


ISSN: 0973-7510
E-ISSN: 2581-690X
Bacteriocins are proteins secreted by many species of bacteria to inhibit other bacteria, thus eliminating competitors to gain resources. Bacteria from the Lactobacillus group are known for their applications as probiotics and food preservatives. They have earned a reputation for producing substances that inhibit the growth of other microorganisms, which include organic acids, diacetyl, and bacteriocins. Produced by the ribosomes, bacteriocins are cationic proteins that inhibit other bacteria coexisting within a shared ecological habitat. Due to their potential uses in a variety of applications large-scale production of Bacteriocins would be necessary. The study aimed to identify and characterize Lactobacillus bacteria that produce potent bacteriocins and to analyze the antimicrobial activity and stability of the isolated bacteriocin under various physical and biochemical conditions. A total of 50 samples including buffalo dung, cheese, and rhizospheric region of plants were screened to isolate 8 Lactobacillus Li-1, Li-2, Li-3, Li-4, Li-5, Li-6, Li-7, and Li-8, confirmed by gram staining and other biochemical tests. The cell free supernatant from the Li-3 strain showed higher inhibition of Escherichia coli and Staphylococcus aureus, as compared to the other isolated strains. Li-3 strain was further identified as Levilactobacillus brevis strain ABRIINW-K by 16S rRNA gene sequencing. The bacteriocin isolated from this strain is a thermostable peptide (~6kDa), which is characteristic of class II bacteriocins, with potent antibacterial activity against Lactobacillus rhamnosus, Escherichia coli, and Salmonella enterica.
Bacteriocin, Lactobacillus brevis, Antimicrobial Activity, 16S rRNA, Antimicrobial Peptides
In recent years, the exploration of natural compounds with antimicrobial properties has garnered significant attention due to the escalating global concern over antibiotic resistance and its potential impact on public health. Bacteriocins, a class of ribosomally synthesized antimicrobial peptides, have emerged as a promising substitute for traditional antibiotics.1-3 These peptide molecules exhibit a broad spectrum activity against pathogenic microorganisms, including bacteria, making them a subject of intense research within the fields of food preservation, pharmaceuticals, and clinical therapeutics.4,5
Among the sources of bacteriocins, lactic acid bacteria (LAB) have gained special attention for their ability to produce these bioactive compounds.6-8 Lactobacillus species, a prominent group of LAB, has received wide attention for their probiotic properties and their role in maintaining gut health. In addition to their probiotic attributes, certain strains of Lactobacillus have shown the capacity to produce potent bacteriocins that exhibit inhibitory effects against closely related and even distantly related pathogenic bacteria.4,9,10 This dual functionality of some Lactobacillus strains has spurred a growing interest in the characterization of the bacteriocins produced by them.11-13
The strain Lactobacillus brevis ABRIINW-K, isolated from an unexplored ecological niche, presents an intriguing opportunity for bacteriocin discovery. The genetic diversity of LAB strains, reflected in their distinct 16S rRNA sequences, contributes to variations in their metabolic profiles, antimicrobial capabilities, and probiotic potential. Understanding the genetic and functional attributes of L. brevis ABRIINW-K’s bacteriocin holds promise for harnessing its antimicrobial properties for applications in food safety, microbial ecology, and biomedical interventions.13
This research paper aims to delve into the isolation and characterization of bacteriocin accomplished by L. brevis ABRIINW-K. By employing a combination of biochemical, molecular biology, and analytical techniques, we seek to elucidate the structural properties, mode of action, spectrum of activity, and potential applications of the isolated bacteriocin. The findings of this study may contribute to expanding the repertoire of natural antimicrobial agents available for combating bacterial infections and provide insights into the broader ecological roles of bacteriocin-producing LAB strains.14-16
In the following sections, we present the methodology employed for bacteriocin isolation, purification, and characterization from Lactobacillus brevis ABRIINW-K isolated from buffalo dung. Additionally, we will discuss the results of our investigations, drawing comparisons to existing literature and highlighting the significance of our findings in the context of antimicrobial research and application.17-19
All laboratory-based experimental studies were conducted at the Microbiology laboratory, IFTM University, Moradabad, Uttar Pradesh, India. The indicator organisms used in the study are Lactobacillus rhamnosus (MTCC 1408), Escherichia coli (MTCC 1687), Staphylococcus aureus (MTCC 737), and Salmonella enterica (MTCC 3858) were obtained from MTCC, Chandigarh, India.
Isolation and screening of Lactobacillus strains
50 samples including buffalo dung, cheese, and rhizospheric region of plants were examined for the presence of bacteriocin-producing Lactobacillus. Buffalo dung is rich in organic matter and provides a nutrient-rich environment. Bacteria that produce bacteriocins can survive and thrive in such environments by limiting competition from other microorganisms that might utilize the available resources. Similarly, some bacteriocin-producing bacteria are naturally present in milk, the primary ingredient in cheese. These bacteria can survive the cheese-making process and become part of the cheese microbiota. In addition, the rhizosphere is a highly competitive environment with a diverse microbial community vying for resources such as nutrients and space. Bacteriocin-producing bacteria can inhibit or kill competing microbial species, giving them an advantage in establishing and maintaining their populations that’s why these habitats were selected to isolate the bacteria. Strains were identified by gram staining and other biochemical tests like Gram staining to confirm its Gram-positive nature, the catalase test, and the catalase test, both of which should yield negative results. Carbohydrate fermentation, H2S, indole, motility, urease, MR-VP, and Oxidase were also assessed and developed in 50.0 mL MRS broth (HiMedia) at 30°C for 48 hours in a rotary shaker. After incubation, cell-free supernatant was obtained from each culture for screening of bacteriocin production. The broth was centrifuged at 10,000 rpm for 15 minutes at 4°C and the antibacterial capabilities of the supernatant against E. coli and S. aureus was assumed by well diffusion assay in Mueller Hinton agar (HiMedia) plates as described earlier.20-22 As per our findings, Lactobacillus strain Li-3 was used for further studies due to the higher activity of its cell supernatant against both indicator organisms.
Molecular identification of Li-3 strain
Genomic DNA of the Li-3 strain has been purified utilizing a QIAamp DNA kit (Qiagen) and selective 16S rRNA gene has been amplified utilizing the forward as well as reverse primers 5’-GGATGAGCCCGCGGCCTA-3’ and 5’-CGGTGTGTACAAGGCCCGG-3’ respectively.23 The PCR conditions were in the following order: the 5 minutes of initial denaturation at 96oC, denaturation at 96°C for 30 seconds, 30 seconds needed for annealing at 50°C, 1 minute and 30 seconds for the prolongation at 60°C.24 Sanger dideoxy sequencing was carried out as previously described.25,26 Applied Biosystems’ ABI 3130 genetic analyzer was used for the sequencing process and chemistry big dye terminator version 3.1 cycle sequencing kit using a 50 cm capillary array, POP-7 Polymer, and BDTv3KB Denovo v5.2 protocol. Data was performed on the Applied Biosystems SeqScape v5.2 software. The organism has been observed by comparing the sequencing data to the NCBI database utilizing the BLAST (BasiciLocal Alignment Search Tool).24-27
Bacteriocin isolation and antibacterial activity
For bacteriocin isolation, 50 mL MRS broth with the Li-3 strain, hereon designated as L. brevis ABRIINW-K, was grown at 30°C for 48 hours with continuous shaking. To partially purify bacteriocin from the bacterial fermentation broth, begin by centrifuging the broth at 10,000 rpm for 10 minutes at 4°C to pellet the bacterial cells, and then filtered the supernatant through a 0.22 µm membrane filter to remove residual cells and debris. Next, gradually added solid ammonium sulfate to the filtered supernatant to achieve 60% saturation, stirring gently until fully dissolved. Allowed the solution to precipitate at 4°C for overnight, then centrifuged at 10,000 rpm for 10 minutes at 4°C to collect the protein pellet. Dissolved the pellet in a minimal volume of Tris-HCl buffer (10 mM, pH 7.5) and dialyzed against a large volume of distilled water or Tris-HCl buffer at 4°C, changing the buffer several times over 24 hours to remove ammonium sulfate.22
The activity of the L. brevis ABRIINW-K bacteriocin was assayed by the well diffusion test method. 50 µL crude bacteriocin was added into 8 mm diameter wells in Muller Hinton agar (HiMedia) plates. Inoculum size was estimated by the McFarland method. Approximately 1.0 × 108, L. rhamnosus, E. coli, and Salmonella enterica were plated onto the agar surface and incubated at 37°C for 24 to 48 hours. Further, loss of activity after treatment with proteases confirmed the active component was a protein. Activity units per milliliter (AU mL -1) were determined by serial two-fold dilution of the Bacteriocin. One AU mL– 1 shows the reciprocal of the maximum dilution at which a distinct zone of inhibition was observed.22,27,28
Effect of temperature, pH, and proteases on bacteriocin activity
The inhibitory activity of L. brevis ABRIINW-K bacteriocin was assayed against indicator organisms upon exposure to different physical and biochemical conditions. To study the effect of temperature bacteriocin was heated at 50, 60, 70, 80, 90, and 100°C for 2 hours and at 121°C for 20 minutes, then antibacterial activity was assayed against indicator organisms. The stability of purified bacteriocin was assayed at pH 4, 4.5, 5, 5.5, 6, 6.5, 7.0, and 9.0, pH was adjusted with sterile 1M HCL and 1M NaOH. The impact of proteases on Bacteriocin was studied by treatment with 0.1% w/v each of pepsin, proteinase K, chymotrypsin, and trypsin (all from HiMedia) at 30°C for 2 hrs in separate reactions. The impact of detergents like Triton X-100, Tween 20, and Tween 80 was tested at 0.5% v/v concentration. Antibacterial activity for all tests was assayed in triplicates.21,22,27
Size estimation of Bacteriocin by SDS-PAGE
Polyacrylamide gel electrophoresis (SDS-PAGE) with sodium dodecyl sulfate was used to separate the partially purified bacteriocin with a 16% resolving gel utilizing the tris glycine buffer system, in a Mini-Protean Electrophoresis System (Bio-rad).29 A pre-stained 2.5 to 45 kDa molecular weight marker (Amersham Biosciences) was used. Following electrophoresis, the gel was predicted by staining with Coomassie BrilliantiBlue R-250 (HiMedia).21,30
Isolation and screening of Lactobacillus strains
In this study, eight Lactobacillus strains labeled Li-1 to Li-8 were isolated and identified through Gram staining and various biochemical tests. All strains demonstrated the ability to produce bacteriocins, aligning with the findings of previous studies on Lactobacillus bacteriocin production. The identification of these strains contributes to the growing body of research emphasizing the potential of Lactobacillus species in antimicrobial applications.
The bacteriocin production observed in our strains is consistent with earlier reports by Todorov et al.,13 who characterized bacteriocin-producing Lactobacillus strains isolated from traditional fermented foods. Similar to our findings, they reported the inhibitory activity of these bacteriocins against a range of pathogenic bacteria, suggesting a broad-spectrum antimicrobial potential.
The bacteriocin production observed in our Lactobacillus brevis strain UN is consistent with the findings reported by Gautam et al.31, who also characterized bacteriocin production by Lactobacillus brevis isolated from Dhulliachar, a traditional food product of North East India. Their study demonstrated the purification and characterization of this bacteriocin, revealing its broad-spectrum antimicrobial activity against several pathogenic bacteria, which aligns with our observations. This suggests that the specific conditions and microbial communities in traditional fermented foods like Dhulliachar may foster the development of potent bacteriocin-producing strains with significant antimicrobial properties.
In addition, the study by Jang et al.32 on Lactobacillus brevis KU15153, isolated from kimchi, supports our findings by highlighting the broad antimicrobial effects of bacteriocins produced by Lactobacillus brevis. They characterized the bacteriocin’s inhibitory effects against a variety of pathogens, similar to the spectrum observed in our study. The work by Jang et al. also emphasized the probiotic characteristics of Lactobacillus brevis, further underlining the potential health benefits of strains isolated from traditional fermented foods.
Moreover, Elayaraja et al.33 studied the bacteriocin produced by Lactobacillus murinus AU06 and reported a broad antibacterial spectrum, which echoes the broad-spectrum activity found in our Lactobacillus brevis strain UN. Their research, like ours, involved the purification and characterization of bacteriocins, adding to the body of evidence that bacteriocins from different Lactobacillus species can effectively inhibit a wide range of pathogens.
Finally, the work by Mounier et al.34 on bacteriocins in smear-ripened cheeses provides additional context by exploring the role of bacteriocins in the complex microbial ecosystems of fermented foods. Although their focus was on cheese, the principles of bacteriocin interaction within microbial communities and their preservation qualities are relevant to our study, as they highlight the versatility and importance of bacteriocins in food safety and microbial balance.
In a comparative study by Ogunbanwo et al.,35 the bacteriocins produced by Lactobacillus strains isolated from fermented food products exhibited notable antimicrobial activities. Their strains, like ours, were identified and characterized using Gram staining and biochemical tests, affirming the reliability and consistency of these methods for identifying bacteriocinogenic Lactobacillus species.
Furthermore, the study by Parvez et al.36 highlighted the importance of bacteriocin-producing Lactobacillus in probiotic applications. Our isolated strains may have similar probiotic potential, given their ability to inhibit pathogenic bacteria. This potential is further supported by our preliminary observations on the stability and activity of the bacteriocins under various conditions, akin to the stability characteristics reported by Yang et al.37
Our study adds to the existing literature by not only confirming the bacteriocin-producing capability of these eight Lactobacillus strains but also by preserving these strains for future research. This preservation is crucial for detailed genetic and proteomic analyses to understand the mechanisms underlying bacteriocin production and their applications in food preservation and human health.
In this study, we isolated eight Lactobacillus strains from diverse environments: the rhizospheric region of plants (Li-1 and Li-2), fecal samples of buffalo dung (Li-3 and Li-4), and cheese samples (Li-5 to Li-8). All strains demonstrated the ability to produce bacteriocins, as evidenced by the inhibitory activity of their supernatants against pathogenic bacteria. The protease sensitivity of these bacteriocins suggests that their antibacterial activity is proteinaceous, aligning with the characteristics of known bacteriocins.
Comparative studies have similarly reported the isolation of bacteriocin-producing Lactobacillus strains from varied sources. For instance, Todorov et al.13 isolated Lactobacillus strains from traditional fermented foods, which exhibited significant antimicrobial activity, akin to our findings with strains from cheese samples. Their study also emphasized the importance of the source in determining the bacteriocin spectrum and potency, a trend observed in our results, particularly with the superior activity of Li-3.
The inhibitory activity of Li-3, isolated from buffalo dung, was notably high against both Escherichia coli and Staphylococcus aureus, outperforming the other strains. This is consistent with Ogunbanwo et al.,35 who reported robust bacteriocin activity from Lactobacillus strains isolated from animal sources, suggesting that such environments might harbor strains with potent antimicrobial properties. The enhanced activity of Li-3 could be attributed to the unique microbial interactions and competitive pressures in the gut microbiota of buffalo, which select for highly antagonistic bacteria.
Our findings are also in agreement with the work of Parvez et al.,36 who highlighted the probiotic potential of bacteriocin-producing Lactobacillus strains. The inhibition of common pathogens by Li-3 underscores its potential application in probiotic formulations aimed at improving gut health and preventing infections. Moreover, the study by Yang et al.37 on the stability and activity of Lactobacillus bacteriocins under various conditions further supports the feasibility of using Li-3 in diverse applications, ranging from food preservation to clinical therapeutics.
The protease sensitivity of the bacteriocins produced by our strains indicates that their antimicrobial components are proteins, as demonstrated by previous research.35,36 This characteristic is crucial for their functional categorization and potential application in environments where protein stability can be managed.
The exceptional performance of Li-3, in particular, warrants further investigation. Future research will focus on the molecular characterization of its bacteriocin, assessing its genetic basis, mode of action, and spectrum of activity. Additionally, its application in real-world scenarios, such as food safety and as a probiotic, will be explored. Understanding the precise mechanisms by which Li-3 exerts its antimicrobial effects will be crucial for developing effective strategies to harness its potential.
In conclusion, this study not only confirms the bacteriocin-producing capabilities of Lactobacillus strains from diverse sources but also identifies the Li-3 strain as a particularly potent candidate for further exploration. These findings contribute to the expanding knowledge of Lactobacillus-derived bacteriocins and their applications, with the potential to develop natural and effective antimicrobial solutions.
Isolation and screening of Lactobacillus strains
Eight Lactobacillus cultures were isolated and identified by gram staining and other biochemical tests (Table 1). All 8 strains were found to produce Bacteriocins and were preserved for future research. These strains were labelled from Li-1 to Li-8.
Table (1):
Biochemical identification of Lactobacillus strains
Characteristics |
Lactobacillus rhamnosus (MTCC 1408) |
Li-3-strain Levilactobacillus brevis |
|---|---|---|
Gram stain |
Gram-positive |
Gram-positive |
H2S |
Negative |
Negative |
Catalase |
Negative |
Negative |
Indole |
Negative |
Negative |
Motility |
Negative |
Negative |
Urease |
Negative |
Negative |
MR-VP |
Negative |
Negative |
Oxidase |
Negative |
Negative |
Li-1 and Li-2 were isolated from the rhizospheric region of plants, Li-3 and Li-4, were isolated from fecal samples of buffalo dung, and Li-5, Li-6, Li-7, and Li-8 were isolated from cheese samples. The inhibitory activity of the supernatant preparations from each organism containing crude bacteriocins is presented in Table 2. Treatment of the supernatants with proteases resulted in inhibition of their antibacterial activity. Lactobacillus strain Li-3 showed the highest inhibition of both E. coli and S. aureus
(Figure 1). Therefore, the Li-3 strain was considered for further exploration of its bacteriocin properties.
Table (2):
Screening of Bacteriocin activity of Lactobacillus strains
Indicator strain |
Li-1 |
Li-2 |
Li-3 |
Li-4 |
Li-5 |
Li-6 |
Li-7 |
Li-8 |
|---|---|---|---|---|---|---|---|---|
E. coli |
+ |
+ |
+++ |
+ |
++ |
– |
– |
– |
S. aureus |
– |
– |
+++ |
– |
– |
– |
– |
– |
(-); No inhibition, (+); 1-5 mm low inhibition, (++); 5-10 mm moderate inhibition, (+++); 10 mmi high inhibition

Figure 1. Antibacterial activity of Lactobacillus isolated from buffalo dung, (1) Well A: Li-3, Well B: Li-4, Well C: Li-4, and Well D: negative control. (2) Well A: negative control, Well B: Li-4, Well C: Li-3, and Well D: Li-3
Molecular identification of Li-3 strain
About 1,500 bp region of the Li-3 16S rRNA gene has been amplified (Figure 2) and sequenced. Analysis of the amplified sequence using BLAST search on the NCBI server showed 99.52% similarity to Lactobacillus brevis ABRIINW-K strain (Sequence ID JN368471.1). Phylogenetic analysis of the strain is presented in Figure 3. Owing to their probiotic potential, bacteriocin-producing L. brevis has also been isolated and characterized from diverse sources like traditional food products, various plants, animals, and human sources.31,32

Figure 2. Agarose gel electrophoresis of (1) Amplified region of 16S rRNA gene of the Li-3 strain with ~1500bp size, (L) 500bp DNA ladder

Figure 3. Phylogenetic tree of Levilactobacillus brevis strain ABRllNW-K using the Fast Minimum Evolution method in the Blast Tree View Widget on NCBI server
Bacteriocin purification from L. brevis ABRIINW-K and antibacterial activity
The partially purified L. brevis ABRIINW-K bacteriocin obtained by dialysis showed active inhibition of all indicator strains L. rhamnosus, Salmonella enterica, and E. coli upon 24 to 48 hours of incubation at 37°C (Figure 4). Results are presented in Table 3. It is evident from past studies that bacteriocin from Lactobacillus species has the potential to inhibit or kill closely related as well as other groups of bacteria.
Table (3):
Antibacterial activity of L. brevis ABRIINW-K bacteriocin
Indicator strain |
Antibacterial activity (AU mL-1) |
|---|---|
L. rhamnosus |
6400 |
Salmonella enterica |
3200 |
E. coli |
3200 |
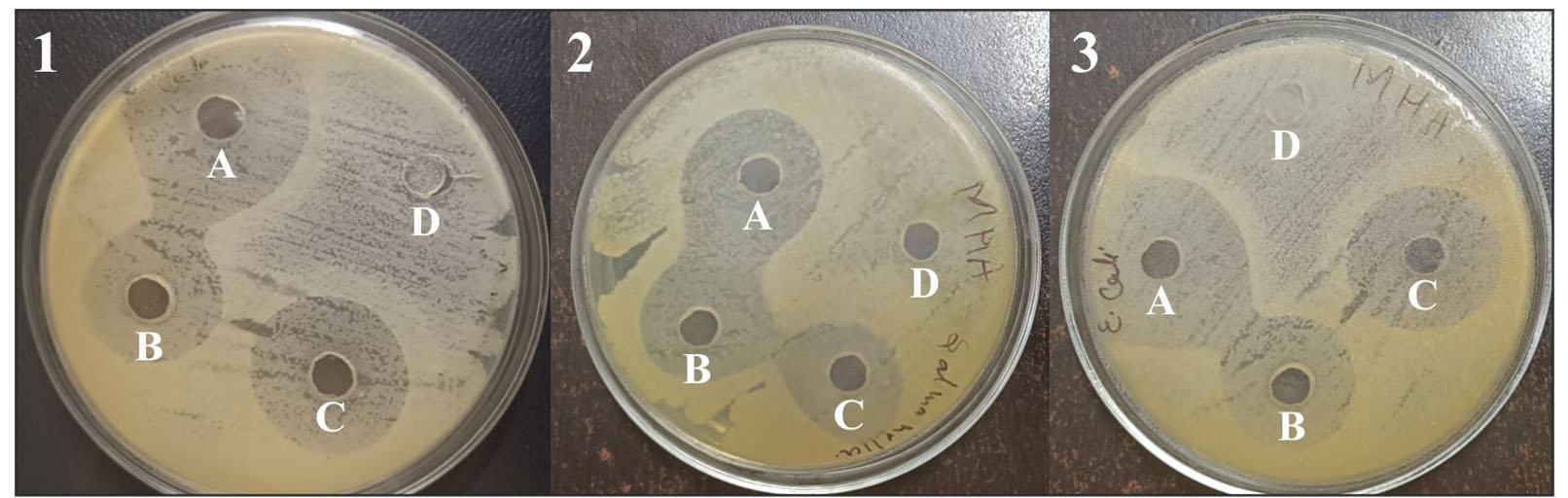
Figure 4. Antibacterial activity of L. brevis ABRIINW-K bacteriocin showing zone of inhibition against indicator strains (1) L. rhamnosus, (2) Salmonella enterica, and (3) E. coli. 0.1 mL of the bacteriocin was added to wells A, B, and C in each plate. 0.1 mL sodium acetate buffer was added to well D as a negative control
Effect of temperature, pH, and proteases on bacteriocin activity
The partially purified L. brevis ABRIINW-K bacteriocin heated at 50, 60, 70, 80, 90, and
100°C for 2 hours, showed no changes in their activity against indicator strains, but lost complete activity against indicator strains when heated at 121°C for 20 min, thus confirming its thermostable characteristics.33 The bacteriocin remained stable between pH 5.5 to 8.5 and showed a decrease in its potency beyond this pH range. Complete bacteriocin activity has been eliminated after treatment with all of the proteases pepsin, proteinase K, chymotrypsin, and trypsin (Figure 5). Since bacteriocins are peptides that are sensitive to proteolytic enzymes.24
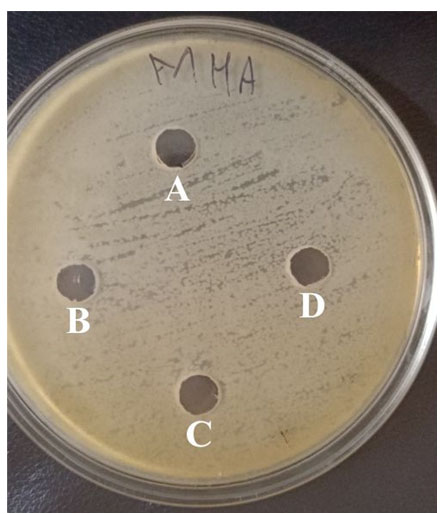
Figure 5. The antibacterial activity of bacteriocin after treatment with proteases. (A) Pepsin, (B) Trypsin, (C) Chymotrypsin, and (D) Proteinase K treated bacteriocin were added to each well in plates spread with E. coli culture
Size estimation by SDS-PAGE and classification of bacteriocin
SDS-PAGE analysis of the L. brevis ABRIINW-K bacteriocin compared to low molecular weight protein marker presented a clear band of approximately 6 kDa (Figure 6). Contaminant proteins were detected at lower concentrations. The low molecular weight and thermostability of the bacteriocin correspond to characteristics of class II bacteriocins as confirmed by earlier studies.34
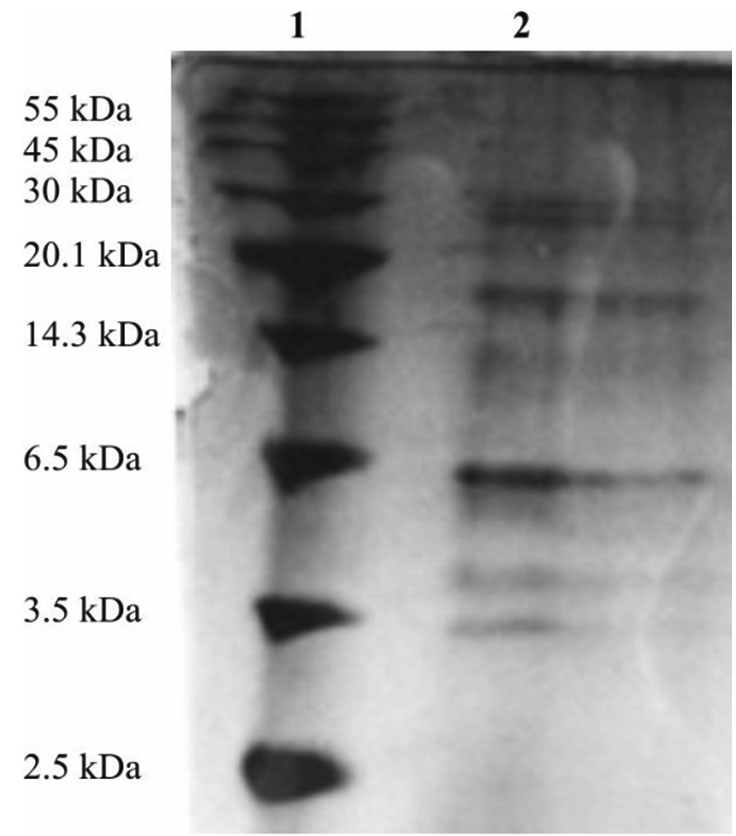
Figure 6. Size estimation of Bacteriocin by SDS-PAGE. Lane 1 Molecular weight marker, and Lane 2: Partially cleaned L. brevis ABRIINW-K bacteriocin
Cell-free supernatants from eight Lactobacillus species showed antibacterial properties against various foodborne pathogens, suggesting potential use as a natural antimicrobial agent in food production. This could be a substitute for preservatives in organic foods. A potent bacteriocin from the Lactobacillus strain, L. brevis ABRIINW-K, was isolated and characterized, confirming its broad-spectrum activity and potential as a novel antimicrobial agent. Future research should focus on purification, structural elucidation, in vivo efficacy studies, and safety assessments. The data collected may also help explore other unexplored bacteriocins and microorganisms, enriching our understanding of their diversity and functional significance within microbial ecosystems.
In summary, these reports add to the expanding knowledge of bacteriocins and their promise as natural substitutes for conventional antibiotics. The combination of molecular, biochemical, and biophysical techniques has allowed us to unravel the intricate details of this bacteriocin’s properties, providing a foundation for further research in areas such as food preservation, medical therapeutics, and biopreservation.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Gunasekaran T, Gopalakrishnan A, Balaraman D, Kathirkaman P. Chitinolytic Bacterial Disease Infected in Scylla serrata (Forsskal, 1775) Fattening Farm of Tamil Nadu. Int J Life Sci. 2018;7(2):90-95.
Crossref - Mathur H, Field D, Rea MC, Cotter PD, Hill C, Ross RP. Bacteriocin-Antimicrobial Synergy: A Medical and Food Perspective. Front Microbiol. 2017;8:1205.
Crossref - Turgis M, Vu KD, Jamshidian M, Maherani B, Lacroix M. Synergistic antimicrobial effect of combined bacteriocins against food pathogens and spoilage bacteria. Microbiol Res Int. 2016;4(1):1-5.
- Pieterse R, Todorov SD. Bacteriocins – exploring alternatives to antibiotics in mastitis treatment. Braz J Microbiol. 2010;41(3):542-562.
Crossref - Ortiz-Rivera Y, Sanchez-Vega R, Gutierrez-Mendez N, Leon-Felix J, Acosta-Muniz C, Sepulveda DR. Production of reuterin in a fermented milk product by Lactobacillus reuteri: Inhibition of pathogens, spoilage microorganisms, and lactic acid bacteria. J Dairy Sci. 2017;100(6):4258-4268.
Crossref - Hammami R, Zouhir A, Le Lay C, Ben Hamida J, Fliss I. BACTIBASE second release: a database and tool platform for bacteriocin characterization. BMC Microbiol. 2010;10:22.
Crossref - Soltani S, Hammami R, Cotter PD, et al. Bacteriocins as a new generation of antimicrobials: toxicity aspects and regulations. FEMS Microbiol Rev. 2021;45(1):fuaa039.
Crossref - Kabuki T, Uenishi H, Watanabe M, Seto Y, Nakajima H. Characterization of a bacteriocin, Thermophilin 1277, produced by Streptococcus thermophilus SBT1277. J Appl Microbiol. 2007;102(4):971-980.
Crossref - Tagg JR, Dajani AS, Wannamaker LW. Bacteriocins of gram-positive bacteria. Bacteriol Rev. 1976;40(3):722-756.
Crossref - Lelise A, Belaynesh G, Mulubrhan M, Kedija S, Endashaw B, Abebe B. Isolation and screening of antibacterial producing lactic acid bacteria from traditionally fermented drinks (“Ergo” and “Tej” ) in Gondar town, Northwest Ethiopia. Global Research Journal of Public Health and Epidemiology; 2014;1(3):18-22
- Asare PT, Greppi A, Stettler M, Schwab C, Stevens MJA, Lacroix C. Decontamination of Minimally-Processed Fresh Lettuce Using Reuterin Produced by Lactobacillus reuteri. Front Microbiol. 2018;9:1421.
Crossref - Cotter PD, Ross RP, Hill C. Bacteriocins – a viable alternative to antibiotics? Nat Rev Microbiol. 2013;11(2):95-105.
Crossref - Todorov SD, Furtado DN, Saad SMI, Tome E, Franco BDGM. Potential beneficial properties of bacteriocin-producing lactic acid bacteria isolated from smoked salmon. J Appl Microbiol. 2011;110(4):971-986.
Crossref - Cocolin L, Foschino R, Comi G, Fortina MG. Description of the bacteriocins produced by two strains of Enterococcus faecium isolated from Italian goat milk. Food Microbiol. 2007;24(7-8):752-758.
Crossref - Mathara JM, Schillinger U, Kutima PM, Mbugua SK, Holzapfel WH. Isolation, identification and characterisation of the dominant microorganisms of kule naoto: the Maasai traditional fermented milk in Kenya. Int J Food Microbiol. 2004;94(3):269-278.
Crossref - Kang JH, Lee MS. Characterization of a bacteriocin produced by Enterococcus faecium GM-1 isolated from an infant. J Appl Microbiol. 2005;98(5):1169-1176.
Crossref - O’Shea EF, O’Connor PM, O’Sullivan O, Cotter PD, Ross RP, Hill C. Bactofencin A, a new type of cationic bacteriocin with unusual immunity. mBio. 2013;4(6):e00498-00413.
Crossref - Serna CL, de Stouvenel AR. Lactic acid production by a strain of Lactococcus lactis subs lactis isolated from sugar cane plants. Electron J Biotechnol. 2006;9(1).
Crossref - Rantsiou K, Drosinos EH, Gialitaki M, Metaxopoulos I, Comi G, Cocolin L. Use of molecular tools to characterize Lactobacillus spp. isolated from Greek traditional fermented sausages. Int J Food Microbiol. 2006;112(3):215-222.
Crossref - Alias R, Idrus ES, Lani M, et al. Antibacterial Activity of Lactic Acid Bacteria Isolated from Goats’ Milk. 2012.
- Hassan MU, Nayab H, Rehman TU, et al. Characterisation of Bacteriocins Produced by Lactobacillus spp. Isolated from the Traditional Pakistani Yoghurt and Their Antimicrobial Activity against Common Foodborne Pathogens. BioMed Res Int. 2020;e8281623.
Crossref - Malheiros PS, Sant’Anna V, Todorov SD, Franco BDGM. Optimization of growth and bacteriocin production by Lactobacillus sakei subsp. sakei2a. Braz J Microbiol. 2015;46:825-834.
Crossref - Verma M, Mishra V. Bioelectricity Generation Using Sweet Lemon Peels as Anolyte and Cow Urine as Catholyte in a Yeast-Based Microbial Fuel Cell. Waste Biomass Valorization. 2023;14(8):2643-2657.
Crossref - Rossi E, Ali A, Efendi R, et al. Characterization of Bacteriocin Produced by Lactic Acid Bacteria Isolated from Solid Waste of Soymilk production. IOP Conf Ser Earth Environ Sci. 2021;709(1):012020.
Crossref - Santos NC, Cabrera MEB, Flores JD, et al. Molecular Identification and Phylogenetic Analysis of Bioluminescent Bacteria Isolated from Selected Marine Crustacean in Manila Bay, Philippines. J Pure Appl Microbiol. 2023;17(4):2119-2130.
Crossref - Singh RK, Tiwari S, Mishra VK, Tiwari R, Dhole TN. Molecular epidemiology of Chikungunya virus: Mutation in E1 gene region. J Virol Methods. 2012;185(2):213-220.
Crossref - Sevim E, Karaoglu SA, Sevim A, Canakci S. Antimicrobial Activity of Bacillus Strains Isolated from Spring Water and A Novel Bacteriocin: RS108. J Pure Appl Microbiol. 2013;7(4):2757-2765.
- Ribeiro SC, O’Connor PM, Ross RP, Stanton C, Silva CCG. An anti-listerial Lactococcus lactis strain isolated from Azorean Pico cheese produces lacticin 481. Int Dairy J. 2016;63:18-28.
Crossref - Merzoug M, Dalache F, Zadi Karam H, Karam NE. Isolation and preliminary characterisation of bacteriocin produced by Enterococcus faecium GHB21 isolated from Algerian paste of dates “ghars.” Ann Microbiol. 2016;66(2):795-805.
Crossref - Tulini FL, De-Martinis ECP. Improved adsorption-desorption extraction applied to the partial characterization of the antilisterial bacteriocin produced by Carnobacterium maltaromaticum C2. Braz J Microbiol Publ Braz Soc Microbiol. 2010;41(2):493-496.
Crossref - Gautam N, Sharma N, Ahlawat OP. Purification and Characterization of Bacteriocin Produced by Lactobacillus brevis UN Isolated from Dhulliachar: a Traditional Food Product of North East India. Indian J Microbiol. 2014;54(2):185-189.
Crossref - Jang HJ, Lee NK, Paik HD. Probiotic characterization of Lactobacillus brevis KU15153 showing antimicrobial and antioxidant effect isolated from kimchi. Food Sci Biotechnol. 2019;28(5):1521-1528.
Crossref - Elayaraja S, Annamalai N, Mayavu P, Balasubramanian T. Production, purification and characterization of bacteriocin from Lactobacillus murinus AU06 and its broad antibacterial spectrum. Asian Pac J Trop Biomed. 2014;4(Suppl 1):S305-311.
Crossref - Mounier J, Coton M, Irlinger F, Landaud S, Bonnarme P. Chapter 38 – Smear-Ripened Cheeses. Cheese (Fourth Edition). 2017:955-996.
Crossref - Ogunbanwo ST, Sanni AI, Onilude AA. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. African Journal of Biotechnology, 2003;2(8):219-227.
- Parvez S, Malik KA, Ah-Kang S, Kim HY. Probiotics and their fermented food products are beneficial for health. Journal of applied microbiology, 2006;100(6): 1171-1185.
Crossref - Yang E, Fan L, Jiang Y, Doucette C, Fillmore S. Antimicrobial activity of bacteriocin-producing lactic acid bacteria isolated from cheeses and yogurts. AMB express, 2012;2:1-12.
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.