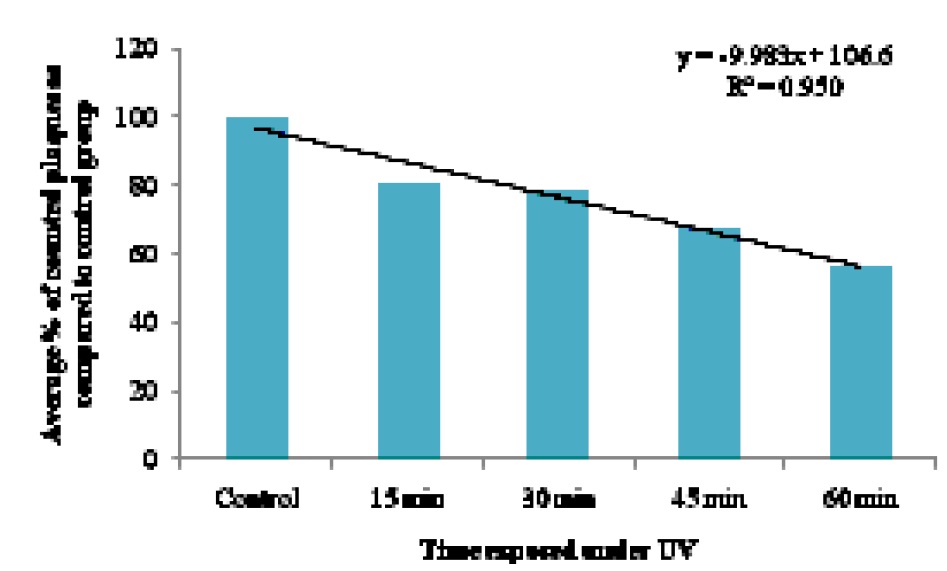 Fig. 5. Trend of phage survival when exposed to UV
Fig. 5. Trend of phage survival when exposed to UVTable (2):
Percentage error of average plate count compared using standard deviation.
Time |
Average plaque count |
% of Control |
|---|---|---|
Control |
149.41 ± 28.63 |
100 |
15 |
120.50 ±28.71 |
80.64 |
30 |
117.33 ± 21.13 |
78.52 |
45 |
101.33± 15.44 |
67.81 |
60 |
84.41± 20.56 |
56.49 |
Somatic phages and bacteriophage f2 were also found to be completely inactivated (3 log unit reduction) on exposure to less than 3h of UV irradiation.32 Also the persistence of somatic coliphages were found to be longer when compared to fecal coliforms, enterococci and F-RNA phages when exposed to UV light.27 So our results suggest that, the compelling biological attributes of coliphages makes them a potential candidate as microbial tracer and as bio-disinfectant.
Sewage in general contains a large diversity of coliforms due to fecal contamination therefore, it is a reservoir of enteric pathogens. The phages obtained in this experiment were found to be lytic due to the development of clear zones of lysis on the host bacterium. Studies on bacteriophages infection have revealed that the process was initiated when the virion interacts with host cell surface receptor molecules. Also the isolated coliphage was presumptively identified as a member of Siphoviridiae family with its morphological and genomic analysis. In conclusion, most of the bacteriophage isolates collected from the sewage samples of Canoli canal area were able to infect Escherichia coli KP005067. However, more precise studies are to be carried out to identify the potential application of these phages as microbial tracers and bio-disinfectants.
- Freifelder, D. Microbial Genetics. Jones and Bartlett Publishers, Boston; 1987
- Callaway, T. R., Edrington, T. S., Braban, A. D., Anderson, R.C.,Rossman, M.L., Engler, M. J.,Carr, M. A., Genovese, K. J. Bacteriophages isolated from feedlot cattle can reduce Escherichia coli O157:H7 populations in ruminant gastrointestinal tracts. Food Pathog Dis; 2008; 5, pp 183-191
- 3. Hagens, S. and Loessner, M. J. Application of bacteriophages for detection and control of foodborne pathogens. Appl Microbial Biotechnol 2007; 76, pp 513-519
- Gill, J. J., and P. Hyman.Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010; 11, pp 2-14.
- Kudva I. T., Jelacic S., Tarr P. I., Youderian P., Hovde C. J. Biocontrol of Escherichia coli O157 with O157-specific bacteriophages. Appl. Environ. Microbiol., 1999; 65, pp 3767–3773.
- Hagens S., Loessner M. J. Bacteriophage for biocontrol of foodborne pathogens: calculations and considerations. Curr. Pharm. Biotechnol., 2010; 11, pp 58–68
- Coffey, B., Mills S., Coffey, A., McAuliffe, O., Ross, R. P. Phage and their lysins as biocontrol agents for food safety applications. Annu. Rev. Food Sci. Technol., 2010;1, pp 449–468
- Guenther, D., Huwyler, S., Richard, M., Loessner, J. Virulent bacteriophage for efficient biocontrol of Listeria monocytogenes in ready-to-eat foods. Applied and Environmental Microbiology., 2009; 75: pp 93–100
- Patel, A. Virus and prokaryote enumeration from planktonic aquatic environments by epifluorescence microscopy with SYBR Green I. Nat. Protoc.,2007; 2: pp 269–276
- Cerveny, K. E., DePaola, A.. Duckworth, D. H., Gulig, P. A. Phage therapy of local and systemic disease caused by Vibrio vulnificus in iron-dextran-treated mice. Infect Immun., 2002; 70(11), pp 6251-62.
- Chang, H. C., Chen. C. R., Lin, J. W., Shen. G. H., Chang, K. M., Tseng, Y. H. Isolation and characterization of novel giant Stenotrophomonas maltophilia phage phiSMA5. Appl Environ Microbiol., 2005; 71(3): pp 1387 93.
- Cornax, R., Moriñigo, M. A., Paez, I. G., Muñoz, M.A., Borrego, J. J. Application of direct plaque assay for detection and enumeration of bacteriophages of Bacteroides fragilis from contaminated-water samples. Appl Environ Microbiol., 1990; 56(10), pp 3170-3177.
- Sillankorva, S., Neubauer, P., Azeredo, J. Isolation and characterization of a T7-like lytic phage for Pseudomonas fluorescens. BMC Biotechnol., 2008; 14(3), pp 592-597
- Adams, M. Bacteriophages. New York: Interscience; 1959; pp 137-59.
- Leuschner, R. G. K., Arendt, E. K., Hammes, W. P. Characterization of a virulent Lactobacillus sake phage PWH2. Appl. Microbiol. Biotechnol., 1993; 39: pp 617–621
- Ellis, E. L., Delbruck, M. The growth of bacteriophage. J. Gen. Physiol., 1939; 22: pp 365–384
- Oliveira, A. Isolation and characterization of bacteriophages for avian pathogenic E. coli strains. Journal of Applied Microbiology, 2009; 106: pp. 1919 – 1927.
- Maniatis, T., Fritsch, E. F., Sambrook, J. Molecular Cloning: A Laboratory Manual. New York: Cold Spring Harbour Laboratory Press; 1982.
- Maria, Ilaria, Leopizzi., Loredana, Stabili., Federica, Lugoli., Antonella, De, Donno. Virioplankton and bacterioplankton dynamic in an italian coastal area of the Southern adriatic sea subjected to pollution. Rapp. Comm. int. Mer Médit., 2010; 39.
- Frank, H., Moebus, K. An electron microscopic study of bacteriophages from marine waters. Helgol Meeresunters 1987; 41: pp 385-414
- Hayes, W. The genetics of bacteria and their viruses (Blackwell Scientific Publications Ltd. London, United Kingdom), 2nd ed; 1968.
- Steele, P. R. M. Morphological manifestations of freezing and thawing injury in bacteriophage T4Bo. J. Hyg., 1971; 77: 119.
- Jensen, E.C. Prevalence of borad-host-range lytic bacteriophaegs of Sphaerotilus natans, Escherichia coli, and Pseudomonas aeruginosa. Applied and Environmental Microbiology., 1998; 64(2), pp. 575 – 580.
- Jarvis, A. W. Bacteriophages of lactic acid bacteria. J Daiy Sci 1989; 72: pp 3406-3428.
- Kaiser, P. , L., Rothwell, E. E., Galyov, P. A., Barrow, J., Burnside., P. Wigley. Differential cytokine expression in avian cells in response to invasion by Salmonella typhimurium, Salmonella enteritidis, and Salmonella gallinarum. Microbiology., 2000; 146: pp 3217–3226.
- Maniloff, J. and Ackermann, H. W. Taxonomy of bacterial viruses: Establishment of tailed virus genera and the order Caudovirales. Arch Virol., 1998; 143(10): pp 2051-2063.
- Muniesa, M., Lucena, F. and Jofre, J. Study of the potential relationship between the morphology of infectious somatic coliphages and their persistence in the environment. J Appl Microbiol., 1999; 87: pp 402–409
- Ackermann, H.W. and Dubow, M.S. Viruses of Prokaryotes. Boca Raton, Florida: CRC Press, Inc,1987.
- Abedon, S.T. Size does matter-distinguishing bacteriophages by genome length (and ‘breadth’). Microbiology Australia., 2011; 32(2), pp. 90 – 91.
- Vipin, Singh., Pranay, Jain., and Swati, Dahiya.Isolation and characterization of bacteriophage from waste water against E. coli, a food borne pathogen. Asian Jr. of Microbiol. Biotech. Env. Sc., 2016; 18(1), pp 163-170.
- Wommack, K.E. Effects of sunlight on bacteriophage viability and structure. Applied and Environmental Microbiology., 1996; 62(4), pp 1336 – 1341.
- Davies-Colley, R. J., Craggs, R. J., Park, J., Sukias, J. P., Nagels, J . W., Stott, R. Virus removal in a pilot-scale ‘advanced’ pond system as indicated by somatic and F-RNA bacteriophages. Water Sci. Technol., 2005; 51: pp 107–110.
© The Author(s) 2017. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.