


ISSN: 0973-7510
E-ISSN: 2581-690X
Endophytic microorganisms reside within the plant tissues in a symbiotic association and generally refer to fungi and bacteria. Endophytes benefit the host plants in several ways. They are known to protect the plants from harsh environmental conditions, produce variety of compounds which boost the growth of the plants, render protection against herbivores and also serve as potent biocontrol agents. Screening of fungal species for implementation on such fields has been the area of interest. The current study aims to isolate fungal endophytes from P. napaulensis and screen for its antagonistic efficiency against common wide host range fungal pathogens via extracellular enzymatic activities and dual culture methods. Thirteen fungal endophytes were isolated which showed varying range of extracellular enzymes production and growth inhibition of the fungal pathogens. The study highlights potent fungal isolates, which can be implemented in bio-agriculture (Trichoderma atroviride, T. koningiopsis and Xylaria feejeensis) and also for mass and sustainable production of hydrolytic enzymes such as cellulase and amylase and which have now gained interest in industrial sectors (Chaetomium globosum, Colletotrichum gloeosporioides and Fusarium graminearum).
Prunus napaulensis, Fungal Endophytes, Antagonists, Extracellular Enzymatic Activities
Endophytes, generally referred to fungi and bacteria growing within the plant tissues in symbiotic association, have been characterized as defenders of the host plants against the attack of other microorganisms, insects and herbivores.1 Besides, they also produce phytohormones, enzymes and other bioactive compounds which are of advantages to the host plant.2 Endophytic fungi have pulled great attention in the recent past due to its efficiency in producing novel secondary metabolites that can be exploited in various sectors such as agriculture, varied industries and medicines.1 The implementation of biocontrols in agriculture have also gained parallel interest. Unlike application of synthetic chemicals, biocontrol agents offer a more sustainable approach by reducing reliance on chemicals which not only deteriorates the environment but also pose ill impact on humans and animals.3,4 Antagonists significantly combat the population of fungal pathogen residing on the aerial parts of the host plant and decrease the intensity of diseases.5,6 Recent researches have been focused on isolation of potent endophytic antagonists. Antagonists exhibits strong colonization ability and have superior environmental adaptability.7 Fungal biocontrol agents have gained popularity alongside bacteria (particularly Bacillus thuringiensis), primarily because of its effectiveness as disease control agent against broad spectrum of pathogens.8,9 Besides, extracellular hydrolytic enzymes are also known to be secreted by these fungi for instance, amylase, cellulase, protease, catalase, etc, which assist the antagonists in combating growth and effect of pathogens.
Prunus napaulensis (Ser.) Steud. (syn. Prunus nepalensis L.), a wild indigenous fruit tree growing in the temperate Himalayan regions of India at altitudes between 1200-3000 m, is popular for its high nutritional and medicinal attributes. It is also considered to have potent antidiabetic and anticancer properties.10-12 In Meghalaya the plant is distributed in the Khasi and Jaintia Hills districts, and is locally known as Sohiong. This study provides input on fungal endophytes isolated from P. napaulensis, their potency to produce various extracellular enzymes and antifungal efficacy against wide host range pathogens.
Study area and collection of samples
Fresh and healthy leaves, twigs, roots and barks of P. napaulensis were collected from Shangpung (Lat. 25.47816°, Long. 92.35328°) West Jaintia Hills, Meghalaya, India. The samples were transferred in sterilized plastic zip bags and brought to laboratory to carry out further studies.
Isolation of fungal endophytes and maintenance of pure culture
The plant samples (leaves, twigs, roots and barks) were washed clean with running water in order to remove any unwanted entities which might have adhered to the surface of the samples. Surface sterilization was done by subjecting the plant samples to 70% ethanol (Analytical AR Grade Ethanol Absolute) for 2-3 minutes, followed by 1% sodium hypochlorite (HiMedia) for 1-2 minutes and 2-3 times washing with sterile distilled water.13 About 1 cm of the explants were cut from the sterilized samples, air dried and inoculated in potato dextrose agar (HiMedia) plates containing 100 mg/L of streptomycin sulphate (HiMedia). The plates were incubated for 10 days at 28 ± 2 °C and the pure cultures for the obtained endophytic fungal isolates were maintained following the same procedure.14
Identification of fungal endophytes
The isolated fungal endophytes were identified morphologically and microscopically based on their vegetative and reproductive structures using standard manuals.15,16 For molecular characterization the fungal isolates were cultured in potato dextrose broth (HiMedia) for 10 days and were then used for identification by adapting the procedures given by National Bureau of Agriculturally Important Microorganisms (NBAIM) by analysing the Internal transcribed Spacer (ITS) region. The amplification of locus analysed (ITS) was performed using universal primers for the ITS region, ITS 1 and ITS 4.17 The obtained PCR products were purified and sequenced at Eurofins Genomics India Pvt. Ltd., Bengaluru, Karnataka, India. Analysis of the sequences were carried out using BLAST program from GenBank database (NCBI), sequence with highest similarity was downloaded. Sequences were aligned using MEGA 11 software and using Clustal W alignment program. The resulting sequences were deposited to the GenBank database and the accession numbers were acquired.
Analysis of enzymatic assay
Amylase test
Qualitative analysis for amylase was done by inoculating isolates in starch agar (HiMedia) plates and incubated at 28 ± 2 °C for 10 days. Following incubation, the plates were flooded with 0.5% iodine solution for a minute. Occurrence of clear zones around the colonies depicted positivity for amylase production.18 Quantification of amylase was done by using the filtrates of 10 days old fungal broth grown in potato dextrose broth (PDB) containing 1% starch. A reaction mixture comprising of 1 ml of filtrate (enzyme broth) and 0.5 ml of soluble starch (1%) was incubated for 30 minutes at 37 °C and then 0.5 ml Dinitrosalicylic acid (DNS) was added. The solution was diluted to 5 ml by adding distilled water. The optical density (OD) of the developed reddish brown colour was recorded at 540 nm using UV/Vis Spectrophotometer (Lambda 35, Perkin Elmer, USA). Readings were taken in triplicate and quantity of sugar produced was read off by considering the standard curve of maltose. The result was expressed in unit per ml (Uml-1), where one unit of amylase is the amount of enzyme releasing a µmol of reducing sugar, expressed as maltose, per min, per ml, under given condition.19
Cellulase test
Qualitative analysis for cellulase was determined by inoculating fungal isolates in PDA plates amended with 1% carboxymethylcellulose (CMC) and incubating for 10 days at 28 ± 2 °C. Following incubation, plates were flooded with congo red solution (0.1%) for 15 minutes, the excess stain was washed with 1M NaCl solution for 10 minutes.20 Occurrence of clear zones around the colonies indicated positive activity for cellulase. Quantification of Endo-β-1,4-glucanase (Carboxymethyl cellulase) was done by centrifuging 10 days grown PDB cultures of endophytic fungi at 10,000 rpm (Eppendorf, Centrifuge 5430 R) for 15 minutes. The reaction mixture comprised of 1 ml supernatant and 1 ml of 1% CMC in 0.05 M sodium citrate buffer with pH 4.8. The mixture was incubated at 50 °C for an hour and the 3 ml of alkaline DNS reagent was added to halt the reaction. Absorbance was measured at 540 nm and readings were taken in triplicates. The result was expressed in Unit per ml (Uml-1), where one unit of enzyme (CMC-ase) is the amount of enzyme releasing a µmol of reducing sugars, expressed as glucose, per min, per ml, under given condition.19
Protease test
Protease activity was screened by inoculating fungal isolates in skim milk agar plates followed by incubating for 10 days at 28 ± 2 °C.21 Occurrence of clear zones around the colonies specified positive activity of protease. Protease quantification was done by centrifuging the fungal cultures grown in PDB amended with 1% casein (10 days) at 10,000 rpm (using Eppendorf, Centrifuge 5430R) for 15 minutes. The reaction mixture comprised of 1 ml of the supernatant and 5 ml of 0.5 N NaOH. The mixture was mixed vortex and 1.5 ml of Folin-Ciocalteu reagent (diluted to 1:1 v/v) was added and incubated for 30 minutes in the dark at room temperature. Development of blue colour indicated protease activity and absorbance was measured at 660 nm using tyrosine standard. The result was expressed in unit per ml (Uml-1), where one unit of protease is the amount of enzyme releasing µmol of tyrosine per ml per min under the given condition.22
Catalase test
Catalase activity was determined by mixing small inoculum of the isolates with 3% hydrogen peroxide solution. Intensity of bubble evolution indicated the catalase producing potential of the fungi: (-) no activity, (+) low activity, i.e. few bubbles, (++) moderate activity, (+++) high activity, i.e. abundant bubbles.23
Hydrogen cyanide (HCN) production test
Production of HCN was detected by inoculating isolates onto PDA plates. The media used for the test contained 0.4% L-Glycine. Thin Whatman filter paper strips were soaked in a solution of 2% Na2CO3 and 0.5% picrate. Single strips were adhered to the lids of petri dish and secured using parafilm. The plates were incubated for 10 days at 28 ± 2 °C. Any alteration in the strips colour after the incubation period indicated the evolution of HCN.24
Urease test
Urease test was done by inoculating fungal isolates in plates containing Christensen’s urea agar (HiMedia) and incubating for 10 days at 28 ± 2 °C. Change in the colour medium denoted the urease activity: no change in colour implies negative (-), orange colour means weak activity (+), pink colour means moderate activity (++) and red colour means strong activity (+++).23
Isolation, identification and culture of fungal phytopathogens
Fungal phytopathogens were isolated from diseased parts of Sechium edule (leaf), Solanum tuberosum (leaf) and Mangifera indica (fruit pericarp and leaf), and identified following the methods as described above for the endophytic fungi.
Assessment of antagonistic potential of endophytic fungi in in vitro condition
The in vitro assessment of antagonistic potential of the isolated endophytic fungi was carried out by dual culture technique. A 6 mm mycelial disc of fungal pathogens and fungal endophytes obtained from 4 days old cultures were placed opposite to each other on PDA plates, secured with parafilm and incubated for 10 days at 28 ± 2 °C.25 Percentage of growth inhibition was determined using the formula:
Growth Inhibition % = R1-R2 / R1 × 100
Where, R1 indicates the radial growth of the pathogen’s control colony and R2 indicates the radial growth of the pathogen in a treated colony.
Determination of the zone of interaction between the fungal endophytes and pathogens’ colonies
The interaction zone between the fungal endophytes and pathogens in dual culture were studied using Scanning Electron Microscopy (SEM) in Sophisticated Analytical Instrument Facility, North-Eastern Hill University, Shillong, India. Samples were prepared based on the method given by NEHU-SAIF. Mycelial cubes of 0.5 cm were cut using sterilized blade at the interaction zone in a dual culture plate. The samples were fixed with 3% Glutaraldehyde for 2 hours and then immersed in 0.1 M Sodium Cacodylate buffer for 15 minutes at 4 °C to wash off the excess fixative (3 changes for 15 minutes each). The samples were then subjected to gradient dehydration using acetone (30%-100%) and followed by immersion in Tetramethylsilane for 10-15 minutes at 4 °C. The samples were oven dried at 26 °C to remove the moisture. The dried samples were mounted on SEM stubs, gold sputter coated and observed under SEM (JEOL JSM-6360 SEM) at 20 kV.
Data analysis
The data analysis (analysis of variance and Tukey’s HSD test at P ≤ 0.05) were done using SPSS, v.25 (Chicago, IL, USA). Evaluation of various extracellular enzymatic activities and antagonistic potential of the isolated endophytic fungi were done in triplicates.
Identification of endophytic fungi isolated from P. napaulensis
Following the isolation of endophytic fungi, 13 isolates were identified; their names and accession number of which are listed in Table 1.
Table (1):
Species identification of fungal endophytes isolated from P. napaulensis
No. |
Fungal endophytes |
Accession No. |
|---|---|---|
1 |
Alternaria alternata |
PP855860 |
2 |
Aspergillus fumigatus |
PP856661 |
3 |
Chaetomium globosum |
PP855861 |
4 |
Colletotrichum gloeosporioides |
PP856664 |
5 |
Curvularia verruciformis |
PP855862 |
6 |
Fusarium graminearum |
PP856667 |
7 |
Fusarium oxysporum |
PP856665 |
8 |
Hypoxylon lividipigmentum |
PP856668 |
9 |
Nemania primolutea |
PP855863 |
10 |
Diaporthe amygdali |
PP856669 |
11 |
Trichoderma atroviride |
PP856670 |
12 |
Trichoderma koningiopsis |
PP855865 |
13 |
Xylaria feejeensis |
PP855866 |
Analysis of enzymatic assay
The isolated fungal endophytes showed varying range of extracellular enzymatic activities (Table 2 and Figure 1). For amylase test 7 out of 13 endophytes tested positive. Chaetomium globosum (6.25 Uml-1) and Fusarium graminearum (6.02 Uml-1) were found to produce higher amount of amylase in comparison to the other isolates. In cellulase test 8 isolates showed positive response, Colletotrichum gloeosporioides was found to produce significantly higher cellulase (7.06 Uml-1) followed by Chaetomium globosum (3.75 Uml-1) and Fusarium graminearum (3.78 Uml-1). For the protease test only 4 isolates showed positive activity. Nemania primolutea was found to produce maximum (27.84 ± 0.5 Uml-1) of the enzyme. Aspergillus fumigatus, Trichoderma atroviride and Xylaria feejeensis produced protease in the range 2.80 ± 0.3 Uml-1, 4.06 ± 0.1 Uml-1 and 6.28 ± 0.1 Uml-1, respectively. In the qualitative screening, 6 isolates tested positive for catalase, only the Trichoderma species reported positive for HCN production and 5 isolates showed positive response for urease production (Table 3). Alternaria alternata showed negative response for all the extracellular enzymatic activities and HCN production.
Table (2):
Quantitative assessment of amylase, cellulase and protease produced by the fungal endophytes isolated from P. napaulensis
No. |
Fungal endophytes |
Amylase (Uml-1) |
Cellulase (Uml-1) |
Protease (Uml-1) |
|---|---|---|---|---|
1 |
Alternaria alternata |
0 |
0 |
0 |
2 |
Aspergillus fumigatus |
5.79 ± 0.1bc |
0.8 ± 0.1g |
2.80 ± 0.3c |
3 |
Chaetomium globosum |
6.25 ± 0.1a |
3.75 ± 0.1b |
0 |
4 |
Colletotrichum gloeosporioides |
0 |
7.06 ± 0.0a |
0 |
5 |
Curvularia verruciformis |
3.73 ± 0.0e |
0 |
0 |
6 |
Fusarium graminearum |
6.02 ± 0.4ab |
3.78 ± 0.0b |
0 |
7 |
Fusarium oxysporum |
4.27 ± 0.0d |
0 |
0 |
8 |
Hypoxylon lividipigmentum |
0 |
2.62 ± 0.4d |
0 |
9 |
Nemania primolutea |
0 |
1.47 ± 0.1f |
27.84 ± 0.5a |
10 |
Diaporthe amygdali |
4.64 ± 0.0d |
1.91 ± 0.7e |
0 |
11 |
Trichoderma atroviride |
0 |
0 |
4.06 ± 0.1bc |
12 |
Trichoderma koningiopsis |
0 |
0 |
0 |
13 |
Xylaria feejeensis |
5.48 ± 0.1c |
3.14 ± 0.1c |
6.28 ± 0.1b |
Data shows the mean ± S.D and Tukey’s HSD test at p<0.05. Values followed by different letters in the same column are significantly different
Table (3):
Qualitative assessment of catalase, HCN and urease production by the fungal endophytes isolated from P. napaulensis
No. |
Fungal endophytes |
Catalase |
HCN |
Urease |
|---|---|---|---|---|
1 |
Alternaria alternata |
– |
– |
– |
2 |
Aspergillus fumigatus |
+ |
– |
– |
3 |
Chaetomium globosum |
– |
– |
– |
4 |
Colletotrichum gloeosporioides |
++ |
– |
– |
5 |
Curvularia verruciformis |
+ |
– |
– |
6 |
Fusarium graminearum |
++ |
– |
+ |
7 |
Fusarium oxysporum |
– |
– |
+++ |
8 |
Hypoxylon lividipigmentum |
– |
– |
– |
9 |
Nemania primolutea |
– |
– |
+ |
10 |
Diaporthe amygdali |
++ |
– |
++ |
11 |
Trichoderma atroviride |
+++ |
+ |
+ |
12 |
Trichoderma koningiopsis |
– |
+ |
– |
13 |
Xylaria feejeensis |
– |
– |
– |
Data indicates positive and negative responses of fungal endophytes for the enzymatic tests: ‘-‘ for negative response; ‘+’ for positive response; ‘++’ for moderate response; ‘+++’ for high response
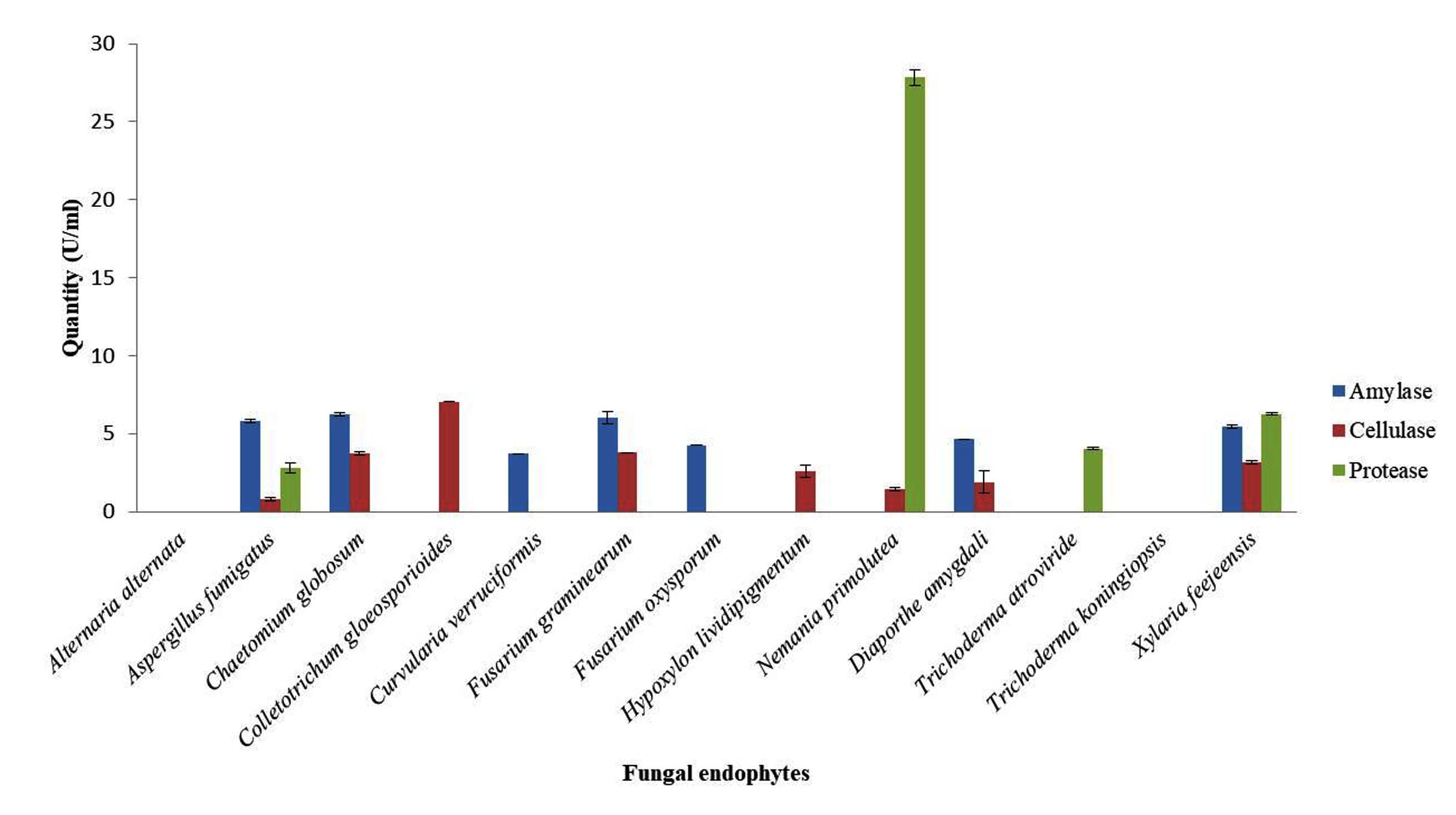
Figure 1. Graphical presentation of quantitative analysis of extracellular enzymes produced by the isolated fungal endophytes
Isolation and identification of fungal phytopathogens
The fungal pathogens isolated from selected crop plants were identified as Alternaria tenuissima, Colletotrichum asianum, Rhizopus sp. and Fusarium equiseti. The accession numbers obtained for identified pathogens have been specified in Table 4.
Table (4):
List of fungal pathogens isolated from crop plants
No. |
Fungal pathogen |
Host Plant |
Plant part |
Accession No. |
|---|---|---|---|---|
1 |
Alternaria tenuissima |
Solanum tuberosum |
Leaf |
PP856662 |
2 |
Colletotrichum asianum |
Mangifera indica |
Leaf |
PP856663 |
3 |
Rhizopus sp. |
Mangifera indica |
Fruit |
PP855864 |
4 |
Fusarium equiseti |
Sechium edule |
Leaf |
PP856666 |
Assessment of antagonistic potential of isolated fungal endophytes
The growth inhibition percentage for fungal endophytes varied from 11.33%-88.76%, depicting that all the isolated endophytes had some inhibitory potential against the test fungal phytopathogens (Figure 2 and Table 5). Trichoderma atroviride and T. koningiopsis showed the highest growth inhibition against Alternaria tenuissima (77.65%), Colletotrichum asianum (85.13% and 70.26%, respectively) and Fusarium equiseti (77.65%). While highest growth inhibition against Rhizopus sp. was observed by Fusarium graminearum (88.76%). Scanning electron microscopy between the endophytes and hosts showed morphological alterations in the pathogens’ hyphae, viz. distortion of pathogens’ hyphae, coiling of antagonist hyphae around the pathogens’ hyphae and also perforations in pathogens’ conidia (Figure 3).
Table (5):
Antagonistic potential of endophytic fungi against fungal phytopathogens
| No. | Endophytic fungal isolates | Growth Inhibition % of Fungal Phytopathogens | |||
|---|---|---|---|---|---|
| Alternaria tenuissima | Colletotrichum asianum | Rhizopus sp. | Fusarium equiseti | ||
| 1 | Alternaria alternata | 43.23 ± 1.2e | 33.37 ± 0.4f | 11.07 ± 0.1i | 35.83 ± 0.9g |
| 2 | Aspergillus fumigatus | 55.39 ± 0.3d | 11.33 ± 0.4h | 72.84 ± 0.6b | 32.43 ± 1.3h |
| 3 | Chaetomium globosum | 55.39 ± 0.3d | 41.69 ± 0.4e | 22.15 ± 0.1h | 38.80 ± 0.3f |
| 4 | Colletotrichum gloeosporioides | 38.93 ± 0.1f | 33.37 ± 0.4f | 44.05 ± 0.6e | 53.47 ± 0.2bc |
| 5 | Curvularia verruciformis | 65.79 ± 1.6b | 55.67 ± 0.2c | 36.31 ± 0.7g | 51.37 ± 0.6c |
| 6 | Fusarium graminearum | 61.07 ± 0.1c | 48.24 ± 0.3d | 88.76 ± 0.2a | 51.67 ± 0.6c |
| 7 | Fusarium oxysporum | 55.39 ± 0.3d | 48.24 ± 0.3d | 22.14 ± 0.1h | 46.53 ± 1.3d |
| 8 | Hypoxylon lividipigmentum | 65.79 ± 1.6b | 55.67 ± 0.2c | 72.84 ± 0.6b | 42.40 ± 0.8e |
| 9 | Nemania primolutea | 53.38 ± 0.1d | 41.45 ± 0.5e | 72.84 ± 0.6b | 38.80 ± 0.3f |
| 10 | Diaporthe amygdali | 62.94 ± 0.4c | 26.32 ± 0.3g | 38.89 ± 0.1f | 42.77 ± 1.4e |
| 11 | Trichoderma atroviride | 77.65 ± 0.2a | 85.13 ± 0.1a | 66.68 ± 0.1c | 70.50 ± 0.5a |
| 12 | Trichoderma koningiopsis | 77.65 ± 0.2a | 70.26 ± 0.3b | 55.39 ± 0.3d | 70.50 ± 0.5a |
| 13 | Xylaria feejeensis | 65.79 ± 1.6b | 55.67 ± 0.2c | 44.05 ± 0.6e | 55.13 ± 0.4b |
Data shows the mean ± S.D and Tukey’s HSD test at p<0.05. Values followed by different letters in the same column are significantly different
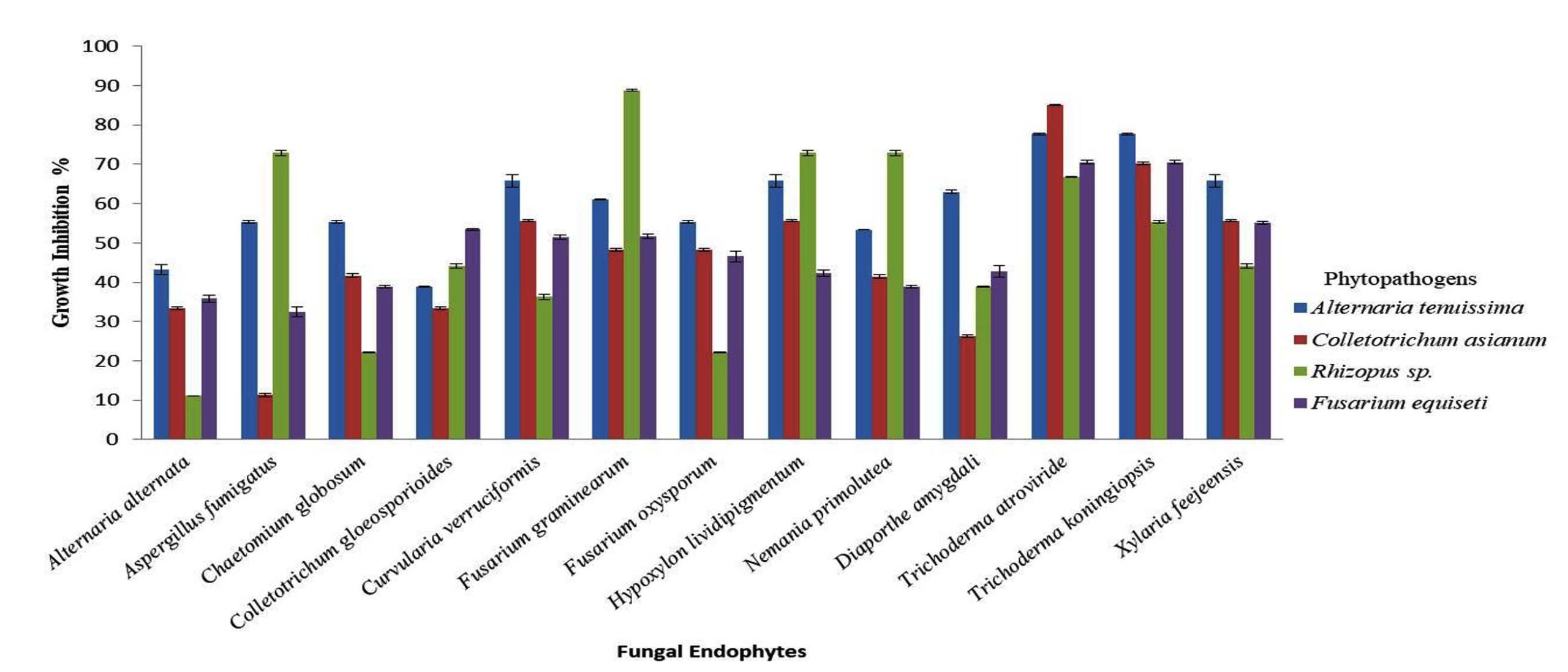
Figure 2. Graphical representation of antagonistic assessment of the isolated fungal endophytes against selected wide host phytopathogens

Figure 3. SEM micrographs of: (a) untreated Alternaria tenuissima (b) Alternaria tenuissima treated with Xylaria feejeensis (c) untreated Fusarium equiseti (d) Fusarium equiseti treated with Trichoderma koningiopsis
Biocontrol agents render crop protection against wide range of pathogens and unlike chemical pesticides and insecticides, biocontrol agents offer significant result as it ensures environmental safety, are target specific, do not produce hazardous residues and are easy to administer.26 Fungal endophytes are known to secrete various hydrolytic enzymes which facilitates its initial colonization in the host tissues, nutrient absorption from the host, hydrolyses complex macromolecules, exhibits mycoparasitism and also elicits defensive mechanisms against various pathogens or other external stress.27,28 In the host tissue, endophytes and the pathogens compete under the same micro ecological niche by means of the varied hydrolytic enzymes secreted by the fungal endophytes.29-31 The commonly produced hydrolytic enzymes by endophytic fungi are amylase, cellulase and protease.32 In the present study, most of the species were found to produce extracellular enzymes: 8 of 13 fungal endophytes produced cellulase in the range of 0.8-7.06 U/ml, 7 of 13 species produced amylase in the range 3.73-6.25 U/ml and 4 out of 13 species were found to produce protease in the range 2.80-27.84 U/ml. In the current study Aspergillus fumigatus, Chaetomium globosum, Fusarium graminearum, Diaporthe amygdali and Xylaria feejeensis were found to produce both amylase and cellulase. The qualitative analysis showed 6 of 13 fungal endophytes positive for catalase activity. Microbial catalase renders protection to the host against oxidative stress. In a recent study, Aspergillus fumigatus was reported to secrete three types of catalases of which one was found to produce by conidia and the other two by its mycelia.33 Microbial cyanides in general, are documented to control several plant diseases by increasing plants’ resistance during pathogenic attack, and by inhibiting energy metabolism or metalloenzyme activity.34-36 The present study showed that only the Trichoderma species were able to produce HCN. Also in the study, urea hydrolysis was observed only for five isolates- Fusarium graminearum, Fusarium oxysporum, Nemania primoletea, Diaporthe amygdali, and Trichoderma atroviride. It was thus observed that not all the isolated fungal endophytes produced all the enzymes. Production and regulation of extracellular enzymes by the fungal endophytes is the outcome of its genetic recombination along with the host elicitation and environmental factors.37 Maccheroni et al., reported in their study that factors such as variation in pH influenced the lytic activity of the endophytes.38 Similar results were also obtained by Shubha and Srinivas, and Hawar that not all the fungal endophytes could produce all the hydrolytic enzymes.39,40 Some produce only one of the hydrolytic enzymes while few produced none.
The isolated fungal endophytes all belong to Ascomycetes. Ascomycetes possess high antagonistic activity as they produce bioactive compounds with antimicrobial properties.41 As a fungal biocontrol agents Trichoderma sp. have been reported to share almost half of the market most preferably as soil or growth enhancers due to its wider antagonism potential against varied plant pathogens.42,43 Chaetomium, Penicillium, Aspergillus etc., have also been reported as commonly used antagonists.44 The antagonist activity can be regarded as the outcome of secondary metabolites production, competition, direct parasitism or development of resistance which can sometimes serve as a defensive tool in reducing the enzymatic activities of pathogens.45,46 In the present study, isolated fungal endophytes from P. napaulensis showed varying growth inhibition percentage against the selected fungal phytopathogens irrespective of their efficiency in producing extracellular enzymes. It can also be noted that Alternaria alternata showed negative response for all the proposed enzymatic activities and it scored the least per cent for growth inhibition. Studies have reported the involvement of microbial lytic enzymes in controlling infection against fungal pathogens like Rhizoctonia solani and Fusarium oxysporum.47,48 Thus, it can be inferred that production of extracellular enzymes enhances the antagonistic efficiency of the fungal endophytes.
Microbial antagonists imparts a significant role in restricting the population of fungal pathogens residing in the aerial parts of the host plants. Colonization by fungal endophytes have been reported in many plants which provides several benefits to the host including enhancing plant growth, increasing resistance against wider pathogens indirectly by some signalling pathways or directly by its antagonistic effect.49,50 Additionally, several industries like biomaterials, food, cosmetics, textile, fine chemicals, leather, etc., have started employing microbial enzymes such as amylase, cellulase and protease.51,52 Thus, these potential fungal endophytes can meet the needs of wider sectors.
Use of chemical fungicides on crop plants apparently makes the plants unfit for human consumptions as it is reported to have ill impact on human health apart from its adverse effect on the environment. These budding issues have let to the exploration of biocontrol systems which are more natural and environmentally acceptable to combat the problems. Fungi and bacteria are the commonly utilized biocontrol agents. The study provides the insights of fungal endophytes associated with P. napaulensis, the extracellular enzymes they produced and their antagonistic potential against various fungal phytopathogens. The tested endophytes showed competitive nature against the pathogens for space, nutrients and also mycoparasitism thereby restricting the growth of the pathogens. The findings add on to the environmentally friendly management strategies to minimise the application of synthetic chemicals for crop plants.
ACKNOWLEDGMENTS
The authors express gratitude to the CSIR-UGC NET Fellowship for providing financial aid, Eurofins Genomics India Pvt. Ltd., Karnataka, India, for performing sequencing of DNA samples and the Sophisticated Analytical Instrument Facility, NEHU, Shillong, for granting permission to conduct the Scanning Electron Microscopy.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
Both authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
This study was supported by CSIR-UGC NET Fellowship.
DATA AVAILABILITY
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Tan RX, Zou WX. Endophytes: a rich source of functional metabolites. Nat Prod Rep. 2001;18(4):448-459.
Crossref - Azevedo JL, Maccheroni Jr W, Pereira JO, de Araujo WL. Endophytic microorganisms: a review on insect control and recent advances on tropical plants. Electron J Biotechnol. 2000;3(1):15-16.
Crossref - Agbessenou A, Akutse KS, Yusuf AA, Khamis FM. The endophyte Trichoderma asperellum M2RT4 induces the systemic release of methyl salicylate and (Z)-jasmone in tomato plant affecting host location and herbivory of Tuta absoluta. Front Plant Sci. 2022;13:860309.
Crossref - Whipps JM, Lumsden RD. Commercial use of fungi as plant disease biological control agents: status and prospects. In: Butt TM, Jackson C, Magan N, ed. Fungi as biocontrol agents: progress, problems and potential. Wallingford UK. 2001:9-22.
Crossref - Ahmed AS, Sanchez CP, Candela ME. Evaluation of induction of systemic resistance in pepper plants (Capsicum annuum) to Phytophthora capsici using Trichoderma harzianum and its relation with capsidiol accumulation. Eur J Plant Pathol. 2000;106(9):817-824.
Crossref - Sempere F, Santamarina MP. In vitro biocontrol analysis of Alternaria alternata (Fr.) Keissler under different environmental conditions. Mycopathologia. 2007;163(3):183-190.
Crossref - Wilson CL, Wisniewski ME. Biological control of postharvest diseases of fruits and vegetables:an emerging technology. Annu Rev Phytopathol. 1989;27(1):425-441.
Crossref - Copping LG, Menn JJ. Biopesticides: a review of their action, applications and efficacy. Pest Manag Sci. 2000;56(8):651-676.
Crossref - Kushwaha M, Verma AK. Antagonistic activity of Trichoderma spp. (a bio-control agent) against isolated and identified plant pathogens. IJCBS. 2014;1(1):1-6
- Maynard DN. Underutilized and underexploited horticultural crops. Hortscience. 2008;43(1):279.
Crossref - Patel RK, Singh A, Yadav RK, Sharma YP, De LC. Enjoying sohiong an unexploited fruit in Meghalaya. Indian Horticulture. 2008;47-48
- Rymbai H, Roy AR, Deshmukh NA, et al. Analysis study on potential underutilized edible fruit genetic resources of the foothills track of Eastern Himalayas, India. Genet Resour Crop Evol. 2016;63(1):125-139.
Crossref - Fisher PJ, Petrini OSBC, Sutton BC. A comparative study of fungal endophytes in leaves, xylem and bark of Eucalyptus in Australia and England. Sydowia. 1993;45(2):338-345
- Suryanarayanan TS, Senthilarasu G, Muruganandam V. Endophytic fungi from Cuscuta reflexa and its host plants. Fungal Divers. 2000;4(11):117-123
- Barnett HL, Hunter BB, eds. Illustrated genera of Imperfect fungi, 3rd Ed. Minneapolis, Minn.: Burgess Publishing Company. 1972;225
- Domsch KH, Gams W, Anderson. Compendium of soil fungi. Academic press. 1980.
- White TJ, Bruns T, Lee SJWT, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, eds. PCR Protocols: A Guide to Methods and Applications, Academic Press, New York. 1990;18(1):
Crossref - Aneja KR. Experiments in microbiology, plant pathology and biotechnology. New Age International. 2007
- Patil MG, Pagare J, Patil SN, Sidhu AK. Extracellular enzymatic activities of endophytic fungi isolated from various medicinal plants. Int J Curr Microbiol Appl Sci. 2015;4(3):1035-1042
- Cattelan AJ, Hartel PG, Fuhrmann JJ. Screening for plant growth–promoting rhizobacteria to promote early soybean growth. Soil Sci Soc Am J. 1999;63(6):1670-1680.
Crossref - Smibert RM, Krieg NR. Phenotypic characterization. In Gerhardt P, Murray RGE, Wood WA, Krieg NR (eds) Methods of General and Molecular Bacteriology. American Society of Microbiology, Washington DC. 1994;607-654
- Tsuchida O, Yamagata Y, Ishizuka T, et al. An alkaline proteinase of an alkalophilic Bacillus sp. Curr Microbiol. 1986;14:7-12.
Crossref - Dubey S, Jain SK. Extracellular enzymatic profile of fungal deteriogens of historical Palace of Ujjain. Int J Curr Microbiol Appl Sci. 2015;4(5):122-132
- Lorck H. Production of hydrocyanic acid by bacteria. Physiol Plant. 1948;1(2):142-146.
Crossref - Skidmore AM, Dickinson CH. Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Trans Br Mycol Soc. 1976;66(1):57-64.
Crossref - Rajapakse RHS, Ratnasekera D, Abeysinghe S. Biopesticides Research: Current Status and Future Trends in Sri Lanka. In: Singh H, Sarma B, Keswani C, eds. Agriculturally Important Microorganisms. Springer, Singapore. 2016;219-234.
Crossref - Adeleke BS, Ayilara MS, Akinola SA, Babalola OO. Biocontrol mechanisms of endophytic fungi. Egypt J Biol Pest Control. 2022;32(1):46.
Crossref - Akram S, Ahmed A, He P, et al. Uniting the Role of Endophytic Fungi against Plant Pathogens and Their Interaction. J Fungi. 2023;9(1):72.
Crossref - Guthrie JL, Castle AJ. Chitinase production during interaction of Trichoderma aggressivum and Agaricus bisporus. Can J Microbiol. 2006;52(10):961-967.
Crossref - Almeida FBDR, Cerqueira FM, Silva RDN, Ulhoa CJ, Lima AL. Mycoparasitism studies of Trichoderma harzianum strains against Rhizoctonia solani: evaluation of coiling and hydrolytic enzyme production. Biotechnol lett. 2007;29:1189-1193
- Sanchez V, Rebolledo O, Picaso RM, et al. In vitro antagonism of Thielaviopsis paradoxa by Trichoderma longibrachiatum. Mycopathologia. 2007;163:49-58.
Crossref - Mamangkey J, Mendes LW, Harahap A, Briggs D, Kayacilar C. Endophytic bacteria and fungi from Indonesian medicinal plants with antibacterial, pathogenic antifungal and extracellular enzymes activities: a review. Int J Sci Technol Manag. 2022;3(1):245-255.
Crossref - Paris S, Wysong D, Debeaupuis JP, et al. Catalases of Aspergillus fumigatus. Infect immun. 2003;71(6):3551-3562.
Crossref - Singh P, Singh RK, Guo DJ, et al. Whole Genome Analysis of Sugarcane Root-Associated Endophyte Pseudomonas aeruginosa B18-A Plant Growth-Promoting Bacterium With Antagonistic Potential Against Sporisorium scitamineum. Front Microbiol. 2021;12:628376.
Crossref - Bhadra F, Gupta A, Vasundhara M, Reddy MS. Endophytic fungi: a potential source of industrial enzyme producers. 3 Biotech. 2022;12(4):86.
Crossref - Sarkar M, Tiru Z, Pal A, Mandal P. Antagonistic Role Of Fungal Biocontrol Agents For Sustain Able Management Of Rice Blight Caused By Curvularia Lunata. Appl Biol Res. 2022;24(2):191-202.
- Khan AL, Al-Harrasi A, Al-Rawahi A, et al. Endophytic Fungi from Frankincense Tree Improves Host Growth and Produces Extracellular Enzymes and Indole Acetic Acid. PLoS One. 2016;11(6):e0158207.
Crossref - Maccheroni W Jr, Araujo WL, Azevedo JL. Ambient pH-regulated enzime secretion in endophytic and pathogenic isolates of the fungal genus Colletotrichum. Sci Agric. 2004;61(3):298-302.
Crossref - Shubha J, Srinivas C. Diversity and extracellular enzymes of endophytic fungi associated with Cymbidium aloifolium L. Afr J Biotechnol. 2017;16(48):2248-2258.
- Hawar SN. Extracellular enzyme of endophytic fungi isolated from Ziziphus spina leaves as medicinal plant. Int J Biomate. 2022;2022(1):2135927.
Crossref - Zhao X, Hou D, Xu J, Wang K, Hu Z. Antagonistic activity of fungal strains against Fusarium crown rot. Plants. 2022;11(3):255.
Crossref - Muhorakeye MC, Namikoye ES, Khamis FM, Wanjohi W, Akutse KS. Biostimulant and antagonistic potential of endophytic fungi against Fusarium wilt pathogen of tomato Fusarium oxysporum f. sp. lycopersici. Sci Rep. 2024;14(1):15365.
Crossref - Vinale F, Sivasithamparam K, Ghisalberti EL, et al. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol Mol Plant Pathol. 2008;72(1-3):80-86.
Crossref - Verma M, Brar SK, Tyagi RD, Surampalli RN, Valero JR. Antagonistic fungi, Trichoderma spp.: panoply of biological control. Biochem Eng J. 2007;37(1):1-20.
Crossref - Jin L, Huang R, Zhang J, et al. Identification and Characterization of Endophytic Fungus DJE2023 Isolated from Banana (Musa sp. cv. Dajiao) with Potential for Biocontrol of Banana Fusarium Wilt. J Fungi. 2024;10(12):877.
Crossref - Zimand G, Elad Y, Chet I. Effect of Trichoderma harzianum on Botrytis cinerea pathogenicity. Phytopathology. 1996;86(11):1255-1260.
Crossref - Nagarajkumar M, Bhaskaran R, Velazhahan R. Involvement of secondary metabolites and extracellular lytic enzymes produced by Pseudomonas fluorescens in inhibition of Rhizoctonia solani, the rice sheath blight pathogen. Microbiol Res. 2004;159(1):73-81.
Crossref - Monda EO. Biological control of Fusarium wilt of tomato–a review. JTMB. 2002;1(1):74-78.
Crossref - Harrison MJ. Signaling in the arbuscular mycorrhizal symbiosis. Annu Rev Microbiol. 2005;59:19-42.
Crossref - Wang X, Radwan MM, Tarawneh AH, et al. Antifungal activity against plant pathogens of metabolites from the endophytic fungus Cladosporium cladosporioides. J Agric Food Chem. 2013;61(19):4551-4555.
Crossref - Bhat MK. Cellulases and related enzymes in biotechnology. Biotechnol Adv. 2000;18(5):355-383.
Crossref - Pandey A, Nigam P, Soccol CR, Soccol VT, Singh D, Mohan R. Advances in microbial amylases. Biotechnol Appl Bioc. 2000;31(2):135-152.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.