


ISSN: 0973-7510
E-ISSN: 2581-690X
The global challenge of antibiotic resistance among gastrointestinal pathogens is increasing daily. Gastroenteritis is a major health challenge in developing countries and is ranked 8th among the top 10 causes of death in Namibia. While medicinal plants used in Namibia could be the source of medicine for gastroenteritis, there is a gap in knowledge regarding the efficacy and safety of most medicinal plants used in Namibia. The study aimed to evaluate the antibacterial and cytotoxicity of medicinal plants used in Namibia to manage gastroenteritis. Aqueous and methanol extracts were prepared from Lantana camara, Grewia tenax, Corchorus tridens, Albizia anthelmintic, and Terminalia sericea. Agar disc diffusion and resazurin-based microtiter dilution assay were used to determine the antibacterial activity. Antibiogram assay was used to determine the susceptibility of gastrointestinal pathogens to 11 clinically used antibiotics. Normal mouse embryo fibroblast cells using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay were used for the cytotoxicity test. A 70% multidrug resistance to ampicillin, cephalosporin, sulphamethoxazole, erythromycin, penicillin, vancomycin, and ampicillin was observed among clinical isolates of Salmonella and Escherichia coli. Meanwhile, L. camara methanol seed extracts showed potent antibacterial activity against clinical strains of E. coli and Salmonella spp. (20.67 ± 1.53 and 21.67 ± 0.58 mm, respectively), with MICs between 31.25 and 62.5 µg/mL. The IC50 of all extracts was ≥65.85.00 ± 0.58 µg/mL. The study showed that methanol extracts of L. camara, C. tridens, and T. sericea have potent antibacterial activity against multidrug-resistant clinical Salmonella and E. coli. Moreover, methanol extracts did not show significant toxicity to NIH/3T3 cells.
Multidrug Resistance, Antibacterial, Cytotoxicity, Gastroenteritis, Salmonella, Escherichia coli
Diarrheal diseases are a major global health concern in pediatrics accounting for over 1.3 million mortality cases in children under the age of 5 annually.1-3 Rotavirus is the most common cause of acute diarrhea among children; however, important bacteria such as Salmonella, Shigella, Yersinia, Campylobacter, and Escherichia coli are responsible for a significant number of deaths among children, especially in immunocompromised paediatrics in Sub-Africa.4-7
Bacterial diarrhea occurs mainly via the ingestion of contaminated food or water. Salmonella invades the mucosal membrane of the distal ileum and proximal colon. Salmonella can survive within a hostile gastric environment with low pH and avoid lysis by bile salts in the upper small intestine.8 It specifically colonizes the M-cells that are located at the top of the lymphoid follicles of Peyer’s patches.9 If not eliminated, the microbes then penetrate the sub-mucosa and enter the bloodstream via the lymphatics. Transient bacteremia follows and the Salmonella spread to the endothelium of the liver, spleen, bone marrow, gall bladder, and kidneys. The organisms will then re-enter the intestine from the gallbladder resulting in the inflammation of the Peyers’ patches and ulceration.10 E. coli invade the mucosal membrane of the small intestine where; it binds to the host cell membranes. It further releases the toxin that gets endocytosed and translocated through the cell in a process involving trans-Golgi vesicular transport leading to systemic complications such as hemolytic uremic syndrome, which can lead to kidney failure and death.11,12
In many cases, oral rehydration is recommended to manage acute diarrhea in children instead of antibiotic therapy.13 However, in immune-compromised children with persistent diarrhea treatment using probiotics and antibacterial agents are highly recommended.14,15 The use of antibiotics and antidiarrheal agents are fraught with complications due to undesired side effects of the treatments that include vomiting, nausea, severe diarrhea, and loss of appetite largely due to large dosages administered. All those challenges limit the effectiveness of existing antibiotics and antibacterial agents and have resulted in increased mortality and morbidity cases due to acute diarrhea, especially in children and immunocompromised patients.16 Moreover, antibiotic resistance has become a global health crisis estimated to be responsible for 7,00,000 deaths annually worldwide. Higher rates of antibiotic resistance to first-line antibiotics such as amoxicillin, erythromycin, gentamycin, and ampicillin in gastrointestinal pathogens are reported daily. Hence, the World Health Assembly adopted a Global Action Plan on antimicrobial resistance in 2015, with national governments in more than 120 countries acting on the challenges presented by antimicrobial resistance.17,18 Limitations presented by existing antibiotics and antibacterial agents call for alternative medicine with fewer complications and significant therapeutic effects.
In recent years, there have been an increasing number of studies to discover new bioactive compounds of plant origin with the hope of controlling antibiotic-resistant bacteria.19 This is because medicinal plants have been used in all age groups for a long period, with 80% of populations in developing countries relying on medicinal plants as an alternative treatment option for diarrhea and other ailments.20,21 Medicinal plants such as Terminalia sericea, Lantana camara, Grewia tenax, and Albizia anthelmintica are used in Iikokola village to treat different gastrointestinal conditions such as diarrhea, stomach pain, vomiting, bloating, and indigestion.22 Moreover, these plants are also reported to contain phytochemical compounds such as alkaloids, saponins, anthraquinones, terpenoids, tannins, and flavonoids; and their total phenolic and flavonoids contents are reported by Shatri and Mumbengegwi.22 However, despite the long history of use, there is limited scientific data to validate the efficacy and safety of medicinal plants especially those used of plants to manage diarrhea. The escalating global cases of antibiotic resistance especially among gastrointestinal bacteria require alternative treatment options for diarrhea from bacteria such as Salmonella, Shigella, Campylobacter, and verotoxigenic Escherichia coli species, especially in African countries such as Nigeria, Namibia, Ethiopia, Chad, Mali, and the Democratic Republic of Congo.6,23 Hence, the objective of this study was to determine the antibacterial activity of Terminalia sericea (BRL 40), Lantana camara (BRL 99), Grewia tenax (BRL 33), Corchorus. tridens (BRL 30), and Albizia anthelmintica (BRL 34) and the antibiogram activity of selected antibiotics against clinical isolates of Salmonella spp. and verotoxigenic Escherichia coli 157:H7 and reference strains of Salmonella Typhimurium ACCT 13311 and Escherichia coli ATCC 25922 as well as to determine the cytotoxicity of the selected extracts to the of NiH/3T3 normal mouse embryo fibroblast cells. This study contributes to a better understanding of the potential therapeutic applications of the plants in the treatment of bacterial gastroenteritis.
Plant material
Terminalia sericea (twigs and seeds), Corchorus tridens (whole), Lantana camara, (twigs and seeds), Grewia tenax (roots and twigs), and Albizia anthelmintica (twigs and roots) plant materials were selected for this study based on their frequency of ethnomedicinal use by people in Iikokola village, Omusati region in Namibia to manage different gastrointestinal conditions including gastroenteritis. The identification of the plants was conducted by the National Herbarium of Namibia (permit number: 2221/2017). Ethical clearance to conduct this study was obtained from the University of Namibia’s ethical committee (SOS-0039).
Preparation of crude extract
T. sericea (twigs and seeds), C. tridens (whole), L. camara (twigs and seeds), G. tenax (roots and twigs), and A. anthelmintica (twigs) plant materials were collected, washed with distilled water, and air-dried in the shade for 4 weeks. The dried plant materials were ground to a fine powder using an industrial grinder. For the preparation of organic extracts, 20,000 µg/mL of the plant materials were added to 200 mL of methanol and mixed in a shaker for 48 hours at room temperature for maceration. Aqueous extracts were prepared by adding 20,000 µg/mL of the plant materials to 200 mL of distilled water and keeping the mixture in a water bath set at 60°C for 4 hours. All extracts were filtered using Whatman no 1 filter papers and the filtrates were concentrated by rotary evaporation at 40°C and then freeze-dried to form a powder. The powdered extracts were stored at -20°C. The percentage yield was calculated using the formula:
% Yield = (Mass of plant extract/Mass of plant material) x 100
Clinical isolates and reference strain
Four bacterial isolates were used in this study. Clinical Salmonella spp. and E. coli 157:H7 VTEC were isolated from stool samples as described by Iikasha et al.6 while the reference strains of Salmonella Typhi ATCC 13311 and Escherichia coli ATCC 25922 were used. The bacterial isolates were used for the comparative evaluation of the antibacterial activity of organic and aqueous crude plant extract as well as for the antigram assay.
Preparation of bacterial culture
Using aseptic techniques, test tubes of Muller Hinton broth were inoculated with the reference and clinical bacterial isolates. The inoculated broths were incubated for 18 hrs at 37°C. After incubation, the turbidity of the bacteria cultures was adjusted to match the 0.5 McFarland standard by serially diluting the cultures with Muller Hinton broth. The viability graph was used to calculate the actual number of colony-forming units. The dilution factor was calculated, and the dilution was performed to obtain a final concentration of 5 x 106 CFU/mL.
Antibiotic susceptibility testing of clinical and reference strains of Salmonella and E. coli
Antibiotic susceptibility test of the clinical isolates and reference strains; and the antibacterial activity of each plant extract were determined using the modi ed Kirby-Bauer disc difusion method in agreement with the guidelines of the Clinical and Laboratory Standards Institute.15 The antibiotics used for the antigram assay included: cipro oxacin (5 µg/mL), erythromycin (15 µg/mL), tetracycline (30 µg/mL), vancomycin (30 µg/mL), cephalothin (30 µg/mL), ampicillin (10 µg/mL), sulphathiazole (25 µg/mL), gentamicin (10 µg/mL), penicillin G (10 µg/mL), and amoxicillin (10 µg/mL). The plant extract (1000 µg) was re-dissolved in 2 mL of methanol and vortexed. Sterile Whatman filter paper discs were placed in falcon tubes containing the plant extracts of the concentration of 500 µg/mL and macerated for 24 hours at 4°C. Petri dishes containing 8 mL of solidified Mueller-Hilton agar were inoculated with 10 µl of the inoculums containing clinical isolates and reference strains of Salmonella and E. coli. The discs were allowed to warm to ambient temperature before application to prevent condensation. The impregnated discs and antibiotic discs were placed on Mueller-Hilton agar plates. The plates were allowed to stand for 30 minutes at 4°C to allow plant extracts and antibiotics to diffuse into the media and then incubated at 37°C for 24 hours. Distilled water and 99% methanol were used as negative controls.
Validation of the resazurin-based microtiter dilution assay
To establish the accuracy of this method, the performance of 3 standard antibiotics namely tetracycline, which is bacteriostatic as well as gentamycin, and ciprofloxacin, which are bactericidal against reference strains of Salmonella Typhi ATCC 13311 and Escherichia coli ATCC 25922 were determined. The minimum inhibitory concentration (MIC) values obtained were compared with those published by the National Committee for Clinical Laboratory Standards,15 and the Global Salm-Surv.23 The MIC was determined based on the color change observed. With resazurin-based MIC assay, active living cells cause a reduction of resazurin (purple-blue) to resorufin (pink-colorless).
Resazurin-based Microtiter Dilution Assay to determine the minimum inhibitory concentrations of the plant extracts
Resazurin dye (600 mg) was dissolved in 80 mL of sterile distilled water. A vortex mixer was used to homogenize from the Resazurin dye solution. Under aseptic conditions, 96 well microtiter plates were used for the Resazurin-based Microtiter Dilution Assay. For each plate, the first row of the microtiter plate was filled with 100 µl of organic or aqueous plant extract stock solution of 500 µg/mL. All the wells of microtiter plates were filled with 100 µl of nutrient broth. Two-fold serial dilution (throughout the column) was achieved by starting to transfer 250 µl test material from the first row to the subsequent wells in the next row of the same column so that each well has 250 µl of plant extracts in serially descending concentrations. 10 µl of resazurin solution as an indicator was added to each well. Finally, a volume of 10 µl was taken from bacterial suspensions of clinical Salmonella spp. and E. coli 157:H7 VTEC; and laboratory isolates of Salmonella Typhi ATCC 13311 and Escherichia coli ACCT 25922 (5×106 CFU/mL) was added to each well respectively. To avoid the dehydration of bacterial culture, each plate was covered loosely with the plate lid. Each microtiter plate had a set of 3 controls: (a) a column with Sulphathiazole (25) as a positive control, (b) a column with all solutions except for the test extract, and (c) a column with all solutions except bacterial solution replaced by 10 µl of nutrient broth. The plates were incubated at 37°C for 24 hours. The color change in the well was then observed visually. Resazurin is an oxidation-reduction indicator used for the evaluation of cell growth, particularly in various cytotoxicity assays. The dye remains purple in the absence of bacteria growth (non-fluorescent); however, when there is bacteria growth the dye becomes pink and fluorescent as it is reduced to resorufin by oxidation-reduction within viable cells. Resorufin is further reduced to hydro-resorufin (uncolored). Any color change observed from purple to pink or colorless was taken as positive. The lowest concentration of plant leaf extract at which no color change was recorded was the MIC value. All the experiments were performed in triplicates. The average values were calculated for the MIC of the test material.24
Cytotoxicity assay
Culturing of normal mouse embryo fibroblast cells (NiH/3T3) cells
The 3T3 cells used in this study were initially cultured and maintained as described by Pour et al.25 with minor modifications. Frozen cells were removed from -80°C, thawed at 37°C, and grown for 1 week at 90% Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS) and 1% streptomycin at 37°C in 95% humidity with 5% CO2. The cultured cells were viewed under the inverted microscope and the confluence of the cell monolayer was monitored until it was about 80-90%. Under sterile conditions in the biosafety cabinet, the flasks with over 90% confluence were selected for further analysis. The culture media was aspirated from the cells and washed with warm sterile phosphate buffer saline (PBS). After aspirating out PBS, 10% of the enzymatic digestion trypsin in PBS was added to each flask, and the flasks were incubated in the CO2 incubator at 37°C for 3 minutes to allow the cells to detach from the flask surface. Skipper was used to gently stripe off the cells from the flask surface. Briefly, fresh syringe-filtered media was added to each flask with cells and swirled gently. The cell suspensions were transferred to a falcon tube and centrifuged for 7 minutes at 1000 rpm. After discarding the media, the pellet was resuspended in fresh media and mixed gently. Cells were sub-cultured once a week and used before the 10th passage.
Cell count and seeding density
Briefly, 200 µl of trypan blue stock solution was transferred to a 15 mL falcon tube and 9.8 mL of 1% acetic acid (working solution). Exactly 100 µl of the cell suspension was transferred into a 1.5 mL falcon tube and add 100 µl of trypan blue.26 Briefly, 10 µl of the mixture was loaded onto the haemocytometers and viewed at the lower magnification and the cells were counted inside each of the 4 large corner squares (the total number of cells counted should be ≥50). The cell density was calculated using the equation below:
Total cells/ml = Total cell count x (Dilution factor/Number of squares) x 10000
Using the obtained total cell count, and the desired volume the seeding density was calculated. The cells were seeded at 2.85 x 105. Briefly, 100 µl of the 2.85 x 105 cell suspension was seeded onto all the 96-well microtiter tissue culture plates except for the wells used for blank. The 96 well plates were incubated at 37°C for 24 hours in the CO2 incubator for 48 hours to allow 80-90% confluence in the 96-well microtiter tissue culture plates.
Extracts preparation
For each extract, a stock solution (SS) of 10 mg/mL concentration was prepared in 1:9 of dimethyl sulphoxide (DMSO) and media, respectively and vortexed for 5 minutes at room temperature. Exactly 1 mg/mL concentration was prepared by adding 100 µl of the SS to 900 µl of the fresh media. A desired initial working concentration of 100 µg/mL was prepared by adding 20 µl of the 1 mg/mL concentration to 980 µl of the fresh media. The solution was then syringe filtered. Serial ten-fold dilutions were made from SS to give working concentrations of 100-3.125 µg/mL. Confluent monolayers of the cells were grown in 96 well-microtiter plates for 48 hours. Cells were incubated with various concentrations of the test extracts in triplicate at 37°C in a CO2 incubator for 48 hours. The negative control was performed using a growth medium alone instead of plant extract, while Triton-x was used as the positive control.
Determination of cell viability by MTT assay
Cell viability was examined by the ability of the cells to cleave the tetrazolium salt MTT [3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyl tetrazolium bromide] (Sigma, Chem, St. Louis, MO), by the mitochondrial enzyme succinate dehydrogenase following the procedure as described van Tonder et al. with minor modifications. After 48 hours, supernatants were removed from the wells and 30 µl of 5 mg/mL MTT solution was immediately put into each well, followed by an incubation period of 2 hours at room temperature in the dark. The plates were incubated for 2 hours at room temperature. Afterward, the media in the plates was carefully discarded. For dissolving formazan blue crystals, 100 µl of dimethyl sulfoxide (DMSO) was added to each well and incubated at room temperature for 15 minutes in the dark. The plates were placed in the spectrophotometer and shaken for 5 minutes. The optical density was determined at 570 nm using a multi-well spectrophotometer. The percentage of cell growth inhibition was calculated based on a comparison with the untreated cells.
The percentage of cell viability was calculated as per the following formula:
Cell viability (%) = [(Absorbance of sample – Absorbance of blank)/(Absorbance of untreated cells – Absorbance of Blank)] × 100
The 50% Inhibitory concentration (IC50) was defined as the extract concentration required for the reduction of cell viability by half. The IC50 value and the standard error mean (SEM) were calculated using a non-linear regression curve contained in Excel (Concentrations 100-3.125 µg/mL).
Statistical analysis
All experiments were done in triplicates and statistical analysis was performed employing Graph Pad Prisms software version 7.0. Comparison between groups was done using Two-way ANOVA, followed by the Bonferroni post-tests test in which the P < 0.05 was considered significant.
Effect of extracting solvent on the percentage yields of crude extracts
Organic and aqueous plant extracts were prepared from selected four plants using methanol and distilled water, respectively. The percent extract yield of organic and aqueous plant extracts varies between 1.0 to 13.5% (Table 1). In most cases, the amount of residue extracted with methanol was higher than that of distilled water. However, the percentage yields for L. camara organic and aqueous seeds and twigs extracts and organic and aqueous twigs extracts were equal, respectively. Seed extracts yielded higher than twigs and root extracts (Table 1). Studies have linked the percentage yield of the crude extract to its efficacy and bioprospecting properties.27
Table (1):
Percent (%) yield of various organic and aqueous plant extracts
| Sample number | Plant name | Part used | The percent yield of plant extract | |
|---|---|---|---|---|
| Aqueous extracts | Organic extracts | |||
| BRL 33 | G. tenax | Roots Twigs |
1.0 3.0 |
6.5 3.0 |
| BRL 34 | A. anthelmintica | Seeds Twigs |
1.5 9.5 |
8.0 1.5 |
| BRL 40 | T. sericea | Seeds Twigs |
10.0 4.0 |
12.0 8.5 |
| BRL 99 | L. camara | Seeds Twigs |
13.5 12.5 |
13.5 12.5 |
| BRL56 | C. tridens | Whole | 1.0 | 8.0 |
Antibacterial activity of aqueous and organic extracts
The antibacterial activity of organic and aqueous extracts of G. tenax, A. anthelmintica, T. sericea, L. camara, and C. tridens were assessed against clinical and reference strains of E. coli and Salmonella. Organic extracts showed higher and broad-spectrum antibacterial activity in comparison to aqueous extracts (Table 2 and 3). Higher antibacterial activity was observed in organic seed extracts of L. camara against both clinical and reference strains of E. coli (20.67 ± 1.53 mm and 22.00 ± 1.00 mm) and Salmonella species (21.67 ± 0.58 mm and 20.33 ± 0.58 mm), respectively (Table 3). The L. camara (seeds) and C. tridens were the only aqueous extracts that showed antibacterial activity against clinical isolates of Salmonella spp. (11.33 ± 0.58 mm) and E. coli 157:H7 VTEC (11.67 ± 0.58 mm), respectively (Table 2). T. sericea twigs and seeds, L. camara seeds, and C. tridens also showed antibacterial activity against reference strains of E. coli, with higher activity depicted by L. camara seeds (12.33 ± 0.58 mm) and C. tridens whole plant (12.00 ± 1.00 mm). Among other, aqueous extracts, T. sericea twigs, and L. camara seeds, were the only aqueous extracts that showed antibacterial activity against the reference strain of S. Typhimurium (Table 2).
Table (2):
Antibacterial activity of aqueous extracts against clinical isolates and reference strains of Salmonella spp. and E. coli
Plant extracts |
Average inhibition of clinical E. coli (mm) |
Average inhibition of reference strain of E. coli (mm) |
Average inhibition of clinical Salmonella isolates (mm) |
Average inhibition of reference strain of Salmonella isolates (mm) |
|---|---|---|---|---|
G. tenax twigs |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
G. tenax roots |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
A. anthelmintica twigs |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
A. anthelmintica roots |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
T. sericea seeds |
0 ± 0.0 |
8.3 ± 0.58 |
0 ± 0.0 |
8 ± 1.0 |
T. sericea twigs |
0 ± 0.0 |
8 ± 0.0 |
0 ± 0.0 |
7.3 ± 0.58 |
L. camara seeds |
9 ± 0.0 |
12.3 ± 0.58 |
11 ± 0.58 |
8.7 ± 0.58 |
L. camara twigs |
0 ± 0.0 |
10.7 ± 1.2 |
0 ± 0.0 |
0 ± 0.0 |
C. tridens whole |
11.7 ± 0.58 |
12.0 ± 1.0 |
0 ± 0.0 |
0 ± 0.0 |
Methanol |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
Distilled water |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
0 ± 0.0 |
Table (3):
Antibacterial activity of organic extracts against clinical isolates and reference strains of Salmonella spp. and E. coli
Plant extracts |
Average inhibition of clinical E. coli (mm) |
Average inhibition of reference strain of E. coli (mm) |
Average inhibition of clinical Salmonella isolates (mm) |
Average inhibition of reference strain of Salmonella isolates (mm) |
|---|---|---|---|---|
G. tenax twigs |
11.7 ± 1.58* |
10.7 ± 0.6* |
9.3 ± 0.58* |
7 ± 0.00* |
G. tenax roots |
0 ± 0.00 |
0 ± 0.00 |
8.7 ± 0.58* |
0 ± 0.00 |
A. anthelmintica twigs |
8.3 ± 0.58* |
0 ± 0.00 |
0 ± 0.00 |
7.7 ± 0.58* |
A. anthelmintica roots |
0 ± 0.00 |
8.7 ± 1.10 |
0 ± 0.00 |
0 ± 0.00 |
T. sericea seeds |
0 ± 0.00 |
14.7 ± 1.58* |
17.3 ± 1.53* |
15 ± 1.00* |
T. sericea twigs |
0 ± 0.00 |
13.3 ± 1.20* |
0 ± 0.00 |
0 ± 0.00 |
L. camara seeds |
20.7 ± 1.58 |
22 ± 1.00 |
21.7 ± 0.58 |
20.3 ± 0.58 |
L. camara twigs |
13.7 ± 1.58* |
12 ± 1.00* |
13.7 ± 0.58* |
14.7 ± 1.15* |
C. tridens whole |
12.3 ± 1.58 |
12.3 ± 1.58 |
13 ± 1.00 |
12.7 ± 1.15 |
Methanol |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
Distilled water |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
Tetracycline |
20 ± 0.60 |
20.7 ± 0.6 |
18 ± 0.00 |
25.7 ± 0.58 |
The results of this study help to validate the efficacy of 18 extracts prepared from medicinal plants used as alternatives to treat gastrointestinal conditions in Northern Namibian. In this study, L. camara organic extracts displayed higher antibacterial activity against Salmonella and E. coli strains. These findings are consistent with that of Sharma et al.,28 who reported significantly higher activity in extracts of L. camara and C. tridens, respectively. Moreover, other studies have been conducted globally on the efficacy of T. sericea and G. tenax against diarrheal pathogens, but none of these studies tried to compare the efficacy in clinical and reference strains. This is important because, as observed in this study, there is a significant difference in the efficacy of extracts against clinical and reference strains; hence, the use of laboratory isolates alone will not fully reflect the efficacy of the extract in clinical isolates. Although reference strains are useful in susceptibility tests, their uses should not replace clinical isolates, and this is supported by the findings observed in this study. This is important, especially when analyzing the antibacterial effects of extracts that are being considered in clinical applications.
Isaiah and Arun et al.27 reported that C. tridens had antibacterial activity against Staphylococcus aureus, Bacillus subtilis, Klebsiella aerogenes, and Escherichia coli, the findings of this study also demonstrated broad-spectrum antibacterial activity against clinical and reference strains of Salmonella and E. coli were observed in its aqueous and organic extracts. Based on statistical analysis, there was a significant difference between the antibacterial activity of G. tenax roots, A. anthelmintica twigs, and T. sericea seeds (p < 0.001) to clinical and reference strains of Salmonella. Meanwhile, G. tenax roots, A. anthelmintic twigs, A. anthelmintic roots, T. sericea seeds, and T. sericea twigs showed a significant difference in their antibacterial activity against reference and clinical isolates of E. coli.
The minimum inhibitory concentration of different extracts with antibacterial activity
Results obtained from the validation and quality control assay for MIC by resazurin were within the range of recommended MIC values of the Clinical and Laboratory Standards Institute15 as shown in Table 4. Clinical diarrheal pathogens showed higher sensitivity to organic extracts in comparison to reference strains. Among the aqueous extracts, L. camara showed the lowest MIC value of 125 µg/mL against the clinical isolate of Salmonella (Table 5). Moreover, L. camara organic seed extract showed the lowest MIC and minimum bactericidal concentration (MBC) of 31.25 µg/mL against clinical E. coli 157:H7 VTEC and reference strain of E. coli (Table 5). A. anthelmintica organic and aqueous extracts showed less sensitivity to clinical and reference strains in this study. Distilled water and 99% methanol that were used as negative controls did not show any inhibitory properties against the test organisms.
Table (4):
Determination of the MIC for ATCC Gram-negative strains for quality control purposes
| Bacteria | Antibiotic | MIC from this study | MIC recommended by CLSI (µg/ml) |
|---|---|---|---|
| Salmonella Typhi ATCC 13311 | Gentamycin | 1.0 | 2-32 |
| Ciprofloxacin | 2 | 0.03-4 | |
| Tetracycline | 3.0 | 2-32 | |
| Escherichia coli ATCC 25922 | Gentamycin | 1.0 | 0.25-1 |
| Ciprofloxacin | 0.005 | 0.004-0.015 | |
| Tetracycline | 2.0 | 0.5-2 |
While aqueous extracts are normally used in the ethnomedicinal setting to prepare decoctions for treating gastrointestinal, the findings of this study showed minimal efficacy in aqueous extracts in comparison to organic extracts. This could be because bioactive compounds extracted using water require further metabolism to perform bacterial inhibitory properties.28,29 Moreover, while roots are reported to have potent antibacterial activity in other studies, in this study, seeds of the studied plants have shown potent antibacterial activity with significantly lower MIC values. This is important especially since according to the global conservative estimate, the current loss of plant species is between 100 and 1000 times higher than the expected natural extinction rate, and the Earth is losing at least one potential major drug every 2 years29; hence, using plant parts that prevent uprooting and killing plants while still offering maximum therapeutic benefits is very important.
Sharma et al.28 reported the MIC of G. tenax root extracts to be 62.5 and 125 µg/ml for aqueous and organic extracts, respectively. However, in this study, only the organic twig extract of G. tenax showed antibacterial activity with the lowest MIC of 62.5 against clinical Salmonella isolate (Table 5). Moreover, other studies have reported MIC values of L. camara than the MIC of 31.5 µg/mL that are reported in this study. Opportunistic infections such as gastroenteritis are quite inevitable among children under the age of 5 and immunocompromised patients; hence, finding alternative medicine to eliminate them is crucial and one way to mitigate the challenge of microbial resistance.30,31 The findings of this study are the first to use a resazurin assay to evaluate MIC values of methanol and aqueous plant extracts. These findings have shown the easy reproducibility and accuracy of MIC evaluation by resazurin and agree with the findings of Elshikh et al.31 These findings also show the potential of the resazurin-based 96-well plate microdilution method as an interpretational tool for testing the MIC of natural products.
Table (5):
MIC of organic and aqueous plant extracts against clinical and laboratory isolates of Salmonella and E. coli by Resazurin microtiter dilution assay
| Plant name | Part used | MIC of plant extract | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Aqueous extracts | Organic extracts | ||||||||
| C.E | R.E | C.S | R.S | C.E | R.E | C.S | R.S | ||
| G. tenax | Seeds | – | – | – | – | 250 | – | 62.5 | 125 |
| Roots | – | – | – | – | – | – | – | – | |
| A. anthelmintica | Twigs | – | – | – | – | 500 | 500* | 500 | 250 |
| Roots | – | – | – | – | – | 250* | – | – | |
| T. sericea | Seeds | – | 500 | – | 500 | – | 125* | 62.5 | 62.5 |
| Twigs | – | 500 | – | 500 | – | 250* | – | – | |
| L. camara | Seeds | 250 | 250 | 250 | 250 | 31.25 | 31.25* | 62.5 | 62.5* |
| Twigs | – | 250 | – | – | 31.25 | 62.5* | 62.5 | 125* | |
| C. tridens | Whole | 250 | 500 | – | – | 125 | 125 | 62.5 | 62.5 |
| Tetracycline | – | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 |
C.E= Clinical E. coli; R.E= Reference E. coli; C.S= Clinical Salmonella; R.S= Reference Salmonella; MIC values are expressed in µg/mL; SEM: standard error of the mean; n = 3; (-): mean MIC above 500 µg/mL, *p < 0.05 significant between the difference in MICs different parts of the same plant
A comparison of the antibiogram activity of selected antibiotics against clinical and reference bacteria causing gastroenteritis
A higher antibiotic multidrug resistance pattern was observed among all 4 pathogens tested in this study. The reference strain of E. coli showed resistance to antibiotics such as amoxicillin 10 µg, ciprofloxacin 10 µg, tetracycline 30 µg/mL, erythromycin, penicillin, vancomycin, and ampicillin (Table 6). Among the 10 antibiotics used in the antigram assay, clinical E. coli 157:H7 VTEC has shown resistance to 70% with sensitivity only observed in tetracycline (20.33 ± 0.58 mm), gentamycin (10.00 ± 0.00 mm), and ciprofloxacin (15 ± 0.00 mm) (Table 6). Meanwhile, the S. Typhimurium reference strain showed resistance to 50% of the standard antibiotics. Furthermore, a 70% resistance was also observed in clinical isolates of Salmonella with sensitivity only observed in ciprofloxacin (10 ± 0.00 mm), gentamicin (9 ± 0.00 mm), and tetracycline (18 ± 0.00 mm) (Table 6). Overall, the clinical isolates of Salmonella and E. coli 157:H7 VTEC showed the highest resistance pattern with activity only observed in 3/10 antibiotics tested in comparison to the reference strains. This agrees with the findings of the lowest MIC value for tetracycline that was used as a positive control in the MIC test was 2.0 ± 0.00 µg/mL (Table 2).
Table (6):
Antigram activity of standard antibiotics against clinical and reference strains of Salmonella spp. and E. coli spp.
Antibiotics (µg/mL) |
Average inhibition of reference strain of Salmonella isolates (mm) |
Average inhibition of clinical Salmonella isolates (mm) |
Average inhibition of reference strain of E. coli |
Average inhibition of clinical E. coli isolates (mm) |
|---|---|---|---|---|
Amoxicillin 10 |
9.7 ± 0.58* |
0 ± 0.00* |
0 ± 0.00 |
0 ± 0.00 |
Cephalothin 30 |
34 ± 1.73* |
0 ± 0.00* |
18.7 ± 0.58* |
0 ± 0.00* |
Sulphathiazole 25 |
30.3 ± 0.58* |
0 ± 0.00* |
20.3 ± 0.58 |
0 ± 0.00 |
Ciprofloxacin 10 |
0 ± 0.00* |
10 ± 0.00* |
0 ± 0.00* |
15 ± 0.00* |
Gentamicin 10 |
28.3 ± 0.58 |
9 ± 0.00 |
20.3 ± 0.58* |
10.3 ± 0.58* |
Tetracycline 30 |
25.7 ± 0.58* |
18 ± 0.00* |
20.7 ± 0.58 |
20 ± 0.00 |
Erythromycin |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
penicillin G 10 |
20.3 ± 0.58* |
0 ± 0.00* |
0 ± 0.00 |
0 ± 0.00 |
Vancomycin 30 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
Ampicillin 10 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
0 ± 0.00 |
In Namibia, morbidity, and mortality due to drug-resistant Gram-negative gastrointestinal pathogens are of great concern due to the lack of surveillance data for antibiotics. Moreover, antimicrobials prescription without following the Namibian treatment guidelines for local and regional antimicrobial sensitivity data is also reported in Namibia.32 Although antibiotics such as amoxicillin, sulphathiazole, ciprofloxacin, and ampicillin are used in Namibia as first-line drugs to treat E. coli 157:H7 VTEC and Salmonella infection, their uses may not always be effective due to the higher resistance pattern observed in this study. Comparative studies looking at the effectiveness of medicinal plants and antibiotics against both clinical isolates and reference strains are therefore important as they help to eliminate the limitation possessed using only reference strains in drug development. Antibiotic resistance develops as an evolutionary response; hence, it must be visualized as an evolving phenomenon that demands constant surveillance and continuous efforts to identify the degree of resistance to available antibiotics while creating strategies to combat this problem.33,34 The higher antibiotic resistance pattern against 10 antidiarrheal antibiotics, especially against clinical isolate reported in this study, is alarming and this makes the plant extracts especially organic extracts of L. camara, T. sericea, G. tenax, and C. tridens with broad-spectrum and potent inhibitory activity against MDR gastrointestinal pathogens worth further analysis in formulating alternative plant-based medicine for gastroenteritis however the toxicity profile of these extracts have to be determined.
Cytotoxicity effect against NIH/3T3 cell line
Making assumptions that plant-based extracts are safe due to their long history of use could be misleading and dangerous. Hence, cell-based assays are often used for screening novel formulations to determine, if the test molecules are having direct cytotoxic effects. MTT assay was used in the present study. With this assay, viable cells with active metabolism can convert MTT into a purple-colored formazan with a maximum absorbance of 590 nm. When cells die, they lose the ability to convert MTT into formazan; thus, color formation is the marker of only viable cells.35,36
The cytotoxic activities of the methanol extracts of L. camara, C. tridens, G. tenax, and T. sericea were determined at various concentrations on 3T3 cells. Using the absorbance from the microplate reader values of percentage cell viability of the 3T3 cells at different concentrations of the extracts after 48 hours’ incubation are shown in Figure 1. The values for cell viability with methanol extracts at the highest concentration of 100 µg/mL (Figure 1) were 38.78% for G. tenax, 63.08% for T. sericea, and 50.52% for C. tridens, and 51.68% for L. camara. These values were higher than the recorded 29.81% cell viability of the negative control (10% triton-x + media) which is known to kill cells by lysing the cell membrane and solubilizing mitochondrial membranes releasing mitochondrial matrix proteins. At the lowest concentration of 1.56 µg/mL for all extracts, the percentage viability was between 80.05 ± 0.06% and 97.63 ± 0.14%.
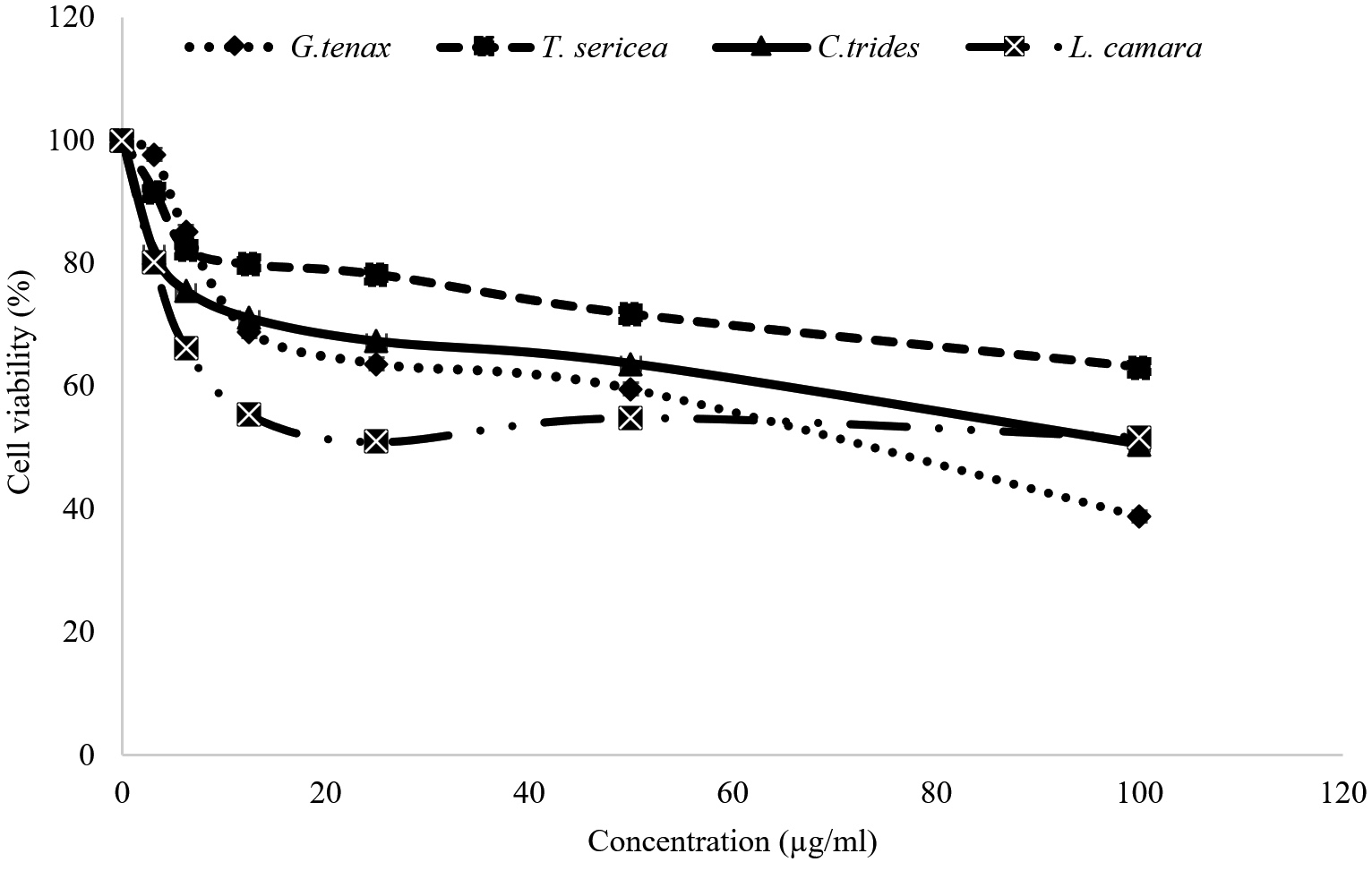
Figure 1. MTT assay on 3T3 cells to evaluate the effect of the L. camara, C. tridens, G. tenax, and T. sericea methanol extract on the cell growth after 48 hours. Values are the means of three independent experiments
This study is the first to demonstrate the effect of L. camara, C. tridens, G. tenax, and T. sericea methanol extracts on 3T3 cell lines. The results showed that all L. camara, C. tridens, G. tenax, and T. sericea extracts showed a decrease in cell viability of the 3T3 cell line as the concentration increased. In this study, a threshold of the cytotoxic concentration (IC50) below
20 µg/mL was considered to be toxic, and above 20 µg/mL to be non-toxic.37-39 A study by Koley and Bard,40 on the effect of Triton-x on HeLa, has also demonstrated toxicity due to changes in cell morphology, membrane permeability, and viability caused by the presence of Triton-x. Among the four studied extracts, G. tenax showed the highest average half-maximal inhibitory concentration (IC50) values of 91.50 ± 0.14 µg/mL after 48 hours. Meanwhile, the T. sericea extract showed a lower IC50 of 65.85 ± 0.07 µg/mL as depicted in Figure 2. It is important to note that while L. camara is used in Namibia in the traditional setting to treat different diseases, other studies such as Pour et al.25 have reported on the toxicity of L. camara in livestock, because it contains secondary metabolites such as naphthoquinones, irdoid, glycosides, and the most poisonous lantadenes. Lantadenes are pentacyclic triterpenes that are also reported to cause hepatotoxicity and jaundice, as well as photosensitization. However, since the IC50 of all the extracts is above 20 µg/mL which was set as the threshold for cytotoxicity, the extracts are considered not toxic.
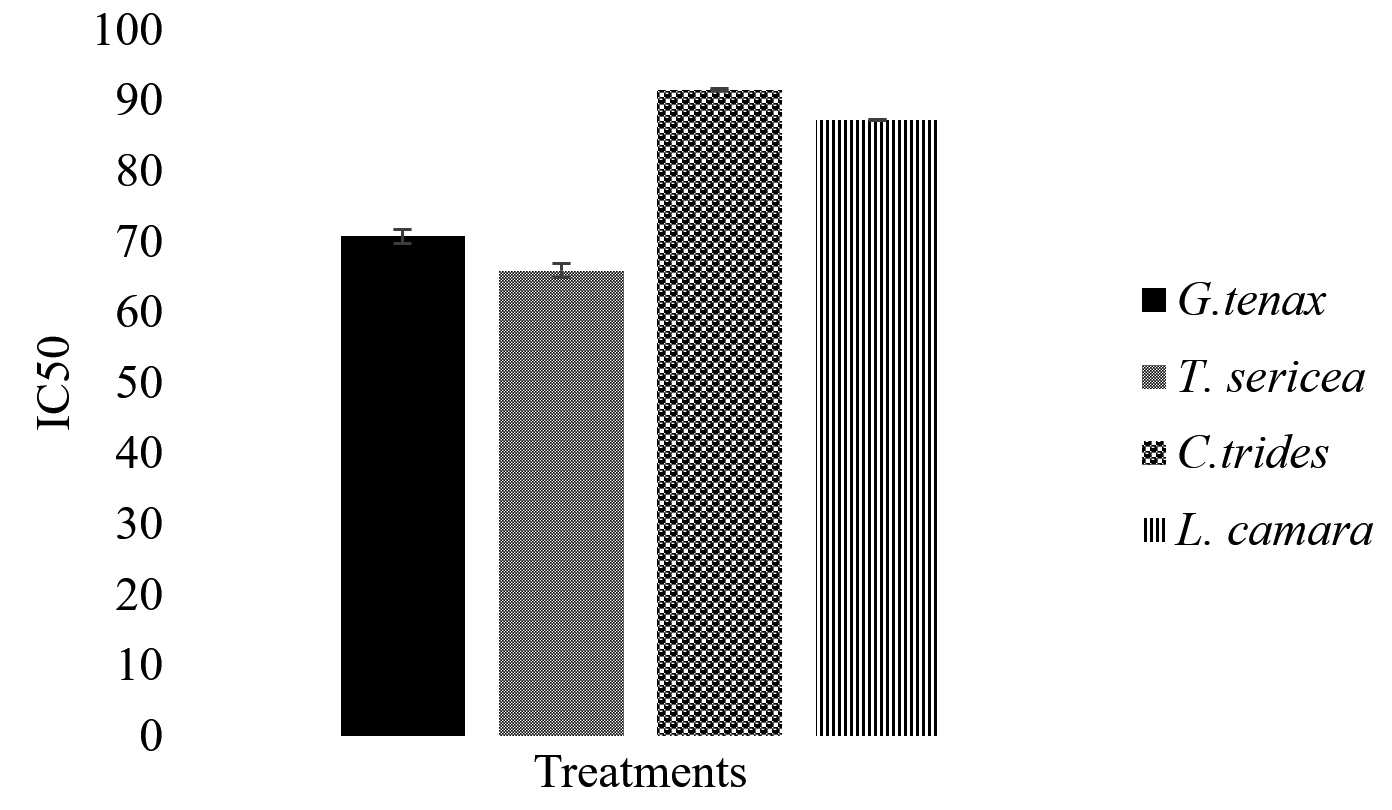
Figure 2. In vitro cytotoxic activity of selected antidiarrheal medicinal plant extracts on 3T3 cell line
This is the first study to demonstrate the outstanding antibacterial activity of L. camara, C. tridens, and G. tenax against multidrug-resistant clinical strains of Salmonella and E. coli and their toxicity profiles to 3T3 cells. The study has shown that the extracts have potent broad-spectrum antibacterial properties. Moreover, these extracts have proven not to be toxic to 3T3 cells at concentrations ≤100 µg/mL. This makes these extracts potential sources for further complementary and alternative medicine for bacterial gastroenteritis, but only after a safe dosage regimen is validated in vivo.
ACKNOWLEDGMENTS
The authors would like to acknowledge the University of Namibia for providing the facilities to conduct the study. Authors also would like to thank the Omusati region knowledge holders for providing guidance in collecting the medicinal plants.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
Both authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study was approved by the University of Namibia’s ethical committee (SOS-0039). The identification of the plants was conducted by the National Herbarium of Namibia (permit number: 2221/2017).
- Giannattasio A, Guarino A, Lo Vecchio A. Management of children with prolonged diarrhea. F1000Research. 2016;5:F1000.
Crossref - Ochoa TJ, Contreras CA. Enteropathogenic E. coli infection in children. Curr Opin Infect Dis. 2011;24(5):478-483.
Crossref - World Health Organization. International organizations unite on critical recommendations to combat drug-resistant infections and prevent staggering number of deaths each year. 2019. https://www.who.int/news/item/29-04-2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis. Accessed on July 2021.
- Mokomane M, Kasvosve I, de Melo E, Pernica JM, Goldfarb DM. The global problem of childhood diarrhoeal diseases: emerging strategies in prevention and management. Ther Adv Infect Dis. 2018;5(1):29-43.
Crossref - Fletcher SM, Stark D, Ellis J. Prevalence of gastrointestinal pathogens in Sub Saharan Africa: systematic review and meta-analysis. J Public Health Africa. 2011;2(2):e30.
- Iikasha AMN, Quaye IK, Mumbengegwi DR. Evaluation of Antibacterial Activity of Medicinal Plant Extracts Against Clinical Isolates of Pathogens from Children with Acute Gastroenteritis at Katutura State Hospital¸ Windhoek, Namibia. Afr J Tradit Complement Altern Med. 2020;16(2):15-23.
- Shen H, Zhang J, Li Y, et al. The 12 Gastrointestinal Pathogens Spectrum of Acute Infectious Diarrhea in a Sentinel Hospital, Shenzhen, China. Front Microbiol. 2016;7:1926.
Crossref - Crum-Cianflone NF. Salmonellosis and the gastrointestinal tract: More than just peanut butter. Curr Gastroenterol Rep. 2008;10(4):424-431.
Crossref - Mabbott NA, Donaldson DS, Ohno H, Williams IR, Mahajan A. Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal Immunology. 2013;6(4): 666-677.
Crossref - Ugboko H, De N. Mechanisms of Antibiotic resistance in Salmonella typhi. Int J Curr Microbiol Appl Sci. 2014;3(12):461-476.
- Nataro JP, Kaper JB. Diarrheagenic Escherichia coli. Clin Microbiol Rev. 1998;11(1):142-201.
Crossref - Ameer MA, Wasey A, Salen P. Escherichia Coli (E. Coli 0157 H7). StatPearls [Internet]. Treasure Island (FL): StatPearls. 2021. https://www.ncbi.nlm.nih.gov/books/NBK507845/
- Bruzzese E, Giannattasio A, Guarino A. Antibiotic treatment of acute gastroenteritis in children. F1000Research. 2018;7:193.
Crossref - Pavlinac PB, Tickell KD, Walson JL. Management of diarrhea in HIV-affected infants and children. Expert Rev Anti Infect Ther. 2015;13(1):5-8.
Crossref - CLSI M100-Ed34 includes updated tables for the Clinical and Laboratory Standards Institute antimicrobial susceptibility testing standards CLSI M02| Performance Standards for Antimicrobial Disk Susceptibility Tests, 14th Edition (2024), CLSI M07| Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 12th Edition (2024), and CLSI M11 | Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria, 9th Edition (2018). https://clsi.org/standards/products/microbiology/documents/m100/
- Prabhuswamy B, Kuruni N, Mallikarjun N. Antibiogram profile and antibacterial efficacy of medicinal plant extracts on Streptococcus mutans: An in vitro study. Int J Clin Dent Sci. 2016;7:1-7.
- Baekkeskov E, Rubin O, Munkholm L, Zaman W. Antimicrobial Resistance as a Global Health Crisis. Oxford Research Encyclopedia of Politics. 2022.
Crossref - Fair RJ, Tor Y. Antibiotics and bacterial resistance in the 21st century. PMC. 2014;6:25-64.
Crossref - Dhama K, Tiwari R, Chakraborty S, et al. Evidence based antibacterial potentials of medicinal plants and herbs countering bacterial pathogens especially in the era of emerging drug resistance: An integrated update. Int J Pharmacol. 2014;10(1):1-43.
Crossref - Njume C, Goduka NI. Treatment of diarrhoea in rural african communities: An overview of measures to maximise the medicinal potentials of indigenous plants. Int J Environ Res Public Health. 2012;9(11):3911-3933.
Crossref - Ugboko HU, Nwinyi OC, Oranusi SU, Oyewale JO. Childhood diarrhoeal diseases in developing countries. Heliyon. 2020;6(4):e03690.
Crossref - Shatri AMN, Mumbengegwi DR. Ethnomedicinal use and phytochemical analysis of medicinal plants used to treat gastrointestinal conditions by Awambo people in Iikokola Village, Namibia. Scientific African. 2022;18:e01428.
Crossref - Global Salm-Surv. MIC susceptibility testing of Salmonella and Campylobacter. 4th edition. 2003. http://antimicrobialresistance.dk/CustomerData/ Files/Folders/6-pdf-protocols/54_anti4-pdf.pdf
- Gahlaut A, Chhillar AK. Evaluation of the antibacterial potential of plant extracts using resazurin based microtiter dilution assay. Int J Pharm Pharm Sci. 2013;5(2):372-376.
- Pour BM, Latha LY, Sasidharan S. Cytotoxicity and oral acute toxicity studies of Lantana camara leaf extract. Molecules. 2011;16(5):3663-3674.
Crossref - Zhang, QW, Lin LG, Ye WC. Techniques for extraction and isolation of natural products: a comprehensive review. Chin Med. 2018;13:20.
Crossref - Isaiah S, Senthamizh SN, Arun KC. Phytochemical screening, anti-microbial activity and GC-MS analysis of Corchorus tridens L. Int J Pharm Res. 2016;6(12):353-357.
- Sharma C, Malgaonkar M, Sangvikar S, Murthy S, Pawar S. In vitro evaluation of antimicrobial and antioxidant profile of Grewia L. root extracts. J Appl Life Sci Int. 2016;7(1):1-9.
Crossref - Chen SL, Yu H, Luo HM. Conservation and sustainable use of medicinal plants: problems, progress, and prospects. Chin Med. 2016;11:37.
Crossref - Voravuthikunchai SP, Phongpaichit S, Subhadhirasakul S. Evaluation of antibacterial activities of medicinal plants widely used among AIDS patients in Thailand. Pharm Biol. 2005;43(8):701-706.
Crossref - Elshikh M, Ahmed S, Funston S, et al. Resazurin-based 96-well plate microdilution method for the determination of minimum inhibitory concentration of biosurfactants. Biotechnol Lett. 216;38(6):1015-1019.
Crossref - Mapiye S. Analysis of the antibacterial activity of Terminalia sericea and Combretum imberbe two Combretaceae species from Namibia (Master’s thesis, University of Namibia, Windhoek, Namibia). 2019. https://repository.unam.edu.na/bitstream/handle/11070/2872/samson_2019.pdf?sequence=1&isAllowed=y
- Munita JM, Bayer AS, Arias CA. Evolving resistance among gram-positive pathogens. Clin Infect Dis. 2015;61(Suppl 2):S48-S57.
Crossref - Mohamed MA, Nasr M, Elkhatib WF, Eltayeb WN. In vitro evaluation of antimicrobial activity and cytotoxicity of different nanobiotics targeting multidrug resistant and biofilm forming Staphylococci. BioMed Res Int. 2018;2018:7658238.
Crossref - Njeru SN, Muema JM. In vitro cytotoxicity of Aspilia pluriseta Schweinf. extract fractions. BMC Res Notes. 2021;14(1):57.
Crossref - Supino R. MTT Assays. In: O’Hare, S., Atterwill, C.K. (eds) In Vitro Toxicity Testing Protocols. Methods Mol Boil. 1995.
Crossref - Mongalo NI, McGaw LJ, Finnie JF, Van Staden J. Pharmacological properties of extracts from six South African medicinal plants used to treat sexually transmitted infections (STIs) and related infections. S Afr J Bot. 2017;112:290-295.
Crossref - Afagnigni AD, Nyegue MA, Djova SV, Etoa F-X. LC-MS analysis, 15-lipoxygenase inhibition, cytotoxicity, and genotoxicity of dissotis multiflora (Sm) triana (melastomataceae) and Paullinia pinnata Linn (Sapindaceae). J Trop Med. 2020;2020:5169847.
Crossref - Elisha IL, Botha FS, McGaw LJ. The antibacterial activity of extracts of nine plant species with good activity against Escherichia coli against five other bacteria and cytotoxicity of extracts. BMC Complement Altern Med. 2017;17:133.
Crossref - Koley D, Bard AJ. Triton X-100 concentration effects on membrane permeability of a single HeLa cell by scanning electrochemical microscopy (SECM). Proc Natl Acad Sci. 2010;107(39):16783-16787.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.