


Virus has the ability to cause health problems and even death in humans. Therefore, this review aims to assess the potential of metabolites derived from Bacillus species as viral protease inhibitors, specifically targeting Mpro/3CLpro and PLpro, in SARS-CoV-2 infection. During infection, SARS-CoV-2 enters host cells and initiates replication by translating viral proteases. The major protease (Mpro), also known as 3CLpro, and the papain-like protease (PLpro) are both encoded by SARS-CoV-2. Protease inhibitors (PIs) disrupt the formation of new viral particles by suppressing protease activity. Metabolites capable of acting as protease inhibitors found in Bacillus spp. include chondrillasterol, cholestane, trifluoroacetic acid, octadecenoic acid, stigmasterol, 9-octadecenoic acid, hexadecanoic acid, Macrolactin A, Subtilosin A, Leodoglucomide, Gramicidin S, and Tyrocidine A. Molecular docking analysis presented effective binding of these compounds to the active sites of Mpro or PLpro. The results showed that various compounds identified in Bacillus spp. had the potential to be developed as alternative drugs for combating SARS-CoV-2.
Virus, SARS-CoV-2, Bacillus, Proteases, Protease inhibitors
Virus has the ability to cause health problems and even death in humans. History shows that various viral outbreaks have led to widespread morbidity and mortality across different countries. Coronaviridae is popular among the virus families known to cause respiratory, neurological, and gastrointestinal problems in humans. This comprises the subfamily Coronavirinae,1 which includes the genera Alphacoronavirus (CoV), Betacoronavirus (CoV), Gammacoronavirus (CoV), and Deltacoronavirus (CoV).2 Furthermore, a coronavirus (CoV) contains an enveloped single-positive-strand RNA and is capable of infecting vertebrates, such as mammals and poultry birds. This virus group has been reported to initiate several pandemics over the years.3
Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) was first identified in November 2002 in the southern region of China. Subsequently, it experienced a rapid widespread transmission across 29 countries through international travel. By July 2003, the epidemic had resulted in 8,098 cases and 774 fatalities, representing a mortality rate of approximately 9.6%. In September 2012, Middle East Respiratory Syndrome Coronavirus (MERS-CoV) was diagnosed in a patient from Jeddah, Saudi Arabia, with symptoms of acute pneumonia and multi-organ failure. This virus continued to spread both in and outside the Middle East, with clusters of cases detected on several continents. On October 9, 2015, the total number of individuals infected by MERS-CoV reached 1,593, among which 568 deaths were recorded.3
In March 2020, the World Health Organization (WHO) declared the novel coronavirus disease 2019 (COVID-19) a global pandemic, with infections reported in over 100 countries. Patients with this disease commonly show symptoms such as fever, cough, fatigue, headache, hemoptysis, and diarrhea.4 The microorganism responsible for COVID-19 is SARS-CoV-2, a betacoronavirus5 containing a positive-sense single-stranded RNA genome of approximately 32 kilobases in length. SARS-CoV-2 ranges from a size of 80 to 160 nm, and it can cause severe pneumonia.5
On April 8, 2022, WHO reported a total of 493,392,853 confirmed cases of COVID-19 worldwide, with 6,165,833 deaths. Europe has recorded the highest number of cases, followed by America and Southeast Asia. Moreover, several strains of SARS-CoV-2 including B.1.1.7, B.1.351, P.1,6 and the omicron variant B.1.1.529,7 have been identified. B.1.1.7 was first detected in the UK in October 2020, while B.1.351 and P.1 were first identified in South Africa and Brazil, respectively.6 The earliest case of the omicron variant was announced in November 2021 from South Africa.7
Significant advancements have been made in medical research and therapeutics since the onset of the global pandemic in 2020. These include Remdesivir, an RNA polymerase inhibitor, endorsed by the Food and Drug Administration (FDA) for COVID-19 treatment.8 The continuous evolution of virus necessitates the development of additional antiviral agents. Despite several vaccines and drug candidates being in clinical trials, novel antivirals and antibodies need to be designed to address newly identified variants for effective control of the pandemic.9
The pathogenesis of SARS-CoV-2 infection essentially depends on viral or host proteases, which play crucial roles in viral replication, protein maturation, and disease progression.10 Specifically, papain-like protease (PLpro) and major protease (Mpro/3CLpro) are essential for viral replication and development. Therefore, restraint of viral protease activity using inhibitors presents a promising therapeutic method for COVID-19. Protease inhibitors function by disrupting viral replication through the prevention of polyprotein cleavage. These can be small molecules, proteins, or peptides that target specific protease enzymes.11 Numerous metabolites with potential viral protease inhibitory properties have been isolated from various sources, including plants,12 algae,13 bacteria, and fungi.14 Research showed that metabolites derived from Bacillus spp. manifested antiviral.15,16 and protease inhibitory activity.17 Bacillus spp. are ubiquitous bacteria found in soil, sediment, and marine environments,18 known for being easily culturable as well as capable of producing spores and different metabolites.19,20 Therefore, this review aims to explore the bioactivity of Bacillus spp. metabolites by focusing on the potential to serve as viral protease inhibitors with high specificity against PLpro and Mpro/3CLpro in SARS-CoV-2 infection.
Mechanism of SARS-CoV-2 infection
SARS-CoV-2 is known to enter host cells using viral spike (S) proteins which consist of subunits S1 and S2. The subunit S1 is responsible for binding to the angiotensin-converting enzyme 2 (ACE2) receptor on the cell surface, and S2 facilitates fusion with the host cell membrane,21 while the whole spike proteins may join heparan sulfate proteoglycans (HSPGs). Additionally, extracellular host cell proteases such as trypsin, factor Xa, and elastin assist in the activation of viral particles and the attachment process.10
Following attachment, the virus penetrates host cells and replicates its genetic material through the help of proteases. Among other roles, proteases catalyze the cleavage of specific peptide bonds in viral polyprotein precursors, a process essential for the production of functional components and the assembly of viral particles.22 Subsequently, viral components are packaged into a complete virus in the endoplasmic reticulum, Golgi apparatus, and cytosol,10 and this infection mechanism is presented in Figure 1.
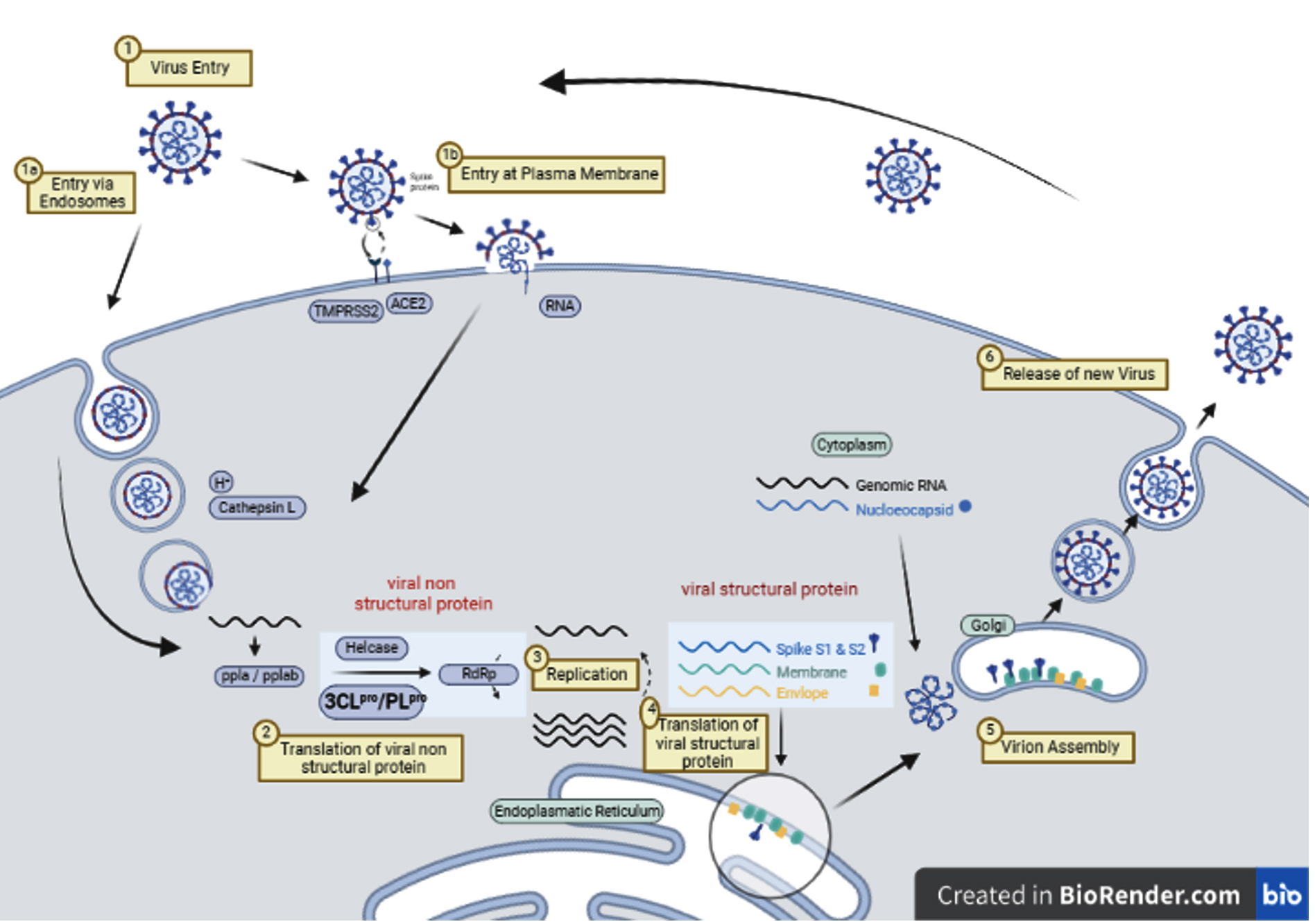
Figure 1. Mechanism of Infection of SARS-CoV-210 modified and redrawn using BioRender (https://www.biorender.com/)
SARS-CoV-2 proteases
Two important proteases encoded by the genetic material of SARS-CoV-2 in the primary translation region include Mpro and PLpro. Both play a fundamental role in the processing of polyproteins synthesized from viral RNA (Figure 2).
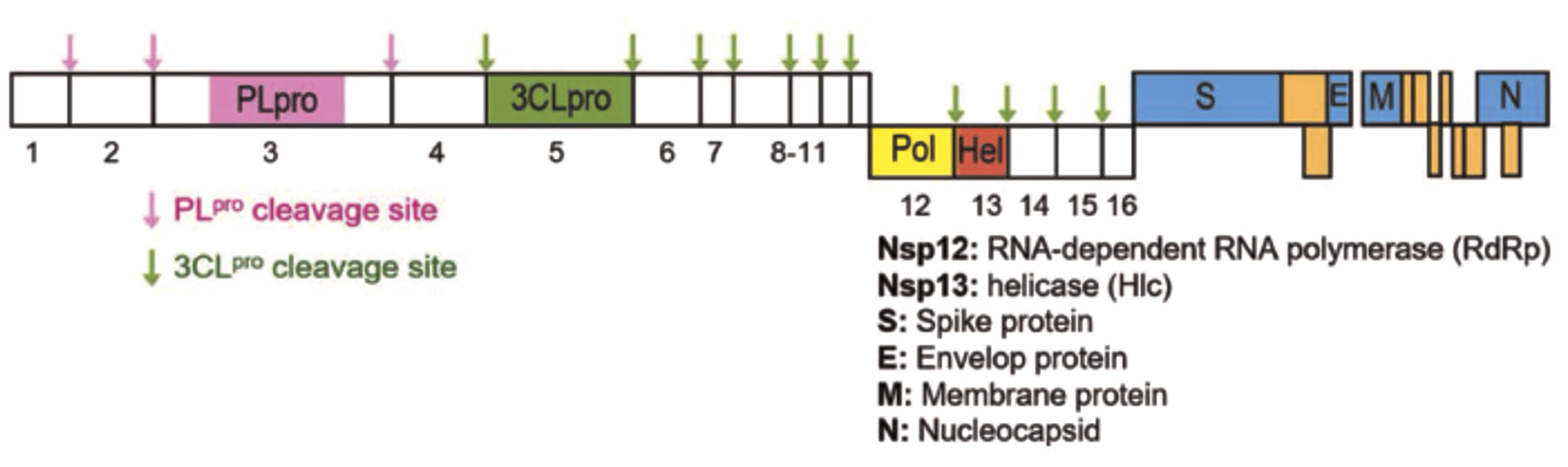
Figure 2. The SARS-CoV polyproteins are depicted in a schematic with two viral protease cleavage sites. The immature polyproteins are cleaved into non-structural proteins (designated 1-16) by the viral proteases PLpro and 3CLpro. SARS-CoV PLpro cut sites are shown by pink arrows, while SARS-CoV 3CLpro cleavage sites are indicated by green arrows29
Mpro cleaves a total of 12 non-structural proteins, including nsp4-nsp16, which comprise essential proteins and enzymes such as RNA-dependent RNA polymerase (nsp12) and helicase (nsp13). Furthermore, it manifests enzyme activity at many cleavage sites on the gigantic 1ab polyprotein (clone 1ab, about 790 kDa).10 The predominant cleavage site configuration observed is Leu-Gln↓ (Ser, Ala, and Gly), with the downward arrow signifying the specific cleavage site.23 SARS-CoV-2 Mpro protein has a molecular weight of 67.6 kDa and homophilic cysteine characteristics, showing a tendency to form clusters similar to Mpro. This protein produces a dimeric structure, with each monomer unit being composed of an N-terminal catalytic domain and a C-terminal region. Additionally, each monomer comprised three distinct regions, namely spaces I, II, and III comprising protons numbered 8 – 101, 102 – 184, and 201 – 303, respectively. Two critical amino acid residues, including histidine (H41) and cysteine (C145), participate in the catalytic activity manifested at the active site.
PLpro also cleaves the N-terminal of the polyprotein transcript to generate nsp1, nsp2, and nsp3, which are crucial for facilitating viral replication.10 Three essential amino acid residues, cysteine (C111), histidine (H272), and aspartic acid (D286), contribute to the catalytic activity of PLpro.
Mpro and PLpro from SARS-CoV-2 and SARS-CoV possess significant sequence similarities. Consequently, potential cross-inhibitory effects tend to occur between inhibitors targeting these two proteases.
Mechanism of action of protease inhibitors
Protease inhibitors (PIs) play a crucial role in regulating proteolytic processes across various biological systems24 and can be used to treat or prevent viral infection. Those developed as antivirals over the years include saquinavir, ritonavir, indinavir, nelfinavir, and amprenavir.11
Many minor protease inhibitors derived from various sources, including plants, organisms, and microbes has been identified.24 Protease inhibitors contain varying restraint components and target different types of proteases, such as serine, cysteine, aspartyl, and matrix metalloproteinase (MMP). These are often classified as competitive inhibitors based on effective binding to the active site of the target proteases, thereby obstructing access to the substrate.
The mechanism of protease inhibition includes reaction loop attachment to the active site and binding to a part of the protein structure, leading to serine protease restriction. Additionally, cystatins are a protein superfamily that serves as inhibitors by blocking papain-like cysteine proteases. Cytokine retention results from the integration of the wedge-shaped face of inhibitors, comprising the protein N-terminal and two clamping rings, into the dynamic V-shaped cleft of cysteine proteases. Moreover, the N-terminal of four human tissue inhibitors of metalloproteases (TIMPs) binds to P1-P3 pockets of proteases, thereby sequestering the catalytic Zn2+ particle and preventing access to the water molecule at the active site. The second loop of TIMPs links both P3 and P2 pockets with the N-terminal region of MMP.30
Potential metabolites from Bacillus spp. as SARS-CoV-2 protease inhibitors
Bacillus spp. produce various bioactive compounds, including lipopeptide antimicrobial substances such as surfactin, iturin, fengycin, and subtilisin. These lipopeptide substances prevent the replication of Newcastle disease virus (NDV) and infectious bursitis virus (IBDV).4 Surfactin as a lipopeptide, with antiviral activity against porcine diarrhea virus (PEDV) or gastroenteritis virus transmissible in pigs (TGEV) has been reported.31 Another research also found that it successfully inhibited cell and viral membrane fusion.23
The antiviral mechanisms of lipopeptide antimicrobial substances include acting as protease inhibitors, a characteristic observed in. surfactin.23 Molecular docking analysis showed that surfactin, bacillomycin, and iturin could bind to the spike glycoprotein of SARS-CoV-2 with strong affinity, suggesting the potential possessed as therapeutic agents against COVID-19.32 An additional mechanism of lipopeptide antimicrobial substances as antivirals is the inhibition of the membrane fusion process, preventing viral morphogenesis.33
Bacillus spp. are known to also produce alkaloids, steroids, saturated fatty acids, and lipids. The methanol extract from Bacillus subtilis often contains octadecenoic acid, hexadecanoic acid, aspidospermidine, stearic acid, and phenol. Paenibacillus dendritiformis methanol extract comprises 2-hydroxyl-1,3-propanedyl ester, 4-tetrahydro-6-methyl, 9-Octadecenoic acid(z), cholestan-3-ol,2-methylene (3a,5a), oxalic acid, stearic acid, chondrillasterol, naphthalene, 1,2,3-propanoic acid, trifluoracetic acid, and stigmasterol. The methanol extract of Brevibacillus formosus contains 1,3,5-trioxane, cyclobutane, dasycarpidan, and phenol.17
The numerous compounds derived from bioactive components of Bacillus spp. were investigated using molecular docking analysis to identify the inhibition potential manifested against Mpro, the primary protease in SARS-CoV-2. These included chondrillasterol, cholestan, trifluoroacetic acid, octadecenoic acid, stigmasterol, 9-octadecenoic acid, and hexadecanoic acid. Stigmasterol (-8.3 kcal/mol), hexadecanoic acid (-6.9 kcal/mol), and chondrillasterol (-7.9 kcal/mol) are expected to fit into Mpro binding site.17 Compared to the docked complexes of stigmasterol and chondrillasterol, hexadecanoic acid-Mpro is the most stable complex form, thereby suggesting hexadecanoic acid as a form of suitable inhibitors of Mpro in COVID-19.
Research reported the potential of Macrolactin A produced by Bacillus amyloliquefaciens to serve as a drug candidate for combating COVID-19. The results of molecular docking analysis showed that Macrolactin A could bind to the active site of SARS-CoV-2 Mpro, initiating catalytic activity inhibition.25 Moreover, Tables 1 and 2 present the entire binding affinities and structures of bioactive compounds from Bacillus spp. with the potential as Mpro and PLpro inhibitors.
Table (1):
Bioactive compounds from Bacillus spp. potential as Mpro and PLpro inhibitors
No. |
Compounds |
Source |
Target |
Binding affinities |
Reference |
|---|---|---|---|---|---|
1. |
Chondrillasterol |
Paenibacillus dendritiformis |
Mpro |
−7.9 kcal/mol |
17 |
2. |
Cholestane |
Paenibacillus dendritiformis |
Mpro |
−7.8 kcal/mol |
17 |
3. |
Trifluoroacetic acid |
Paenibacillus dendritiformis |
Mpro |
−7.7 kcal/mol |
17 |
4. |
Octadecenoic-acid |
Bacillus subtilis |
Mpro |
−9.9 kcal/mol |
17 |
5. |
Stigmasterol |
Paenibacillus dendritiformis |
Mpro |
−8.3 kcal/mol |
17 |
6. |
9 octadecenoic-acid |
Paenibacillus dendritiformis |
Mpro |
−9.7 kcal/mol |
17 |
7. |
Hexadecanoic acid |
Bacillus subtilis |
Mpro |
−6.9 kcal/mol |
17 |
8. |
Macrolaktin A |
Bacillus subtilis |
Mpro |
−9.22 kcal/mol |
25 |
9. |
Subtilosin-A |
Bacillus subtilis |
Mpro |
-146.0 kcal/mol |
26 |
10. |
Leodoglucomide |
Bacillus licheniformis |
PLpro |
− 6.7 |
27 |
11. |
Gramicidin S |
Bacillus brevis |
PLpro |
−11.4 kcal/mol |
28 |
12. |
Tyrocidine A |
Bacillus brevis |
Plpro |
−13.1 kcal/mol |
28 |
Table (2):
Summary of the structure of natural compounds that are potential as protease inhibitors for Mpro and PLpro
No |
Compound |
Structure and Molecular formula |
Sources (PubChem ID) |
|---|---|---|---|
1 |
Chondrillasterol |
C29H48O 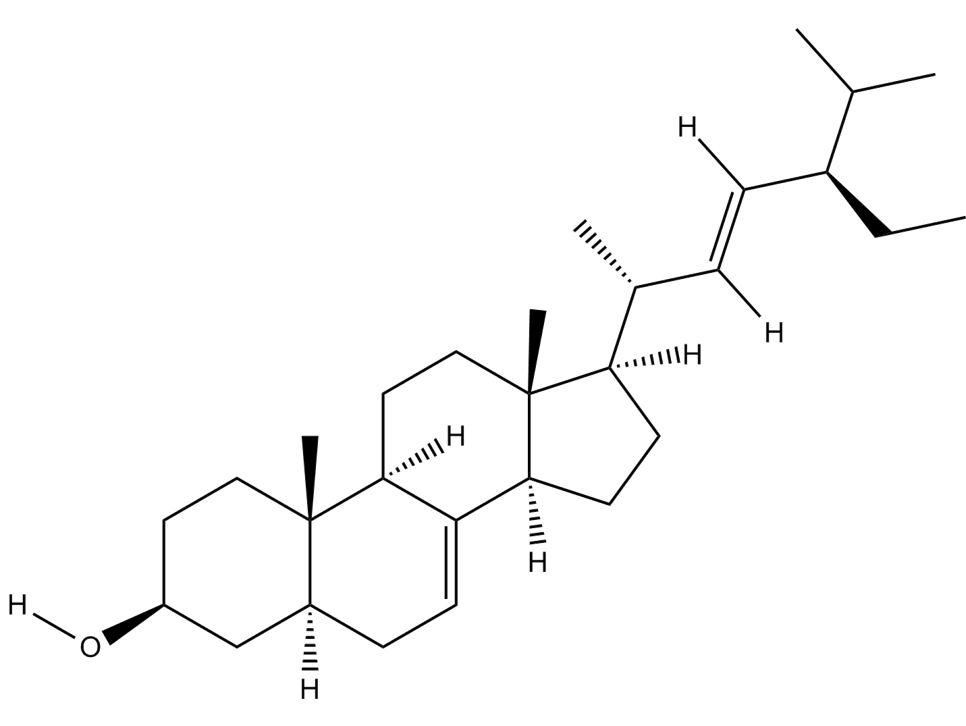 |
5283663 |
2 |
Cholestane |
C27H48 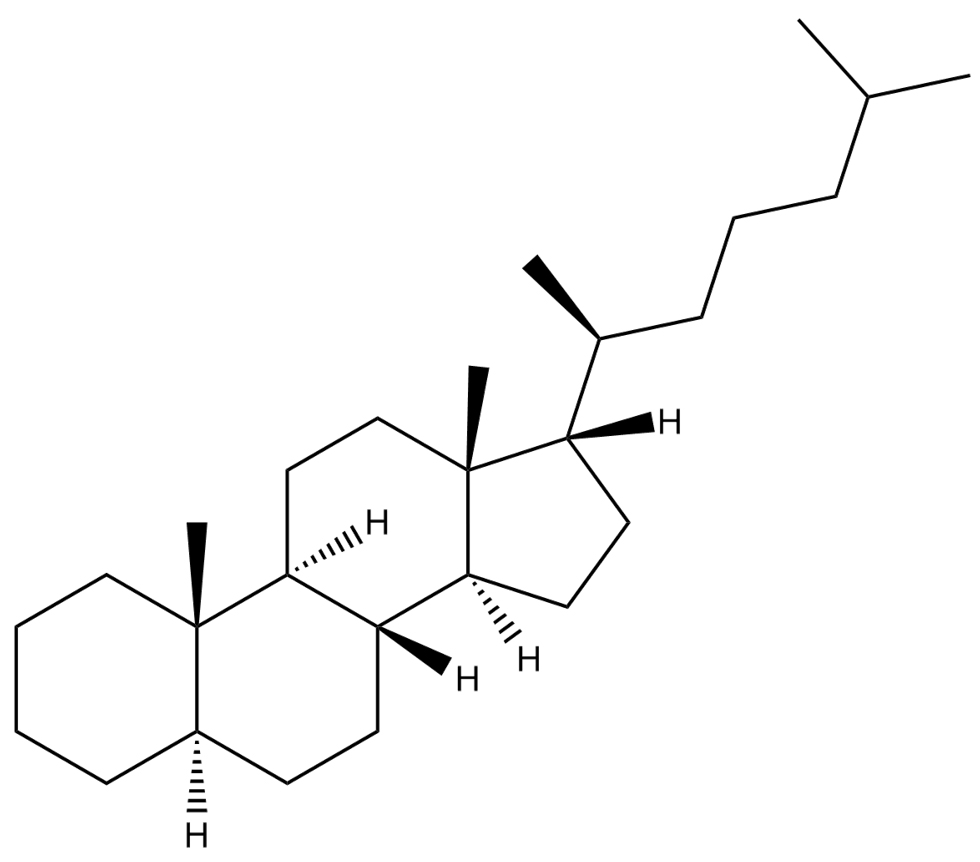 |
637620 |
3 |
Trifluoroacetic acid |
C2HF3O2 or CF3COOH 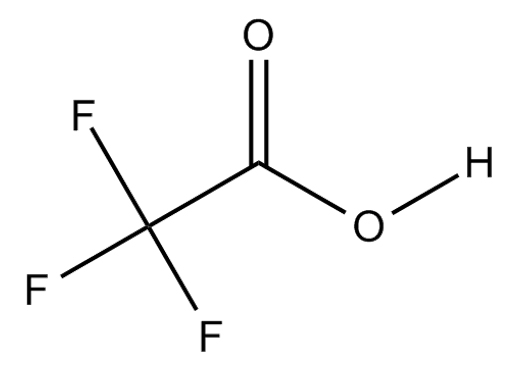 |
6422 |
4 |
Octadeccenoic-acid |
C18H34O2 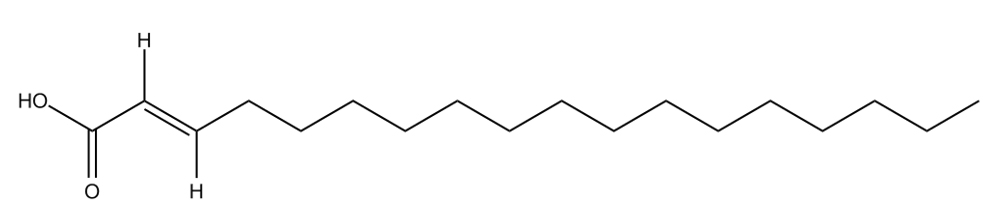 |
5282750 |
5 |
Stigmasterol |
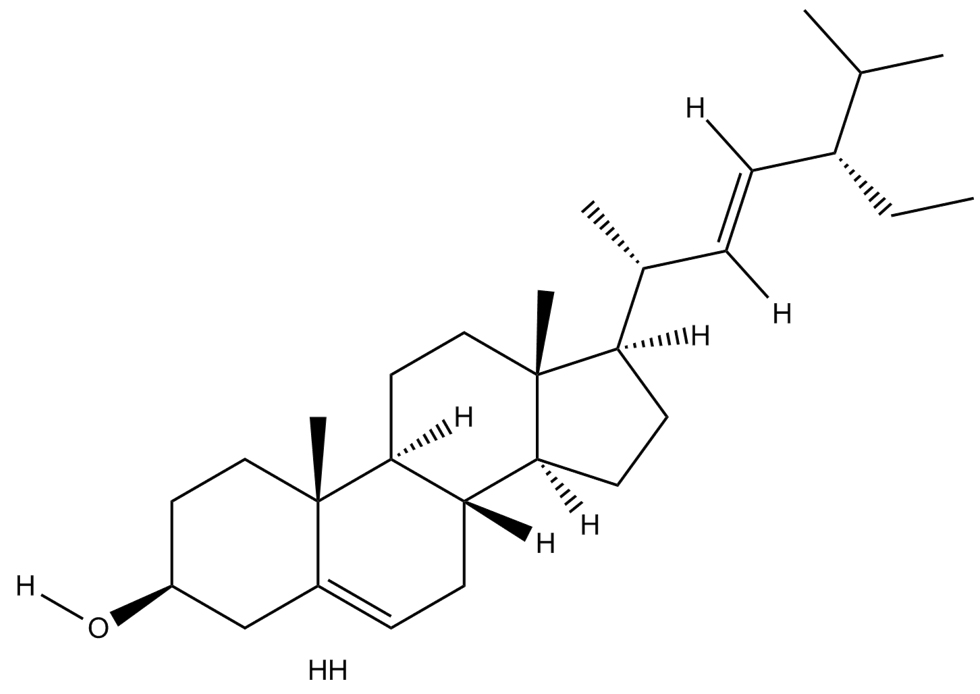 |
5280794 |
6 |
9 octadecenoic-acid |
C18H34O2 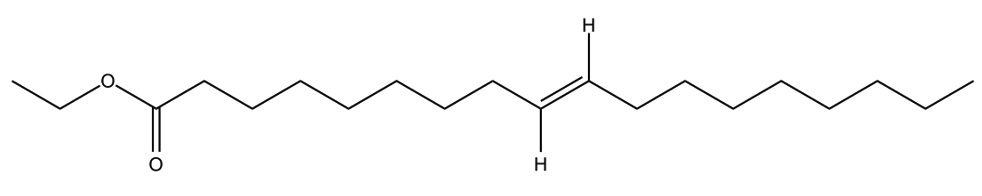 |
5364430 |
7 |
Hexadecanoic acid |
C16H32O2 |
985 |
8 |
Macrolaktin A |
C24H34O5 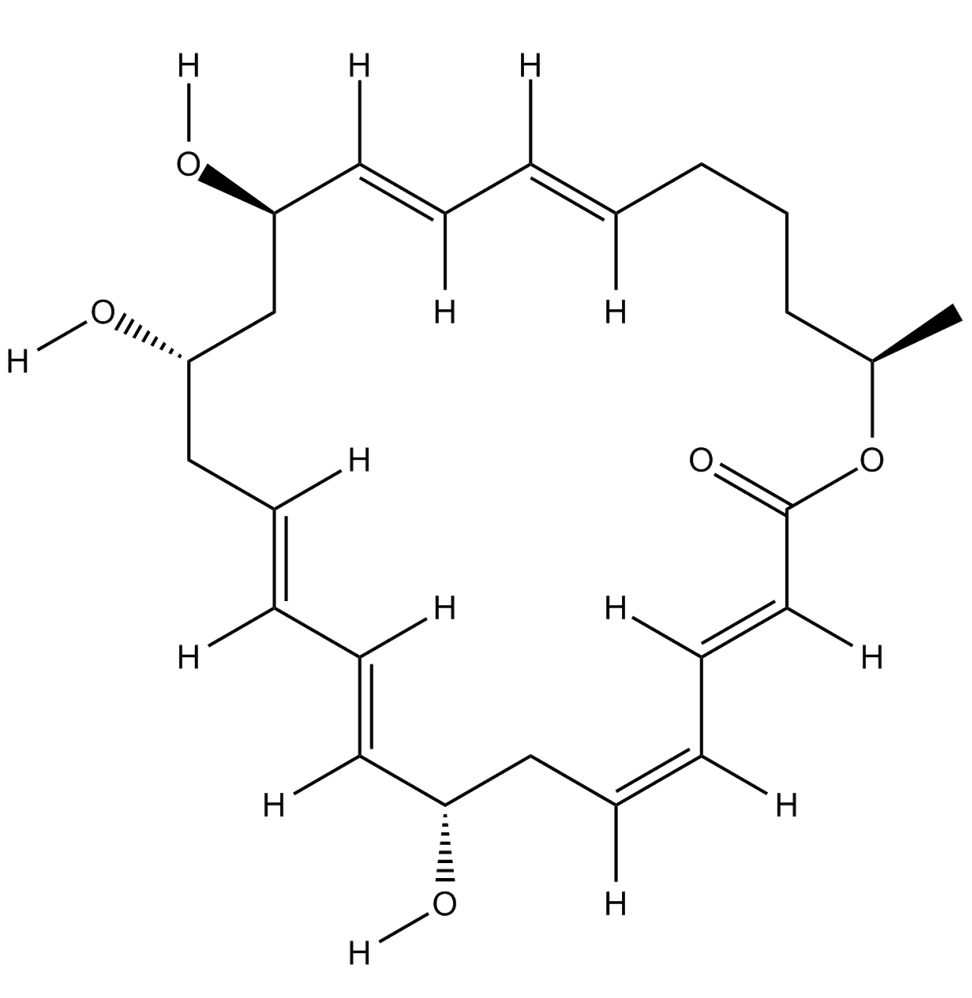 |
14504311 |
9 |
Subtilosin-A |
C129H208N36O41S3  |
16132390 |
10 |
Gramicidin S |
C60H92N12O10 (homodetic cyclic peptide) 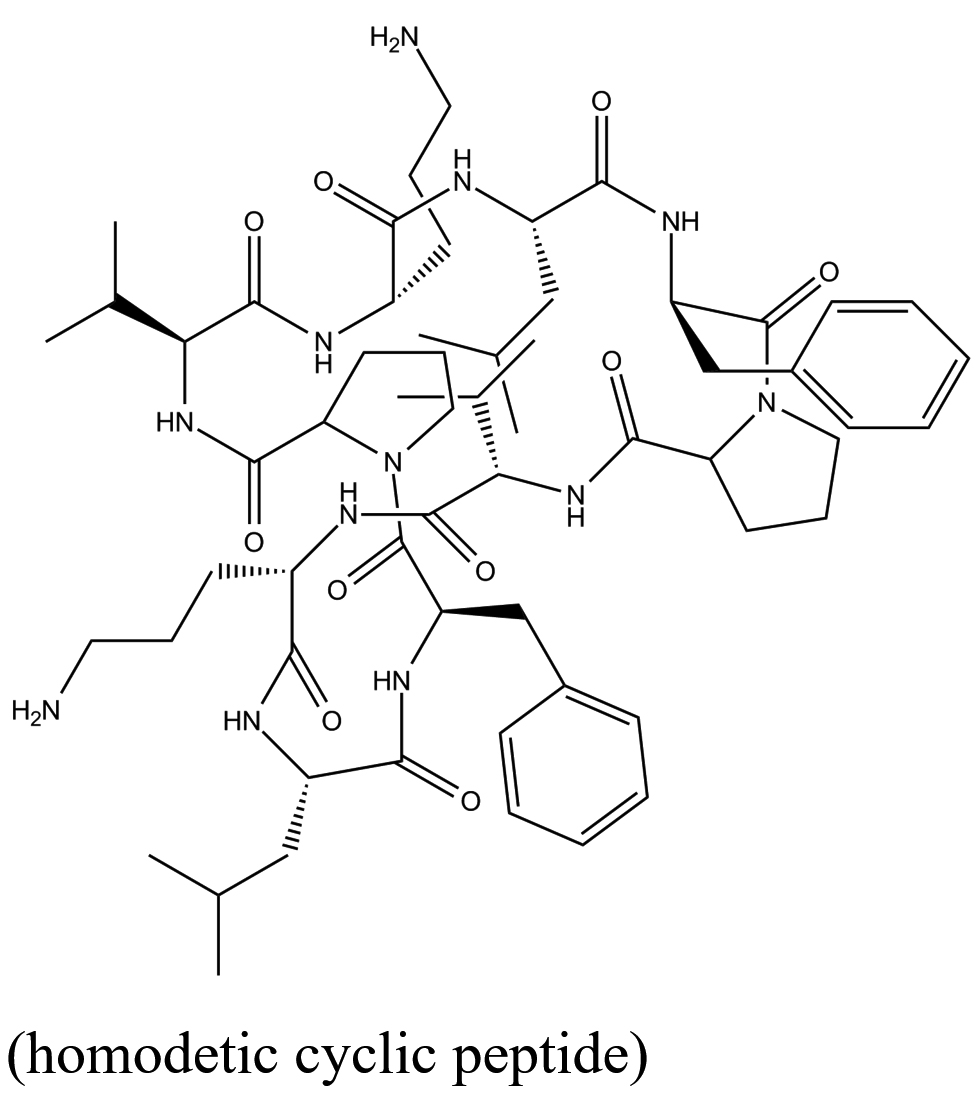 |
73357 |
11 |
Tyrocidine A |
C66H87N13O13 (homodetic cyclic decapeptide) 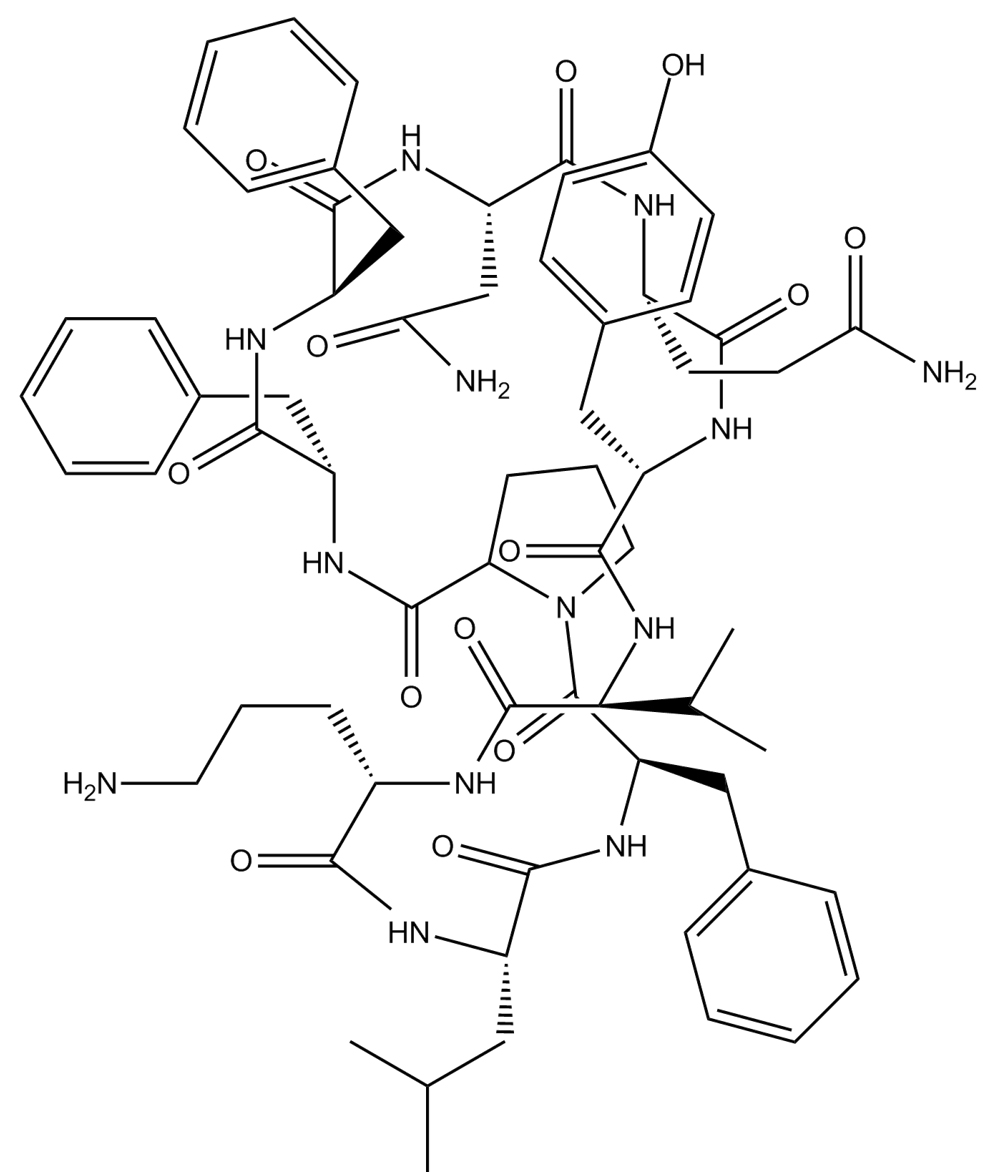 |
16129635 |
In conclusion, this review identified that the global COVID-19 pandemic caused by SARS-CoV-2 prompted research into alternative antiviral drugs to inhibit viral replication. Specific antiviral drugs targeting viral proteases were considered important to achieve effective therapy, since SARS-CoV-2 was found to infect host cells and replicate through the help of Mpro, and PLpro. The activity of viral proteases could be inhibited using compounds such as chondrillasterol, cholestane, trifluoroacetic acid, octadecenoic acid, stigmasterol, 9-octadecenoic acid, hexadecanoic acid, Macrolactin A, Subtilosin A, Leodoglucomide, Gramicidin S, and Tyrocidine A derived from Bacillus spp. The results of molecular docking analysis showed that these compounds bound specifically to the active sites of Mpro and PLpro.
ACKNOWLEDGMENTS
The authors are grateful to Universitas Padjadjaran for the financial support provided through the RDPD Research Grant in 2020.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Payne S. Family Coronaviridae. Viruses. 2017:149-158.
Crossref - Goyal B., Goyal D. Targeting the dimerization of main protease of coronaviruses: a potential broad-spectrum therapeutic strategy. ACS Comb.Sci. 2020;22:297–305.
Crossref - Zumla A, Chan JF, Azhar EI, Hui DS, Yuen KY. Coronaviruses-Drug Discovery and Therapeutic Options. Nat Rev Drug Discov. 2016;15(5):327-347.
Crossref - Huang C, Wang Y, Li X, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet (London, England). 2020;395(10223):497-506.
Crossref - Wu D, Wu T, Liu Q, Yang Z. The SARS-CoV-2 Outbreak: What We Know. Int J Infect Dis. 2020(94):44-48.
Crossref - Zhou D, Dejnirattisai W, Supasa P, et al. Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. Cell. 2021;184(9):2348-2361.e6.
Crossref - World Health Organization. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. 2021: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern, accessed on May 22, 2023
- Kokic G, Hillen HS, Tegunov D, et al. Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nat Commun. 2021;12:279.
Crossref - Pardi N, Weissman D. Development of vaccines and antivirals for combating viral pandemics. Nat Biomed Eng. 2020;4:1128-1133.
Crossref - Luan B, Huynh T, Cheng X, Lan G, Wang HR. Targeting Proteases for Treating COVID-19. J Proteome Res. 2020;19(11):4316-4326.
Crossref - Sapkale PV, Jadhav SB, Sable P. Protease inhiBitors : a review. Indian Drugs. 2013;50(2):5-19
Crossref - Sayed AM, Khattab AR, AboulMagd AM, Hassan MH, Rateb ME, Zaid H, Abdelmohsen UR. Nature as a treasure trove of potential anti-SARS-CoV drug leads: a structural/ mechanistic rationale. RSC Adv. 2020;10:19790-19802.
Crossref - Chakravarti R, Singh R, Ghosh A, et al. A review on potential of natural products in the management of COVID-19. RSC Adv. 2021;11:16711-16735.
Crossref - Sabotic J, Kos J. Microbial and fungal protease inhibitors–current and potential applications. Appl Microbiol Biotechnol. 2012;93(4):1351-1375.
Crossref - Bastos JCSB, Kohn LK, Fantinatti-Garboggini F, et al. Antiviral activity of Bacillus sp. isolated from the marine sponge Petromica citrina against bovine viral diarrhea virus, a surrogate model of the hepatitis C virus. Viruses. 2013;5(5):1219-1230.
Crossref - Song K, Luo F, Chen W, Qi X, Shen Y, Zha J, Ling F, Wang G. Evaluation on the antiviral activity of Bacillus velezensis extract against spring viremia of carp virus. Aquaculture. 2022;547:737477.
Crossref - Alam S, Sadiqi S, Sabir M, Nisa S, Ahmad S, Abbasi SW. Bacillus species; a potential source of anti-SARS-CoV-2 main protease inhibitors. J Biomol Struct Dyn. 2021;40(13):5748-5758.
Crossref - Polonca S. Environment Shape the Intra-species Diversity of Bacillus subtilis Isolates. Microb Ecol. 2020;79(4):853-864.
Crossref - Turnbull PCB. Bacillus (Chap. 15) in Medical Microbiology 4th Edition (Ed. Baron). University of Texas Medical Branch at Galveston, USA. 1996: https://www.ncbi.nlm.nih.gov/books/NBK7699/
- Mondol MAM, Shin HJ, Islam MT. Diversity of Secondary Metabolites from Marine Bacillus Species: Chemistry and Biological Activity. Mar. Drugs. 2013:2846-2872.
Crossref - Jackson CB, Farzan M, Chen B, Choe H. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol. 2022;23(1):3-20.
Crossref - Sharma A, Gupta SP. Fundamentals of Viruses and Their Proteases. Viral Proteases and Their Inhibitors. 2017:1-24.
Crossref - Zhang L , Gao Z, Zhao X, Qi G. A natural lipopeptide of surfactin for oral delivery of insulin. Drug Delivery. 2016;23(6):2084-2093.
Crossref - Marathe KR, Patil RH, Vishwakarma KS, Chaudhari AB, Maheshwari VL. Chapter 6 – Protease Inhibitors and Their Applications: An Overview. Studies in Natural Products Chemistry. 2019;62:211-242.
Crossref - Bharadwaj KK, Sarkar T, Ghosh A, et al. Macrolactin A as a Novel Inhibitory Agent for SARS-CoV-2 Mpro: Bioinformatics Approach. Appl Biochem Biotechnol. 2021;193(10):3371-3394.
Crossref - Balmeh N, Mahmoudi S, Fard NA. Manipulated bio antimicrobial peptides from probiotic bacteria as proposed drugs for COVID-19 disease. Inform Med Unlocked. 2021;23:100515.
Crossref - Bansal P, Kumar R, Singh J, Dhanda S. In silico molecular docking of SARS-CoV-2 surface proteins with microbial non-ribosomal peptides: identification of potential drugs. J Proteins Proteom. 2021;12(3):177-184.
Crossref - Razali R, Asis H, Budiman C. Structure-Function Characteristics of SARS-CoV-2 Proteases and Their Potential Inhibitors from Microbial Sources. Microorganisms. 2021;9(12):2481.
Crossref - Anirudhan V, Lee H, Cheng H, Cooper L, Rong L. Targeting SARS-CoV-2 viral proteases as a therapeutic strategy to treat COVID-19. J Med Virol. 2021;93(5):2722-2734.
Crossref - Farady CJ, Craik CS. Mechanisms of macromolecular protease inhibitors. Chembiochem : A European J Chem Biol. 2010;11(17):2341-2346.
Crossref - Yuan L, Zhang S, Peng J, Li Y, Yang Q. Synthetic surfactin analogues have improved anti-PEDV properties. PLoS ONE. 2019;14(4):e0215227.
Crossref - Chowdhury T, Baindara P, Mandal SM. LPD-12: a promising lipopeptide to control COVID-19. Int J Antimicrob Agents. 2021;57(1):106218.
Crossref - Huang X, Lu Z, Zhao H, Bie X, Lu F, Yang S. Antiviral Activity of Antimicrobial Lipopeptide from Bacillus subtilis fmbj Against Pseudorabies Virus, Porcine Parvovirus, Newcastle Disease Virus and Infectious Bursal Disease Virus in Vitro. Int J Pept Res Ther. 2006;12:373-377.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.