


ISSN: 0973-7510
E-ISSN: 2581-690X
An arsenic-resistant bacteria (strain PKA 200) was isolated from the Hindon River (Ghaziabad) after analyzing its water sample. Strain PKA 200 belongs to the Pseudomonas genus and can convert harmful arsenite [As (III)] into less harmful arsenate [As(V)]. Strain PKA 200 exhibits a remarkable ability to sense and move towards arsenite. Strain PKA 200 is a promising candidate for studying how bacteria sense and transform arsenic. Here, we report a novel circular plate assay to demonstrate both chemotaxis and biotransformation of arsenite by Pseudomonas sp. PKA 200.
Detoxification, Arsenic, Biotransformation, Chemotaxis
Arsenic is a naturally occurring metalloid that can be found in the environment, both as a result of natural processes and human activities. It exists in various chemical forms, including organic and inorganic species, which have different levels of toxicity.1,2 Most common forms are arsenite [As(III)] and arsenate [As(V)]. Arsenite is generally more toxic and mobile than arsenate.3-5 Biotransformation and chemotaxis play significant roles in the environmental behavior and impact of arsenic.
The arsenic biotransformation involves the microbial conversion of arsenic compounds from one form to another.6,7 This process can significantly influence the toxicity, mobility, and bioavailability of arsenic in the environment. Microorganisms, particularly bacteria, play a crucial role in arsenic biotransformation.6,7 They can metabolize arsenic through several pathways, including reduction, oxidation and methylation.6,7
Microbial reduction of arsenate [As(V)] to arsenite [As(III)] is a common biotransformation pathway.8-11 Arsenate-reducing bacteria, such as members of the genus Shewanella and Geobacter, utilize arsenate as an electron acceptor in anaerobic respiration.8-11 This reduction increases the solubility and mobility of arsenic, making it more bioavailable and toxic.
Some bacteria can oxidize arsenite to arsenate depending on the specific bacterial species.12 The oxidation of highly toxic arsenite to less toxic arsenate is considered as a detoxification mechanism.13,14 These bacteria are crucial in controlling the speciation and mobility of arsenic in the environment. By transforming arsenic compounds, they can influence the bioavailability and toxicity of arsenic to other organisms. Arsenic-oxidizing bacteria are found in a range of environments, including arsenic-contaminated soils, groundwater, surface waters, and hydrothermal vent systems. They are also found in extreme environments like arsenic-rich hot springs.13,14 This process can immobilize arsenic by converting it to less soluble and less toxic forms. Arsenite-oxidizing bacteria have potential applications in bioremediation efforts to reduce arsenic contamination in water and soil.13,14 By converting toxic arsenite into less toxic arsenate, they can help mitigate the health and environmental risks associated with arsenic contamination.15
Arsenic can also undergo methylation, a process where methyl groups are added to arsenic compounds.16 Certain microorganisms, such as Methanobacterium and Methanosarcina, can methylate arsenic to produce organic forms like monomethylarsonic acid (MMA) and dimethylarsinic acid (DMA).16-18 Methylation can reduce arsenic toxicity, but it can also produce volatile arsenic compounds, which may lead to environmental dispersion.16-18
Some arsenic-oxidizing bacteria exhibit chemotactic responses to different concentrations of arsenic compounds in their environment.14,15 For example, they might move toward areas with higher concentrations of arsenite to facilitate its oxidation and subsequent detoxification.15 Alternatively, if arsenate is more favorable for their metabolism, they may exhibit chemotaxis towards regions with higher concentrations of arsenate.15
Understanding the interplay between arsenic oxidation and chemotaxis in bacteria is crucial for environmental scientists and microbiologists studying bioremediation strategies or natural attenuation of arsenic-contaminated sites. This communication details the discovery of a new bacterium exhibiting both the ability to convert toxic arsenite into a less harmful form and a remarkable movement towards it. Furthermore, in this communication, we report a novel circular plate assay to demonstrate both chemotaxis and biotransformation of arsenite by a newly isolated Pseudomonas sp. PKA 200.
Isolation of Arsenite-resistance bacteria
A water sample from the Hindon River was collected in sterile bottles for isolating arsenite-resistant bacteria. The study used broth culture enrichment followed by selective agar plating with sodium arsenite. Initially, 1 mL sample was added to 500 mL of nutrient broth containing 200 ppm sodium arsenite, and the bacteria were cultured for two days. Following this, the sample underwent serial dilution and was plated on selective agar media supplemented with the same concentration of arsenite. These plates were then incubated for an additional two days. Ultimately, eight different bacterial isolates, each displaying unique morphologies, were selected, and preserved for further analysis.
Screening of bacteria for Arsenic resistance
To evaluate the tolerance levels of each bacterial isolate, the eight strains were individually streaked onto nutrient agar plates with sodium arsenite concentrations ranging from 200 to 1000 ppm. Following a 48-hour incubation period at 30°C, only one strain, labeled PKA 200, demonstrated significant resistance, successfully growing even at the maximum concentration of 500 ppm. As a result, strain PKA 200 was selected for further study.
Identification and biochemical characterization of bacterial strain
The bacterial strain PKA 200 was identified by sequencing its 16S ribosomal RNA gene, utilizing a method described in a previous study.19 Phylogenetic tree was constructed as described previously.19 All biochemical tests and morphological characteristics were performed by the standard methods as described previously.20
Arsenite transformation assay
Pseudomonas sp. PKA 200 was grown on nutrient agar plates containing 500 ppm sodium arsenite or sodium arsenate. The oxidation of arsenite to arsenate and the reduction of arsenate to arsenite were determined by the silver nitrate method.21
Determination of minimum inhibitory concentrations
Strain PKA 200 was grown on nutrient agar plates containing different concentrations of arsenite (200 ppm to 1000 ppm) and plates were incubated at 30°C. The MIC was considered as the lowest concentrations of arsenite that inhibits the complete growth of strain PKA 200 within 48 hours.
Chemotaxis towards Arsenite
Drop plate assay and circular plate assay were used to demonstrate the chemotactic potential of Pseudomonas sp. PKA 200 towards arsenite.
Drop plate assay
Drop plate assay was performed as described previously.22 The bacteria were grown overnight in a minimal medium containing 200 ppm sodium arsenite (the attractant) and 20 mM sodium succinate (the energy source). Subsequently, the cells were harvested by centrifugation, washed twice with a saline solution, and resuspended in minimal medium containing 0.3% agar. This cell suspension was then poured onto a Petri plate to solidify. To test if the bacteria are attracted to arsenic, small amounts of sodium arsenite, sodium arsenate, or aspartic acid (a known attractant) were placed in the center of Petri dishes. The dishes were then incubated for a few hours. If the bacteria are attracted, they will gather around the crystals in visible rings.
A circular plate assay
Two rigs of 0.4% bacto agar with increasing concentration of sodium arsenite (100 ppm to 200 ppm) towards center were casted on glass petri-dishes. The outermost ring was inoculated with cells of strain PKA 200, whereas the inner ring remained uninoculated. This experiment was carried out in triplicate and plates were collected at 0 h, 24 h and 36 h, respectively and flooded with 0.1 M silver nitrate solution. Chemotaxis and transformation of arsenite was determined by the color development with 0.1 M silver nitrate solution.21 Arsenite provides yellow color with silver nitrate whereas arsenate give red-brown color.21
Characterization of an Arsenite-oxidizing bacteria
An arsenite-oxidizing bacterium, strain PKA 200 was isolated from a polluted water sample collected from the Hindon river, Ghaziabad. Strain PKA 200 was subjected to the 16S rRNA gene sequencing and identified as a member of genus Pseudomonas. The 16S rRNA gene sequence of strain PKA 200 was submitted to the NCBI Genbank under accession number MW440700. The phylogenetic tree, constructed using the neighbour-joining method, showed that strain PKA 200 forms a clade with Pseudomonas allupotida, supported by a high bootstrap value (Figure 1).

Figure 1. Phylogenetic analysis of Pseudomonas sp. PKA 200
Cells of strain PKA 200 were Gram-negative, strictly aerobic, motile and rod shaped. Cells were 1.4-2.0 µm long and 0.7-1.2 µm in diameter. The various biochemical tests of strain PKA 200 were summarized in Table.
Table:
Biochemical and Morphological characteristics of Pseudomonas sp. PKA 200
Characteristics |
Results |
Gram stain |
Negative |
Shape |
Rods |
Colony colour |
White |
Catalase |
Positive |
Spore formation |
Negative |
Oxidase |
Positive |
Motility |
Positive |
Casein hydrolysis |
Negative |
Gelatin hydrolysis |
Negative |
Citrate utilization |
Positive |
Indole formation |
Positive |
Urease test |
Negative |
Growth on MacConkey Agar |
Positive |
H2S production |
Negative |
Aesculin hydrolysis |
Negative |
Fermentation of carbohydrates |
|
Raffinose |
Negative |
Mannitol |
Positive |
Arabinose |
Negative |
Maltose |
Negative |
Sucrose |
Negative |
Lactose |
Negative |
Growth studies
The growth of Pseudomonas sp. PKA 200 was assessed under two different conditions: in the presence and absence of arsenite [As(III)]. In this study, bacterial growth was measured by monitoring optical density (OD) at a specific wavelength (600 nm) (Figure 2). In the absence of arsenite, Pseudomonas sp. PKA 200 showed better growth, indicating that without the toxic stress of arsenite, the bacteria were able to proliferate efficiently. The normal growth conditions allowed for rapid cell division and biomass accumulation. In the presence of arsenite, bacterial growth was significantly reduced. This suggests that arsenite had an inhibitory effect on the metabolism or cellular processes of Pseudomonas sp. PKA 200. The bacterium may have been exposed to oxidative stress, disruption in protein synthesis, or impaired energy production, which slowed down or halted its growth. The growth studies highlights how arsenite, as a toxic substance, impacts bacterial growth, and provides insight into the bacterium’s potential tolerance limits or mechanisms of resistance. The data in Figure 2 likely show slower growth rates or lower final biomass in arsenite-containing conditions, illustrating the stress imposed by this toxic compound.
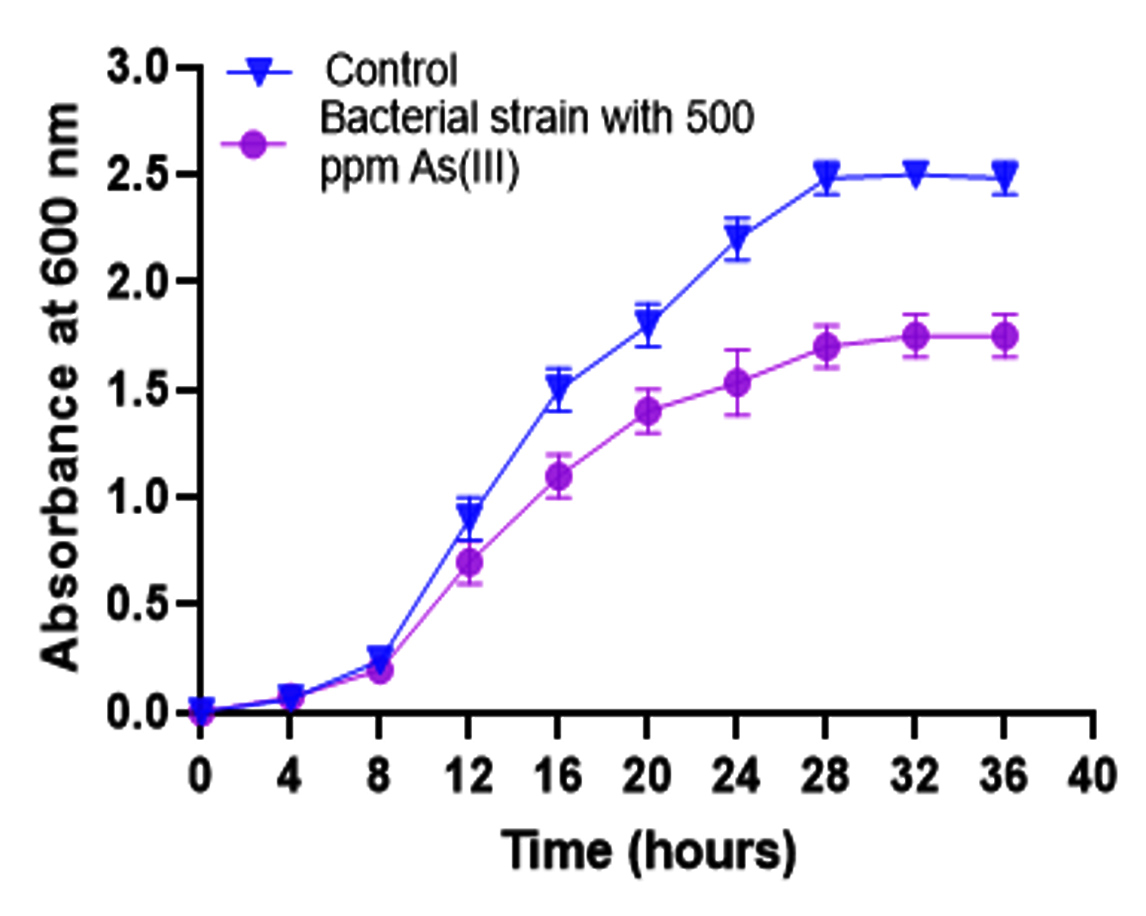
Figure 2. Growth curve of Pseudomonas sp. PKA 200 in the absence (control) or presence of sodium arsenite
Arsenite transformation assay
The potential of Pseudomonas sp. PKA 200 for arsenite [As (III)] oxidation and arsenate [As (V)] reduction was determined qualitatively using silver nitrate (AgNO3) test that is based on reaction between AgNO3 and As (III) or As (V). When 0.1M silver nitrate solution was added into 48 h old bacterial nutrient agar culture plate containing 500 ppm sodium arsenite, the plate colour was red-brown due to formation of silver arsenate, confirming complete bacterial oxidation of arsenite [As(III] to arsenate [As (V)] within 48 h. Whereas, the 48 h old control uninoculated nutrient agar plate containing 500 ppm sodium arsenite produced a yellow colour after adding silver nitrate solution due to presence of silver arsenite. To monitor the ability of cells of strain PKA 200 to reduce arsenate, the silver nitrate solution was added into 48 h old bacterial nutrient agar plate containing sodium arsenate, the plate colour was red-brown due to presence of silver arsenate, suggesting that cells of PKA 200 were unable to reduce arsenate to arsenite. Minimal inhibitory concentration of As(III) for strain PKA 200 was determined as 700 ppm.
Chemotaxis towards Arsenite
Drop plate assay
Drop plate assay exhibited the bacterial ring of cells of strain PKA 200 around aspartic acid which is known attractant (Figure 3A). Bacterial ring was also observed where chemotactic material was sodium arsenite, suggesting movement of the PKA 200 cells towards arsenite (Figure 3B). However, there is no bacterial ring towards sodium arsenate (Figure 3C). This data indicates that strain PKA exhibited chemotaxis only towards As (III) and did not show chemotactic behaviour towards As (V).

Figure 3. Chemotaxis of Pseudomonas sp. PKA 200. (A) aspartic acid, (B) sodium arsenite, and (C) sodium arsenate
Circular plate assay
A circular plate assay was carried out to show the capability of strain PKA 200 to oxidize arsenite and chemotaxis towards it at regular intervals. Figure 4 showed that there is no biotransformation and bacterial chemotaxis towards arsenite in 0 h plate (at time of experiment start) and the colour of the plate was yellow after flooding with 0.1M silver nitrate. In 24 h plate, most of arsenite around the outer bacterial ring was converted to arsenate, indicating by the red-brown colour after flooding with 0.1M silver nitrate. The red-brown colour was due to formation of silver arsenate (Figure 4). In 36 h plate, bacteria completely moved to higher concentrations of arsenite and transformed into arsenate, confirming by complete red-brown colour of plate (after flooding with 0.1M silver nitrate) and disappearance of outermost ring where bacteria was inoculated. This assay also suggests that arsenite oxidation was associated with arsenite chemotaxis in Pseudomonas sp. PKA 200.
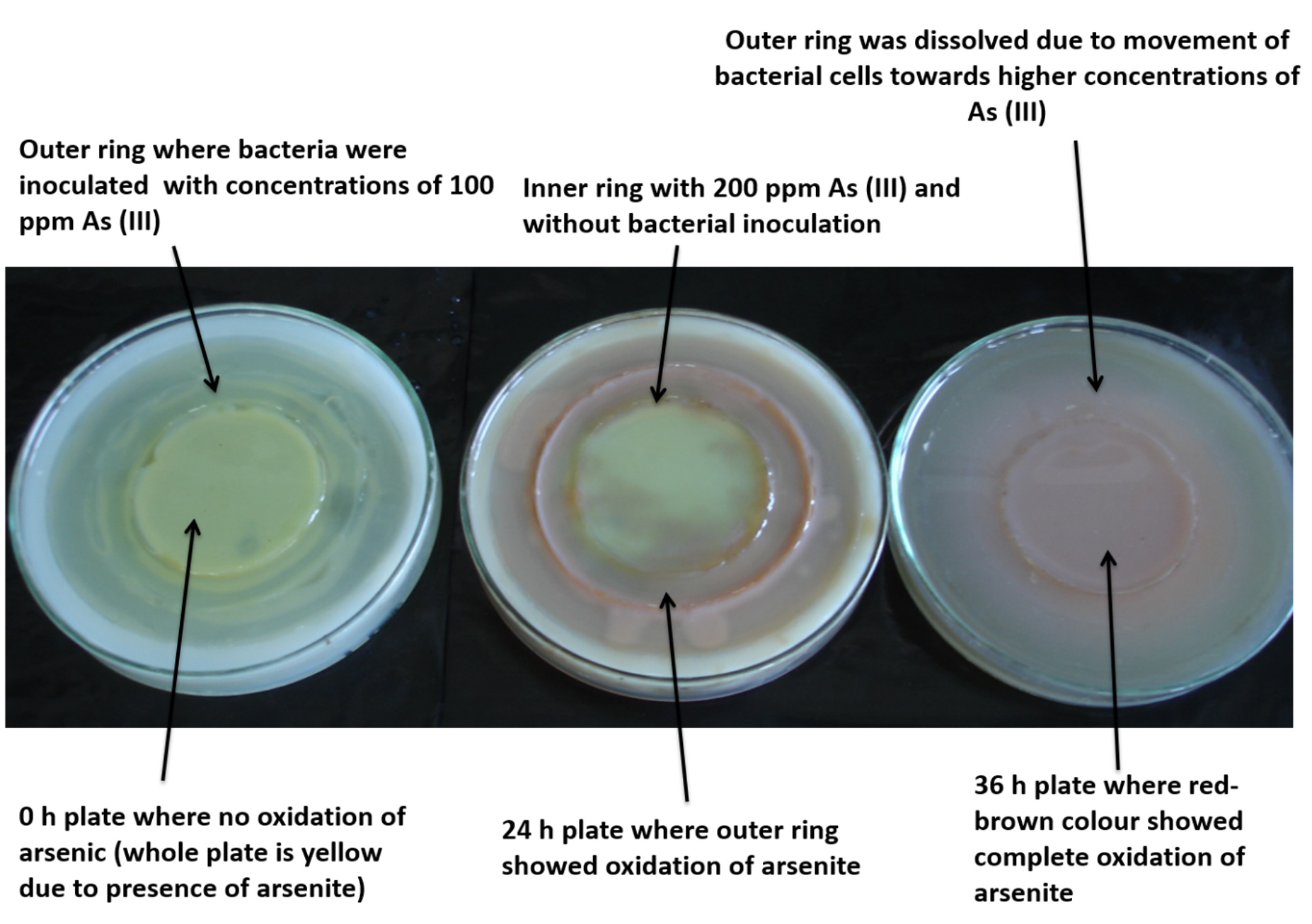
Figure 4. A Circular plate assay showing bacterial chemotaxis and transformation towards arsenite
In this study, we have isolated and identified a new arsenite-oxidizing bacterium, Pseudomonas sp. PKA 200. Literature studied showed that many As(III) oxidizing bacteria including Pseudomonas,23 Agrobacterium,24 Alkalilimnicola,25 Microbacterium,26 Micrococcus,27 Bosea,28 Delftia,29 Thiobacillus,30 Anaeromyxobacter,30 and Ensifer31 have already been characterized for As(III) oxidation. Few bacteria grew well in arsenic containing media and gained energy during As(III) oxidation.32-35 As(III) served as a sole electron donor during their growth.32,35 In this study, the growth of Pseudomonas sp. PKA 200 was monitored in presence of As(III) and results showed that Pseudomonas sp. PKA 200 was unable to gain energy during As(III) oxidation as there was no inductive effect of As(III) on growth of Pseudomonas sp. PKA 200. Our study clearly showed that the growth of strain PKA 200 was reduced in the presence of arsenite as compared to control where no arsenite was present in the media. Furthermore, an arsenic transformation assay using silver nitrate demonstrated that strain PKA 200 detoxified arsenite [As(III)] into arsenate [As(V)], which is less toxic than arsenite.
In this study, we reported that Pseudomonas sp. PKA 200 exhibited chemotaxis toward arsenite. Earlier research indicates that only a few bacteria are known to show positive or negative chemotaxis towards heavy metals.36,37 Positive chemotaxis of heavy metals is considered as a part of bioremediation mechanism in which bacteria move towards heavy metals to transform them.36 On the other hand, negative chemotaxis is a survival mechanism in which bacteria remove themselves from heavy metals mediated toxicity areas.36 Research investigations have reported that four marine strains of Pseudomonas species exhibited negative chemotaxis of heavy metals.37 In this report, the positive chemotaxis towards arsenite by a Pseudomonas sp. PKA 200 was demonstrated using various methods including drop plate assay and a circular plate assay. The drop plate assay has already been used for successful demonstration of chemotaxis towards various xenobiotic compounds using Pseudomonas species.38 For example, Arora and Bae38 reported that Pseudomonas sp. strain JHN exhibited positive chemotaxis towards 4-chloro-2-nitrophenol using drop plate assay. Similarly, the five isolates of Pseudomonas, taxonomically related to P. jessenii, P. moorei, P. vancouverensis, P. reinekei, P. koreensis, and P. moraviensis exhibited chemotaxis towards sodium dodecyl sulphate.39 Another strain BUR11 of Pseudomonas sp. was reported to exhibit chemotaxis towards parathion, chlorphrifos and their hydrolytic intermediates.40 Grim and Harwood41 used drop plate assay to exhibit chemotaxis towards naphthalene by Pseudomonas putida G7 and Pseudomonas sp. strain NCIB 9816-4.
In this study, we introduce a novel circular plate assay to demonstrate chemotaxis toward arsenite by Pseudomonas sp. PKA 200. The assay indicates that arsenite oxidation is associated with chemotaxis in Pseudomonas sp. PKA 200. Previous investigations have shown that no single method in the literature can simultaneously assess both chemotaxis and arsenite oxidation.14,15 Therefore, this assay is the only method that can monitor both arsenite oxidation and chemotaxis toward arsenite. This method can also be used to detect arsenate reduction and chemotaxis toward arsenate.42 Additionally, it may be applicable for studying bacterial chemotaxis and the reduction or oxidation of other heavy metals, such as the reduction of selenite to red-colored selenium and chemotaxis toward it.43 This method could also be suitable for investigating bacterial degradation of colored organic compounds, such as nitrophenols and chloronitrophenols.44
To date, only few As(III) oxidation bacteria exhibited chemotaxis towards arsenite.14,15 The common feature of these bacteria is that they gained energy during arsenite oxidation. However, in this study Pseudomonas sp. PKA 200 exhibited chemotaxis towards arsenite, but it was unable to gain energy during arsenite oxidation.
Pseudomonas strain PKA-200 shows a distinctive capability to biotransform the toxic arsenite [As(III)] into less harmful arsenate [As(V)]. Its unique ability to sense and move toward arsenite underscores its potential as an effective agent in bioremediation of arsenic-contaminated environments. This strain holds significant promise for further research into microbial arsenic sensing and detoxification mechanisms, offering a valuable foundation for developing sustainable biotechnological solutions for arsenic mitigation.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Mukherjee A, Coomar P, Sarkar S, et al. Arsenic and other geogenic contaminants in global groundwater. Nat Rev Earth Environ. 2024;5(4):312-328.
Crossref - Issanov A, Adewusi B, Saint-Jacques N, Dummer TJB. Arsenic in drinking water and lung cancer: A systematic review of 35 years of evidence. Toxicol Appl Pharmacol. 2024;483:116808.
Crossref - Ganie SY, Javaid D, Hajam YA, Reshi MS. Arsenic toxicity: Sources, pathophysiology and mechanism. Toxicol Res. 2024;13(1):tfad111.
Crossref - Sevak P, Pushkar B. Arsenic pollution cycle, toxicity and sustainable remediation technologies: A comprehensive review and bibliometric analysis. J Environ Manage. 2024;349:119504.
Crossref - Rajendran S, Rathinam V, Sharma A, Vallinayagam S, Muthusamy M. Arsenic and Environment: A Systematic Review on Arsenic Sources, Uptake Mechanism in Plants, Health Hazards and Remediation Strategies. Top Catal. 2024;67(1):325-341.
Crossref - Mohsin H, Shafique M, Rehman Y. Genes and biochemical pathways involved in microbiatransformation of arsenic. Arsenic Toxicity: Challenges and Solutions. 2021:391-413.
Crossref - Yan G, Chen X, Du S, Deng Z, Wang L, Chen S. Genetic mechanisms of arsenic detoxification and metabolism in bacteria. Curr Genet. 2019;65(2):329-338.
Crossref - Drewniak L, Stasiuk R, Uhrynowski W, Sklodowska A. Shewanella sp. O23S as a driving agent of a system utilizing dissimilatory arsenate-reducing bacteria responsible for self-cleaning of water contaminated with arsenic. Int J Mol Sci. 2015;16(7):14409-14427.
Crossref - Ohtsuka T, Yamaguchi N, Makino T, et al. Arsenic dissolution from Japanese paddy soil by a dissimilatory arsenate-reducing bacterium Geobacter sp. OR-1. Environ Sci Technol. 2013;47(12):6263-6271.
Crossref - Tsuchiya T, Ehara A, Kasahara Y, et al. Expression of genes and proteins involved in arsenic respiration and resistance in dissimilatory arsenate-reducing Geobacter sp. strain OR-1. Appl Environ Microbiol. 2019;85(14):e00763.
Crossref - Dang Y, Walker DJF, Vautour KE, Dixon S, Holmes DE. Arsenic detoxification by Geobacter species. Appl Environ Microbiol. 2017;83(4):e02689-16.
Crossref - Herrera C, Moraga R, Bustamante B, et al. Characterization of arsenite-oxidizing bacteria isolated from arsenic-rich sediments, Atacama Desert, Chile. Microorganisms. 2021;9(3):483.
Crossref - Mujawar SY, Shamim K, Vaigankar DC, Naik MM, Dubey SK. Rapid arsenite oxidation by Paenarthrobacter nicotinovorans strain SSBW5: Unraveling the role of GlpF, aioAB and aioE genes. Arch Microbiol. 2023;205(10):333.
Crossref - Shi K, Fan X, Qiao Z, et al. Arsenite oxidation regulator AioR regulates bacterial chemotaxis towards arsenite in Agrobacterium tumefaciens GW4. Sci Rep. 2017;7(1):43252.
Crossref - Shi K, Wang Q, Wang G. Microbial oxidation of arsenite: Regulation, chemotaxis, phosphate metabolism and energy generation. Front Microbiol. 2020;11:569282.
Crossref - Hemmat-Jou MH, Liu S, Liang Y, Cheng G, Fang L, Li F. Microbial arsenic methylation in soil-water systems and its environmental significance. Sci Total Environ. 2024;944:173873.
Crossref - Li P, Wang Y, Jiang Z, et al. Microbial diversity in high arsenic groundwater in Hetao Basin of Inner Mongolia, China. Geomicrobiol J. 2013;30(10):897-909.
Crossref - Wang YH, Li P, Dai XY, et al. Abundance and diversity of methanogens: Potential role in high arsenic groundwater in Hetao Plain of Inner Mongolia, China. Sci Total Environ. 2015;515:153-161.
Crossref - Kumar A, Mishra R, Srivastava A, et al. Diversity of hexavalent chromium-reducing bacteria and physicochemical properties of the Kanpur tannery wastewater. Environ Chem Ecotoxicol. 2023;5:205-212.
Crossref - Arora PK, Chauhan A, Pant B, et al. Chryseomicrobium imtechense gen. nov., sp. nov., a new member of the family Planococcaceae. Int J Syst Evol Microbiol. 2011;61(8):1859-1864.
Crossref - Simeonova DD, Lievremont D, Lagarde F, Muller DAE, Groudeva VI, Lett MC. Microplate screening assay for the detection of arsenite-oxidizing and arsenate-reducing bacteria. FEMS Microbiol Lett. 2004;237(2):249-253.
Crossref - Pandey J, Sharma NK, Khan F, Ghosh A, Oakeshott JG, Jain RK, Pandey G. Chemotaxis of Burkholderia sp. strain SJ98 towards chloronitroaromatic compounds that it can metabolise. BMC Microbiola. 2012;12:1-9
- Satyapal GK, Kumar R, Kumar S, et al. Cloning and functional characterization of arsenite oxidase (aoxB) gene associated with arsenic transformation in Pseudomonas sp. strain AK9. Gene. 2023;850:146926.
Crossref - Salmassi TM, Venkateswaren K, Satomi M, et al. Oxidation of arsenite by Agrobacterium albertimagni AOL15, sp. nov., isolated from Hot Creek, California. Geomicrobiol J. 2002;19(1):53-66.
Crossref - Zargar K, Hoeft S, Oremland R, Saltikov CW. Identification of a novel arsenite oxidase gene, arxA, in the haloalkaliphilic, arsenite-oxidizing bacterium Alkalilimnicola ehrlichii strain MLHE-1. J Bacteriol. 2010;192:3755-3762.
Crossref - Mokashi SA, Paknikar KM. Arsenic (III) oxidizing Microbacterium lacticum and its use in the treatment of arsenic contaminated groundwater. Lett Appl Microbiol. 2002;34(4):258-262.
Crossref - Paul T, Chakraborty A, Islam E, Mukherjee SK. Arsenic bioremediation potential of arsenite-oxidizing Micrococcus sp. KUMAs15 isolated from contaminated soil. Pedosphere. 2018;28(2):299-310.
Crossref - Lu X, Zhang Y, Liu C, Wu M, Wang H. Characterization of the antimonite- and arsenite-oxidizing bacterium Bosea sp. AS-1 and its potential application in arsenic removal. J Hazard Mater. 2018;359:527-534.
Crossref - Biswas R, Vivekanand V, Saha A, Ghosh A, Sarkar A. Arsenite oxidation by a facultative chemolithotrophic Delftia spp. BAs29 for its potential application in groundwater arsenic bioremediation. Int Biodeterior Biodegrad. 2019;136:55-62.
Crossref - Li Y, Guo L, Yang R, et al. Thiobacillus spp. and Anaeromyxobacter spp. mediate arsenite oxidation-dependent biological nitrogen fixation in two contrasting types of arsenic-contaminated soils. J Hazard Mater. 2023;443(Pt A):130220.
Crossref - Li X, Li J, Zhao Q, Qiao L, Wang L, Yu C. Physiological, biochemical, and genomic elucidation of the Ensifer adhaerens M8 strain with simultaneous arsenic oxidation and chromium reduction. J Hazard Mater. 2023;441:129862.
Crossref - vanden Hoven RN, Santini JM. Arsenite oxidation by the heterotroph Hydrogenophaga sp. str. NT-14: The arsenite oxidase and its physiological electron acceptor. Biochim Biophys Acta. 2004;1656(2-3):148-155.
Crossref - Wang Q, Qin D, Zhang S, et al. Fate of arsenate following arsenite oxidation in Agrobacterium tumefaciens GW4. Environ Microbiol. 2015;17(6):1926-1940.
Crossref - Santini JM, vanden Hoven RN. Molybdenum-containing arsenite oxidase of the chemolithoautotrophic arsenite oxidizer NT-26. J Bacteriol. 2004;186(6):1614-1619.
Crossref - Silver S, Phung LT. Genes and enzymes involved in bacterial oxidation and reduction of inorganic arsenic. Appl Environ Microbiol. 2005;71(2):599-608.
Crosssref - Arora PK, Jeong MJ, Bae H. Chemotaxis away from 4-chloro-2-nitrophenol, 4-nitrophenol, and 2, 6-dichloro-4-nitrophenol by Bacillus subtilis PA-2. J Chem. 2015;2015(1):1-6.
Crossref - Young LY, Mitchell R. Negative chemotaxis of marine bacteria to toxic chemicals. Appl Microbiol. 1973;25(6):972-975.
Crossref - Arora PK, Bae H. Biotransformation and chemotaxis of 4-chloro-2-nitrophenol by Pseudomonas sp. JHN. Microb Cell Fact. 2014;13:1-6.
Crossref - Furmanczyk EM, Kaminski MA, Spolnik G, et al. Isolation and characterization of Pseudomonas spp. strains that efficiently decompose sodium dodecyl sulfate. Front Microbiol. 2017;8:1872.
Crossref - Pailan S, Saha P. Chemotaxis and degradation of organophosphate compound by a novel moderately thermo-halo tolerant Pseudomonas sp. strain BUR11: evidence for possible existence of two pathways for degradation. PeerJ. 2015;3:1378
Crossref - Grimm AC, Harwood CS. Chemotaxis of Pseudomonas spp. to the polyaromatic hydrocarbon naphthalene. Appl Environ Microbiol. 1997;63(10):4111-4115.
Crossref - He X, Xiao W, Zeng J, Tang J, Wang L. Detoxification and removal of arsenite by Pseudomonas sp. SMS11: Oxidation, biosorption and bioaccumulation. J Environ Manage. 2023;336:117641.
Crossref - Rehan M, Alsohim AS, El-Fadly G, Tisa LS. Detoxification and reduction of selenite to elemental red selenium by Frankia. Antonie Van Leeuwenhoek. 2019;112(2):127-139.
Crossref - Arora PK, Srivastava A, Garg SK, Singh VP. Recent advances in degradation of chloronitrophenols. Bioresour Technol. 2018;250:902-909.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.