


ISSN: 0973-7510
E-ISSN: 2581-690X
Greater wax moth (GWM), Galleria mellonella (Lepidoptera: Pyralidae), is a destructive pest of honeybee hives. They lay eggs in the bee hives; and the hatched larvae feed on beeswax. The unique ability of GWM to digest the beeswax that contains ethene (CH2=CH2) bond has attracted attention because the same bond is also present in plastic polymers. Polymer-digesting ability, being a very uncommon among animals, we suspected a assistive role of gut bacteria of the insect in the process. The present study aimed to isolate and characterize potential polymer-degrading bacteria from GWM gut, following a culture-dependent approach, and to characterize them morphologically, biochemically, and with 16S rDNA sequences. We also intended to study polymer-degrading abilities of those bacteria, and check the presence of esterase, one of the most potent enzymes involved in plastic polymer degradation, using tributyrin agar test. We found nine bacterial isolates from GMW gut. Out of them, six showed positive results for plastic degradation to varying extents (19.3-31.2%), after 30 days of co-culture with plastic sheets. The esterase enzyme was found to be present in all of them in a qualitative test. Through
16S rDNA sequencing, four isolates were identified as gram-negative Serratia marcescens strains, one as gram-negative Ralstonia pickettii and one as gram-positive Bacillus cereus, some of which are known potential polymer degraders. Therefore, our hypothesis of involvement of gut bacteria in the digestion of polymers by Greater wax moth larvae was perhaps correct. There is possibility of exploiting the bacterial isolates for plastic-pollution remediation; after deeper and further experimentation.
Greater Wax Moth, Galleria mellonella, Polymer-degradation, Gut Bacteria, 16S rDNA Sequence
Greater wax moth (GWM; Galleria mellonella Linnaeus) is an important lepidopteran insect that has drawn researchers’ attention due to its rare ability to eat and digest beeswax. It was first reported in colonies of the Asian honeybee, A. cerana1 and considered an economically important pest of honeybees due to its destructive feeding habit.2 Almost 60-70 percent of economic losses are faced by beekeeping industries in developing countries because of this infamous pest.3 The adults of G. mellonella emerge in the bee hives and lays eggs. The hatched larvae bores into the combs and starts feeding on the wax of the comb and other materials such as pollen, dead bees, propolis, and pupal cases of bees.4 Severe greater wax moth infestation is encountered throughout the year, especially from May to September. Almost 90 percent of GWM infestations in India are reported in the colonies of Apis dorsata.5
Beeswax is a naturally occurring polymer composed of monoesters, fatty acids and other substances. Beeswax shares its structure with common plastic polymers; both having the same CH2=CH2 bonds.6 Recent studies indicate that two invertebrates, Indian meal moth (Plodia interpunctella) and GWM, have the capacity to degrade plastics.7,8 During biodegradation of plastic by these worms, their gut microbes presumably play key roles in plastic depolymerisation. The role of gut microbes in plastic depolymerisation by yellow meal worm was confirmed earlier by feeding the worms with gentamycin. Upon feeding with the antibiotic, the plastic depolymerisation was inhibited.9
Meanwhile, it is also known that some microorganisms are capable of utilizing polythene as a carbon source10 by adhering to the surface of the polythene film. On degradation, the long polymeric chains are cleaved to form short, low-molecular weight chains. Such microorganisms were reported to produce certain enzymes involved in this plastic degradation process.11 These enzymes are referred to as hydrolytic enzymes that functions in the presence of water and degrades plastic polymers into simpler units. Some of the enzymes include esterases, lipases, depolymerases, and PETases.12 The present study was undertaken to characterize the potential polymer-degrading bacteria with beneficial enzyme activity from the gut of GWM.
Collection and maintenance of GWM
The GWM larvae were collected from the Department of Entomology, Assam Agricultural University, Jorhat. The larvae were reared at the optimum temperature of 30°C, and were maintained for long term feeding with the natural diet, bees wax, obtained from commercial beekeepers. Usually large polyethylene terephthalate (PET) containers (HDPE, grade 34), 30 cm (length) × 15 cm (diameter), were used for rearing and maintenance. Tiny wholes were made in the lids for aeration.
Rearing of larvae on artificial feed and plastic
For our experiments, the larvae were reared in similar containers, and were fed on an artificial media that was prepared following previous reports.13 This mixture of artificial media was highly nutritious for the growth and development of the larvae.
Isolation of gut bacteria from GWM larvae
The standard procedure by Anand et al.14 was followed to isolate the GWM gut bacteria, with a few modifications. In a nutshell, ten healthy larvae were starved for 24 hours to ensure good feeding by them. The larvae were then surface sterilised with 0.1% (v/v) sodium hypochlorite (Himedia, India) or 70% (v/v) ethanol (Himedia), by wiping with soft brushes dipped in the respective reagent. After immediate rinsing in sterilized water, the larvae were carefully dissected with sterile forceps and micro scissors, to get the complete guts. The isolated guts were crushed in phosphate buffer using a sterile mortar and pestle to achieve uniform homogenization. These suspensions were then collected in micro centrifuge tubes containing 0.9% NaCl (Himedia). After homogenising the mixture, several dilutions, such as 10-1, 10-2, 10-3, 10-4, 10-5, 10-6 and 10-7, were made. Using the spread plate method, the homogenates were then plated on Nutrient agar media (NA; Himedia). Inoculated plates were incubated at 30°C and checked every 24 hours for of bacterial colonies, for up to 72 hours. The isolated individual colonies were maintained as pure cultures by streak plate method on fresh NA plates.
Morphological and biochemical characterization
Morphological and biochemical characterization were performed on pure cultured bacterial colonies. Observations were made on the basis of five colony characters: shape, margin, colour, elevation, and texture of the colony surface.15 This was done either visually or using a basic stereo microscope (SZX7, Olympus, USA), if needed. Gram-staining was done using the standard methodology. Several biochemical assays such as catalase reduction test, motility test, and citrate utilisation test were performed using available kits (Himedia, India). For carrying out these assays, the manufacturer’s instructions were followed.
Bacterial screening based on plastic degradation assay
Plastic sheets were prepared from commercial polythene bags. The sheets were washed with distilled water, followed by acetone and ethanol, in order to remove traces of dirt. Thereafter, those were rinsed in sterile water, dried and cut into nearly circular discs (of nearly 2 cm diameter), each weighing 15 mg. Then they were transferred to conical flasks containing 30 mL of carbon-free artificial media prepared with 0.1% (NH4)2SO4, 0.1% NaNO3, 0.1% K2HPO4, 0.1% KCl, 0.02% MgSO4, and 0.001% yeast extract (Himedia) in distilled water. The plastic sheets serve as sole carbon source for the microbes. Single bacterial colonies were picked and inoculated in Nutrient broth (NB) medium, and incubated at 30°C in 200 rpm in an incubator-shaker for 12 hours to obtain an optical density of 0.5 at 600 nm wave-length. One millilitre of this inoculum was added to 30 mL of the above carbon-free media containing the plastic discs. A control (E. coli) was maintained in similar conditions (30°C in 200 rpm) with plastic discs, to nullify any non-specific effect. These flasks were individually maintained for each treatment and left in a shaker for 30 days. After that period, the plastics discs were collected, washed, dried and re-weighed for final weight. Subsequently, the loss of weight of each disc was calculated.
Determination of esterase enzyme activity
Tributyrin agar (TBA) is a type of agar plate that’s commonly used to screen for lipolytic bacterial strains by looking for a clear zone around colonies. This clear zone indicates the presence of lipase or esterase activity. Qualitatively, a thicker zone indicates production of more enzymes and vice-versa. In our study, TBA was prepared from 5 g peptone, 3 g beef extract, 0.1 g CaCl2/MgCl2, 20 g agar and 10 mL tributyrin (Himedia) in 1000 mL dislilled water. Sterile TBA plates were inoculated with the bacterial samples by spot culture technique. These plates were then incubated at 30°C for 24-48 hours. The bacterial isolates were observed to grow in those plates, creating zones of hydrolysis surrounding the growth of each organism.16 The loss of opacity of the TBA media is an indication of occurrence of hydrolytic reaction yielding free fatty acids and soluble glycerol. Therefore, appearance of clear zones indicate activity of esterase enzyme produced by the bacterial colonies. Any such clear zone was not observed in the media inoculated with E. coli, which was considered as a negative control.
Genomic DNA isolation
The genomic DNA of the bacterial isolates was isolated using the protocol given by Wilson.17 A single bacterial colony picked from an overnight grown plate was inoculated in NB medium (Himedia) and incubated overnight at 30°C to obtain an optical density of approx. 1 at 600 nm. Two millilitres of the bacterial culture was centrifuged (12,000 rpm at 4°C, 2 min) and the supernatant was discarded. The pellet was then suspended in Tris (10 mmol/L)-EDTA (1 mmol/L) buffer (Himedia) followed by addition of 30 µL of 10% sodium dodecyl sulphate, 3 µL of 20 mg/mL Proteinase K and 3 µL RNase (1 µg/mL; Himedia, India). The tubes were mixed thoroughly and incubated for 1 hr at 37°C. Later, 80 µL of cetyltrimethylammonium bromide (CTAB)/NaCl solution (Himedia) was added to this suspension, and thoroughly mixed. It was followed by incubation at 65°C for 10 min. Equal volume of chloroform/isoamyl alcohol (Himedia) was added and mixed gently by inversion. The mixture was centrifuged in the same condition for 4-5 min. The aqueous phase was transferred to a fresh tube and the DNA was extracted using equal volume of phenol/chloroform/isoamyl alcohol (25:24:1) (Himedia) followed by spinning in a microcentrifuge at 12,000 rpm for 5 min. The supernatant was transferred to a fresh tube and double volume of ice-cold isopropanol (Himedia) was added to precipitate the nucleic acids. The tube was shaken back and forth until a whitish DNA precipitate appeared. The tube was then centrifuged (4°C at 12,000 rpm, 10 min) and the DNA pellet were washed once with 70% ethanol (Himedia) to remove any residual CTAB. The tubes were centrifuged again (12,000 rpm) at room temperature for 5 min to re-pellet the nucleic acid. The supernatant was discarded; the pellet was dried and dissolved in 100 µL of Tris-EDTA buffer (10 mM Tris, 0.1 mM EDTA; pH 8.0).
PCR amplification of 16S rRNA gene and sequencing of amplicons
A part of 16S rRNA gene of the bacteria was amplified using the universal primers
27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TACCTTGTTACGACTT-3′). The sequences of the primers were taken from Thakur et al.18 A PCR reaction of 40 µL composed of 20 µL PCR master mix (EmeraldAMP®Max, Takara, Japan), 4 µL each of the primers (1 µmol/L; Sigma-Aldrich, USA), 4 µL of the extracted DNA template (100 ng) and 8 µL of sterile water. The PCR profile was programmed with initial denaturation at 94°C for 7 min, denaturation at 94°C for 1 min, annealing at 51°C for 1 min, extension at 72°C for 1 min 30 s, and final extension at 72°C for 5 min; total 35 cycles. The amplified PCR products were detected on a 1.2% agarose gel using a 1 kb DNA ladder (Dye Plus, Takara, Japan) at 70 V for 45 min. Using the GenElute Gel Extraction Kit (Sigma-Aldrich), the PCR products of size nearing 1.5 kb were purified. The purified amplicons were sequenced by Sanger sequencing (Bioserve Technologies, India). Sequencing was performed using the aforementioned primers, and the final contigs were assembled with BioEdit 7.2 biological sequence alignment editor using the two counter-reads. The sequences were examined using BLASTn (www.ncbi.nlm.nih.gov) and probable bacterial genera and species were identified.
Phylogenetic analysis of the 16S rDNA sequence contigs
The sequence contigs obtained above were subjected to phylogenetic analysis in order to assign a taxonomic position to their respective source bacterial isolates. The NCBI database was used to identify the closest neighbours of the isolates followed by a comparative BLAST analysis (www.ncbi.nlm.nih.gov) of each contig. The software used for construction of the phylogenetic tree was Molecular Evolutionary Genetic Analysis (MEGA11), using the MUSCLE tool and the neighbour-joining method of Saitou and Nei.19
Isolation and characterization of GWM gut bacteria
When the GWM larvae were provided with sheets of plastic along with artificial diets, they were found to feed on plastic, creating holes in the plastic sheets (Figure 1). Several such larvae, each of them around 18-22 mm in length, were randomly selected for further studies. These larvae were surface sterilized and dissected under sterile conditions. Individual colonies were obtained in different dilutions of the homogenized contents of GWM gut, for different bacterial isolate. The pure cultured colonies were characterized morphologically based on five colony characters. Their characteristics were tabulated, and a total of nine bacterial isolates of considerable diversity were successfully isolated based on these characteristics (Table 1). The bacterial isolates were further subjected to Gram staining (Table 1). Out of the nine isolates, seven were found to be gram-negative and two gram-positive. The isolates, GMB1 and GMB9, were Gram-negative cocci-shaped bacteria; the isolates, GMB2, GMB4, GMB5, GMB6, GMB8 isolates were Gram-negative rod-shaped bacteria; the isolate, GMB3, was a Gram-positive cocci-shaped bacteria, and the isolate, GMB7, was identified as Gram-positive rod-shaped bacteria. The isolates were thereafter characterized based on catalase test, motility test and citrate utilization test. Table 1 presents the results of all the tests. In a nutshell, all the nine bacterial isolates showed positive response to catalase test. On the other hand, from the motility test, the isolates GMB1, GMB3 and GMB6, were found to be non-motile, while the isolates GMB2, GMB4, GMB5, GMB7, GMB8 and GMB9, were highly motile. All the isolates, except GMB3, showed positive results in the citrate utilization test.
Table (1):
Colony and cellular characteristics of the bacteria isolated from the gut of Greater wax moth
| Isolate No. | Laboratory code | Colony characteristics | Gram reaction | Shape test | Catalase test | Motility test | Citrate utilization test | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Form | Elevation | Margin | Colour | Surface | |||||||
| 1 | GMB1 | Circular | Raised | Entire | White | Smooth | Gram -ve | Cocci | +ve | Non-motile | +ve |
| 2 | GMB2 | Filamentous | Umbonate | Convex | Pink | Rough | Gram-ve | Rods | +ve | Motile | +ve |
| 3 | GMB3 | Circular | Raised | Entire | Creamy-white | Smooth | Gram +ve | Cocci | +ve | Non-
motile |
-ve |
| 4 | GMB4 | Filamentous | Umbo-nate | Convex | Red | Rough | Gram-ve | Rods | +ve | Motile | +ve |
| 5 | GMB5 | Rhizoid | Raised | Convex | Light pink | Rough | Gram-ve | Rods | +ve | Motile | +ve |
| 6 | GMB6 | Circular | Raised | Entire | White | Smooth | Gram-ve | Rods | +ve | Non-motile | +ve |
| 7 | GMB7 | Circular | Raised | Entire | Creamy-white | Smooth | Gram +ve | Rods | +ve | Motile | +ve |
| 8 | GMB8 | Circular | Raised | Convex | Dark pink | Smooth | Gram-ve | Rods | +ve | Motile | +ve |
| 9 | GMB9 | Irregular | Raised | Entire | Creamy-white | Smooth | Gram-ve | Cocci | Weakly +ve | Motile | +ve |
*+ve: positive, -ve: negative
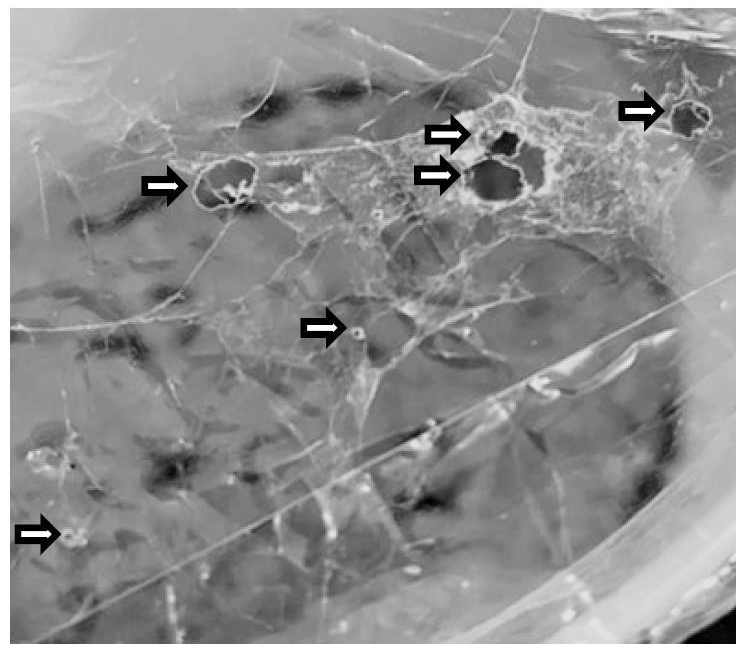
Figure 1. Holes made by larvae of Greater wax moth in a plastic sheet (shown with arrows)
Microbial degradation of plastic in laboratory
The weight of the plastic discs after subjecting to the microbial degradation in shaker culture, with respective bacterial isolates in artificial media for 30 days, were measured to detect any microbial degradation of the discs. It was observed that in case of GMB2, GMB4, GMB5, GMB6, GMB7 and GMB8, the plastic discs showed significant weight loss, albeit to a varying extent, after 30 days of incubation. This indicated considerable degradation of the plastic sheets by the action of the respective bacterial isolates present in the media (Table 2). Isolate GMB6 (Ralstonia pickettii) showed the least amount of PE degradation (2.90 ± 0.04 mg; 19.33 ± 0.248%) and isolate GMB7 (Bacillus cereus) showed the highest PE degradation (4.40 ± 0.031; 31.21 ± 0.183%). Three isolates, GMB1, GMB3 and GMB9 showed no evidence of biodegradation of the plastic polymers, as shown by no loss of the weight of the provided plastic sheets. E. coli, used as a negative control, as expected, did not show any considerable weight loss of the plastic sheets.
Table (2):
Determination of plastic degradation, esterase production and results of 16S rDNA sequence analysis of the nine gut bacterial isolates obtained from GWM gut
| Isolate | Absolute decrease in plastic disc weight (mg) | % decrease in plastic disc weight (%)* | Diameter of zone of clearance (cm) | BLASTn results of 16S rDNA sequences | |||
|---|---|---|---|---|---|---|---|
| Most similarity with (Genbank accession No.) | Maximum identity (%) | ||||||
| Control | 0 | 0.00 | No zone | – | – | ||
| GMB1 | 0.00 | 0.00 | No zone | – | – | ||
| GMB2 | 3.50 ± 0.035 | 22.15 ± 0.18 | 0.2 | Serratia marcescens strain XT218 (MT424800.1) | 99.54 | ||
| GMB3 | 0.00 | 0.00 | No zone | – | – | ||
| GMB4 | 3.60 ± 0.017 | 21.69 ± 0.11 | 0.35 | Serratia marcescens strain 16Sr (KJ729604.1) | 99.43 | ||
| GMB5 | 3.90 ± 0.029 | 22.67 ± 0.15 | 0.3 | Serratia marcescens JW-CZ2 (CP055161.1) | 99.7 | ||
| GMB6 | 2.90 ± 0.04 | 19.33 ± 0.25 | 0.25 | Ralstonia pickettii strain CHP10 (MT341804.1) | 100 | ||
| GMB7 | 4.40 ± 0.031 | 31.21 ± 0.18 | 0.6 | Bacillus cereus strain KF-3 (MF977355.1) | 99.89 | ||
| GMB8 | 3.20 ± 0.006 | 20.65 ± 0.04 | 0.1 | Serratia marcescens strain FY (CP053378.1) | 99.89 | ||
| GMB9 | 0.00 | 0.00 | No zone | – | – | ||
*values are average of three biological replicates ± SD with a statistical significance of p < 0.05
Determination of esterase enzyme production
The isolates, GMB2, GMB4, GMB5, GMB6, GMB7 and GMB8, were observed to show zones of clearance in TBA plates after incubation for one to two days (Figure 2). Nevertheless, the thickness of the zone was found to vary among the isolates. Isolates, GMB1, GMB3 and GMB9, however, did not show any clear zone. This indicates the hydrolysis of tributyrin by the esterase enzyme produced by the former group of isolates. On the other hand, E. coli, which was used as a negative control, also showed no zone of clearance in the TBA plates. The thickness of the zones of clearance was measured in order to have a quantitative and comparative observation for possible esterase production (Table 2).
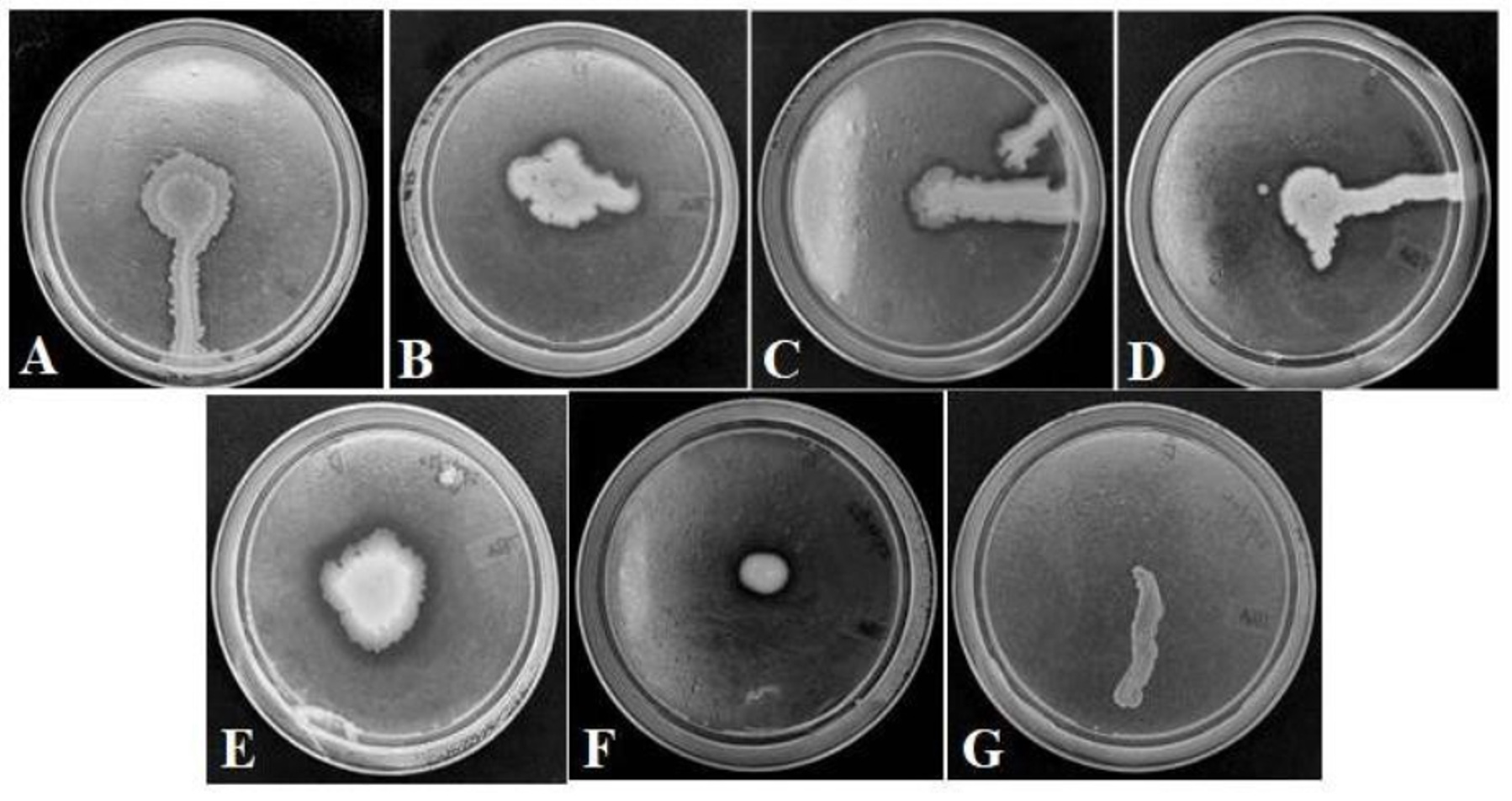
Figure 2. Esterase enzyme activities of the six bacterial isolates obtained from the gut of Greater wax moth. (A). GMB2 (B). GMB4 (C). GMB5 (D). GMB6 (E). GMB7 (F). GMB8 (G). Negative control (E. coli)
Sequence analyses of 16S rDNA
In all the six bacterial isolates, the 16S rDNA universal primer-pair could successfully amplify the expected size of the target DNA. The amplified partial 16S rDNA fragments were sequenced after eluting from agarose gels. The identity of the sequences was ascertained through analysis using the BLASTn programme of NCBI (Table 2). The Gram-negative rod-shaped motile isolates, GMB2, GMB4, GMB5 and GMB8, were confirmed to belong to the species Serratia marcescens. The Gram-negative rod-shaped non-motile isolate, GMB6, was identified as Ralstonia pickettii. Lastly, Gram-positive rod-shaped motile isolate, GMB7, was confirmed as Bacillus cereus strain. All these bacteria were potential esterase producers, thereby possibly aiding in microbial degradation of plastic.
Plastic production has been increasing exponentially for last more than 50 years; and PE and polypropylene contributes nearly 92% of the total plastic production. Bombelli et al.7 was the first to report PE biodegradation by the larva of G. mellonella. In their study, wax worms were observed to make holes on to the PE film at a rate of 2.2 ± 1.2 holes per worm per hour. In our study, we also observed that GWM larvae could survive, and also gain weight on polyethylene, albeit lower than their principal natural diet, the beeswax. Besides, there were visible holes made on the polyethylene sheet that was provided to them along with the artificial media. Esterase, cutinase, lipase, and PETase are the key enzymes that have lately become the subjects of interest so far as the enzymatic breakdown of plastics is concerned. We could not detect any mistake. The citation might perhaps be deleted, only keeping the number.20 All of these enzymes contribute to the breakdown of various types of polymer.21 In our study, out of the nine candidate bacterial isolates, six isolates, GMB2, GMB4, GMB5, GMB6, GMB7 and GMB8, showed the presence of the esterase enzyme. Interestingly, the same six isolates showed ability of plastic biodegradation after 30 days of co-culture. Therefore, this correlation indicates possible involvement of the esterase enzyme in the observed biodegradation of plastic and other similar polymers by the GWM larvae. Meanwhile, the same species of bacteria isolated and identified from our studies were reported to produce the same (esterase) enzyme that are capable of cleaving the ester bonds in complex high molecular weight polymers. For example, Bhardwaj et al.22 purified high molecular weight thermo-tolerant esterase from Serratia sp. isolated from Himachal Pradesh, India. Similarly, Ghati and Paul23 purified and characterized an extremely benzene-tolerant esterase from a Bacillus cereus strain isolated from West Bengal, India. A novel esterase from Ralstonia sp. was also reported by Quyen et al.24 The observed microbes in our study, i.e. the isolates of Serratia marcescens, Ralstonia picketti and Bacillus cereus, were found to be also present in several Lepidopteran guts. Some such recent reports include presence of Serratia marcescens in Lymantria dispar larva,25 Bacillus sp. in the larval gut of Discladispa armigera,26 and Ralstonia pickettii in the gut of armyworm27 etc. We ourselves had characterized 12 culturable bacterial species from GWM gut recently.28 However, to our knowledge, this is the first report of the presence of Ralstonia pickettii as a gut bacterium of GWM. The larvae of G. mellonella feed on some uncommon feeding materials such as paper, wood, polyethylene, occasionally polystyrene, and even muslin clothes. This uncommon and extreme eating habit may promote the growth of unique bacteria in its intestine having plastic-polymer degrading properties. Engel et al.29 elaborately reviewed about gut microbes of insects and stated that relatively fewer species of gut microbes are present in insects; and their diversity depends on the insect’s diet. They also concluded that many of the insect gut-bacteria are usually opportunists. Later, Lou et al.30 studied the effect of supplying co-diet (wheat bran, beeswax) and plastic diet (polyethylene, polystyrene) upon the physiological properties and the gut microbiomes of the GWM larvae. They found that larva fed on polyethelene and polystyrene showed lower community diversity of microbiomes in their gut. However, they also found that the gut microbiome of the polyethelene-fed larva did constitute of Serratia marcescens and Bacillus cereus; both of them together contributing to nearly 94% of the population. The results of our study led to a similar conclusion, with detectable presence of Serratia marcescens and Bacillus sp. in the gut of GWM larvae. Beeswax posses the ethylene hydrocarbon bond, CH2-CH2, as in case of polyethylene. Considering this fact, and based on the available information, it could be presumed that this bond could be the target of digestion of the plastic-polymer digesting bacteria. It was even reported that Bacillus sp., found in the gut of Plodia interpunctella wax moth, can depolymerise long chains of PE into water soluble low molecular weight products.31 There are many other reports of degradation of low-density PE film by various microorganisms like Bacillus cereus,32 Serratia marcescens,33 Enterobacter asburiae, Bacillus sp.34 and Ralstonia sp.35. Ryan et al.36 had demonstrated the ability of Ralstonia pickettii to breakdown several toxic polymers such as trichlorothylene, some aromatic hydrocarbons, chlorinated phenolic compounds etc. It is, indeed, a model organism for bioremediation studies and other potential applications.
In conclusion, our exploration of the gut-harboring bacteria of GWM leads to interesting and important discovery of useful bacterial species living in the gut of its larvae; which could find further useful application in tackling the menace of plastic pollution. However, further and more detailed research would be needed before a novel technology could be designed for the same.
ACKNOWLEDGMENTS
The authors are thankful to Richita Saikia and Ricky Raj Paswan for their assistance.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
BKB, RCB, BKS, AB, MKD and PD conceived the study. BKB, BD and GSHV designed the experiments. BD and GSHV executed the work. BKB obtained the funding. BD, GSHV and BKB wrote the manuscript. RCB, BKS, AB, MKS and PD edited the manuscript. GSHV and BKB approved the manuscript.
FUNDING
This study was supported by Assam Agricultural University, Jorhat, under the theme, “Generation of processes and products for the benefit of farming community and agripreneurs” (Grant No.: 7/297/DRA(A)/2022/1095-109).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Honeybee diseases and enemies in Asia: a practical guide. Rome, Italy: Food & Agriculture Organization. 1987. https://openknowledge.fao.org/server/api/core/bitstreams/3cef7eea-ad03-4a7e-80a4-e5c90f76af10/content
- Burges HD. Control of wax moths: physical, chemical and biological methods. Bee World. 1978;59(4):129-138.
Crossref - Kapil RP, Sihag RC. Wax moth and its control. Indian Bee J. 1983;45:47-49
- Hosamani V, Hanumanthaswamy BC, Kattimani KN, Kalibavi CM. Studies on biology of greater wax moth (Galleria mellonella L.). Int J Curr Microbiol Appl Sci. 2017;6(11):3811-3815.
Crossref - Mahindre DB. Handling rock bee colonies. Indian Bee J. 1983;45:72-73.
- Austin HP, Allen MD, Donohoe BS, et al. Characterization and engineering of a plastic-degrading aromatic polyesterase. Proc Natl Acad Sci U S A. 2018;115(19):E4350-E4357.
Crossref - Bombelli P, Howe CJ, Bertocchini F. Polyethylene bio-degradation by caterpillars of the wax moth Galleria mellonella. Curr Biol. 2017;27(8):R292-R293.
Crossref - Yang J, Yang Y, Wu WM, Zhao J, Jiang L. Evidence of polyethylene biodegradation by bacterial strains from the guts of plastic-eating waxworms. Environ Sci Technol. 2014;48(23):13776-13784.
Crossref - Yang SS, Brandon AM, Flanagan JCA, et al. Biodegradation of polystyrene wastes in yellow mealworms (larvae of Tenebrio molitor Linnaeus): factors affecting biodegradation rates and the ability of polystyrene-fed larvae to complete their life cycle. Chemosphere. 2018;191:979-989.
Crossref - Tokiwa Y, Suzuki T. Hydrolysis of polyesters by lipases. Nature. 1977;270(5632):76-78.
Crossref - Technical report on the PCR-DGGE analysis of bacterial and fungal soil communities. Tsukuba, Japan: National Institute for Agro-Environmental Sciences. 2010. https://www.naro.affrc.go.jp/archive/niaes/project/edna/edna_jp/manual_bacterium_e.pdf
- Pometto 3rd AL, Lee BT, Johnson KE. Production of an extracellular polyethylene-degrading enzyme (s) by Streptomyces species. Appl Environ Microbiol. 1992;58(2):731-733.
Crossref - Walia RK, Bajaj HK. Textbook of Introductory Plant Nematology (2nd Edition), New Delhi, India: Kalyani Publishers. 2014.
- Anand AAP, Vennison SJ, Sankar SG, et al. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J Insect Sci. 2010;10(1):1-20.
Crossref - Breakwell D, Woolverton C, MacDonald B, Smith K, Robison R. Colony Morphology Protocol. American Society for Microbiology. 2007:1-7.
- Palafox JC, Rivera-Chavira BE, Ramirez-Baca N, Manzanares-Papayanopoulos LI, Nevarez-Moorillon GV. Improved method for qualitative screening of lipolytic bacterial strains. MethodsX. 2018;5:68-74.
Crossref - Wilson K. Preparation of Genomic DNA from Bacteria. Curr Protoc Mol Biol. 2001;56(1):2.4.1-2.4.5.
Crossref - Thakur A, Dhammi P, Saini HS, Kaur S. Pathogenicity of bacteria isolated from gut of Spodoptera litura (Lepidoptera: Noctuidae) and fitness costs of insect associated with consumption of bacteria. J Invertebr Pathol. 2015;127:38-46.
Crossref - Saitou N, Nei M. The Neighbor-Joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406-425.
- Raoufi H, Taqwa S, Fagiryaar F. Enzymatic degradation of polyethylene and polyethylene terephthalate: a mini review. Am J Environ Climate. 2023;2(3):41-50.
Crossref - Yamashita Y, Tsurumi J, Ohno M, et al. Efficient molecular doping of polymeric semiconductors driven by anion exchange. Nature. 2019;572(7771):634-638.
Crossref - Bhardwaj KK, Kishen S, Mehta A, Sharma A, Gupta R. Purification of high molecular weight thermotolerant esterase from Serratia sp. and its characterization. 3Biotech. 2021;11(6):308.
Crossref - Ghati A, Paul G. Purification and characterization of a thermo-halophilic, alkali-stable and extremely benzene tolerant esterase from a thermo-halo tolerant Bacillus cereus strain AGP-03, isolated from ‘Bakreshwar’ hot spring, India. Process Biochem. 2015;50(5):771-781.
Crossref - Quyen DT, Dao TT, Nguyen SLT. A novel esterase from Ralstonia sp. M1: gene cloning, sequencing, high-level expression and characterization. Protein Expr Purif. 2007;51(2):133-140.
Crossref - Broderick NA, Raffa KF, Goodman RM, Handelsman J. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Appl Environ Microbiol. 2004;70(1):293-300.
Crossref - Thakur D, Bhuyan M, Majumder S, et al. Isolation, characterization, in-vitro antibiotic susceptibility and pesticide tolerance of gut bacteria from rice hispa, Dicladispa armigera (Olivier). Ind J Microbiol. 2005;45(3):217-221.
- He C, Nan X, Zhang Z, Li M. Composition and diversity analysis of the gut bacterial community of the Oriental armyworm, Mythimna separata, determined by culture-independent and culture-dependent techniques. J Insect Sci. 2013;13(1):165.
Crossref - Saikia SS, Borah BK, Baruah G, Rokozeno, Deka MK. Characterization of the gut microbes of greater wax moth (Galleria mellonella Linnaeus) shows presence of potential polymer degraders. Folia Microbiol (Praha). 2022;67(1):133-141.
Crossref - Engel P, Moran NA. The gut microbiota of insects-diversity in structure and function. FEMS Microbiol Rev. 2013;37(5):699-735.
Crossref - Lou Y, Ekaterina P, Yang SS, et al. Biodegradation of polyethylene and polystyrene by greater wax moth larvae (Galleria mellonella L.) and the effect of co-diet supplementation on the core gut microbiome. Environ Sci Technol. 2020;54(5):2821-2831.
Crossref - Yang Y, Yang J, Wu WM, et al. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: part 2. Role of gut microorganisms. Environ Sci Technol. 2015;49(20):12087-12093.
Crossref - Sudhakar M, Doble M, Murthy PS, Venkatesan R. Marine microbe-mediated biodegradation of low-and high-density polyethylenes. Int Biodeterior Biodegradation. 2008;61(3):203-213.
Crossref - Azeko ST, Etuk-Udo GA, Odusanya OS, Malatesta K, Anuku N, Soboyejo WO. Biodegradation of linear low-density polyethylene by Serratia marcescens subsp. marcescens and its cell free extracts. Waste Biomass Valorization. 2015;6(6):1047-1057.
Crossref - Yang Y, Yang J, Wu WM, et al. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: Part 1. Chemical and physical characterization and isotopic tests. Environ Sci Technol. 2015;49(20):12080-12086.
Crossref - Biki SP, Mahmud S, Akhter S, et al. Polyethylene degradation by Ralstonia sp. strain SKM2 and Bacillus sp. strain SM1 isolated from land fill soil site. Environ Technol Innov. 2021;22:101495.
Crossref - Ryan MP, Pembroke JT, Adley CC. Ralstonia pickettii in environmental biotechnology: potential and applications. J Appl Microbiol. 2007;103(4):754-764.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.