


ISSN: 0973-7510
E-ISSN: 2581-690X
Stimulation of resistance by the activity of defense enzymes viz., peroxidase, polyphenol oxidase, catalase and superoxide dismutase up on treatment with azoxystrobin and chaetoglobosin was studied in this experiment. The potato plants inoculated with Phytophthora infestans showed various levels of peroxidase activity with respect to the treatments. The maximum increase of peroxidase activity (0.954) was noted in combined application of azoxystrobin (Willowood) with chaetoglobosin at 0.2 per cent concentration. The individual application of azoxystrobin (0.908), chaetoglobosin (0.711) and metalaxyl (0.702) was also increased the peroxidase activity to a significant level when compared to inoculated (0.327) and uninoculated (0.259) control. The highest (PPO) polyphenol oxidase (0.898) activity was noticed when combined application of azoxystrobin and chaetoglobosin biomolecule. The same combination also showed the maximum induction of catalase (1.042). Combination of azoxystrobin with metalaxyl was recorded the second highest increase (0.783) of PPO. The individual application of azoxystrobin (0.725) chaetoglobosin (0.719) and metalaxyl (0.653) also increased the PPO activity to a significant level when compared to inoculated (0.301) and uninoculated (0.193) control. In the case of superoxide dismutase, the highest (8.01) activity was recorded in combination of azoxystrobin with metalaxyl. The combination of azoxystrobin with chaetoglobosin also recorded the considerable increase in SOD activity (7.72) which is on par with the best treatment.
Azoxystrobin. Chaetoglobosin, Resistance, Enzymes.
Induced resistance is defined as an enhancement of the plant’s defensive capacity against a broad spectrum of pathogens and pests that is acquired after appropriate stimulation. The resulting elevated resistance due to an inducing agent upon infection by a pathogen is called Induced Systemic Resistance (ISR) or Systemic Acquired Resistance (SAR) (Hammerschmidt and Kuc, 1995). The activity of defense enzymes viz., peroxidase (PO), polyphenol oxidase (PPO), phenylalanine ammonia lyase (PAL), â-1,3 glucanase, chitinase, catalase and defense inducing chemicals (total phenols) was found to be increased in azoxystrobin and P. fluorescens treated grapevine plants (Vinothini et al., 2014). Increased expression of specific isoforms of PO and PPO was observed due to Induced Systemic Resistance (ISR) induction in grapevine (Archana et al., 2011). Peroxidase polymorphism could be used as a biochemical marker related to different levels of field resistance (Lebeda et al., 1999). Peroxidases also participate in synthesis of phenolic compounds and in the building of intermolecular bonds during the organization of the cell wall at the sites of infection by the pathogens (Ahmed et al., 2016). Many studies have shown that PPO is induced in response to mechanical wounding; fungal and bacterial infection; treatment with signaling molecules such as jasmonic acid / methyl jasmonate (MeJA); systemin and salicylic acid (Constabel et al., 2000). Sundravadana (2008) reported that, azoxystrobin had efficiently activated the defense enzymes viz., PO, PPO, and PAL which are increased the lignin content in P. grisea inoculated rice seedlings. Systemic induction of PPO in response to wounding and pathogen infection might provide an additional line of defense to protect the plants against further attack by pathogen and insects (Thipyapong et al., 1995). Application of salicylic acid on bluegrass plants increased the activity of catalase and super oxide dismutase (Mckersie et al., 1996). Babitha (2002) reported the higher SOD activity in resistant pearl millet seedlings than the susceptible seedlings upon inoculation with Sclerospora graminicola. The fungicides such as carbendazim, mancozeb and tebuconazole increased the production of antioxidant enzymes viz., superoxide dismutase, catalase, and peroxidases in mulberry (Narayanan et al., 2016).
Induction of defense related enzymes in potato up on treatment with azoxystrobin and chaetoglobosin was assayed by using the methodologies given below. In all the experiments, metalaxyl was included for comparison purpose.
Sample collection and enzyme extraction
The biomolecules azoxystrobin and chaetoglobosin at 0.2 per cent concentration were compared with 0.2 per cent of metalaxyl for the induction of defense related enzymes. The potato plants sprayed with above treatments were inoculated with P.infestans. The leaf samples were collected at 0, 1, 3, 5, 7, 9 d after inoculation of the pathogen and used for enzyme assay.
One g of potato leaf sample was homogenized with one ml of 0.1M Sodium phosphate buffer (pH 7.0) at 4°C. The homogenate was centrifuged for 20 min at 10000 rpm. The supernatant was used as enzyme extract for assaying of Peroxidase (PO) and Poly Phenol Oxidase (PPO). For Catalase and Super oxide Dismutase (SOD) the sample was extracted in 5 ml of 0.05 M sodium acetate buffer (pH 5.0). The homogenate was centrifuged at 20,000 rpm for 10 min at 4°C and the supernatant was used as enzyme source.
Assay of peroxidase (PO)
Assay of PO activity was carried out as per the procedure described by Hammerschmidt et al. (1995). The reaction mixture consisted of 2.5 ml of the mixture containing 0.25% (v/v) guaiacol in 0.01 M sodium phosphate buffer, pH 6.0 and 0.1 M hydrogen peroxide. Enzyme extract (0.1ml) was added to initiate the reaction, which was followed calorimetrically at 470 nm. Crude enzyme preparations were diluted to give changes in absorbance at 470 nm of 0.1 to 0.2 absorbance units / min. The boiled enzyme was used as blank. Activity was expressed as the increase in absorbance at 420 nm min-1 mg-1 of protein.
Assay of polyphenoloxidase (PPO)
The polyphenoloxidase activity was determined as per the procedure given by Mayer et al., (1965). The reaction mixture consisted of 1.5 ml of 0.1 M sodium phosphate buffer (pH 6.5) and 200 ml of the enzyme extract. To start the reaction, 200 ml of 0.01 M catechol was added and the activity was expressed as change in absorbance at 495 min-1mg-1 of protein.
Assay of catalase (CAT)
CAT activity was assayed spectrophotometrically as described by Chaparro-Giraldo et al. (2000) using 3 ml assay mixture containing 100 mM potassium phosphate buffer (pH 7.5) and 2.5 mM H2O2 prepared immediately before use and 100 µl enzyme extract. The activity was measured by monitoring the degradation of H2O2 using UV-Visible Spectrophotometer (Varian Cary 50) at 240 nm over 1 min, against a plant extract-free blank. The decrease in H2O2 was followed as the decline in optical density at 240 nm, activity was calculated using the extinction coefficient (å240nm = 40 mM-1 cm-1) for H2O2 and expressed in mmol min-1 mg-1 of sample.
Assay of superoxide dismutase (SOD)
The enzyme extract was prepared by homogenizing 1 g of potato leaf tissue in 2 ml of 0.2 M citrate phosphate buffer (pH 6.5) at 4°C. The homogenate was centrifuged at 15,000 g at 4°C for 30 min. The supernatant served as enzyme source and SOD activity (EC 1.15.1.1) was determined as its ability to inhibit the photochemical reduction of NBT. The assay mixture (3 ml) contained 50 mM sodium phosphate buffer (pH 7.8), 13 mM methionine, 75 µM NBT, 2 µM riboflavin. 0.1 mM EDTA and 100 µl of the enzyme extract and the riboflavin was added at the end. Tubes were shaken and placed under a 40-W fluorescent lamp at 25°C. The reaction was initiated and terminated by turning the light on and off respectively. The absorbance at 560 nm was measured against identical non-illuminated in parallel to the sample tubes for blank. Each extract was subtracted from the blank and mathematical difference was then divided by blank and multiplied by 100 to obtain the percentage inhibition of NBT photo-reduction. The SOD activity was expressed in SOD units mg-1 tissue (50% NBT inhibition = 1 unit) (El-Moshaty et al., 1993).
Induced resistance is a “physiological state of enhanced defensive capacity” elicited by specific environmental stimuli, whereby the plant’s innate defenses are potentiated against subsequent biotic challenges. This enhanced state of resistance is effective against broad range of pathogens and parasites (Van Loon, 2000).
Exposing plants to abiotic or biotic stresses lead to improved resistance to subsequent pathogen attack both locally and systemically (Walter et al., 2005). Applying fungicides on plants was also found to induce the resistance against the pathogens. For example, pyraclostrobin (strobilurin class fungicide) enhanced resistance of tobacco plants by activation of pathogenesis related protein (PR 1) against Tobacco Mosaic Virus and Pseudomonas syringae pv tabaci (Herms et al., 2002). The defense enzymes such as superoxide dismutase, catalase and ascorbate peroxidase activities increased after the application of metalaxyl on Solanum nigrum (Alexandra et al.,2013). In the present study, the maximum increase of peroxidase (0.954), polyphenoloxidase (0.898) and catalase activity (1.042) was noted in combined application of azoxystrobin (Willowood) with chaetoglobosin at 0.2 per cent concentration in the potato plants inoculated with P.infestans. In the case of SOD, the combination of azoxystrobin with metalaxyl showed the highest (8.01) activity. Through this study it is evident that, the individual application of fungicides showed lesser increase in defense enzymes as compared to combination treatments. Among the combinations, azoxystrobin with chaetoglobosin showed the maximum induction of defense enzymes on potato. Similar reports have already been made by Anand et al., (2008). They reported that the activity of the defense enzymes such as peroxidase (PO), polyphenol oxidase (PPO), phenyl alanine ammonia lyase (PAL) and Chitinase increased in the azoxystrobin treated cucumber plants. The bioactive compounds, trichotoxin A50 extracted from Trichoderma harzianum PC01 and chaetoglobosin C extracted from Chaetomium globosum have also been reported to elicit resistance or immunity in plants by inducing oxidative burst in plant cells (Nuchadomrong et al., 2004). Enhanced activities of defense related enzymes polyphenol oxidase, peroxidase, phenyl alanine lyase and catalase revealed the role in Induction of systemic resistance in wheat (Aggarwal, 2015). Inducing resistance in plants due to application of biomolecules is an additional advantage through which the disease management cost and quantity of application of fungicidal biomolecules can be reduced to conserve the environment from the contagion.
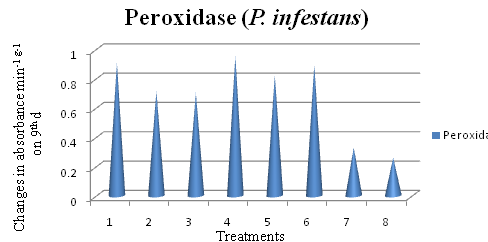
Table (1):
Effect of azoxystrobin, chaetoglobosin and metalaxyl on peroxidase activity in potato plants inoculated with P. infestans.
| Treatment | Absorbance at 420 hm min-1 g-1 at different intervals (d) | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 5 | 7 | 9 | |
| Azoxystrobin 0.2 % | 0.580a | 0.804a | 0.865a | 0.964a | 0.931ab | 0.908a |
| Chaetoglobosin 0.2 % | 0.312c | 0.568d | 0.619b | 0.736b | 0.718c | 0.711c |
| Metalaxyl 0.2% | 0.308c | 0.526d | 0.608b | 0.721b | 0.708c | 0.702c |
| Azoxystrobin 0.2% + Chaetoglobosin 0.2 % | 0.441b | 0.634c | 0.862a | 0.984a | 0.962a | 0.954a |
| Metalaxyl 0.2% + Chaetoglobosin 0.2 % | 0.424b | 0.651c | 0.629b | 0.923a | 0.864b | 0.821b |
| Azoxystrobin 0.2% + Metalaxyl 0.2 % | 0.548a | 0.713b | 0.829a | 0.936a | 0.913ab | 0.892a |
| Inoculated Control | 0.294cd | 0.316e | 0.337c | 0.381c | 0.352d | 0.327d |
| Un inoculated control | 0.265d | 0.283e | 0.302c | 0.297d | 0.264e | 0.259e |
In a column, means followed by same letter are not significantly different at the 5 per cent Level by DMRT
Table (2):
Effect of azoxystrobin, chaetoglobosin and metalaxyl on polyphenol oxidase (PPO) activity in potato plants inoculated with P. infestans
| Treatment | Absorbance at 495hm min-1 g-1 at different intervals (d) | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 5 | 7 | 9 | |
| Azoxystrobin 0.2 % | 0.282 b | 0.586b | 0.642b | 0.751b | 0.746b | 0.725bc |
| Chaetoglobosin 0.2 % | 0.296b | 0.684a | 0.797a | 0.802b | 0.786b | 0.719c |
| Metalaxyl 0.2% | 0.326a | 0.369d | 0.643b | 0.791b | 0.722b | 0.653d |
| Azoxystrobin 0.2% + Chaetoglobosin 0.2 % | 0.294b | 0.642a | 0.802a | 0.902a | 0.906a | 0.898a |
| Metalaxyl 0.2% + Chaetoglobosin 0.2 % | 0.220c | 0.462c | 0.752a | 0.914a | 0.910a | 0.739bc |
| Azoxystrobin 0.2% + Metalaxyl 0.2 % | 0.287b | 0.496c | 0.684b | 0.891a | 0.865a | 0.783b |
| Inoculated Control | 0.242c | 0.294e | 0.300c | 0.492c | 0.427c | 0.301e |
| Un inoculated control | 0.284 b | 0.289e | 0.297c | 0.212d | 0.206d | 0.193f |
In a column, means followed by same letter are not significantly different at the 5 per cent level by DMRT
Table (3):
Effect of azoxystrobin, chaetoglobosin and metalaxyl on catalase activity in potato plants inoculated with P. infestans
| Treatment | changes in absorbance at 240 nm min-1g-1at different intervals (d) | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 5 | 7 | 9 | |
| Azoxystrobin @ 0.2% | 0.562bc | 0.815a | 0.867a | 0.891b | 0.836bc | 0.814b |
| Chaetoglobosin 0.2 % | 0.558 bc | 0.647c | 0.781b | 0.801c | 0.769c | 0.652d |
| Metalaxyl 0.2% | 0.517bcd | 0.574d | 0.698c | 0.799c | 0.782c | 0.676cd |
| Azoxystrobin 0.2% + Chaetoglobosin 0.2 % | 0.498d | 0.693bc | 0.924a | 1.138a | 1.119a | 1.042a |
| Metalaxyl 0.2% + Chaetoglobosin 0.2 % | 0.569b | 0.672bc | 0.791b | 0.883b | 0.865b | 0.721c |
| Azoxystrobin 0.2% + Metalaxyl 0.2 % | 0.621a | 0.718b | 0.787b | 0.896b | 0.841bc | 0.803b |
| Inoculated Control | 0.531bcd | 0.565d | 0.572d | 0.581d | 0.551d | 0.544e |
| Un inoculated control | 0.516cd | 0.532d | 0.543d | 0.568d | 0.534d | 0.521e |
Mean of three replications
In a column, means followed by same letter are not significantly different at the 5 per cent level by DMRT
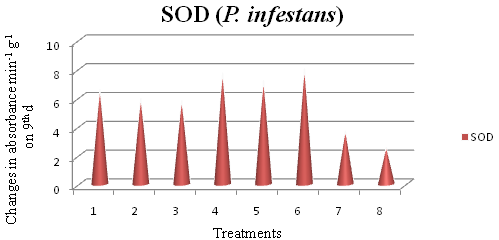
Table (4):
Effect of azoxystrobin, chaetoglobosin and metalaxylon superoxide dismutase (SOD) activity in potato plants inoculated with P. infestans
| Treatment | Unit / min / g of sample at 560 nm in different intervals (d) | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 5 | 7 | 9 | |
| Azoxystrobin 0.2% | 4.03ab | 4.74bc | 6.15bc | 6.89b | 6.73c | 6.58c |
| Chaetoglobosin 0.2 % | 3.85abc | 4.91b | 5.67c | 6.36bc | 6.07d | 5.94d |
| Metalaxyl 0.2% | 3.98abc | 4.37c | 4.90d | 5.87c | 5.96d | 5.81d |
| Azoxystrobin 0.2% + Chaetoglobosin 0.2 % | 3.64c | 5.72a | 6.58ab | 8.24a | 8.06ab | 7.72ab |
| Metalaxyl 0.2% + Chaetoglobosin 0.2 % | 4.16a | 4.84b | 6.03c | 7.98a | 7.54b | 7.19b |
| Azoxystrobin 0.2% + Metalaxyl 0.2% | 3.69bc | 5.02b | 6.69a | 8.30a | 8.13a | 8.01a |
| Inoculated Control | 3.68bc | 3.59d | 3.71e | 3.70d | 3.68e | 3.69e |
| Un inoculated control | 2.84d | 2.95e | 2.77f | 2.63e | 2.61f | 2.55f |
Mean of three replications
In a column, means followed by same letter are not significantly different at the 5 per cent level by DMRT
- Aggarwal, R. Chaetomium globosum: A potential biocontrol agent and its mechanism of action. Indian Phytopath, 2015; 68 (1): 8-24.
- Archana, S., Prabakar, K., Raguchander, T., Manjunath Hubballi., Valarmathi., P and Prakasam, V. Defense response of grapevine to Plasmopara viticola induced by azoxystrobin and Pseudomonas fluorescens. International journal of sustainable agriculture, 2011; 3(1): 30-38.
- El-Moshaty, B., Pike, S.M., Novacky, A.J., Sehgal, O.P. Lipid peroxidation and superoxide production in cowpea (Vigna unguiculata) leaves infected with tobacco ring spot virus or southern bean mosaic virus. Physiol. Mol. Plant Pathol. 1993; 43: 109-119.
- Gonias, E. D., Oosterhuis, D. M. and Bibi, A. C. Physiological response of cotton to the insecticide imidacloprid under high – temperature stress. J. Plant Growth Regul., 2008; 27: 77″82.
- Hammerschmidt, R. and Kuc, J. Induced resistance to disease in plants. Kluwer Academic Publishers, Dordrecht, The Netherlands. 1995; p. 182.
- Herms, S., Seehaus, K., Koehle, H., Conrath, U. A strobilurin fungicide enhances the resistance of tobacco against tobacco mosaic virus and Pseudomonas syringae pv tabaci. Plant Physiol., 2002; 130: 120-27.
- Lebeda, A. Pseudoperonospora cubensis on Cucumis spp. and Cucurbita spp. – resistance breeding aspects. Acta Hort., 1999; 492: 363-370.
- Mayer, A.M., Harel, E. and Shaul, R.B. Assay of catechol oxidase a critical comparison of methods. Phytochemistry, 1965; 5: 783-789.
- McKersie, B.D., Bowley, S.R., Harjanto, E., Leprice, O. Water-deficit tolerance and field performance of transgenic alfalfa over expressing superoxide dismutase. Plant Physiology, 1996; 111: 1321–1326.
- Mosaad F. Ahmed, Hanafy A. Hamza, Ibrahiem A. Ibrahim, A.A. Nower and Mostafa Alansary. Biochemical Detection of Phenolic Compounds and Peroxidase in New Locally Improved Tomato Lines by using HPLC. Int.J.Curr.Microbiol.App.Sci. 2016; 5(5): 275-283.
- Narayanan, P., Parthasarath, S., Rajalakshmi, J., Arunkumar, K and Vanitha. Systemic elicitation of defense related enzymes suppressing Fusarium wilt in mulberry. African Journal of Microbiology Research. 2016; 10(22): 813-819
- Nuchadomrong, S., Buthongsri, P., KanoKmedhalkul, S. and Soytong, K. Trichotoxin A50: Induction of an oxidative defence mechanism in tomato and its celular site of action. In proceedings 30th congress on Science and Technology of Thailand, 19-21 Oct. Impact Muang Thong Thani, Bangkok, Thailand 2004.
- Repka,V and Slovonosa. Structure, expression and synthesis induction of proteins related to pathogenic processes in plant cell. Biol. Listy., 1994; 58: 81-100.
- Sundravadana,S., Alice, D., Kuttalam, S. and Samiyappan, R. Azoxystrobin induces lignification-related enzymes and phenolics in rice (Oryza sativa L.) against blast pathogen (Pyricularia grisea). J. Plant Interactions, 2007; 2: 219 – 224.
- Thipyapong, P., Hunt, M.D., Steffens, J.C. Antisense down regulation of polyphenol oxidase results in enhanced disease susceptibility. Planta, 1995; 220: 105–117
- Van Loon, L.C. Systemic induced resistance. In: Mechanisms of resistance to plant diseases (Slusarenko AJ, Fraser RSS & van Loon LC eds.) Kluwer: Dordrechet, 2000; pp 521-574
- Vinothini.K, Ahila Devi.P., Latha.P., and Prakasam,V. Defense enzyme activation and enhancement of quality parameters by azoxystrobin 8.3 % W/W and mancozeb 64.75 % w/w and biocontrol agent in grapevine against P. viticola, 2014; 3(3): 2319-1473.
- Walter, D., Walsh, D., Newton, A. and. Lyon, G. Induced resistance for plant disease control: maximizing the efficacy of resistance elicitors. Phytopathology, 2005; 95: 1368–1373.
© The Author(s) 2016. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.