


ISSN: 0973-7510
E-ISSN: 2581-690X
The increasing prevalence of multidrug-resistant (MDR) pathogens necessitates the discovery of novel antibacterial agents. In this study, culturable soil bacteria from the Dead Sea region (Jordan) were isolated and the antibacterial activity of their extracellular metabolites were evaluated. Twenty-one isolates were screened using a perpendicular-streak primary assay; three stable producers (GH-15, GH-20, GH-21) were prioritized. Cell-free supernatants were extracted with n-hexane and ethyl acetate, tested using agar well diffusion, and profiled using thin-layer chromatography (TLC) coupled with bioautography (GH-15). Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) were determined by broth microdilution against Bacillus subtilis ATCC 11774, B. cereus ATCC 10876, Staphylococcus aureus ATCC 29213, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, and Proteus mirabilis ATCC 29906. The ethyl acetate extract of GH-15 showed the lowest MIC against E. coli (0.073 mg/mL). The MIC values were equal to MBC for several strains, indicating bactericidal activity. For GH-15 hexane extract, the lowest MIC was 2.625 mg/mL (E. coli and B. cereus), and the lowest MBC was 5.25 mg/mL (E. coli and B. cereus). Dead Sea soils yielded bacterial isolates with broad antibacterial activity; solvent-dependent activity and TLC-bioautography suggested multiple bioactive metabolites, supporting further purification and chemical identification.
Antibiotics, Soil, Dead Sea, Bacteria, Bioautography, MIC, MBC
Antibiotics are natural or synthetic substances that are primarily produced as secondary metabolites by microorganisms with antimicrobial properties.1 A major source of antibiotics is the bacterial community, which is widely used in human healthcare. Each year, approximately 500 antibiotics are discovered, most of which are obtained from soil bacteria.1-3 Antibiotics have revolutionized pharmacology and medicine and have successfully treated many infectious diseases.4 However, over time, many microbes have developed resistance to these drugs due to excessive use,5-7 which has severely reduced treatment options, making multidrug-resistant (MDR) pathogens a global public health problem.8 The misuse of antibiotics can result in the spread of resistant strains, posing public health risks worldwide; antibiotic-resistant bacteria (ARB) could lead to 10 million deaths by 2050.9,10
The Dead Sea is unique in comparison to other extremely salty environments. Its salinity exceeds 34%, and, in addition to other harsh conditions, such as low pH and a distinct ionic composition different from seawater, the Dead Sea supports a diverse ecosystem of halophilic microorganisms that have adapted to its extreme environment.11,12 The ability of these microorganisms to thrive under harsh conditions has led to the production of natural microbial products with considerable potential for developing new antibiotics to address the growing issue of antibiotic resistance.13 In particular, bacteria have shown promise as a source of bioactive compounds and are likely to continue to be an important resource.14
Despite the unique environmental conditions of the Dead Sea, few studies have investigated the antibacterial activity of Dead Sea isolates. Accordingly, the current study aimed to isolate antibacterial agents from the Dead Sea soil bacteria and test the antibacterial activity of their extracts. Moreover, the activity of the secondary metabolites produced by a single isolate was investigated against different Gram-positive and Gram-negative bacteria. This study prioritized the isolation of stable antibacterial-producing bacteria from multiple Dead Sea sites and combined solvent extraction, TLC bioautography, and MIC/MBC determination to provide an initial bioactivity profile of Dead Sea-derived bacterial metabolites against both Gram-positive and Gram-negative reference strains. Hypothesizing that soil bacteria isolated from Dead Sea region would provide a source for secondary metabolites with promising antibacterial activity.
Nutrient broth was obtained from Sachar Lau (Spain), and nutrient agar was obtained from Biolab (Hungary). Additionally, soybean casein digestion medium (tryptone broth) and yeast malt broth (YM broth) were obtained from HiMedia (India). For antibacterial activity testing, reference strains including B. subtilis ATCC (11774), B. cereus ATCC (10876), S. aureus ATCC (29213), E. coli ATCC (25922), P. aeruginosa ATCC (27853), and P. mirabilis ATCC (29906) were obtained from Kwikstik (France). Tween 80 was purchased from SIGMA (France). For antibiotic extraction, n-hexane was purchased from ROMIL (Jordan) and ethyl acetate was purchased from Carlo Erba (France).
Methods
Soil sample collection
Nineteen samples were collected from nine different locations in the Dead Sea, Jordan (December 2019) according to a previously described method.13 Approximately 30 g of soil was collected from a 5-15 cm depth and preserved in a sterile disposable cup at room temperature (25 °C).
Bacterial isolation from soil samples
Rubble from the soil samples was removed before weighing 1 g of each collected sample. Serial dilutions of soil samples were prepared using normal saline (0.9% NaCl). From each dilution, (100 µL) was incubated in a nutrient agar plate for 24 hrs at 37 °C. Colonies with different morphologies were selected and streaked onto a new nutrient agar plate to obtain a pure isolate, according to the method described previously, with some modifications.14,2
Primary screening
The isolated bacteria were tested to determine their ability to produce antimicrobial activity against a group of pathogenic bacterial reference strains: B. subtilis ATCC (11774), B. cereus ATCC (10876), E. coli ATCC (25922), P. aeruginosa ATCC (27853), P. mirabilis ATCC (29906), and S. aureus ATCC (29213), according to the perpendicular streaking method.15 Each isolate was streaked in the middle of a nutrient agar plate and then incubated for 48 hrs at 37 °C. Pathogenic bacterial suspensions were adjusted to match 0.5 McFarland standard and streaked perpendicularly around the soil isolates. All plates were incubated again at 37 °C for another 24 hrs. Each experiment was repeated three times.
Morphological characterization
The colony morphology of the isolated bacteria with antimicrobial production potential was studied, including colony color and shape. The microscopic features of the isolated bacteria, including Gram staining and spore formation, were examined according to the Clinical and Laboratory Standards Institute (CLSI) guidelines.
Secondary screening
Production and downstream isolation of antibacterial substances
Three potential bacterial isolates that produced promising antibacterial activity were chosen for further production and testing, and they were coded as GH-15, GH-20, and GH-21. These isolates were chosen because of their stability in producing antimicrobial agents during primary screening. Each potential isolate was cultivated in a 500 ml flask containing 200 ml of tryptone soy broth medium and then incubated for up to 21 days at 37 °C in an orbital shaker. Cultured bacterial suspensions were then centrifuged at 11,000 rpm at 5 °C for > 5 min. The supernatant was filtered through sterile 0.45-µm cellulose membrane filters.16-18
Extraction of supernatant filtrate and testing of antimicrobial activity using the agar well diffusion method
Extraction was conducted according to a previously described method, with some modifications.19,20 The filtrates were extracted using a separating funnel with two organic solvents: hexane, followed by ethyl acetate. Each organic phase was collected separately and allowed to dry completely under a fume hood. Stock solutions (84 mg/mL hexane and 75 mg/mL ethyl acetate) were prepared and tested against B. subtilis ATCC (11774), B. cereus ATCC (10876), S. aureus ATCC (29213), E. coli ATCC (25922), and P. aeruginosa ATCC (27853) using the agar-well diffusion method.
Thin-layer chromatography (TLC)
TLC was conducted on hexane and ethyl acetate extracts of one isolate (GH-15) that showed the highest antibacterial activity during secondary screening, using (8 cm × 4 cm) aluminum sheet TLC plates under a ultraviolet-visible range of 250-366 nm.21 Extracts were applied to the sheets using Minicaps capillary pipettes (10 µL). The sheets were dipped into a mobile phase (toluene:acetone; 1:1) for separation. All TLC separations were conducted at room temperature (25 °C) and dried for 5 min to completely remove the solvent. The sheets were observed under UV light (366 nm). The cells were then stained with iodine.
Detection of the antibacterial activity by using the bioautography technique
Direct bioautography was conducted as described previously.22 Semi-solid nutrient agar was prepared and inoculated with bacterial suspension matching 0.5 McFarland standard for B. subtilis ATCC (11774) and E. coli ATCC (25922). Each TLC plate (8 × 4 cm) was transferred to an empty Petri dish. The inoculated agar was poured into each TLC plate and incubated at 37 °C for 24 hrs. Inhibition zones were examined, and the Rf values were used to determine the position of the active spots.
Minimum Inhibitory Concentration (MIC) and minimum Bactericidal Concentration (MBC)
The MIC was determined using the two-fold broth microdilution method in a 96-well plate.23 Each experiment was conducted in triplicate. A stock solution of the GH-15 extract was prepared by dissolving the extract in five drops of Tween 80 and distilled water to obtain a final concentration of 84 mg/mL for the hexane and 75 mg/mL for the ethyl acetate extract. The lowest concentration of the extract with no visible turbidity was considered as the MIC. Each extract with no visible growth was cultured on nutrient agar plates and incubated for 24 hrs at 37 °C to determine the MBC. The lowest concentration with no bacterial growth was considered as the MBC.
Collection of soil samples
One hundred different types of bacteria were successfully isolated from soil samples based on their colony morphology (Figure 1).
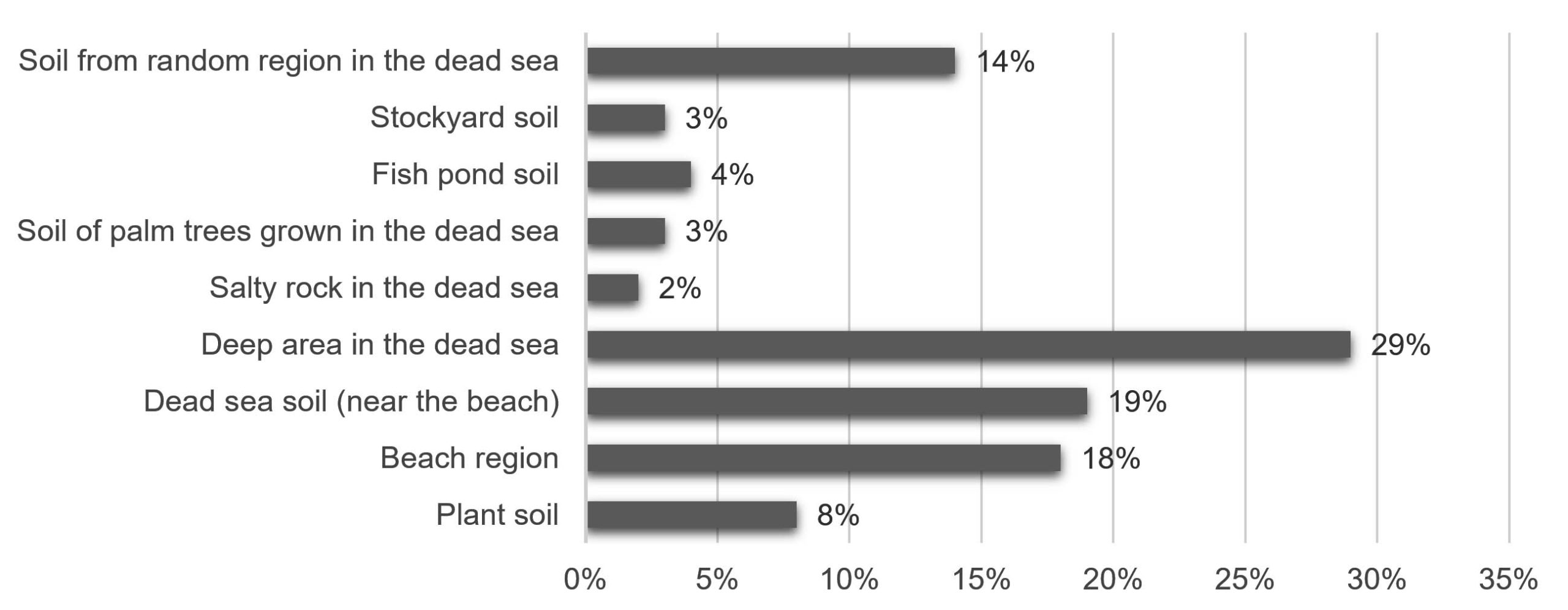
Figure 1. Percentage of bacteria isolated (100 isolates) from each collection site in the Dead Sea region
Primary screening
Each soil-isolated bacterium was cultured on the middle surface of the nutrient agar plates, and the test bacteria were cultured perpendicularly to evaluate their ability to produce antimicrobial agents (Table 1). Twenty-one of the 100 bacterial isolates successfully produced antibacterial substances against the reference strains. Most isolates were active against S. aureus and B. cereus whereas no activity was detected against P. aeruginosa or P. mirabilis.
Table (1): Antibacterial activity of 21 soil isolates against a group of Gram-positive and Gram-negative bacteria- primary screening using perpendicular streak
Isolates |
B. subtilis |
B. cereus |
S. aureus |
E. coli |
P. aeruginosa |
P. mirabilis |
|---|---|---|---|---|---|---|
GH-1 |
+ |
– |
+ |
– |
– |
– |
GH-2 |
+ |
– |
+ |
– |
– |
– |
GH-3 |
+ |
– |
+ |
± |
– |
– |
GH-4 |
+ |
– |
+ |
– |
– |
– |
GH-5 |
+ |
– |
+ |
– |
– |
– |
GH-6 |
+ |
– |
+ |
– |
– |
– |
GH-7 |
– |
– |
– |
± |
– |
– |
GH-8 |
+ |
– |
+ |
– |
– |
– |
GH-9 |
+ |
– |
+ |
– |
– |
– |
GH-10 |
+ |
– |
± |
± |
– |
– |
GH-11 |
+ |
– |
– |
– |
– |
– |
GH-12 |
+ |
– |
+ |
– |
– |
– |
GH-13 |
+ |
– |
+ |
– |
– |
– |
GH-14 |
+ |
– |
+ |
– |
– |
– |
GH-15 |
+ |
– |
+ |
– |
– |
– |
GH-16 |
+ |
– |
+ |
– |
– |
– |
GH-17 |
+ |
– |
+ |
– |
– |
– |
GH-18 |
+ |
– |
+ |
– |
– |
– |
GH-19 |
+ |
+ |
+ |
± |
– |
– |
GH-20 |
+ |
+ |
+ |
– |
– |
– |
GH-21 |
+ |
+ |
+ |
– |
– |
– |
(+) Active against tested bacteria; (-) Inactive against tested bacteria; ( ± ) Reduced activity against test bacteria
Morphological characterization
Morphological determination was conducted for bacterial isolates that had the potential to produce antibacterial substances during primary screening (Table 2).
Table (2): Morphology of Potential Bacterial Soil Isolates
Isolate |
Gram Stain |
Morphological Description |
|---|---|---|
GH-1 |
Gram-positive bacillus |
Cream-colored, round colony with raised margins; flat, moist, and adherent to the surface. |
GH-2 |
Gram-positive bacillus |
Cream-colored, irregular-shaped colony; flat with curled edges, shiny, and mucoid. |
GH-3 |
Gram-positive bacillus |
Light yellow-colored, punctiform, round colony with entire edges; highly adherent to the surface. |
GH-4 |
Gram-positive bacillus |
Cream-colored, round colony with soft texture and entire edges. |
GH-5 |
Gram-positive bacillus |
Cream-colored, mucoid, flat colony with irregular shape and lobate edges. |
GH-7 |
Gram-positive bacillus |
Shiny white-colored, round colony with entire edges; highly viscous. |
GH-9 |
Gram-positive (spore-forming) |
Cream-colored, irregular-shaped, flat colony with undulate edges; matte and moist. |
GH-10 |
Gram-negative bacillus |
Light green-colored, circular colony with entire edges; undulate, shiny, viscous, and transparent. |
GH-11 |
Gram-positive bacillus |
White-colored colony with smooth margins; convex-raised, small, round, shiny, and viscous. |
GH-12 |
Gram-positive bacillus |
Cream-colored, wrinkled, flat colony with lobate edges, curled appearance, irregular shape, slimy, and moist. |
GH-13 |
Gram-positive bacillus |
Yellowish-white colored, wrinkled colony with wavy edges; irregular-shaped, flat, matte, and dry. |
GH-15 |
Gram-positive bacillus |
Brownish cream-colored, punctiform, smooth, round colony with entire edges. |
GH-16 |
Gram-positive (spore-forming) |
Milky-colored colony with entire edges; raised, round, smooth, moist, and matte. |
GH-17 |
Gram-positive (spore-forming) |
Milky-colored colony with entire edges; convex, round, moist, and matte. |
GH-18 |
Gram-positive (spore-forming) |
Milky-colored colony with entire edges; convex, smooth, round, matte, and moist. |
GH-19 |
Gram-positive bacillus |
White-colored, punctiform, convex colony with undulate margins. |
GH-20 |
Gram-positive bacillus |
Yellow-colored, punctiform, flat colony with entire margins. |
GH-21 |
Gram-positive bacillus |
Yellow-colored, punctiform, flat colony with entire margins. |
Secondary screening
Bacterial isolates with promising antibacterial activities (GH-15, GH-20, and GH-21) were selected for secondary screening. Bacterial cultures were grown for 21 days. Five milliliters of the culture flask was collected daily to test the antibacterial activity of the supernatant. Activity was recorded as the diameter of the inhibition zone (Table 3).
Table (3): Antibacterial activity of GH-15, GH-20, GH-21 supernatant as inhibition zone (cm ± SD) against tested Gram-positive bacteria
Isolate |
Cultivation Time (h) |
S. aureus |
B. subtilis |
B. cereus |
|---|---|---|---|---|
GH-15 |
72 |
1.95 ± 0.05 |
2.0 ± 0.16 |
2.1 ± 0.3 |
GH-20 |
408 |
1.50 ± 0.01 |
1.85 ± 0.05 |
1.95 ± 0.05 |
GH-21 |
408 |
1.30 ± 0.01 |
1.80 ± 0.1 |
1.85 ± 0.15 |
Production and downstream isolation of target antibacterial compounds
Extraction of supernatant filtrate and testing of antimicrobial activity
The bacterial isolates were grown for the required time before centrifugation. The supernatant was extracted using hexane and ethyl acetate. The reconstituted extracts (84 mg/mL for hexane and 75 mg/mL for ethyl acetate) were active against the tested Gram-positive and Gram-negative bacteria (Table 4). All tested extracts showed activity against B. cereus and E. coli. However, not all extracts showed activity against B. subtilis, S. aureus, and P. aeruginosa.
Table (4): Antibacterial activity of GH-15, GH-20, and GH-21 extract as inhibition zone (cm ± SD) against Gram-positive and Gram-negative bacteria
Extract (Bacteria) |
B. subtilis |
B. cereus |
S. aureus |
E. coli |
P. aeruginosa |
|---|---|---|---|---|---|
Ethyl acetate extract (GH-15) |
4.05 ± 0.07 |
3.73 ± 0.17 |
4.15 ± 0.07 |
3.95 ± 0.07 |
2.6 ± 0.22 |
Hexane extract (GH-15) |
3 ± 0.01 |
4.78 ± 0.22 |
4.65 ± 0.21 |
5.15 ± 0.2 |
1.2 ± 0.14 |
Ethyl acetate extract (GH-20) |
4.3 ± 0.3 |
3.9 ± 0.17 |
4 ± 0.12 |
4.1 ± 0.1 |
2.4 ± 0.4 |
Hexane extract (GH-20) |
– |
3.25 ± 0.35 |
– |
3.5 ± 0.01 |
– |
Ethyl acetate extract (GH-21) |
3.85 ± 0.12 |
3.15 ± 0.2 |
3.35 ± 2.35 |
3.56 ± 0.2 |
– |
Hexane extract (GH-21) |
– |
3.65 ± 0.2 |
3.33 ± 0.2 |
3.93 ± 0.3 |
– |
(-) No activity
Thin layer chromatography (TLC)
TLC was conducted for the GH-15 extract to determine the number of bacterial metabolites. Both the hexane and ethyl acetate extracts were separated using the same mobile phase. Three spots were detected in the ethyl acetate extract and two spots were detected in the hexane extract.
Bioautography
TLC plates were inoculated with test bacteria grown on semi-solid agar to determine the active spots. Spots with Rf (0.727) for hexane and Rf (0.818) for ethyl acetate indicated the active compounds (Figure 2).
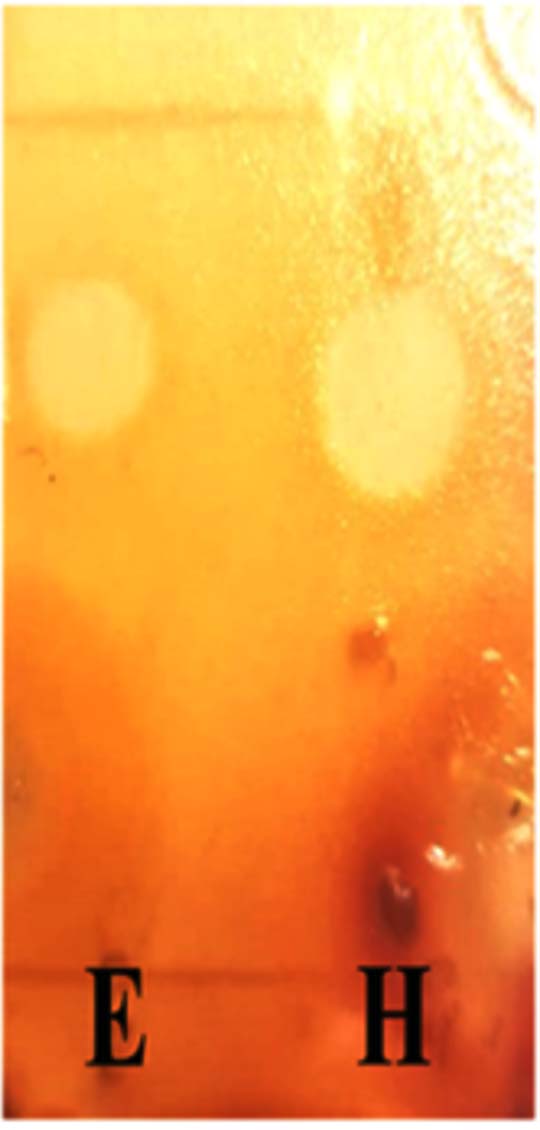
Figure 2. Bioautography. E: Ethyl acetate extract, H: Hexane extract, stained with iodine
MIC
The MIC values were examined against B. subtilis ATCC (11774), B. cereus ATCC (10876), S. aureus ATCC (29213), E. coli ATCC (25922), P. aeruginosa ATCC (27853), and P. mirabilis ATCC (29906) (Table 5). The lowest MIC of the acetate extract was against E. coli (0.073 mg/mL), whereas the hexane extract had the lowest MIC (2.625 mg/mL) against E. coli and B. cereus.
Table (5): MIC values for hexane and ethyl acetate extracts of GH-15 extracts against pathogenic bacteria
Bacterial Reference strains |
Ethyl acetate extract (mg/mL) |
Hexane extract (mg/mL) |
|---|---|---|
B. subtilis |
0.292 |
5.25 |
B. cereus |
0.292 |
2.625 |
S. aureus |
0.292 |
5.25 |
E. coli |
0.073 |
2.625 |
P. aeruginosa |
9.375 |
10.5 |
P. mirabilis |
1.17 |
10.5 |
MBC
All clear wells in the MIC test were cultured in nutrient agar to determine the MBC (Table 6). Ethyl acetate MIC was equal to the MBC against B. subtilis, S. aureus, E. coli and P. aeruginosa. The lowest MBC for the hexane extract was 5.25 mg/mL against E. coli and B. cereus.
Table (6): MBC values for hexane and ethyl acetate extracts of GH-15 extracts against pathogenic bacteria
Pathogenic strain |
Ethyl acetate extract (mg/mL) |
Hexane extract (mg/mL) |
|---|---|---|
B. subtilis |
0.292 |
10.5 |
B. cereus |
0.585 |
5.25 |
S. aureus |
0.292 |
10.5 |
E. coli |
0.073 |
5.25 |
P. aeruginosa |
9.375 |
10.5 |
P. mirabilis |
4.687 |
10.5 |
The rapid increase in antibiotic resistance by pathogenic bacteria has led to the detection of novel antibacterial agents.15 Soil bacteria represent one of the most promising frontiers for antibacterial agents discovery.16,17 Harsh environmental conditions, including high salinity and elevated temperatures, in the Dead Sea can lead to the dominance of tolerant bacteria that can be a source of promising secondary metabolites with antibacterial activity.13 The total number of different bacteria isolated was 100, which were selected based on morphological differences. Of the isolates, 21% passed the primary screening against pathogenic bacteria, a finding similar to that reported previously (Table 1).18,19 The 21 isolates were tested against six types of pathogenic bacteria using the agar-well diffusion method to evaluate the antibiotic activity produced in the supernatant. Potential isolates in the primary screening were examined according to colony shape, Gram staining, and cell shape. All isolates were Gram-positive bacteria (Table 2). The observed dominance of Gram-positive bacteria likely reflects environmental factors.
Secondary screening aimed to test the activity of the potential antibacterial agents produced in the supernatant.20 GH-15 exhibited maximal antibacterial activity after 72 h of culture, whereas GH-20 and GH-21 required prolonged incubation (408 hrs) (Table 3). This delayed activation of secondary metabolite production could be attributed to stress-induced changes in the metabolic pathways.21 Marked differences were observed in the antibacterial profile of the solvent extraction. Both the hexane and ethyl acetate extracts of GH-15 exhibited activities against B. subtilis, B. cereus, S. aureus, E. coli, and P. aeruginosa. The ethyl acetate extract of GH-20 demonstrated activity against all tested pathogenic bacteria, whereas the hexane extract showed activity against B. cereus and E. coli. The hexane extract of GH-21 exhibited activity against B. cereus, E. coli, and S. aureus. The ethyl acetate extract of GH-21 was active against all tested bacteria, except P. aeruginosa. This indicated that each isolate of (GH-20 and GH-21) produced at least two types of secondary metabolites with different antibacterial activities. The diversity of the antibacterial profile was consistent with that observed in other Dead Sea isolates.12,22
Thin-layer chromatography coupled with bioautography confirmed that the antibacterial activity was related to the separation of secondary metabolites. Three different spots were detected in the ethyl acetate extract and two spots were detected in the hexane extract under UV light. A bioautography test was conducted to determine the Rf of spots with antibacterial activity. The hexane extract of G15 demonstrated an active spot at Rf = 0.727, whereas the ethyl acetate extract showed an active spot at Rf =0.818. MIC and MBC were determined for both G15 extracts. The ethyl acetate extract showed better antibacterial activity than the hexane extract against all the tested bacterial strains. Superior activity of the ethyl acetate extract of silage isolated from Bacillus, compared to a hexane extract has been reported.23 Bacillus antimicrobial metabolites include polyketides, lipopeptides, and volatile compounds that are more soluble in ethyl acetate than in hexane.24,25
The lowest MIC for the hexane extract was detected against B. cereus and E. coli (2.625 mg/mL), and the highest MIC value was against P. mirabilis and P. aeruginosa (10.5 mg/mL) (Table 5). The ethyl acetate extract exhibited the highest antimicrobial activity against E. coli (0.073 mg/mL), with an MIC of 0.292 mg/mL against B. subtilis, B. cereus, and S. aureus. Antimicrobial activity was reduced against P. mirabilis (1.17 mg/mL) and demonstrated the weakest activity against P. aeruginosa. Ethyl acetate extract of marine-isolated Bacillus demonstrated greater antibacterial activity, according to MIC values, against E. coli (15.62 µg/mL) than its antimicrobial activity against B. subtilis, B. cereus, and S. aureus (31.25 µg/mL).26 However, MIC values of the ethyl acetate extract of marine-isolated Bacillus against P. aeruginosa and E. coli were equal.26 Ethyl acetate extract of silage-isolated Bacillus demonstrated the same antibacterial activity against B. subtilis and S. aureus (25 µg/mL).23
The MBCs of ethyl acetate extracts against B. subtilis, E. coli, and S. aureus and hexane extracts against P. mirabilis and P. aeruginosa were equal to their MIC (Table 6). The similarity between the MIC and MBC values against some bacterial strains indicates bactericidal activity. This is consistent with the results reported previously by Balogun et al. who tested Bacillus isolates from soil samples collected from a river bank and reported equal MIC and MBC values of the bacterial ethyl extract against S. aureus, B. subtilis, E. coli, and P. aeruginosa.27 Six promising antibacterial agents were identified from the three bacterial isolates. This paves the way for further evaluation of the extracted secondary metabolites for their identification and characterization.
Several bacterial isolates obtained from the Dead Sea were rich sources of antibacterial agents. Three bacterial isolates (GH-15, GH-20, and GH-21) showed superior activity against Gram-positive and Gram-negative bacteria. Secondary metabolite diversity was indicated by differences in antibacterial activity related to different extraction solvents. The antibacterial activity of (GH-15) extract against the tested bacteria was determined using MIC and MBC. Compound identity and purity were not established, and the activity may reflect a mixture of metabolites. Future studies should include molecular identification of the producing isolates (e.g., 16S rRNA sequencing), bioassay-guided fractionation, structural elucidation (liquid chromatography with tandem mass spectrometry, nuclear magnetic resonance), assessment of cytotoxicity, and expanded testing against clinical MDR isolates to confirm translational relevance. The ecological importance of the source of the isolates (representing extreme environmental conditions) and the broad activity of the isolates against different bacterial strains justify further investigation of the secondary metabolites produced by the bacterial isolates, which highlights the Dead Sea soil microbiome as a promising reservoir of antibacterial bacteria. The solvent-dependent variation in activity, low MIC values observed in certain extracts, and ecological uniqueness of the isolates collectively support further investigation into their bioactive metabolites.
ACKNOWLEDGMENTS
The authors would like to thank Isra University, Jordan, for their support.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
SMA and ND conceptualized the study. GHM applied methodology. DNL and AAT performed data validation. GHM performed formal analysis. SMA supervised the study. DNL wrote the original draft. SMA, AAT and ND wrote, reviewed and revised the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This research was funded by Isra University, Amman, Jordan.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Chandra N, Kumar S. Antibiotics producing soil microorganisms. In: Antibiotics and Secondary Metabolites in Soil Microorganisms. Springer; 2017:1-18.
Crossref - Santos-Aberturas J, Vior NM. Beyond soil-dwelling actinobacteria: fantastic antibiotics and where to find them. Antibiotics. 2022;11(2):195.
Crossref - Prashanthi R, Shreevatsa GK, Krupalini S, Manoj L. Isolation, characterization, and molecular identification of soil bacteria showing antibacterial activity against human pathogenic bacteria. J Genet Eng Biotechnol. 2021;19(1):1-14.
Crossref - Al-Turk A, Odat N, Massadeh MI. Isolation and molecular characterization of antibiotic producing Bacillus licheniformis strains isolated from soil. J Pure Appl Microbiol. 2020;14(4):2363-2370.
Crossref - Ahmad MN, Kuldeep R, Manabendra G. Microbial biofilm: formation, architecture, antibiotic resistance, and control strategies. Brazilian J Microbiol. 2021:1701-1718.
Crossref - Baran A, Kwiatkowska A. Antibiotics and Bacterial Resistance – A Short Story of an Endless Arms Race. Published online 2023.
- Yi R, Shi Y, Cao X, Pan C. Actinomycetes: Treasure trove for discovering novel antibiotic candidates. Eur J Med Chem. 2025;286:117317.
Crossref - Mattick P, Gillock E. Screening Environmental Soil Samples For Antibiotic Production. SACAD John Heinrichs Sch Creat Act Days. 2024;2024(2024).
- Ho CS, Wong CTH, Aung TT, et al. Antimicrobial resistance: a concise update. Lancet Microb. 2025;6(1):100947.
Crossref - Ding D, Wang B, Zhang X, et al. The spread of antibiotic resistance to humans and potential protection strategies. Ecotoxicol Environ Saf. 2023;254:114734.
Crossref - Rajab JA, Yogeshwar P, Tezkan B, Al-Halbouni D. Transient electromagnetic imaging of saltwater intrusion at the shrinking Dead Sea. Sci Rep. 2025;15(1):1-16.
Crossref - Al-daghistani HI, Zein S, Abbas MA. Microbial communities in the Dead Sea and their potential biotechnological applications. Commun Integr Biol. 2024;17(1):2369782.
Crossref - Thompson TP, Gilmore BF. Exploring halophilic environments as a source of new antibiotics. Crit Rev Microbiol. 2024;50(3):341-370.
Crossref - Cheema N, Qureshi AW. Isolation and Characterization of Antibiotic producing Lysobacter. Pak Biomed J. 2024;7(3):62-69.
Crossref - Ahmed SK, Hussein S, Qurbani K, et al. Antimicrobial resistance: Impacts, challenges, and future prospects. J Med Surgery Public Heal. 2024;2:100081.
Crossref - Sapkota A, Thapa A, Budhathoki A, Sainju M, Shrestha P, Aryal S. Isolation, Characterization, and Screening of Antimicrobial-Producing Actinomycetes from Soil Samples. Int J Microbiol. 2020;2020:2716584.
Crossref - Usha Nandhini S, Sudha S, Anusha Jeslin V, Manisha S. Isolation, identification and extraction of antimicrobial compounds produced by Streptomyces sps from terrestrial soil. Biocatal Agric Biotechnol. 2018;15:317-321.
Crossref - Demisie S, Oh D, Wolday D, et al. Diversity of culturable bacterial isolates and their potential as antimicrobial against human pathogens from Afar region, Ethiopia. Microbiol Spectr. 2024;12(11):e0181024.
Crossref - Perrier F, Morice J, Gueulle S, et al. Assessing Normandy Soil Microbial Diversity for Antibacterial Activities Using Traditional Culture and iChip Methods. Microorganisms. 2024;12(12):2422.
Crossref - Santos ACC, Malta SM, Dantas RCC, Coelho Rocha ND, Ariston de Carvalho Azevedo V, Ueira-Vieira C. Antimicrobial activity of supernatants produced by bacteria isolated from Brazilian stingless bee’s larval food. BMC Microbiol. 2022;22(1):127.
Crossref - Pardo-este C, Cortes J, Castro-severyn J, et al. Secondary metabolites with antimicrobial activity produced by thermophilic bacteria from a high-altitude hydrothermal system. Front Microbiol. 2024;15:1477458.
Crossref - Al-Karablieh N. Antimicrobial Activity of Bacillus Persicus 24-DSM Isolated from Dead Sea Mud. Open Microbiol J. 2017;11:372-383.
Crossref - Kadaikunnan S, Rejiniemon TS, Khaled JM, Alharbi NS, Mothana R. In-vitro antibacterial, antifungal, antioxidant and functional properties of Bacillus amyloliquefaciens. Ann Clin Microbiol Antimicrob. 2015;14(1):9.
Crossref - Sumi CD, Yang BW, Yeo I, Hahm YT. Antimicrobial peptides of the genus Bacillus: a new era for antibiotics. Can J Microbiol. 2015;62(2):93-103.
Crossref - Caulier S, Nannan C, Gillis A, Licciardi F. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front Microbiol. 2019;10:1-19.
Crossref - Ahmed E, Shawky H, Eissa MA, El-Rahim WMA, El-Refy A. Isolation and characterization of some potential antibacterial metabolites from marine microorganisms collected in Egypt. Egypt J Microbiol. 2024;59:51-61.
Crossref - Balogun OO, Ugoh SC, Oladosu PO. GC-MS Analysis and Antimicrobial Activity of the Extract and Fractions of Bacillus subtilis subsp. subtilis 168 Isolated from a River Bank in Nigeria. Microbiol Res J Int. 2022;32(2):46-59.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.