


ISSN: 0973-7510
E-ISSN: 2581-690X
The global surge in antimicrobial resistance has critically weakened the effectiveness of existing antibiotic regimens, highlighting an immediate need to identify and develop new antimicrobial compounds. Soil ecosystems, characterized by intense microbial competition and high metabolic diversity, depicts promising reservoir for antibiotic producing microorganisms. In the current study, soil samples were collected from diverse environments market, agricultural, industrial, and hill regions around Vaniyambadi, Thirupattur District, Tamil Nadu, India, to isolate and characterize bacteria with antimicrobial potential. Serial dilution coupled with nutrient agar cultivation yielded 42 morphologically heterogeneous bacterial isolates. Subsequent preliminary antimicrobial assessment employing the agar well diffusion technique against selected Gram-positive (Staphylococcus aureus, Bacillus subtilis) and Gram-negative (Escherichia coli, Pseudomonas aeruginosa) indicator strains identified five isolates (ICBT-FZAB4, ICBT-FZAB6, ICBT-FZAB9, ICBT-FZMB30, and ICBT-FZIB36) exhibiting considerable antibacterial efficacy. Molecular identification via 16S rRNA gene amplification coupled with sequence-based analysis established that the active isolates showed strong homology with Bacillus tropicus, Bacillus tequilensis, Brevibacillus borstelensis and Bacillus subtilis. Phylogenetic inference validated the taxonomic identities of the isolates, and their sequence data were archived in the GenBank database. The outcomes demonstrate a marked prevalence of Bacillus species among soil-derived antibiotic-producing bacteria, reinforcing the concept that diverse soil niches serve as rich repositories of unexplored antimicrobial resources. The present study advocates systematic bioprospecting of soil microorganisms for the discovery of novel therapeutic agents targeting multidrug-resistant pathogens. Additionally, it provides valuable contributions toward elucidating soil microbial diversity and its translational significance in biotechnological innovation.
Antimicrobial Resistance, Soil Microorganisms, Antibiotic Producing Bacteria, Bacillus Species, 16S rRNA Gene Sequencing
Antibiotics are biologically derived compounds capable of inhibiting or eliminating pathogenic microorganisms and remain essential tools in the treatment of infectious diseases. Most antibiotics originate as secondary metabolites produced by bacteria and fungi, where they function as competitive survival mechanisms in natural environments like soil. Since the discovery of penicillin, soil microorganisms particularly bacteria and Actinomycetes have been identified as major contributors to antibiotic development due to their exceptional metabolic diversity. A significant number of clinically important antibiotics have been isolated from soil-derived genera including Streptomyces and Bacillus.1 The effectiveness of existing antibiotics has been highly compromised by the fast urgencies of antimicrobial resistance resulting from their excessive and improper use. The emergence and global spread of multidrug-resistant microorganisms, notably methicillin-resistant Staphylococcus aureus and β-lactamase expressing Escherichia coli, have substantially compromised the clinical efficacy of conventional antimicrobial therapies. This growing resistance crisis necessitates the exploration of innovative antimicrobial agents possessing novel targets and mechanisms of action. Among natural habitats, soil represents a prolific and largely untapped source for isolating unique antibiotic-producing microorganisms.2 Soil environments sustain remarkably heterogeneous microbial assemblages governed by diverse physicochemical properties. Such ecological complexity promotes competitive and cooperative interactions among microorganisms, thereby inducing the production of structurally diverse secondary metabolites, many of which exhibit significant antimicrobial potential.
Members of the actinomycete group, especially the genus Streptomyces, are acknowledged as leading producers of antibiotics; however, other bacterial taxa such as Bacillus, Pseudomonas, Arthrobacter, and Micrococcus have also been identified as possessing significant antimicrobial activity.3 The recovery of antibiotic-producing bacteria generally entails structured soil collection, followed by serial dilution and inoculation onto selective culture media. Preliminary evaluation of antimicrobial activity is commonly performed using antagonistic techniques, including cross-streak assays and agar-based diffusion methods. Strains demonstrating inhibitory effects are subsequently subjected to comprehensive characterization through morphological observation, biochemical profiling, and molecular analysis. In this context, sequencing of the 16S rRNA gene serves as a reliable approach for precise taxonomic assignment, while also facilitating insights into phylogenetic relationships and the potential for secondary metabolite biosynthesis.4
Overall, the continued exploration of diverse soil habitats represents a promising and effective approach for the discovery of novel antibiotic-producing bacteria with significant potential for pharmaceutical development. Soil environments constitute highly complex microbial reservoirs, where intense ecological competition drives the biosynthesis of diverse bioactive secondary metabolites, including antimicrobial compounds. Investigating these environments can therefore yield new antimicrobial compounds capable of combating a broad range of pathogenic microorganisms. Such research is particularly important in the current era of increasing antimicrobial resistance, as the identification of new antibiotic sources is essential for the development of alternative and effective therapeutic agents. In addition to their medical relevance, these studies also contribute to a deeper understanding of soil microbial diversity, ecological interactions, and their broader biotechnological applications. In this context, the objective of the present study was to isolate antibiotic-producing bacteria from soil samples and to evaluate their antimicrobial potential against selected Gram-positive and Gram-negative test organisms. The antimicrobial efficacy of the obtained isolates was evaluated against representative clinically and environmentally relevant bacterial strains, including Gram-positive species such as Staphylococcus aureus and Bacillus subtilis, and Gram-negative organisms including Escherichia coli and Pseudomonas aeruginosa. The outcomes of the present investigation emphasize the promise of soil-associated microbial communities as a rich reservoir for novel bioactive compounds and reinforce their significance in advancing the discovery and development of next-generation antimicrobial therapeutics.
Sampling strategy
Soil were systematically obtained from multiple locations within and surrounding the Vaniyambadi region, which is located in Thirupathur District, Tamil Nadu, India, in order to obtain samples from different local environments such as market, agricultural, industrial and hill soil samples. The precise sampling sites along with their corresponding georeferenced spatial coordinates are provided in Table 1 and Figure 1. Prior to sampling, the soil surface was cleared of visible debris such as stones, plant residues, and organic matter to minimize external contamination soil sample were aseptically obtained using a sterilized spatula from both the superficial crust and underlying strata, extending to depths of approximately 12 centimeters, in order to capture the heterogeneity of microbial communities distributed across vertical soil profiles. Following collection, samples were immediately transferred into sterile, pre-identified plastic bags under aseptic conditions, tightly sealed to maintain sample integrity, and transported to the laboratory for comprehensive microbiological evaluation. Experimental procedures were performed following previously established protocols as outlined by Bhuyan et al.,5 ensuring consistency and reliability of the sample collection process were followed by sampling methodology.
Preparation of soil sample
From each sampling site, 1 g of soil was aseptically weighed using a sterile analytical balance to ensure accuracy and consistency. A series of six sterile test tubes, each containing 9 mL of distilled water, was prepared and sterilized by autoclaving at 121 °C for 15 min prior to use. The first dilution (10-1) was prepared by transferring the soil sample into the initial tube, followed by vortex mixing for 2-3 min to obtain a homogeneous suspension. Serial dilutions were subsequently carried out by transferring 1 mL of the suspension into successive tubes containing sterile diluent, resulting in stepwise dilutions up to 10-6.
Isolation of antibiotic producing bacteria
For the isolation of bacteria, culture media were prepared under standard laboratory conditions, with nutrient agar serving as the primary medium due to its suitability for the growth of a wide range of non-fastidious bacteria. To prepare the medium, 28 g of nutrient agar powder was accurately weighed and dissolved in 1000 mL of distilled water in a clean beaker. The medium components were dissolved by gentle heating with constant agitation to achieve complete solubilization. Subsequently, the prepared medium was sterilized by autoclaving at 121 °C for 15 min under a pressure of 15 psi to ensure the removal of microbial contaminants. Following sterilization, the medium was cooled to approximately 45-50 °C to minimize condensation and prevent thermal damage to culture plates. Under aseptic conditions, the molten nutrient agar was dispensed into sterile Petri dishes and allowed to solidify at ambient temperature. Thereafter, 1 mL aliquots from the 10-4 dilution were aseptically spread onto the agar surface. The inoculum was uniformly distributed using a sterile L-shaped glass spreader to promote even colony formation. The plates were incubated aerobically at 37 °C for 24 hrs distinct well-isolated colonies were subsequently selected for further characterization, following the methodology described by Kavitha et al.6
Subculturing of bacterial isolates
Following overnight incubation, the culture plates were systematically inspected to assess the emergence and morphology of bacterial colonies. Distinct, well-isolated colonies differing in size, shape, color, and margin were identified. Using a sterile inoculating loop under aseptic conditions, a loopful of each selected colony was carefully picked and transferred onto freshly prepared nutrient agar media plates. Following inoculation, culture plates were incubated at 37 °C for 24 hrs under controlled conditions to promote proper colony development. This sub-culturing process was repeated to ensure the elimination of mixed populations and to obtain pure bacterial cultures. The purified isolates were subsequently preserved and used for further analyses.
Extraction of secondary metabolites
Bacterial isolates were cultured in nutrient broth and incubated at 30-37 °C for 48-72 hrs under shaking conditions (120-150 rpm). Cultures were harvested during stationary phase. Secondary metabolites were extracted using methanol and chloroform as solvents. Intracellular and extracellular fractions were obtained as described previously.7 For intracellular extraction, cultures were centrifuged and the pellet was resuspended in methanol (0.5 mL), homogenized, and centrifuged (10,000 rpm, 10 min); the supernatant was retained. For extracellular extraction, chloroform (0.5 mL) was added to the culture, mixed, and centrifuged to separate phases, and the lower organic layer was collected. The extracts were combined and used for antagonistic assays.
Initial screening of isolates for antibiotic production
Initial screening for antimicrobial metabolite production was carried out through the assessment of inhibitory activity exhibited by the isolated bacterial strains against selected microorganisms. The test panel comprised two Gram-positive bacteria (Staphylococcus aureus and Bacillus subtilis) and two Gram-negative bacteria (Escherichia coli and Pseudomonas aeruginosa), selected to represent common pathogenic groups and to evaluate broad-spectrum inhibitory activity. Fresh overnight cultures of the indicator strains were cultivated in nutrient broth and uniformly inoculated onto sterile Mueller–Hinton agar (MHA) media plates to establish a confluent bacterial lawn. Antibacterial potential was assessed using the agar well diffusion method, wherein 50 µL of crude extract obtained from each isolate was aseptically dispensed into wells created in the agar matrix. The inoculated plates were subsequently incubated at 37 °C for 24 hrs to allow effective diffusion and interaction of bioactive compounds with the test organisms. Post-incubation, zones of growth inhibition surrounding the wells were measured and interpreted as indicators of antimicrobial activity. Isolates demonstrating well-defined and quantifiable inhibition zones were considered promising candidates for antibiotic production and were selected for subsequent analyses.5
Identification of most potential isolate
Genomic DNA extraction
Genomic DNA was extracted from bacterial cultures utilizing the Biobee Spin EXpure Microbial DNA Isolation Kit (Bogar Bio Bee Stores Pvt. Ltd.), following the manufacturer’s recommended protocol. Briefly, newly grown bacterial cultures were collected and processed for cell disruption using the lysis buffer supplied with the kit. This buffer promotes effective breakdown of the bacterial cell envelope, enabling the release of genomic DNA into the surrounding solution. The lysate was then treated to remove cellular debris, proteins, and other contaminants through a series of binding and washing steps. The liberated DNA was preferentially adsorbed onto the silica membrane of the spin column under optimized buffer conditions, whereas residual contaminants were effectively eliminated through successive washing steps. Subsequently, purified genomic DNA of high quality was recovered using the kit-provided elution buffer. The extracted DNA was evaluated for both purity and structural integrity and was deemed appropriate for subsequent molecular procedures, including PCR amplification, sequencing, and other downstream genetic analyses.
PCR protocol
Amplification of bacterial genomic DNA was carried out using PCR, a method based on primer-directed enzymatic replication. Universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TACGCYTACCTTGTTACGA CTT-3′), were selected to target conserved regions flanking the nearly complete 16S rRNA gene. The amplification process involved repeated cycles of thermal denaturation, primer annealing, and elongation mediated by a thermostable DNA polymerase, leading to exponential accumulation of the desired amplicon. The amplified products were subsequently utilized for sequencing and phylogenetic analysis to enable accurate bacterial identification.
Purification of PCR production
Amplified DNA fragments were purified to remove residual primers, excess deoxynucleotide triphosphates (dNTPs), and other reaction constituents using the Montage PCR Clean-Up Kit (Millipore) in accordance with the manufacturer’s protocol. This purification step is critical for eliminating carryover contaminants that can compromise downstream sequencing efficiency and data accuracy. The cleaned amplicons were subsequently subjected to DNA sequencing employing the same primer sets used during amplification. Sequencing was performed using the ABI PRISM® BigDye™ Terminator Cycle Sequencing Kit together with AmpliTaq® DNA polymerase (FS enzyme) (Applied Biosystems). This method utilizes fluorescently tagged dideoxynucleotide terminators incorporated during cyclic extension reactions, enabling precise base-calling across the target region. Following the sequencing reactions, the labeled extension products were resolved and detected using an automated DNA sequencer, yielding high-quality, high-resolution nucleotide sequence data.
Sequencing protocol
Single-pass DNA sequencing was carried out for each purified PCR template using universal 16S rRNA gene primers to obtain nucleotide sequence information. In this approach, sequencing was performed in one direction per reaction, generating sufficient read length and accuracy for reliable bacterial identification. During the sequencing reaction, fluorescently labeled dideoxynucleotide terminators were incorporated into the newly synthesized DNA strands, producing a mixture of fragments of varying lengths. Following the sequencing reaction, the fluorescently labeled extension products were purified to remove unincorporated dye terminators and salts using a standard ethanol precipitation protocol. This purification step ensured reduction of background noise and improved the quality of the sequencing reads. The purified DNA fragments were then air-dried, resuspended in distilled water, and denatured prior to analysis. Finally, the prepared samples were subjected to capillary electrophoresis using an ABI 3730xl DNA Analyzer (Applied Biosystems). The sequencer separated the DNA fragments based on size and detected the fluorescent signals emitted by the labeled terminators, allowing accurate base calling and generation of high-quality 16S rRNA gene sequence data.
Bioinformatics analysis procedure
The obtained 16S rRNA gene sequence was subjected to comparative analysis using the BLAST algorithm against the NCBI nucleotide database to determine its closest phylogenetic affiliations. Sequences exhibiting high similarity scores were retrieved and used as representative homologs for downstream analyses. These selected sequences were aligned using MUSCLE (version 3.7),8 a robust multiple sequence alignment tool known for its accuracy and efficiency in handling large datasets. To enhance the quality and reliability of the alignment, ambiguous regions and poorly aligned positions were systematically removed using Gblocks (version 0.91b). This refinement step minimizes noise arising from highly divergent or misaligned segments, thereby improving the robustness of subsequent phylogenetic inference.9 Phylogenetic relationships were then inferred using the maximum-likelihood approach implemented in PhyML (version 3.0), applying the HKY85 nucleotide substitution model, which accounts for differences in transition and transversion rates as well as base frequencies. Branch support was evaluated using the approximate likelihood ratio test (aLRT), providing a rapid and reliable measure of node confidence. Finally, the resulting phylogenetic tree was visualized, annotated, and graphically rendered using TreeDyn (version 198.3), enabling clear interpretation of evolutionary relationships and taxonomic positioning.10
Soil sampling
Soils were obtained from multiple ecologically distinct sites across the Vaniyambadi area in Thirupathur District, Tamil Nadu, India. The sampling sites were strategically selected to represent different ecological niches, including market soil, agricultural fields, industrial zones, and hill soil regions. These varied environments were chosen to enhance the likelihood of isolating diverse bacterial populations with potential antimicrobial properties. Soil has been widely recognized as a rich reservoir of antibiotic producing microorganisms, particularly bacteria belonging to genera such as Bacillus, Streptomyces, and related taxa. Numerous studies have reported successful isolation of novel and bioactive antibiotic-producing bacteria from soil ecosystems.11-14 Therefore, soil samples from heterogeneous environments were selected in the present study to maximize microbial diversity and improve the probability of recovering potent antagonistic bacterial strains. The precise locations of soil sampling sites, along with their corresponding georeferenced spatial coordinates, are systematically documented in Table 1 and Figure 1, ensuring reproducibility and providing spatial context for the sampling strategy employed in this investigation. Table 2 presents a comprehensive description of soil samples collected from diverse sites in and around Vaniyambadi, located within Thirupathur District, Tamil Nadu. The table systematically summarizes key details for each sampling location, including the site type (such as market, agricultural, industrial, and hill regions), geographic coordinates, and salient environmental characteristics.
Table (1): GPS Coordinates of Soil Sampling Sites in Vaniyambadi, Tamil Nadu, India
| No. | Sample ID | Location Description | Geographic Coordinates | |
|---|---|---|---|---|
| Latitude (°N) | Longitude (°E) | |||
| 1 | AB | Agriculture | 12°40’28.948”N | 78°38’58.017”E |
| 2 | HB | Hill | 12°37’27.4”N | 78°46’21.6”E |
| 3 | MB | Market | 12°40’52”N | 78°37’2”E |
| 4 | IB | Industrial | 12°42’54.542”N | 78°38’57.343”E |
AB-Agriculture Bacteria, HB-Hill Bacteria, MB-Market Bacteria and IB-Industrial Bacteria
Table (2): Description of soil collected from diverse sites and isolates within the Vaniyambadi region of Thirupathur District, Tamil Nadu, India
| No. | Sample Sites | Specific Soil sample area | Soil depth (cm) | Number of isolates and their codes |
|---|---|---|---|---|
| 1 | Agriculture | Lala Eri | 12 | AB 1, AB 2, AB 3, AB 4, AB 5, AB 6, AB 7, AB 8, AB 9, AB 10, AB 11 & AB 12. |
| 2 | Hill | Kambath Hill, Alangayam | 12 | HB 1, HB 2, HB 3, HB 4, HB 5, HB 6, HB 7, HB 8, HB 9, HB 10, HB 11 & HB 12. |
| 3 | Market | Vaniyambadi | 12 | MB 1, MB 2, MB 3, MB 4, MB 5, MB 6, MB 7, MB 8, MB 9 & MB 10. |
| 4 | Industry | Vadacheri | 12 | IB 1, IB 2, IB 3, IB 4, IB 5, IB 6, IB 7 & IB 8. |
| Total | 42 | |||
*AB-Agriculture Bacteria, HB-Hill Bacteria, MB-Market Bacteria and IB-Industrial Bacteria

Figure 1. Soil sampling locations in the Vaniyambadi region of Thirupathur District, Tamil Nadu, India, representing agricultural, hill, market and industrial soil environments for isolating antimicrobial-producing bacteria
Isolation of bacteria and its antagonistic activity
A total of 42 bacterial isolates displaying distinct colony morphologies were obtained from four soil-derived samples. Initial assessment of the isolates was conducted using observable colony features, including dimensions, structural form, pigmentation, edge characteristics, elevation, and surface texture (Figure 2). Selected colonies exhibiting clear spatial separation were repeatedly streaked onto nutrient agar to establish pure and stable cultures suitable for subsequent analyses (Figure 3). Antibacterial activity of the purified isolates was then investigated against selected indicator strains, including Gram-positive (Staphylococcus aureus, Bacillus subtilis) and Gram-negative (Escherichia coli, Pseudomonas aeruginosa) bacteria, following previously reported procedures.11,12,15
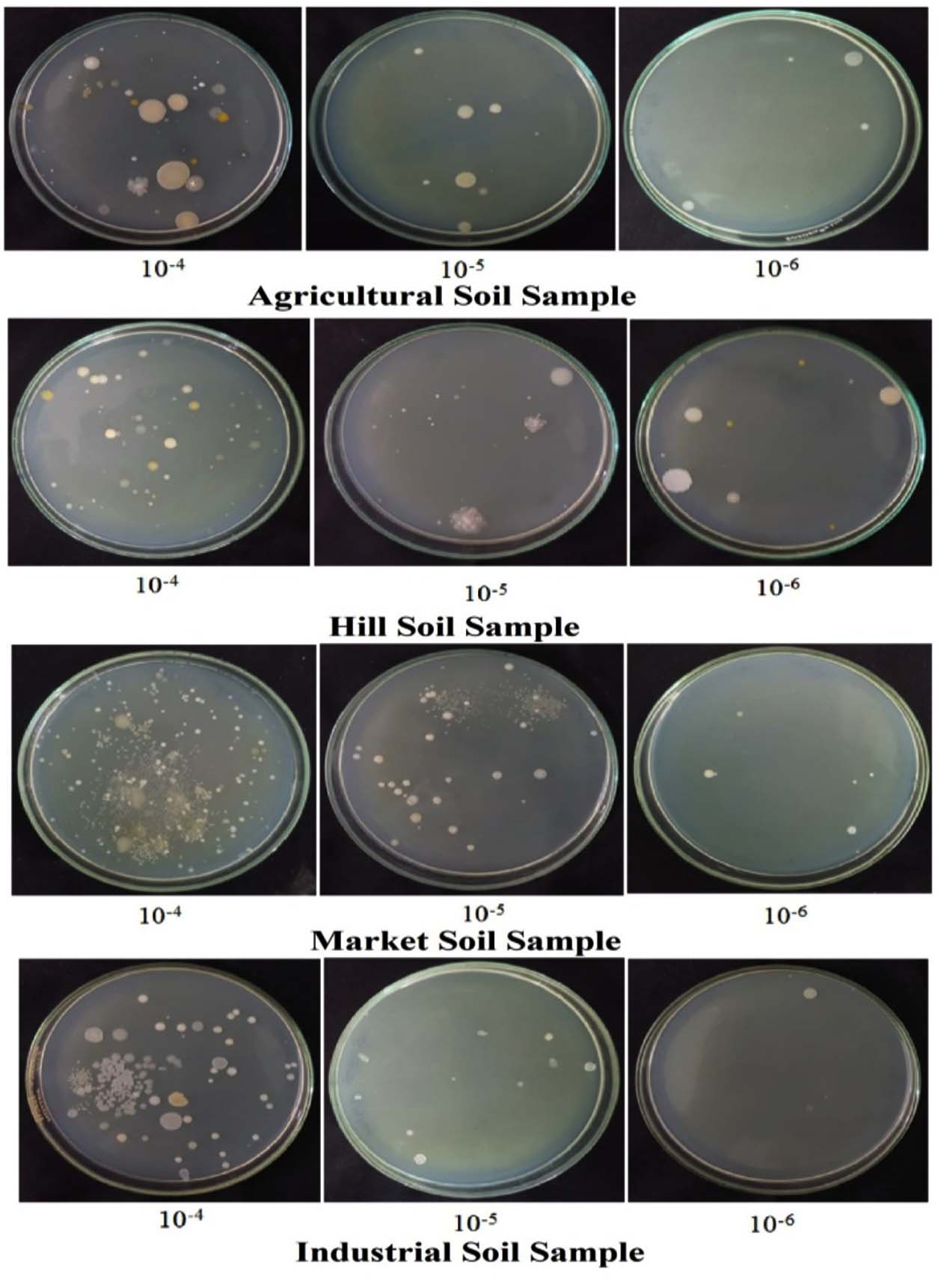
Figure 2. Isolation of morphologically distinct bacterial colonies from four different soil samples using the serial dilution technique

Figure 3. Purification of 42 morphologically distinct bacterial isolates from four soil samples by repeated subculturing
The antagonistic activity of forty-two bacterial isolates was evaluated against two Gram-positive bacteria (Staphylococcus aureus and Bacillus subtilis) and two Gram-negative bacteria (Pseudomonas aeruginosa and Escherichia coli) using the agar well diffusion assay. The results are expressed as mean inhibition zone diameters (mm) ± standard deviation. Among the isolates tested, the majority did not exhibit detectable antagonistic activity against any of the indicator organisms, showing inhibition zones of 0 ± 0 mm. Activity was observed only in a limited number of isolates and was selective toward specific test organisms. Isolates AB 4, AB 6, and AB 9 showed inhibitory activity exclusively against B. subtilis, with inhibition zones of 6.3 ± 0.3 mm, 7.0 ± 0 mm, and 10.3 ± 0.3 mm, respectively. No inhibition was observed against S. aureus, P. aeruginosa, or E. coli for these isolates. Isolate MB6 demonstrated broad antagonistic activity, inhibiting S. aureus (13.0 ± 0 mm), B. subtilis (5.0 ± 0 mm), P. aeruginosa (10.3 ± 0.3 mm), and E. coli (11.6 ± 0.3 mm). Among the industrial soil isolates, IB2 exhibited antagonistic activity against both Gram-negative bacteria, producing inhibition zones of 11.3 ± 0.3 mm against P. aeruginosa and 11.0 ± 0 mm against E. coli. No inhibitory effect was noticed against Gram-positive bacteria for this isolate. Rest of the isolates, including those from hill (HB) soil samples, proven no detectable antagonistic activity against the tested pathogens under the experimental conditions. Overall, the results indicate that only a small proportion of the isolated bacteria possessed measurable antagonistic activity, with selectivity toward either Gram-positive or Gram-negative test organisms, highlighting the targeted antimicrobial potential of certain soil bacterial isolates. These findings are consistent with earlier studies,15,16 which reported maximum inhibitory activity of soil isolates against S. aureus. Similar observations were also reported by Oskay et al.11 The active isolates represented approximately 11.9% of the total bacterial population screened. Overall, the results indicate that only a small proportion of soil derived bacterial isolates exhibit detectable antimicrobial activity under the experimental conditions employed, underscoring the selective nature of antimicrobial potential among soil microorganisms (Table 3 and Figures 4-7).
Table (3): Assessment of the antimicrobial efficacy of bacterial isolates against a panel of selected pathogenic bacterial strains
| No. | Isolates | Inhibition zones (mm) | |||
|---|---|---|---|---|---|
| Gram-positive | Gram-negative | ||||
| S. aureus | B. subtilis | P. aeruginosa | E. coli | ||
| 1 | AB 1 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 2 | AB 2 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 3 | AB 3 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 4 | AB 4 | 0 ± 0 | 6.3 ± 0.3 | 0 ± 0 | 0 ± 0 |
| 5 | AB 5 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 6 | AB 6 | 0 ± 0 | 7. ± 0 | 0 ± 0 | 0 ± 0 |
| 7 | AB 7 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 8 | AB 8 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 9 | AB 9 | 0 ± 0 | 10.3 ± 0.3 | 0 ± 0 | 0 ± 0 |
| 10 | AB 10 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 11 | AB 11 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 12 | AB 12 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 13 | HB 1 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 14 | HB 2 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 15 | HB 3 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 16 | HB 4 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 17 | HB 5 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 18 | HB 6 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 19 | HB 7 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 20 | HB 8 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 21 | HB 9 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 22 | HB 10 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 23 | HB 11 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 24 | HB 12 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 25 | MB 1 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 26 | MB 2 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 27 | MB 3 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 28 | MB 4 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 29 | MB 5 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 30 | MB 6 | 13.0. ± 0 | 5.0 ± 0 | 10.3 ± 0.3 | 11.6 ± 0.3 |
| 31 | MB 7 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 32 | MB 8 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 33 | MB 9 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 34 | MB 10 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 35 | IB 1 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 36 | IB 2 | 0 ± 0 | 0 ± 0 | 11.3 ± 0.3 | 11.0 ± 0 |
| 37 | IB 3 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 38 | IB 4 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 39 | IB 5 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 40 | IB 6 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 41 | IB 7 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 42 | IB 8 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
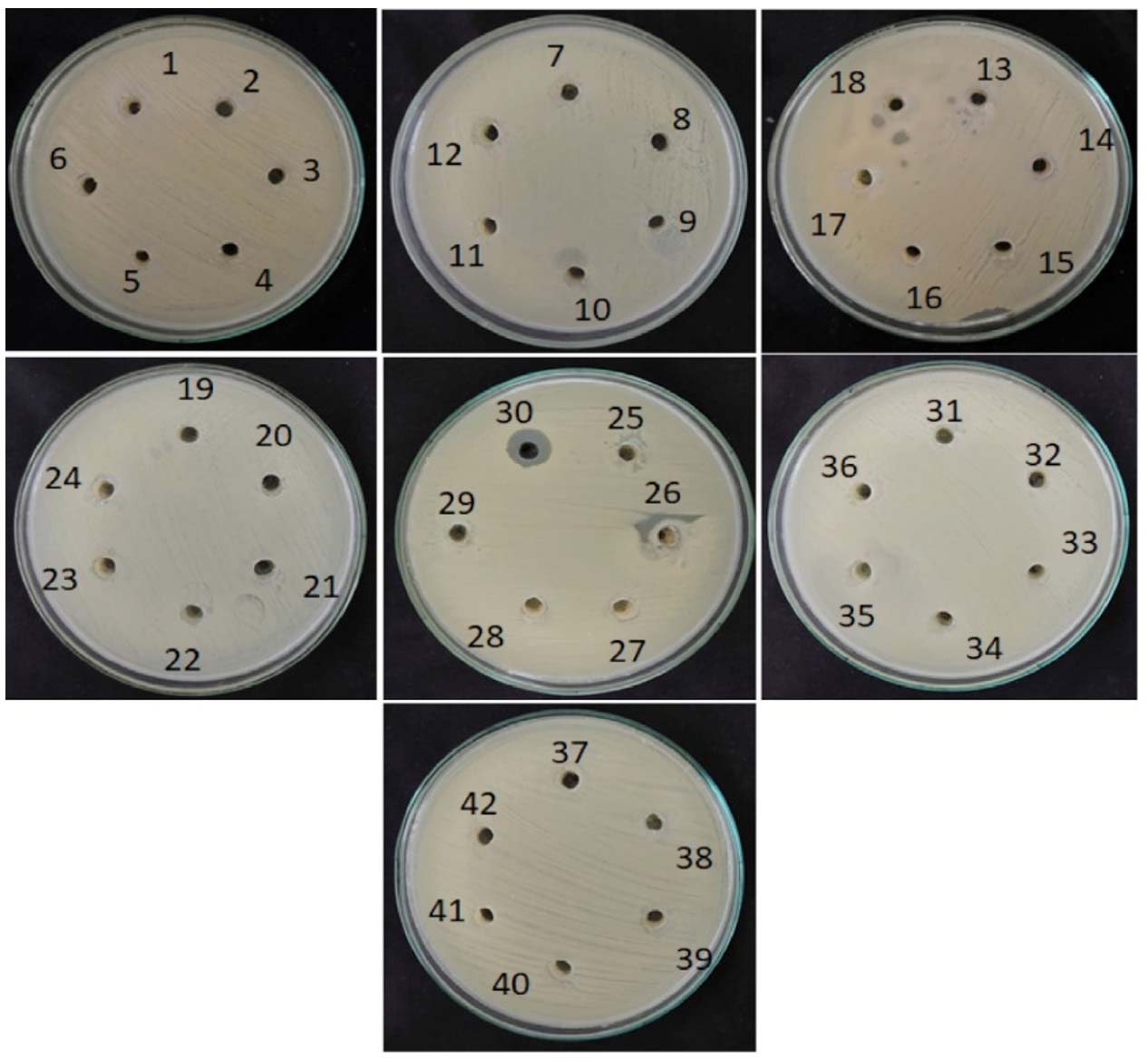
Figure 4. Isolates were screened for antagonism against Staphylococcus aureus

Figure 5. Isolates were screened for antagonism against Bacillus subtilis
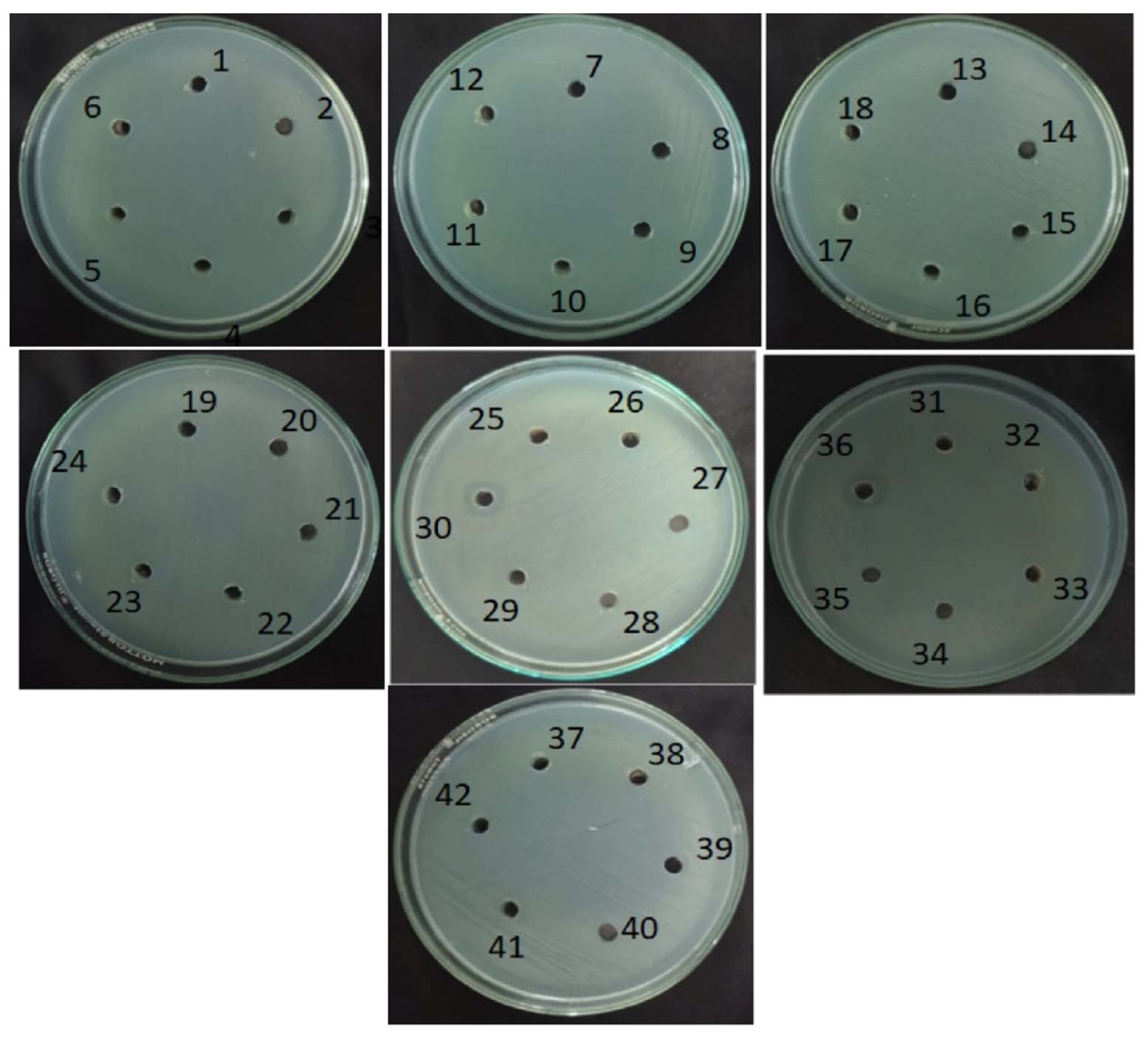
Figure 6. Isolates were screened for antagonism against Pseudomonas aeruginosa
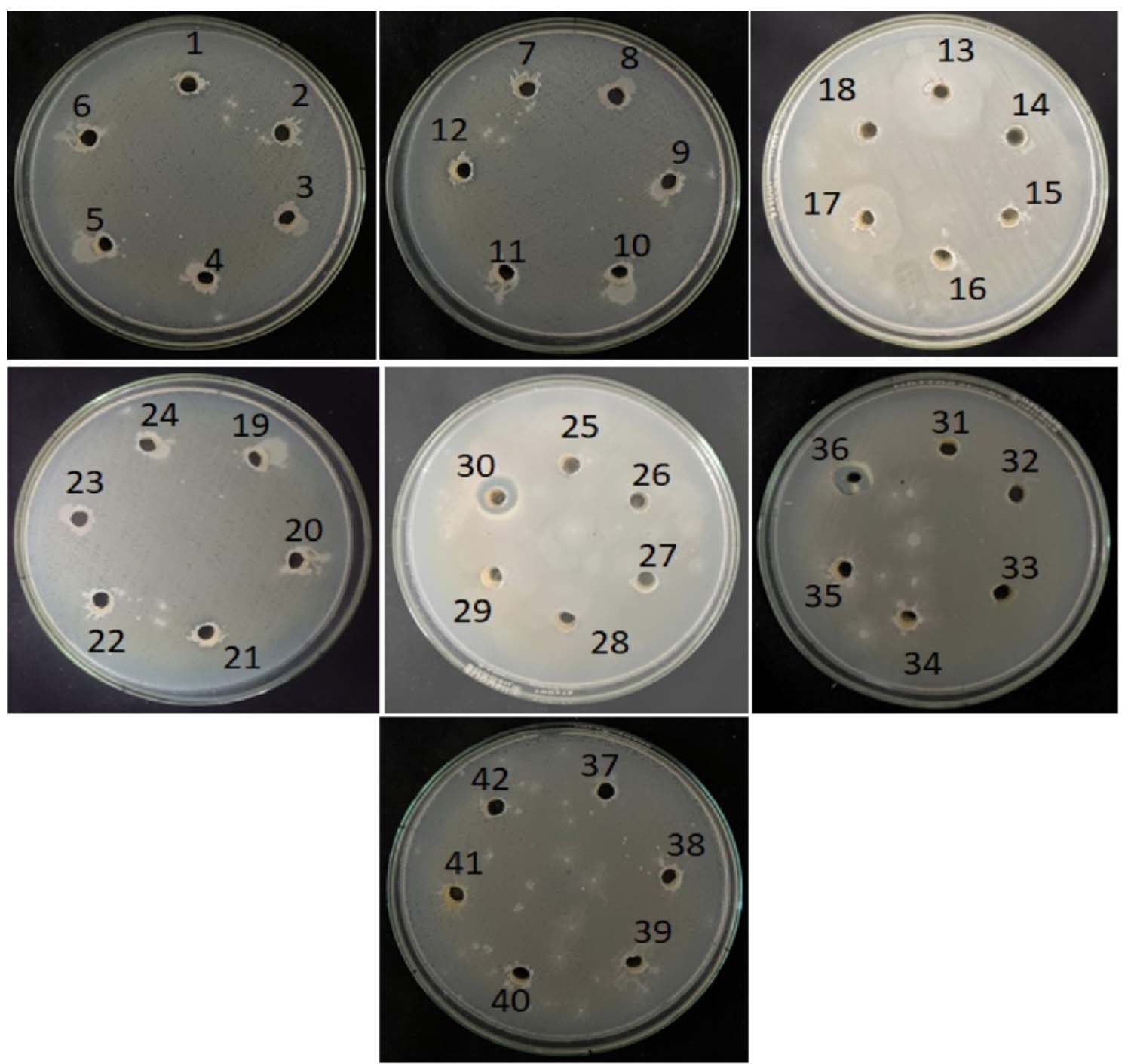
Figure 7. Isolates were screened for antagonism against Escherichia coli
Extraction of genomic DNA from bacterial isolates
Genomic DNA was isolated from bacterial samples following the described protocol to obtain high-quality, high-molecular-weight DNA from five uncharacterized antagonistic bacterial isolates, namely ICBT-FZAB4, ICBT-FZAB6, ICBT-FZAB9, ICBT-FZMB30, and ICBT-FZIB36. The extraction procedure was carried out successfully for all five isolates, yielding intact and purified genomic DNA suitable for downstream molecular analyses. The genomic DNA extraction process involved systematic disruption of the bacterial cell wall, followed by centrifugation to remove cellular debris and insoluble components. Subsequently, nucleic acids were precipitated from the cell lysate, recovered as pelleted DNA, and further purified to eliminate contaminants such as proteins and residual cellular materials. The extracted genomic DNA exhibited satisfactory quality and integrity, indicating its suitability for downstream molecular applications. This DNA preparation was subsequently employed as a template for polymerase chain reaction (PCR) targeting the 16S rRNA gene. Successful amplification of the 16S rRNA region served as a dependable molecular marker for the taxonomic assignment and phylogenetic characterization of the antagonistic bacterial isolates.
Molecular identification of bacteria by 16S rRNA amplification and sequencing
Several previous studies showed that 16S rRNA gene sequence analysis is a more reliable and accurate method for bacterial identification than conventional phenotypic approaches.17-19 In the present study, 16S rRNA gene sequence analysis revealed that strain ICBT-FZAB4 was closely related to Bacillus tropicus, while strain ICBT-FZAB6 showed a close phylogenetic relationship with Bacillus tequilensis. Similarly, strain ICBT-FZAB9 exhibited high sequence similarity with Brevibacillus borstelensis. In addition, strains ICBT-FZMB30 and ICBT-FZIB36 were closely affiliated with Bacillus subtilis. These identifications were confirmed by BLAST analysis through comparison of the obtained sequences with those available in the GenBank database. The partial 16S rRNA gene sequences of all identified strains were deposited in GenBank under the accession numbers PX735844 (ICBT-FZAB4), PX674663 (ICBT-FZAB6), PX674666 (ICBT-FZAB9), PX674673 (ICBT-FZMB30), and PX673725 (ICBT-FZIB36). Phylogenetic analysis based on 16S rRNA gene sequences was performed using MUSCLE, Gblocks, PhyML, and TreeDyn, and the resulting phylogenetic trees illustrating the evolutionary relationships among the isolates and their closely related reference strains Figure 8. These findings are consistent with earlier reports indicating that Bacillus species are among the most predominant bacterial populations in soil.14,20,21 Furthermore, the results strongly support previous studies identifying Bacillus amyloliquefaciens as a potent source of antibiotics.22,23 In addition, Boottanun et al.24 demonstrated that B. amyloliquefaciens is capable of producing a wide range of antimicrobial peptides and secondary metabolites, further corroborating the findings of the present study.

Figure 8. Phylogenetic tree of isolates based on 16S rRNA gene sequences, aligned using MUSCLE, Gblocks, PhyML, and TreeDyn showing their evolutionary relationships with reference strains
A valuable and sustainable source of antibiotic producing bacteria has been demonstrated through this study. Systematic sampling of diverse soil habitats from the Vaniyambadi region resulted in the isolation of multiple bacterial populations, among which a small but significant proportion exhibited antibacterial activity against selected Gram-positive and Gram-negative pathogens. Preliminary screening identified five potent isolates showing inhibitory effects against Staphylococcus aureus, Bacillus subtilis, Escherichia coli, and Pseudomonas aeruginosa. Molecular identification using 16S rRNA gene sequencing revealed that the active isolates were predominantly affiliated with the genus Bacillus, including Bacillus tropicus, Bacillus tequilensis, Bacillus subtilis, and Brevibacillus borstelensis. Phylogenetic analysis confirmed their taxonomic placement and evolutionary relationships with known antibiotic producing strains. In the context of increasing antimicrobial resistance, the present study reinforces the importance of exploring diverse soil environments for the discovery of novel antibiotic producing bacteria and provides a foundation for future studies focused on the characterization and pharmaceutical exploitation of their antimicrobial compounds.
ACKNOWLEDGMENTS
The authors express their sincere gratitude to the Management and the Principal of Islamiah College (Autonomous), Vaniyambadi, for their constant support and for providing the necessary laboratory facilities and institutional infrastructure that enabled the successful completion of this research work.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Rafiq A, Khan SA, Akbar A, et al. Isolation and identification of antibiotic producing microorganisms from soil. Int J Pharm Sci Res. 2018;9(3):1002-1011.
Crossref - Prashanthi R, Shreevatsa GK, Krupalini S, Manoj L. Isolation, characterization, and molecular identification of soil bacteria showing antibacterial activity against human pathogenic bacteria. J Genet Eng Biotechnol. 2021;19(1):120.
Crossref - Elbendary AA, Hessain AM, El-Hariri MD, et al. Isolation of antimicrobial producing Actinobacteria from soil samples. Saudi J Biol Sci. 2017;25(1):44-46.
Crossref - Krishna AR, Aparna Y. Screening and isolation of antibiotic producing microorganisms from soil. Int J Sci Res Biol Sci. 2019;6(Spl Issue 1):46-49.
Crossref - Bhuyan AP, Yadav RNS, Samanta R. Isolation and characterization of antibiotic producing bacteria from soil samples of Dibrugarh District, Assam. Int J Pharm Chem Biol Sci. 2017;7(3):193-201.
- Kavitha R, Dhamodharan N, Dhivya C. Screening, isolation and anti bacterial activity of antibiotic producing bacteria obtained from saprophytic soil samples. Asian J Pharm Clin Res. 2017;10(3):92-96.
Crossref - Pandey A, Srivastava A, Chandra N, Kumar D, Kumar S. Purification and characterization of bioactive secondary metabolites isolated from soil bacteria. Indian J Microbiol Res. 2018;5(2):188-193.
Crossref - Edgar RC. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792-1797.
Crossref - Talavera G, Castresana J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst Biol. 2007;56(4):564-577.
Crossref - Dereeper A, Guignon V, Blanc G, et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008;36(spl 2):W465-W469.
Crossref - Oskay M, Tamer AU, Azeri C. Antibacterial activity of some Actinomycetes isolated from farming soil of Turkey. Afr J Biotechnol. 2004;3(9):441-446.
Crossref - Nike AR, Hassan SA, Ajijolakewu AF. Soil screening for antibiotic-producing microorganisms. Adv Environ Biol. 2013;7(1):7-11.
- Kaur S, Kaur J, Pankaj PP. Isolation and characterization of antibiotic producing microorganisms from soil samples of a certain area of Punjab region of India. Int J Pharm Clin Res. 2014; 6(4):312-315
- Yunus FN, Khalid ZZ, Rashid F, Ashraf A, Iqbal MN, Hussain F. Isolation and screening of antibiotic producing bacteria from soil in Lahore city. PSM Microbiol. 2016;1(1).
- Wadetwar RN, Patil AT. Isolation and characterization of bioactive actinomycetes from soil in and around Nagpur. Int J Pharm Sci Res. 2013;4(4):1428-1433.
Crossref - Saadoun I, Gharaibeh R. The Streptomyces flora of Badia region of Jordan and its potential as a source of antibiotics active against antibiotic-resistant bacteria. J Arid Environ. 2003;53(3):365-371.
Crossref - Clarridge JE III. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin Microbiol Rev. 2004;17(4):840-862.
Crossref - Janda JM, Abbott SL. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J Clin Microbiol. 2007;45(9):2761-2764.
Crossref - Lau SKP, Teng JLL, Ho CC, Woo PCY. Gene amplification and sequencing for bacterial identification. Methods Microbiol. 2015;42:433-464.
Crossref - Singh AP, Singh RB, Mishra S. Studies on isolation and characterization of antibiotic producing microorganisms from industrial waste soil sample. The Open Nutraceuticals Journal. 2012;5(1):169-173.
Crossref - Abbas S, Senthilkumar R, Arjunan S. Isolation and molecular characterization of microorganisms producing novel antibiotics from soil sample. Euro J Exp Bio. 2014;4(5):149-155.
- Arias CA, Murray BE. Antibiotic-resistant bugs in the 21st century: A clinical super-challenge. N Engl J Med. 2009;360(5):439-443.
Crossref - Jeong H, Park SH, Choi SK. Genome sequence of antibiotic-producing Bacillus amyloliquefaciens strain KCTC 13012. Genome Announcements. 2015;3(5):e01121-15.
Crossref - Boottanun P, Potisap C, Hurdle JG, Sermswan RW. Secondary metabolites from Bacillus amyloliquefaciens isolated from soil can kill Burkholderia pseudomallei. AMB Express. 2017;7(1):16.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.