


ISSN: 0973-7510
E-ISSN: 2581-690X
Three lipase genes namely LK2, LK3 and LK5 were successfully sub-cloned into expression vector pET30a. The recombinant vectors were heterologically expressed into Escherichia coli BL21 (DE3) by 1 mM IPTG induced at 37°C. The protein with the size of 32, 31 and 28 kDa with correspond to LK2, LK3 and LK5 clones were over expressed following SDS-PAGE analysis. Further analysis to quantity the level of expression showed that the protein were over expressed at around 30, 44 and 21% of the total protein for LK2, LK3 and LK5 respectively. The lipolytic activity of the proteins by using lauric acid (C12) as substrate at 50°C, appeared that LK2 showed the highest activity among the other (1.49U/mg) followed by LK5 (1,10U/mg) and the lowest activity is LK3 (0.94U/mg).
Thermostable lipase, Heterologous expression, Lipolytic activity.
Lipase is one of hydrolase enzyme which has activity on the interface between water and organic solvent (Jaeger & Eggert, 2002). Lipase also shows many properties such as enantioselectivity, regioselectivity, and a broad of substrat specificity. The enzyme performs number activities such as esterification, transesterification, interesterification, acidolysis, aminolisys, alcoholysis, acylation and racemic resolution (Houde et al., 2004; Salihu and Alam, 2014; Sharma et al., 2001). Since of the above properties, the enzyme are widely used in various fields of industries, such as food, detergents, cosmetics, biomedicine, biopolymers, biosurfactant, biodiesel, agrochemical and pharmaceutical industries (Gupta et al., 2004; Houde et al., 2004; Jaeger and Eggert, 2002; Salihu and Alam, 2014).
Nowadays, thermostable lipases are required by many industries especially for industry with using high temperature process. Enzymatic reactions at high temperature could increase the reaction rate and solubility of the substrate, reduce contaminant and reduce viscosity at the medium (Leow et al., 2004). Therefore, searching of thermostable lipases from thermophilic microorganisms are still extensively been carried out (Madayanti et al., 2008; Widhiastuty et al., 2009; Febriani et al., 2010; Febriani et al., 2013).
Some researches were carried out to isolate thermostable lipase through cultivation of thermophilic microorganisms. A number of thermophilic microorganisms were isolated from many sources and appeared to produce thermostable lipases (Madayanti et al., 2008; Widhiastuty et al., 2009; Febriani et al., 2010; Febriani et al., 2013; Syihab et al., 2015). Another method was also developed to obtain lipase by isolating microbial genomes directly from natural or environmental samples, without cultivation, known as metagenome (Handelsman, 2004). The method was successfully performed to isolate lipase from thermogenic phase of compost (Nurhasanah et al., 2015) and other enzymes (Suhartia et al.,2014).
In previous paper, we reported five clones of lipase isolated from compost through metagenom approach (Nurhasanah et al., 2015). For further characterization in this paper, three of the genes were heterologically expressed in E. coli and the lipolytic activity was examined.
Strain, Vector and Culture Medium
Three sample of lipase genes were cloned from compost within E. coli TOP10 as host cell. E. coli BL21 (DE3) (F- ompT hsdSb (rB-mB-) gal dcm (DE3)) was used for expression. The plasmid for cloning vector was pJET1.2 (Fermentas), while pET-30a(+) (Invitrogen) were used as expression vector. Recombinant cells harbouring the plasmid were grown in LB medium (1% tryptone, 0.5% yeast extract, 1% NaCl) at 37oC for 16 hours. Plasmid isolation was performed using Qiagen kit (Fermentas).
Construction of Expression System
Two restriction sites were added into the lipase genes through amplification using a pair of primers designed to carry NdeI and SalI restriction sites. The PCR product was digested with NdeI and SalI and then ligated with T4 DNA ligase (Promega) into the NdeI and SalI linearized expression vector pET-30a(+). The recombinant plasmid was then introduced into E. coli BL21 (DE3).
Expression of Lipase Gene in E. coli
An overnight culture of transformant was diluted to 1: 100 Luria Bertani (LB) broth and subjected to further incubation at 37oC until the absorbance at 600 nm reached <“0.6. IPTG was added to the culture at final concentration of 1mM. After incubation at 37oC for 4 hours, the bacterial cells were harvested by centrifugation (8000 x g at 4oC for 20 min). The cells were resuspended in 25 mM Tris Buffer Saline (TBS) pH8. The cells were sonicated six times for 30 seconds each time with 30 second interval. The cell debris was removed by centrifugation (12000 x g at 4oC for 30 min). The supernatan was transferred to a 15 mL conical tube.
Gel Electrophoresis
SDS-PAGE was carried out, as described by Laemmli (1970) with Biorad-equipment. SDS-PAGE was performed with gels of 12% (w/v) of acrylamide according to manufacturer’s recommendations. Gels were stained for protein detection by a Comassie Blue procedure.
Determination of Lipolytic Activity and Protein Concentration
Quantitative determination of lipolytic activity was measured by spectrophotometric assay using p-nitrophenyllaurate as substrate (Lee, et al., 1999). All the measurement of lipolytic activities were conducted under standard conditions at 50°C; pH 8. One unit of lipase activity was defined as the amount of enzyme that releases 1 µmolof p-nitrophenol from p-NPL per min under the assay conditions. Protein concentration was determined by Bradford (1976) methods using bovine serum albumin as a standard.
Subcloning of Lipase Genes From Cloning to Expression Vector
The lipase genes (LK2, LK3, and LK5) carried by the recombinant plasmid (pJet-LK) were amplified in vitro by a pair of primers containing NdeI and SalI restriction sites (Table 1). The amplicons were subsequently ligated into pJet 1.2/blunt. The recombinant plasmid from each sample was isolated and restricted by NdeI and SalI restriction enzymes. The fragment containing the gene was ligated into pET-30a expression vector restricted by NdeI/SalI enzymes. The recombinant vectors were transformed into E. coli BL21 (DE3) as host cells. The insertion of the genes into pET-30a was confirmed by restriction analysis and nucleotide sequence (data not shown).
Table (1):
Primers used for amplification of lipase gene with additional of NdeI and SalI restriction sites.
No. |
Primer |
Nucleotide sequence(5ꞌ→3ꞌ) |
∑ basa |
Tm (oC) |
|---|---|---|---|---|
1 |
Feksp |
CAACATATGAACAAGAACAAAACCTTGCTCGCC |
33 |
61 |
2 |
Resksp |
AAAGTCGACGAGCCCCGCGTTCTT |
24 |
60 |
Expression of Recombinant Lipase
E. coli BL21 (DE3) harbouring recombinant plasmid was cultured and induced by addition of IPTG. The cells were harvested and sonicated. The lipases (LK2, LK3, and LK5) were expressed as soluble form of proteins. The cell free extracts were then analysed by SDS-PAGE. The result showed that LK2, LK3, and LK5 were expressed with molecular weight 34, 31, and 28 kDa respectively (Figure 1). The level of expression was seemed in variation. Densitometer analysis showed that LK2, LK3, and LK5 were expressed at the level around 30%, 44%, and 21% respectively from the total of crude extract protein (Figure 2).
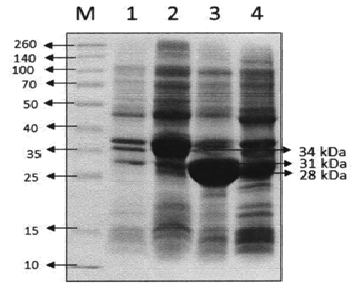 Fig. 1. SDS-PAGE of crude enzyme.-12% polyacrylamide gel was stained by Coomassie blue R-250 after electrophoresis. Lane 1. Broad range marker (Fermentas), Lane 2. Crude extract of cell harbouring pET-30a(+), Lane 3. Crude extract of cell harbouring pET-LK2; Lane 4. Crude extract of LK3 sample harbouring pET-LK3; Lane 5. Crude extract of LK5 sample harbouring pET-LK5.
Fig. 1. SDS-PAGE of crude enzyme.-12% polyacrylamide gel was stained by Coomassie blue R-250 after electrophoresis. Lane 1. Broad range marker (Fermentas), Lane 2. Crude extract of cell harbouring pET-30a(+), Lane 3. Crude extract of cell harbouring pET-LK2; Lane 4. Crude extract of LK3 sample harbouring pET-LK3; Lane 5. Crude extract of LK5 sample harbouring pET-LK5.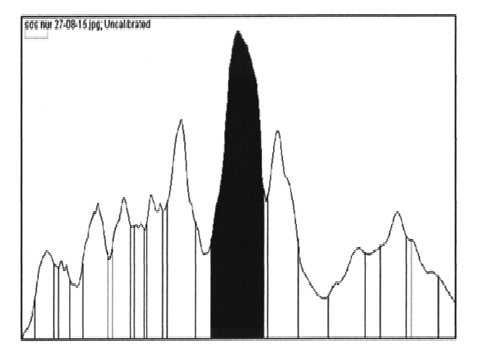
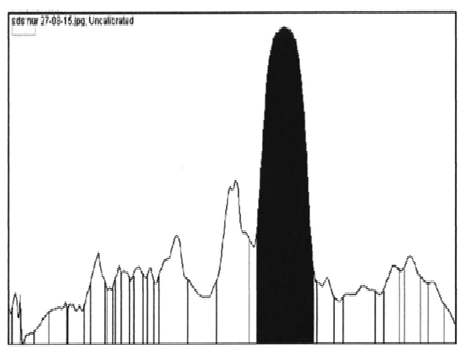
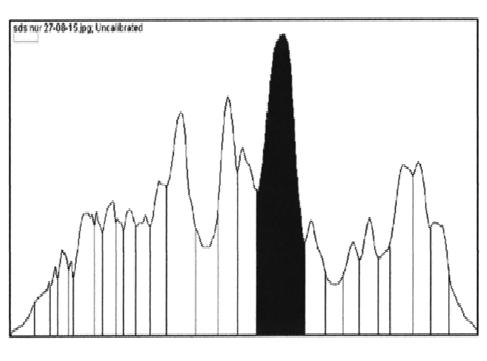
Fig.2. Quantitation of protein sample based on densitometric by using Image J. Software. Sample of LK2 (A); sample of LK3 (B); and sample of LK5 (C)
Lipolytic Activity of the Lipases
The lipolytic activity of the enzymes were measured by using p-nitrophenol laurate (C12) as substrat. All of the enzymes still showed lipolytic activity, however the level of activity are varying. LK5 appeared the most active enzyme with specific activity of 1.10 U/mg, followed by LK2 with specific activity at 1.49 U/mg, while LK3 is the less active enzyme with specific activity of 0.94 U/mg.
Three lipase genes isolated from natural sample (compost) were over expressed in E. coli and assayed the lipolytic activities. The gene sequence of the lipases are varied. LK2 and LK5 contain signal peptides, while LK3 is lack of the sequence. However, all of them are close to P. stutzeri lipase (Nurhasanah et al., 2015).
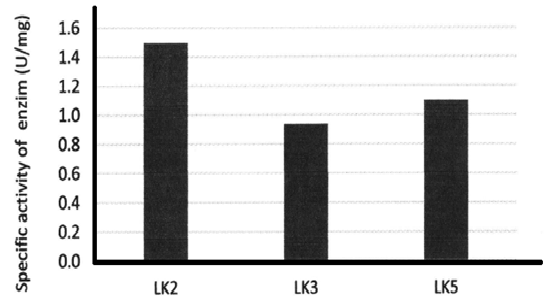 Fig. 3. Specific activity of Lipase samples
Fig. 3. Specific activity of Lipase samplesThe expression of LK3 is the highest (44%) compared to that LK2 (30%) and LK5 (21%). However the LK3 is less active enzyme compared to that LK2 and LK5. This is suggested that the LK3 was probably expressed in the form of more insoluble protein (inclusion body). Meanwhile LK2 and LK5 were expressed in soluble manner. Heterologous expression of some Pseudomonas lipases have been reported (Hirayama et al., 1993; Kojima et al., 2003; An et al., 2003). However most of the lipases were expressed in the form of inclusion bodies. Soluble expression of lipases from Pseudomonas were reported to be needed other proteins functioning for proper folding into its active form and efficient secretion (Jaeger and Eggert, 2002; Quyen et al., 1999).
Lower expression of LK2 and LK5 compared to the LK3 are probably due to activity of the lipase. Since the expressed lipases are more active so that the cells are less convenient for high expression of the enzyme. Furthermore LK2 and LK5 contained signal peptide, low expression of the enzyme might be due to some of the enzyme might be secreted.
- An, S., Kim, S., Choi, Y., Cho, Y., Joo, W., and Lee, Y. Cloning , Expression in Escherichia coli and Enzymatic Properties of a Lipase from Pseudomonassp . SW-3., Journal of Microbiology, 2003; 41(2), 95–101.
- Bradford, M.M. A Rapid and Sensitive Methods for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye-Binding, Analitycal Biochemistry, 1976; 72, 248-254.
- Febriani, Hertadi, R., Kahar, P., Akhmaloka, and Madayanti, F. Isolation and Purification of Thermostable Alkaline Lipase from Local Thermophilic Microorganism, Biosciences Biotechnology Research Asia, 2010; 7 (2) : 617-622.
- Febriani, Ihsanawati, Hertadi, R., Madayanti, F., and Akhmaloka. Thermostable Alkaline Lipase Isolated from ThermusAquaticus, International Journal of Integrative Biology, 2013; 14(2) : 104-112.
- Gupta, R., Gupta, N., and Rathi, P. Bacterial Lipases : An overview of Production, Purification and Biochemical Properties, Applied Microbiology and Biotechnology, 2004; 64, 763-781.
- Handelsman, J. Metagenomic : Application of Genomic to Uncultured Microorganisms, Microbiology and Molecular Biology Reviews, 2004; 68 : 4. 669-685.
- Hirayama, N. O., Yoshikawa, K., Nishioka, T., and Oda, J. Lipase from Pseudomonas aeruginosa Production in Escherichia coli and activation in vitro with a protein from the downstream gene, Eur. J. Biochem, 1993; 215, 239–246.
- Houde, A., Kademi, A., and Leblanc, D. Lipases and Their Industrial Applications: An Overview, Applied Biochemistry and Biotechnology, 2004; 118, 155–170.
- Jaeger, K.E., and Eggert, T. Lipases for Biotechnology, Current Opinion in Biotechnology, 2002; 13, 390-397.
- Kojima, Y., Kobayashi, M., and Shimizu, S. A Novel Lipase from Pseudomonas fluorescens HU380/ : Gene cloning , overproduction , renaturation-activation , two-step purification and characterization, Journal of Bioscience and Bioengineering, 2003; 96 (3), 242–249.
- Lee, D., Koh, Y., Kim, K., Kim, B., Choi, H., Kim, D., and Pyun, Y. Isolation and Characterization of a Thermophilic Lipase from Bacillus thermoleovorans ID-1, FEMS Microbiology Letters, 1999; 179, 393-400.
- Leow, T. C., Rahman, R.N.Z.R.A., Basri, M., and Salleh, A.B. High Level Expression of Thermostable Lipase from Geobacillus sp. Strain T1, Biosci. Biotech. Biochem., 2004; 68 (1), 96-103.
- Madayanti, F., ElVierra, B.V., Widhiatuty, M.P., and Akhmaloka. Characterization and Identification of Thermophilic Lipase Producing Bacteria from Thermogenic Compost. Journal of Pure and Applied Microbiology, 2008; 2(2) : 325-332.
- Nurhasanah, Nurbaiti, S., Warganegara, F.M., and Akhmaloka. Diversity of Gene Encoded Thermostable Lipase from Compost Based on Metagenome Analysis, International Journal of Integrative Biology, 2015; 16 (1), 7-12.
- Quyen, D. T. H. I., Schmidt-dannert, C., and Schmid, R. D. High-Level Formation of Active Pseudomonas cepacia Lipase after Heterologous Expression of the Encoding Gene and Its Modified Chaperone in Escherichia coli and Rapid In Vitro Refolding, Applied and Environmental Microbiology, 1999; 65 (2), 787–794.
- Salihu, A., and Alam, Md.Z. Thermostable Lipases : an Overview of Production, Purification and Characterization, Biosciences Biotechnology Research Asia, 2014; 11 (3), 1095-1107.
- Sharma, S., and Kanwar, S. S. Organic Solvent Tolerant Lipases and Applications, The Scientific World Journal, 2014, 1–16.
- Suhartia, S.S., Hertadia, R., Warganegaraa, F.M., and Akhmaloka. Diversity of Gene Encoded Crenarchaeal DNA Polimerase B from Natural Sample, International Journal of Integrative Biology, 2015; 15(2), 44-48.
- Syihab, S.F., Madayanti,F., and Akhmaloka: Isolation, Characterization and Identification of lipolytic Thermosphiles with Methanol Tolerance from Domestic Compost, Journal of Pure and Aoolied Microbiology, g (Spl Edn. 2), 2015; 385-390.
- Widhiastuty, M.P., Febriani, Yohandini, H., Moeis, M.R., Madayanti, F., and Akhmaloka, Characterization and Identification of Thermostable Alkaline Lipase Producing Bacteria From Hot Spring Around West Java, Journal of Pure and Applied Microbiology, 2009; 3 (1) : 27-40.
© The Author(s) 2017. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.