


ISSN: 0973-7510
E-ISSN: 2581-690X
Pseudomonas aeruginosa, a versatile Gram-negative bacterium, poses a serious threat due to its adeptness at developing resistance mechanisms. Metallo-beta-lactamases produced by P. aeruginosa are pivotal in conferring resistance to beta-lactam antibiotics, compromising treatment effectiveness. The bacterium’s proficiency in forming biofilms further enhances its persistence, especially in clinical settings. In this study conducted at Krishna Hospital and Medical Research Center, Karad, 205 clinical isolates of Pseudomonas aeruginosa were investigated for their metallo-beta-lactamase (MBL) production and biofilm-forming capabilities. The phenotypic detection of MBL production was carried out using the Imipenem-EDTA combination disc test, while biofilm formation was assessed through the microtiter plate technique. Genotypic detection involved conventional polymerase chain reaction targeting MBL and biofilm genes. Results showed that 28.29% of isolates produced MBL phenotypically, with 16.59% possessing MBL genes, notably the blaNDM-1 gene in 15.12% of isolates. Regarding biofilm production, 85.85% of isolates were biofilm producers, and biofilm-encoding genes were present in 80.49% of isolates. The most frequently encoded genes were algD (79.51%), pelF (58.0%), and pslD (46.34%). Comparing phenotypic and genotypic methods for MBL detection, a statistically significant average agreement was observed. While there was an increasing trend in biofilm genotypic positive patterns with stronger biofilm phenotypic patterns, the correlation was not statistically significant. The study concludes that every resistant clinical isolate should be screened for MBL and biofilm production using simple phenotypic tests, with confirmation by PCR if possible. This comprehensive analysis provides insights into the prevalence and genetic basis of MBL and biofilm in P. aeruginosa clinical isolates, contributing to strategies for combating antibiotic resistance.
Pseudomonas aeruginosa, MBL genes, Biofilm Genes, Phenotypic, Genotypic Methods
Pseudomonas aeruginosa is a pathogen that can lead to severe nosocomial and opportunistic infections among immunocompromised patients. It is referred to as the “Superbug,” and is becoming an increasingly dangerous infection due to its inherent and acquired resistance to existing antibiotics. It is listed in the WHO (World Health Organization) priority list of bacterial diseases under the “critical” category. The only carbapenem is a drug that can treat this pathogen. However, these days, carbapenem resistance is also developing quickly. The rapid spread and robust carbapenemase activity of metallo-β-lactamases (MBLs) type carbapenemases in Pseudomonas spp. poses a special concern.1-4
The number of Pseudomonas cases exhibiting MBL production has considerably increased during the past 20 years.5 Due to limited outer-membrane permeability, constitutive expression of different efflux pumps, and synthesis of antibiotic-inactivating enzymes, most isolates show inherent resistance to multiple antibiotics.6 Clinical P. aeruginosa isolates often have multiple β-lactamases coexisting, resulting in resistance to almost all β-lactam antibiotics.7
It was shown that P. aeruginosa harbor multiple forms of MBL genes: (i) Verona integron metallo-β-lactamase encoded by (VIM); (ii) imipenemase (IMP), iii) Seoul imipenemase (SIM); (iv) German imipenemase (GIM); (v) São Paulo metallo-β-lactamase (SPM); (vi) New Delhi metallo-β-lactamase (NDM) types. NDM-1 was initially detected in a patient in Sweden in 2008 who was admitted to the hospital in India and had returned home with the infection.8 Death rates of MBL-producing Pseudomonas aeruginosa vary from 70% to 90%.9,10
The capacity of P. aeruginosa to form biofilms, which shield them from environmental stresses, prevent phagocytosis, and ultimately provide them the capability for colonization and long-term persistence, also makes it difficult to cure these infections. Biofilm development,which adheres to surfaces using self-secreted extracellular polymeric substances (EPS), permits cumulative bacterial growth. Because the EPS is composed of polysaccharides, proteins, and nucleic acids, the organism can survive under unfavorable pH, humidity, and temperature conditions, as well as when exposed to antimicrobial treatments.11
The genetic makeup of strains, dietary requirements, and biofilm development phases all affect how important the EPS matrix is in relation to other factors. P. aeruginosa contains at least three polysaccharides (Psl, Pel, and alginate) that are crucial for the biofilm’s structural maintenance and antibiotic resistance.12 Weak, moderate, and potent biofilms are categorized architecturally as having varying capacities to protect bacteria against antimicrobial agents.11
Biofilms are a major contributor to the development of resistance to antibiotics because very dense populations of bacteria effectively transfer resistance and virulence genes horizontally.13 By exploiting this ability through quorum sensing, an effective cell-to-cell communication mechanism used by microbial populations of P. aeruginosa, can form highly organized biofilms.14
This study aimed to ascertain the prevalence of MBL and biofilm encoded genes among clinical isolates of P. aeruginosa at the Krishna Hospital and Research Centre, Karad.
An observational cross-sectional study was conducted for a period of one year in the Microbiology Department, Krishna Vishwa Vidhyapeeth (Deemed University), Karad. The study included non-repetitive clinical isolates of P. aeruginosa from the in-patient department (IPD) at Krishna Hospital and Medical Research Center, Karad.
Microbiological
Clinical sample collection, Identification & antimicrobial susceptibility test
The study was carried out after ethical clearance from the institutional ethical committee. A total of 205 non-duplicate clinical P. aeruginosa were isolated from Krishna Hospital and Medical Research Center Karad, between September 2021 to September 2022. According to established laboratory protocols, every isolate was collected in an extremely aseptic manner. The strain was identified, and antimicrobial susceptibility testing was carried out using the help of the VITEK-2 system (BioMerieux, France). The results were interpreted in accordance with the Clinical and Laboratory Standards Institute (CLSI, 2021) guidelines. American Type Culture Collection (ATCC) 27853 P. aeruginosa was used as the quality control strain.15
Phenotypic detection of MBL production
The Phenotypic detection of MBL production was done by Imipenem-EDTA combination disc test (CDT). On a Mueller Hinton agar plate, a 10 g Imipenem disc was positioned on a lawn culture of an Imipenem-resistant isolate at a distance of 20 mm from an Imipenem-EDTA disc. Plates were kept in an incubator at 37°C overnight, and the following day, the imipenem and imipenem-EDTA discs inhibition zones were compared. If the zone of inhibition with the imipenem-EDTA disc increased by ≥7 mm compared to imipenem disc, it was considered MBL positive.16
Phenotypic detection of biofilm production
The microtiter plate technique described by Stepanovic et al. was used to quantitatively evaluate biofilm formation.17 The OD (optical density) was measured at 630 nm using a microplate reader.18 Every trial was conducted three times, and a cut-off value (ODc) was established each time. Three standard deviations (SD) above the Mean OD of the negative control, i.e., ODc = Mean OD of the Negative Control + (3 SD of the Negative Control) was established. According to their capacity to form biofilms, the isolates were categories into four groups: non-biofilm producers (OD<ODc); weak producers (ODc ≤ OD < 2 ODc); moderate producers (2 ODc ≤ OD < 4 ODc); and strong producers (4 ODc ≤ OD).
Gene expression analysis
DNA extraction
DNA extraction was carried out from the clinical isolates, for the detection of MBL-producing IMP, NDM, and VIM genes, and biofilm-producing algD, pelF, and pslD genes. Briefly, all isolates of P. aeruginosa were processed for DNA extraction using HiPurA Bacterial Genomic DNA Miniprep Kit (Himedia, Mumbai) as directed by the manufacturer. The colonies of isolates were inoculated in the 10 ml nutrient broth and were incubated for 24 hr at 37°C by using shaker incubator. The cells were harvested from a 1.5 mL culture at 13,000 rpm for 2 minutes. The supernatant was removed, and the cell deposit was used for DNA extraction. Total DNA was extracted from the cells according to the manufacturer’s instructions of the extraction kit. Extracted DNA was examined for its concentration and purity by performing DNA gel electrophoresis (Technosource) on a agarose gel (Sigma-Aldrich) 1.5%, with 1 μl of elute to verify the presence of DNA in the extraction product. The DNA elute was stored at -20°C for further analysis.
Detection of MBL genes
Using primers specified in Table 1, all isolates underwent PCR (polymerase chain reaction) screening for the MBL-encoding genes “blaIMP, blaNDM, and blaVIM “. A PCR reaction with a master mix (19 μl) containing the Taq polymerase enzyme (0.5 μl), dNTPs (0.5 μl), Taq buffer (2 μl), Molecular biology grade water (15 μl), DNA extract (1 μl), and 0.5 μl of each forward and reverse primer (20 pmol) was run in a total volume of 20 μl. The three MBL genes were amplified in a Master cycler Eppendorf after completion of thermal program amplified products were kept at 4°C. BAA 2146 (Klebsiella pneumoniae) was used for blaNDM positive control, NCTC 13476 (Escherichia coli) for blaIMP positive control, a known laboratory strain positive for blaVIM, and ATCC 27853 (Pseudomonas aeruginosa) as a negative control. The cycling conditions for the MBL genes (blaIMP, blaNDM, and blaVIM) as per described in Table 2. The amplified products were analyzed by gel electrophoresis with a 1.5% agarose gel (Sigma-Aldrich) at 100 V in TAE (tris-acetate buffer) buffer with 1.5 μl ethidium bromide and were visualized and photographed under ultraviolet illumination.
Table (1):
Primers used for the amplification of the genes coding for MBL and biofilm exopolysaccharides among P. aeruginosa isolates
| Gene | Primer sequence(5’-3’) | Product size (bp) | Ref. |
|---|---|---|---|
| blaIMP | F-GGAATAGAGTGGCTTAAYTCTC | 232 | [19] |
| R-GGTTTAAYAAAAACAACCACC | |||
| blaNDM | F-GGTTTGGCGATCTGGTTTTC | 621 | [19] |
| R-CGGAATGGCTCATCACGATC | |||
| blaVIM | F-GATGGTGTTTGGTCGCATA | 390 | [19] |
| R-CGAATGCGCAGCACCAG | |||
| algD | F-CTACATCGAGACCGTCTGCC | 593 | [20] |
| R-GCATCAACGAACCGAGCATC | |||
| pelF | F-GAGGTCAGCTACATCCGTCG | 789 | [20] |
| R-TCATGCAATCTCCGTGGCTT | |||
| pslD | F- TGTACACCGTGCTCAACGAC | 369 | [20] |
| R-CTTCCGGCCCGATCTTCATC |
Detection of biofilm genes
All isolates underwent PCR screening for the biofilm genes “algD, pelF, and pslD” using primers specified in Table 1. PCR reaction with a master mix (19 μl) containing Taq polymerase enzyme (0.5 μl), dNTPs (0.5 μl), Taq buffer (2 μl), Molecular biology grade water (15 μl), DNA extract (0.1 μl), 0.5 μl of each forward and reverse primer (20 pmol) was run in a total volume of 20 μl. The three biofilm genes were amplified using a Mastercycler Eppendorf under the conditions provided in Table 2 and amplified products were kept at 4°C after completion of thermal program. The amplified products were analyzed by gel electrophoresis with a 1.5% agarose gel (Sigma-Aldrich) at 100 V in TAE (tris-acetate buffer) buffer with 1.5 μl ethidium bromide and were visualized and photographed under ultraviolet illumination.
Table (2):
Cycling conditions of PCR for different gene identification
Steps in PCR |
IMP |
NDM |
VIM |
algD |
pelF |
pslD |
|---|---|---|---|---|---|---|
Initial denaturation |
95°C 10 min |
95°C 10 min |
95°C 10 min |
95°C 10 min |
95°C 10 min |
95°C 10 min |
Denaturation |
95°C 1 min |
95°C 1 min |
95°C 1 min |
95°C 1 min |
95°C 1 min |
95°C 1 min |
Primer annealing |
52°C 1 min |
52°C 1 min |
45°C 1 min |
58°C 1 min |
58°C 1 min |
56°C 1 min |
Primer extension |
72°C 1 min |
72°C 1 min |
72°C 1 min |
72°C 1 min |
72°C 1 min |
72°C 1 min |
Final extension |
72°C 10 min |
72°C 10 min |
72°C 10 min |
72°C 10 min |
72°C 10 min |
72°C 10 min |
Holding temp. |
4°C ∞ |
4°C ∞ |
4°C ∞ |
4°C ∞ |
4°C ∞ |
4°C ∞ |
Cycles |
35 |
35 |
40 |
30 |
30 |
30 |
Statistical analysis
All information were entered in Microsoft Excel sheet & analyzed by using SPSS 28 version. The Kappa agreement was employed to measure the similarity between phenotypic and genotypic MBL forming ability and Chi-Square test was employed to compare between phenotypic and genotypic biofilm forming ability. Highly statistically significant (HS) was defined as p < 0.001, and non-statistically significant (NS) was defined as p ≥ 0.05.
Metallo-β-lactamase detection
Among 205 total isolates, phenotypic results of P. aeruginosa isolates estimated by the combination disk test, 58 (28.29%) of these isolates produced MBL, while 147 (71.71%) did not, as given in Table 3.
Table (3):
The relation between MBL phenotype and genotype patterns among P. aeruginosa
| Phenotypic no. (%) | Genotyping no. (%) | |
|---|---|---|
| Negative | Positive | |
| Negative 147 (71.71) | 142 (96.60) | 5 (3.40) |
| Positive 58 (28.29) | 29 (50.00) | 29 (50.00) |
| Total 205 | 171 (83.41) | 34 (16.59) |
A total of 34 (16.59%) isolates was found to have MBL genes. This includes 31(15.12%) isolates encoding blaNDM1, which was the most frequently encoded gene among those investigated. Figure 1 shows positive results for it.” followed by the 3 (1.46%) isolates encoding blaIMP and 1 (0.49%) isolate encode blaVIM genes in alone. In combination, a single isolate was discovered to be carrying multiple MBL genes, i.e. blaIMP and blaNDM1.
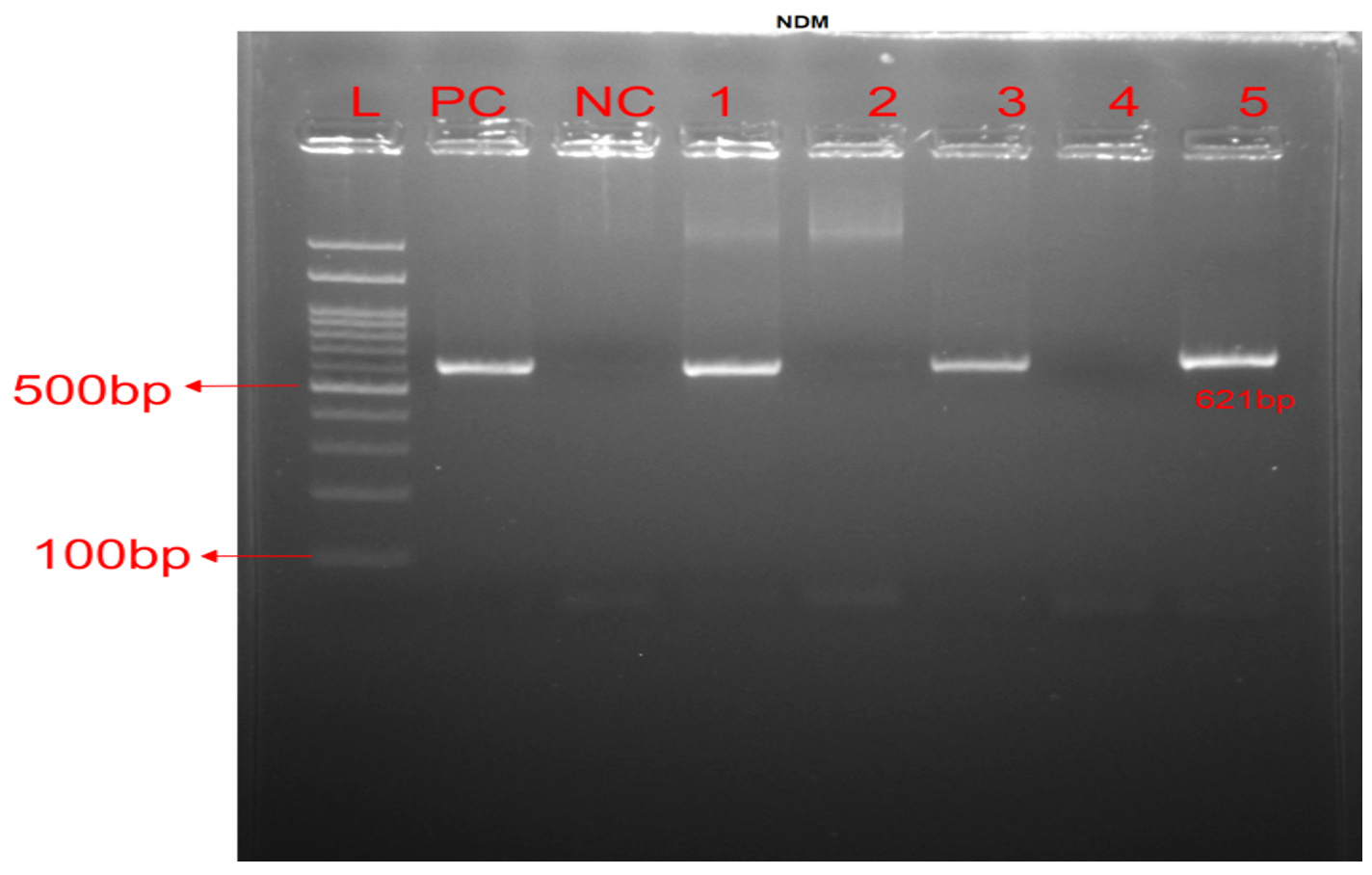
Figure 1. MBL amplification genes: PCR of blaNDM gene. Lane L: 100 bp DNA ladder. Lane PC: Positive control, Lane NC: Negative control, Lanes 1, 3 & 5 were positive, while Lanes 2 & 4 were negative for blaNDM at 621 bp
Biofilm production
Phenotype results of biofilm forming ability using microtiter plate, showed that 176 (85.85%) isolates were biofilm producers and 29 (14.15 %) isolates were non producers. Grading was 19 isolates (9.27%) were strong biofilm producers, 61 isolates (29.76%) were moderate biofilm producers, and 96 isolates (46.83%) were weak biofilm producers. Among 205 isolates, biofilm-encoding genes were present in 165 (80.49%) and 40 (19.51%) were negative for all genes (Table 4). The frequency of genes included 163 (79.51%) algD gene, 119 (58.0%) pelF gene, and 95 (46.34%) pslD gene. The isolates 84 (40.98%) presented all three genes “algD, pelF, pslD” as given in Figure 2, genes “algD, pelF” in 117 (57.07%), and genes “algD, pslD” in 94 (45.85%) isolates.
Table (4):
The relation between biofilm phenotype and genotype patterns among P. aeruginosa
| Biofilm Genotypic Pattern, no. (%) | Biofilm Phenotypic Pattern, no. (%) | |||
|---|---|---|---|---|
| Strong | Moderate | Weak | Non-biofilm | |
| algD+, pelF-, pslD- 36 (17.56) | 3 (8.33) | 12 (33.33) | 18 (50.00) | 3 (8.33) |
| algD+, pelF+, pslD- 33 (16.10) | 2 (6.06) | 17 (51.52) | 11 (33.33) | 3 (9.09) |
| algD+, pelF+, pslD+ 84 (40.98) | 9 (10.71) | 21 (25.00) | 44 (52.38) | 10 (11.90) |
| algD-, pelF+, pslD+ 1 (0.49) | 0 | 0 | 0 | 1 |
| algD-, pelF-, pslD+ 0 | 0 | 0 | 0 | 0 |
| algD-, pelF-, pslD- 40 (19.51) | 3 (7.50) | 10 (25.00) | 19 (47.50) | 8 (20.00) |
| algD+, pelF-, pslD+ 10 (4.88) | 2 (20.00) | 0 | 4 (40.00) | 4 (40.00) |
| algD-, pelF+, pslD- 1 (0.49) | 0 | 1 | 0 | 0 |
| Total 205 | 19 (9.27) | 61 (29.76) | 96 (46.83) | 29 (14.15) |
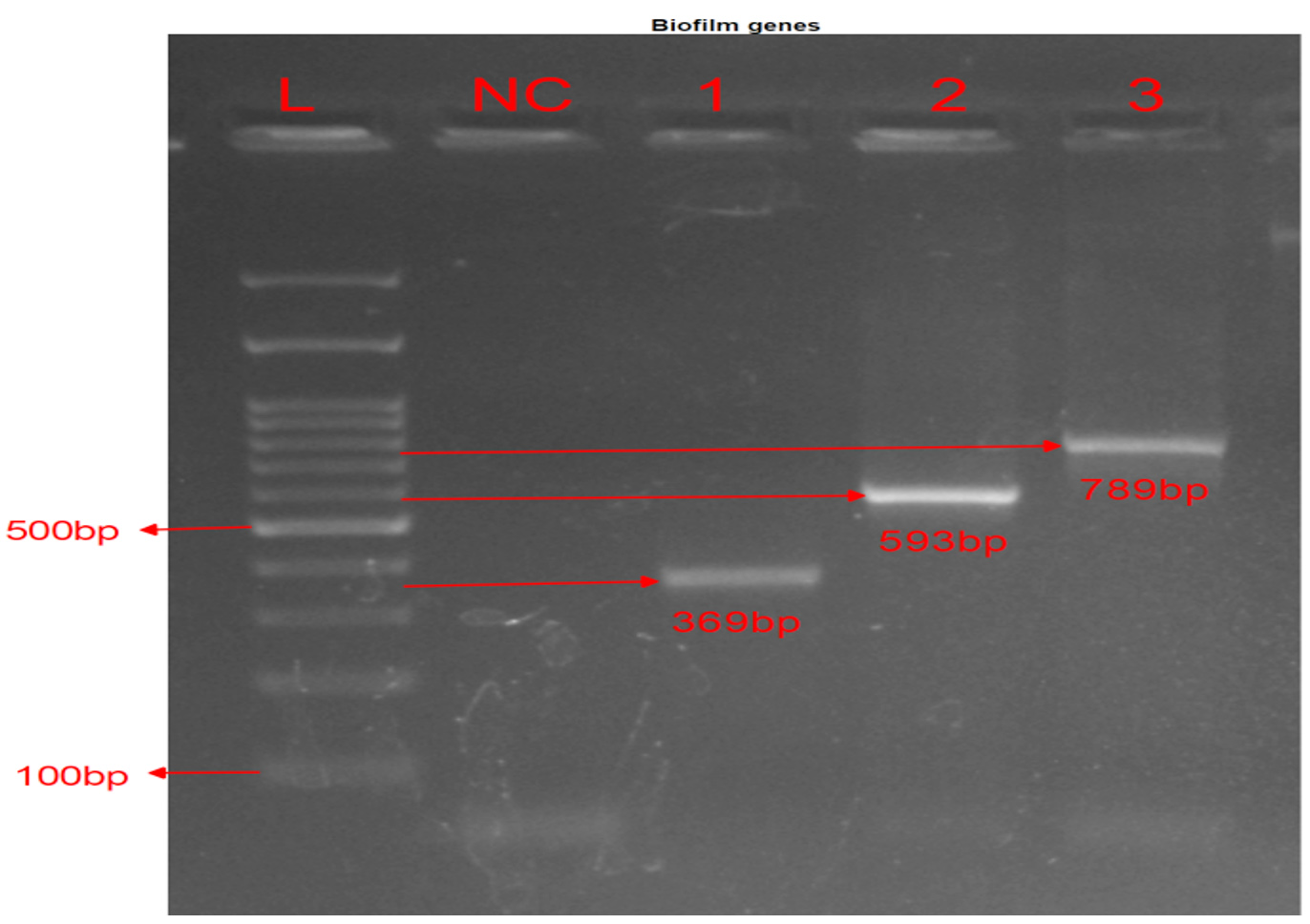
Figure 2. Biofilm amplification genes: PCR of biofilm genes. Lane L: 100 bp DNA ladder. Lane NC: Negative Control for all genes. Lane 1 were positive for pslD at 369 bp, Lane 2 for algD at 593 bp, and Lane 3 for pelF at 789 bp
Multi-drug resistant P. aeruginosa is a leading cause of hospital-acquired infections, capable of causing severe and life-threatening diseases. Carbapenems are the preferred drugs for resistant P. aeruginosa and ESBL-producing organisms. Resistance to carbapenems can occur due to reduced drug uptake, leading to imipenem/meropenem-resistant isolates.
Concerning phenotypic detection of MBL production in our study shows 28.29%. Some studies found it to be as high as 96.97% from Chandigarh, North India, Government Medical College Hospital; Department of Microbiology by Gupta et al. in 2008 and as low as 19.76% from the New Delhi, HIMSR, Jamia Hamdard, Department of Microbiology, India.21,22 A study conducted in Nepal by Sharma et al., using the phenotypic method, the frequency of MBL-producing P. aeruginosa isolates was observed to be 33.3% which is closer to our results.23
In our study, genotypic MBL production were 34 (16.59%) and among them blaNDM-1 15.12% constituted the most common, blaIMP and blaVIM were low 1.46%, and 0.49%, respectively. MBL genes were found in 18.9% of the isolates in a statewide study that were carried out to characterize 301 MBL-producing Pseudomonas species in 10 Indian medical centers.24 The occurrence of MBL genes in India ranges from 7-65 % among P. aeruginosa. In another investigation carried out in India, the MBL production rate was 20.2% among P. aeruginosa isolates, and blaVIM 19.04% was the most prevalent.25 In this investigation, 58 isolates tested positive by the MBL phenotypic test, 34 isolates contained MBL genes (blaVIM, blaIMP, and blaNDM-1), while the remaining isolates were negative, suggesting that there may be other MBL genes such as blaSIM, blaGIM, blaSPM and blaNDM variants (NDM-2 to NDM-7).26,27
We found a reduced prevalence of resistance genes of blaVIM and blaIMP in our setup and higher for blaNDM.28 Therefore, early identification of MBL synthesis in Pseudomonas aeruginosa will aid in limiting the transmission of multidrug resistance to other Gram-negative strains and assist in the proper management of infections caused by them. Antimicrobial resistance levels vary in studies due to differences in antibiotic usage patterns across locations. The prevalence of multidrug-resistant P. aeruginosa strains may result from the overuse of a range of antimicrobials for managing hospital infections or genetic mutations in the P. aeruginosa genome.
Phenotypic detection of biofilm formation in our study showed that among 205 isolates 176 (85.85%) biofilm producers, strong producers were 9.27%, moderate producers were 29.76%, weak producers were 46.83%, and non-producers were 29 (14.15%) of the isolates. Similar results were reported by El-sayed H, Fahmy Y,29 where 84.3% of the producers, among which 12.6% strong, 41.4% moderate, 30.4% weak, and 15.7% were non-biofilm producers. Also, Kamali et al.30 reported that 83.75% were biofilm producers, which were divided into 16.25% strong, 33.75% moderate, and 33.75% weak biofilm producers, while 16.25% biofilm non-producers and Mahmoud et al.31 reported that 70.4% biofilm producers, among them strong 14.8%, moderate 46.3%, 9.3% weak and 29.6% were non-biofilm producers. These results were slightly different from our results.
In this study, the genotypic detection of biofilm formation showed 165 (80.49%) biofilm producer and 40 (19.51%) non-producers. PCR amplification results for the algD, pelF, and pslD genes were present in 79.51%, 58.05%, and 46.34% of P. aeruginosa isolates respectively. In the study conducted by Rajabi et al.32 the frequency of these genes was 78.6%, 70.5%, and 36.6% slightly similar. In another research carried out by Banar et al.,33 the frequency of these genes among 57 P. aeruginosa strains was as follows: algD (100%), pelF (93%), and pslD (54.6%) which is higher than our results. The most frequent pattern in our study is 40.98% “algD+, pelF+, pslD+” this is in consistent with El-sayed H, Fahmy Y.29
P. aeruginosa isolates had statistically significant average agreement between detection of MBL production by phenotypic and genotypic method (kappa = 0.532). As biofilm production phenotypic positive pattern (Strong, Moderate, Weak & Non-biofilm) increases, biofilm genotypic positive pattern increases, however statistically not significant or not proved (chi square trend value= 1.435, p value= 0.231). Therefore, every resistant clinical isolate must be screened for MBL & biofilm production by using simple phenotypic tests and confirmed by PCR if possible.
Limitations of the study
The study did not investigate other MBL genes such as blaSIM, blaGIM, blaSPM, and blaNDM variants (NDM-2 to NDM-7). Additionally, it pointed out that the biofilm-producing capability of clinical isolates of Pseudomonas aeruginosa may vary in vivo. Therefore, the study has limitations, and further research through in vivo studies is needed to better understand these aspects.
It is critical to find out the antibiotic-resistant strains prevalence and their transmission channels. Surveillance data on P. aeruginosa‘s multi-drug resistance in hospitals is vital for health and economic benefits. Detecting Metallo beta-lactamase-producing strains early is essential for prompt treatment initiation. According to this study, a major issue that complicates and makes the treatment of P. aeruginosa infections difficult is the high incidence of biofilm-producing isolates. The algD, pslD, and pelF genes are essential for biofilm development, and our investigation found a strong association between these genes and biofilm formation.
ACKNOWLEDGMENTS
The authors express their sincere gratitude to all the faculties and laboratory assistants of the Department of Microbiology and Molecular Biology, Krishna Institute of Medical Sciences, Krishna Vishwa Vidyapeeth, Deemed University, Karad, Maharashtra, India. Authors are also thankful to the University Grants Commission, Krishna Vishwa Vidyapeeth, Deemed University, Karad, Maharashtra, India, for providing financial support for the research work.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
SAY and SKP designed the study. SAY extracted data. SRP, SAY and KDD performed data analysis. GSK and SRP supervised the study. SAY drafted the manuscript. SAY and SKP reviewed the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This research was funded by Krishna Vishwa Vidyapeeth, Deemed University, Karad, Maharashtra, India, with reference number KIMSDU/DR/633/2021.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
The study was approved by the Institutional Ethics Committee, Krishna Vishwa Vidyapeeth, Deemed University, Karad, Maharashtra, India, with reference number KIMSDU/IEC/06/2021.
INFORMED CONSENT
Written informed consent was obtained from the participants before enrolling in the study.
- Turkina MV, Vikstrom E. Bacteria-host crosstalk: sensing of the quorum in the context of Pseudomonas aeruginosa infections. J Innate Immun. 2019;11(3):263-279.
Crossref - Gale MJ, Maritato MS, Chen YL, Abdulateef SS, Ruiz JE. Pseudomonas aeruginosa cause an inflammatory mass in the nasopharynx of an immunocompromised HIV-infected patient: A mimic of malignancy. IDCases. 2015;2(2):40-43.
Crossref - Jurado-Martin I, Sainz-Mejias M, McClean S. Pseudomonas aeruginosa is an audacious pathogen with an adaptable arsenal of virulence factors. Int J Mol Sci. 2021;22(6):3128.
Crossref - Pragasam AK, Raghanivedha M, Anandan S, Veeraraghavan B. Characterization of Pseudomonas aeruginosa with discrepant carbapenem susceptibility profile. Ann Clin Microbiol Antimicrob. 2016;15(1):1-4.
Crossref - Nandy S, Das AK, Dudeja M. Prevalence of metallo beta lactamase in clinical isolates of Pseudomonas aeruginosa in a tertiary care hospital. Int J Community Med Public Health. 2015;2(4):566-569.
Crossref - Lambert PA. Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. Journal of the Royal Society of Medicine. 2002;95(Suppl 41):22.
- Upadhyay S, Sen MR, Bhattacharjee A. Presence of different beta-lactamase classes among clinical isolates of Pseudomonas aeruginosa expressing AmpC beta-lactamase enzyme. J Infect Dev Ctries. 2010;4(4):239-242.
Crossref - Raghunath D. New metallo-β-lactamase NDM-1. Indian J Med Res. 2010;132(5):478-481.
- Khajuria A, Praharaj AK, Kumar M, Grover N. Emergence of NDM-1 in the clinical isolates of Pseudomonas aeruginosa in India. J Clin Diagn Res. 2013;7(7):1328-1331.
Crossref - Wang W, Wang X. Prevalence of metallo-β-lactamase genes among Pseudomonas aeruginosa isolated from various clinical samples in China. Journal of Laboratory Medicine. 2020;44(4):197-203.
Crossref - Mirzahosseini HK, Hadadi-Fishani M, Morshedi K, Khaledi A. Meta-analysis of biofilm formation, antibiotic resistance patterns, and biofilm-related genes in Pseudomonas aeruginosa isolated from clinical samples. Microb Drug Resist. 2020;26(7):815-824.
Crossref - Wei Q, Ma LZ. Biofilm matrix and its regulation in Pseudomonas aeruginosa. Int J Mol Sci. 2013;14(10):20983-1005.
Crossref - Friedman L, Kolter R. Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J Bacteriol. 2004;186:4457-4465.
Crossref - Simpson JA, Smith SE, Dean RT. Alginate inhibits the uptake of Pseudomonas aeruginosa by macrophages. Microbiology. 1988;134(1):29-36.
Crossref - Weinstein MP, editor. Performance standards for antimicrobial susceptibility testing. Clinical and Laboratory Standards Institute; CLSI document M100-2021; Ed32.
- Prakash MR, Veena M, Sharma A, Basavaraj KN, Viswanath G. The Prospective Evaluation of Four Convenient Methods for Detecting MBLs in the Clinical Isolates. J Clin Diagn Res. 2012;6(7).
- Stepanovic S, Vukovic D, Hola V, et al. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS. 2007;115(8):891-899.
Crossref - Burton E, Yakandawala N, LoVetri K, Madhyastha MS. A microplate spectrofluorometric assay for bacterial biofilms. J Ind Microbiol Biotechnol. 2007;34(1):1-4.
Crossref - Mohanam L, Menon T. Molecular Detection of Extended Spectrum Beta-Lactamases in Clinical Isolates of Pseudomonas aeruginosa. J Pure Appl Microbiol. 2022;16(3):1736-1742.
Crossref - Banar M, Emaneini M, Satarzadeh M, et al. Evaluation of mannosidase and trypsin enzymes effects on biofilm production of Pseudomonas aeruginosa isolated from burn wound infections. PloS One. 2016;11(10):e0164622.
Crossref - Gupta V, Singla N, and Chander J. Use of two double disk synergy tests to detect metallo beta-lactamases in non-fermenters. Indian J Med Res. 2008;128(5):671-672.
- Sharma S, Devkota MD, Pokhrel BM, Banjara MR. Detection of blaNDM− 1, mcr-1 and MexB in multidrugresistant Pseudomonas aeruginosa isolated from clinical specimens in a tertiary care hospital of Nepal. BMC Microbiol. 2023;23(1):1-9.
Crossref - Sharma S, Devkota MD, Pokhrel BM, Banjara MR. Detection of blaNDM− 1, mcr-1 and MexB in multidrug-resistant Pseudomonas aeruginosa isolated from clinical specimens in a tertiary care hospital of Nepal. BMC Microbiol. 2023;23(1):1-9.
Crossref - Castanheira M, Bell JM, Turnidge JD, Mathai D, Jones RN. Carbapenem resistance among Pseudomonas aeruginosa strains from India: evidence for nationwide endemicity of multiple metallo-β-lactamase clones (VIM-2,-5,-6, and-11 and the newly characterized VIM-18). Antimicrob Agents Chemother. 2009;53(3):1225-1227.
Crossref - Sondakar A, Chunchanur SK, Rangaiah A, Shankar SM. Molecular characterization of metallo-beta-lactamase producers among carbapenem-resistant Pseudomonas aeruginosa isolated from cases of hospital-acquired infections in a tertiary care hospital, Bengaluru. Indian J Microbiol Res. 2020;7(2):212-217.
Crossref - Ismail SJ, Mahmoud SS. First detection of New Delhi metallo-β-lactamases variants (NDM-1, NDM-2) among Pseudomonas aeruginosa isolated from Iraqi hospitals. Iran J Microbiol. 2018;10(2):98.
- Hong DJ, Bae IK, Jang IH, Jeong SH, Kang HK, Lee K. Epidemiology and characteristics of metallo-β-lactamase-producing Pseudomonas aeruginosa. Infect Chemother. 2015;47(2):81-97.
Crossref - Pawar SK, Mohite ST, Datkhile KD, Patil MN, Kakade SV. Rising Threat of OXA-48 and other Carbapenemase Encoding Genes among Carbapenem Resistant Enterobacteriaceae in India. J Pure Appl Microbiol. 2020;14(3):1971-1925.
Crossref - El-sayed H, Fahmy Y. Correlation between biofilm formation and multidrug resistance in clinical isolates of Pseudomonas aeruginosa. Microb Infect Dis. 2021;2(3):541-549.
- Kamali E, Jamali A, Ardebili A, Ezadi F, Mohebbi A. Evaluation of antimicrobial resistance, biofilm-forming potential, and the presence of biofilm-related genes among clinical isolates of Pseudomonas aeruginosa. BMC Res Notes. 2020;13:1-6.
Crossref - Mahmoud MF, Fathy FM, Awad DM, Soliman MH. Biofilm formation and quorum sensing lasrgene of Pseudomonas aeruginosa isolated from patients with post-operative wound infections. Eur J Mol Clin Med. 2021;8(2):2177-2189.
- Rajabi H, Salimizand H, Khodabandehloo M, Fayyazi A, Ramazanzadeh R. Prevalence of algD, pslD, pelF, Ppgl, and PAPI-1 genes involved in biofilm formation in clinical Pseudomonas aeruginosa strains. BioMed Res Int. 2022;2022:1716087.
Crossref - Banar M, Emaneini M, Satarzadeh M, Abdellahi N, Beigverdi R, van Leeuwen WB, Jabalameli F. Evaluation of mannosidase and trypsin enzymes effects on biofilm production of Pseudomonas aeruginosa isolated from burn wound infections. PloS One. 2016;11(10):e0164622.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.