


ISSN: 0973-7510
E-ISSN: 2581-690X
Candida albicans biofilms are highly resistant to conventional antifungal agents such as fluconazole, largely due to matrix sequestration, efflux pump activation, and altered sterol metabolism. Among its virulence factors, the secreted aspartyl proteinase family (SAPs) plays a pivotal role in tissue invasion, adhesion, and biofilm development. While SAP1-SAP6 have been extensively characterized, SAP5 remains underexplored despite its strong association with biofilm formation and antifungal resistance. This study aimed to evaluate the antifungal potential of bioactive phytometabolites derived from Astilbe rivularis against the biofilm-associated enzyme SAP5 (PDB ID: 2QZX) using computational approaches. Seven major phytometabolites from A. rivularis, retrieved from the IMPPAT database, were analyzed using AutoDock Vina for molecular docking against SAP5. Protein-protein interaction analysis was conducted through the STRING database to assess SAP5’s functional associations. Top-ranked ligands were visualized in PyMol and Discovery Studio to identify key amino acid interactions. Pharmacokinetic and drug-likeness evaluations were performed using SwissADME, while fluconazole served as the reference control. All seven phytometabolites exhibited strong binding affinities toward SAP5, ranging from -6.4 to -8.3 kcal/mol, indicating stable ligand-protein interactions. Key residues involved in binding included Asp218, Thr221, Tyr222, Ser278, Gly287, and Asp290. The STRING network revealed that SAP5 participates in a dense interaction cluster (11 nodes, 48 edges) associated with fungal virulence pathways. ADME profiling confirmed that all compounds satisfied Lipinski’s Rule of Five, displaying high gastrointestinal absorption and favourable drug-likeness comparable to fluconazole. This study provides the first in silico evidence that Astilbe rivularis phytochemicals can act as natural inhibitors of the Candida albicans SAP5 enzyme, potentially disrupting biofilm formation and virulence. The results highlight SAP5 as a promising antifungal target and position A. rivularis as an underexplored plant source for developing resistance-sparing, plant-based therapeutics.
Antifungal phytoconstituents, Aspartyl protease inhibition, Biofilm-associated virulence, Ethnopharmacology, Quorum sensing modulation, Genetics, Disease, Green product
Oral candidiasis has emerged as more than a simple opportunistic infection, with mounting evidence linking Candida albicans to the pathogenesis and progression of oral cancer.1 Chronic colonization by Candida in the oral cavity not only contributes to mucosal inflammation but also promotes carcinogenesis through the production of carcinogenic metabolites such as nitrosamines and acetaldehyde.2 More critically, the ability of C. albicans to form robust biofilms on oral mucosal surfaces and prosthetic devices enhances its resistance to conventional antifungal agents, creating a reservoir of persistent infection. These biofilms not only impede antifungal penetration but also promote epithelial dysplasia, tumor progression, and even facilitate metastatic dissemination through epithelial–mesenchymal transition (EMT)-like changes.3,4 Thus, Candida biofilms act as hidden cofactors in both tumour initiation and progression of oral squamous cell carcinoma (OSCC).
Invasive fungal disease (IFD) caused by C. albicans and related fungal pathogens has been increasingly documented in immunocompromised populations, particularly in cancer patients undergoing chemotherapy and radiotherapy.5 Approximately 1.7 million deaths annually are caused by the invasive fungal diseases, which is relatively comparable to the 1.5 million deaths caused by tuberculosis and significantly greater than malaria’s annual death toll of 405,000.6 The compromised mucosal barriers and altered immune responses in oral cancer patients create a conducive environment for fungal invasion beyond superficial layers, leading to systemic candidiasis. This interplay between oral cancer and invasive candidal infection not only worsens morbidity but also complicates treatment protocols, as antifungal resistance further reduces therapeutic success. The growing recognition of this association highlights the need for novel antifungal strategies that can target both planktonic and biofilm-associated Candida to improve patient outcomes.7
Despite the limitations of synthetic antifungals and their adverse side effects, attention has shifted toward natural plant-based therapeutics as promising alternatives.8-10 Phytometabolites from medicinal plants have demonstrated broad-spectrum antimicrobial, antioxidant, and anticancer activities, offering a safer and sustainable strategy for managing fungal biofilms.11,12 One promising plant in this context is Astilbe rivularis, a perennial herb belonging to the family Saxifragaceae, widely distributed across the Eastern and Central Himalayan regions of India, including Sikkim, Arunachal Pradesh, and Uttarakhand. In traditional Indian medicine and local Himalayan folk practices, A. rivularis has been valued for its anti-inflammatory, hepatoprotective, and antimicrobial properties.13 The roots and rhizomes are commonly used in Ayurvedic and ethnomedicinal formulations to alleviate fever, body pain, liver ailments, and microbial infections. Decoctions and powdered preparations are traditionally employed for wound healing, rheumatic pain, and digestive disturbances, while topical applications are used to treat skin infections and inflammatory lesions.14 In indigenous healing systems of the Himalayan communities, the plant is also used as a blood purifier and general tonic to promote tissue repair and restore vitality. These diverse therapeutic uses, supported by its rich composition of flavonoids, terpenoids, phenolics, and saponins, highlight Astilbe rivularis as a potential source of natural bioactive compounds for modern drug discovery and biomedical research in India.15 The plant is rich in flavonoids, terpenoids, phenolic compounds, and saponins, which reinforce these traditional uses and provide antimicrobial, antioxidant, and immunomodulatory activities.
Among the virulence factors contributing to C. albicans pathogenicity, the Secreted Aspartyl Proteinase (SAP) family plays a pivotal role in facilitating tissue invasion, adhesion, and immune evasion.16 The SAP gene family comprises at least ten isoenzymes (SAP1-SAP10), which are expressed at different stages of fungal growth and infection. Of these, SAP5 has been extensively implicated in biofilm formation, mucosal colonization, and host cell damage, making it a critical determinant of C. albicans virulence.17 Hence, the present study aims to evaluate the molecular interactions of plant-derived antifungal metabolites from A. rivularis against key biofilm-associated proteins of C. albicans using molecular docking approaches. By exploring the binding affinity and potential inhibitory effects of these phytochemicals, this study seeks to identify novel plant-based antifungal metabolites that could be developed into adjunctive therapies for combating Candida biofilms and reducing their role in oral cancer progression.
Selection and retrieval of phytometabolites
Bioactive phytometabolites of A. rivularis were identified using the IMPPAT (Indian Medicinal Plants, Phytochemistry, and Therapeutics) database (https://cb.imsc.res.in/imppat/). Seven major metabolites previously reported for antimicrobial, antioxidant, and anti-inflammatory activities were selected for molecular docking analysis.18 The three-dimensional (3D) structures of the selected phytometabolites were obtained in Structure Data File (SDF) format and subsequently converted to Protein Data Bank (PDB) format using Open Babel (version 3.1.1). Energy minimization was performed using the MMFF94 force field to generate the most stable conformers before docking.
Target protein selection and preparation
The C. albicans biofilm-associated enzyme Aspartyl Proteinase 5 (SAP5) was selected as the target protein due to its role in fungal adhesion, invasion, and biofilm formation. The crystallographic structure of SAP5 (PDB ID: 2QZX) was retrieved from the RCSB Protein Data Bank (https://www.rcsb.org/).19 Protein preparation was carried out using AutoDock Tools (version 1.5.7) by removing water molecules, heteroatoms, and co-crystallized ligands. Polar hydrogen atoms were added, and Kollman charges were assigned. The processed protein structure was saved in PDBQT format for subsequent docking analysis.
Molecular docking studies
Molecular docking simulations were conducted using AutoDock Vina (version 1.2.3) to predict the binding orientation and affinity of A. rivularis phytometabolites toward the SAP5 protein. The docking grid box was configured to encompass the catalytic region of SAP5, ensuring complete coverage of the active site residues. The exhaustiveness parameter was set to 8 to achieve reliable conformational sampling. The docking output files were analyzed, and ligand-protein complexes were visualized using PyMol version 2.5 and Discovery Studio Visualizer 2021 to examine hydrogen bonds, hydrophobic interactions, and π-π stacking patterns.
Protein-protein interaction (ppi) network construction
The biological network and functional associations of SAP5 were analyzed using the STRING database (https://string-db.org/).20 The analysis was performed with a minimum confidence score of 0.7 to ensure high-confidence interactions. The generated network map displayed nodes representing proteins and edges representing predicted functional associations. The resulting interaction data were used to assess SAP5’s involvement in biofilm-associated and virulence-related pathways in C. albicans.
Molecular interaction profiling
Detailed examination of ligand-receptor interactions was carried out for the top-ranked docking conformations. Active site residues contributing to ligand binding were identified, focusing on hydrogen bonding, hydrophobic contacts, and van der Waals interactions. The interaction profiles and contact maps were visualized using Discovery Studio Visualizer, and key amino acid residues were documented to validate the accuracy of docking conformations.
Pharmacokinetic and drug-likeness evaluation
Pharmacokinetic and drug-likeness analyses were performed using the SwissADME web tool (https://www.swissadme.ch/).21 Each metabolite was evaluated for compliance with Lipinski’s Rule of Five and other essential physicochemical parameters, including molecular weight, Log P, number of hydrogen bond donors and acceptors, topological polar surface area (TPSA), and number of rotatable bonds.22 Parameters related to absorption, distribution, metabolism, and excretion (ADME) were analyzed to assess the suitability of the metabolites as potential orally bioavailable antifungal agents.
Molecular docking analysis
Seven bioactive phytometabolites isolated from A. rivularis were retrieved from the IMPPAT phytochemical database and subjected to molecular docking against the C. albicans biofilm-associated enzyme Aspartyl Proteinase (SAP5) (PDB ID: 2QZX). All metabolites exhibited strong binding affinities ranging from -6.4 to -8.3 kcal/mol, indicating stable and energetically favourable interactions with the target protein (Figure 1). The high negative binding energies reflect robust ligand-receptor complex formation, suggesting that the phytometabolites may effectively inhibit SAP5 activity, which is central to fungal adhesion, invasion, and biofilm maintenance. These docking results support the hypothesis that A. rivularis contains potent antifungal compounds capable of targeting biofilm-associated virulence factors in C. albicans.
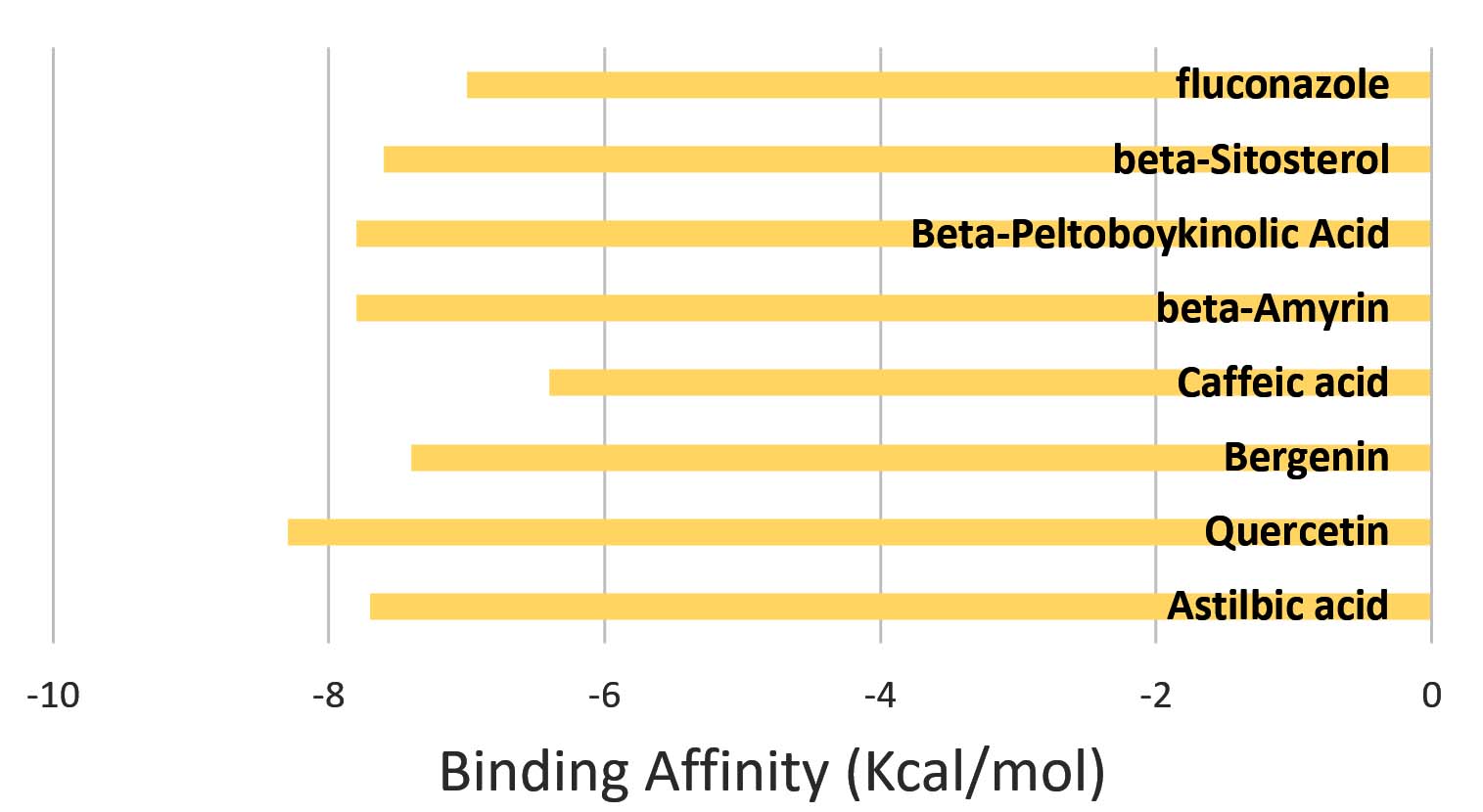
Figure 1. Binding affinity of phytochemicals from A. rivularis against the C. albicans biofilm-associated enzyme SAP5
Interaction network and functional correlation
Protein-protein interaction (PPI) analysis conducted using the STRING database demonstrated that SAP5 is intricately involved in a dense interaction network comprising 11 nodes and 48 edges (Figure 2). This network reveals functional associations between SAP5 and other key virulence-related proteins involved in cell adhesion, proteolysis, and hyphal morphogenesis. The high clustering coefficient observed emphasizes the central regulatory role of SAP5 in maintaining fungal pathogenicity and biofilm stability. Inhibition of SAP5, therefore, could potentially disrupt multiple downstream processes vital for biofilm integrity and infection persistence. The observed multi-protein connectivity reinforces SAP5’s role as a prime therapeutic target for antifungal drug development.
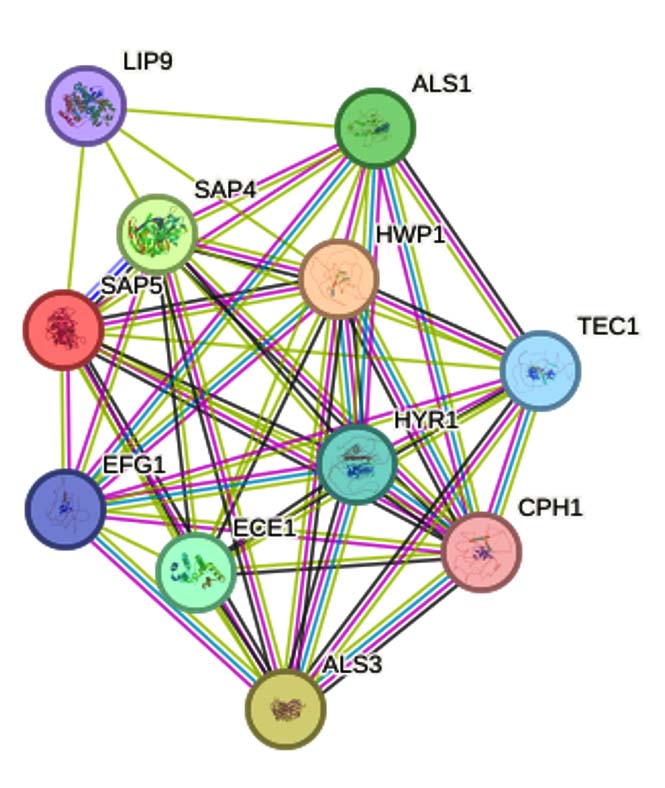
Figure 2. Protein-Protein interaction network of SAP5 obtained from the STRING database, displaying 11 nodes and 48 edges
Binding affinity and amino acid interaction profile
Detailed docking interaction analysis revealed that A. rivularis derived phytometabolites bind specifically within the catalytic groove of SAP5, forming stable non-covalent interactions that strengthen the ligand-protein complex. Key amino acid residues involved included Asp218, Thr221, Tyr222, Ser278, Gly287, and Asp290, which contributed to multiple hydrogen bonds, hydrophobic contacts, and π-π stacking interactions with the active site residues (Table). The extensive hydrogen bonding network stabilized the ligand orientation, while hydrophobic and van der Waals interactions enhanced molecular complementarity and docking stability. Among the metabolites, those with the highest affinity (-8.3 kcal/mol) exhibited multiple-site occupancy within the active pocket, suggesting a high probability of catalytic inhibition. The interaction profiles confirm that these bioactive metabolites can effectively interfere with the proteolytic mechanism of SAP5, potentially attenuating C. albicans biofilm formation and virulence.
Table: Shows key amino acid residues of the phytochemicals derived from A. rivularis binding with SAP5 (C-Control)
| Compound | Binding Affinity (kcal/mol) | Hydrogen Bonds | Hydrophobic Contacts | van der Waals Interactions | π-π Stacking |
|---|---|---|---|---|---|
| Quercetin | -8.3 | Thr13, Thr221, Ser35, Ser88, Asp32 | Ile305, Ile123, Ile30, Leu216 | Gly34, Gly220, Gly85, Asp218, Asp86, Ala119, Lys121 | Tyr84, Tyr225 |
| Beta-Amyrin | -7.8 | Thr221, Thr222 | Ile12, Ile223, Trp51, Phe281 | Gly220, Asp86 | Tyr225, Trp51 |
| Beta-Pelto- boykinolic acid | -7.8 | Ser35, Ser36, Asp32, Asp218 | Ile82, Ala133, Leu216, Tyr84 | Gly34, Gly85, Gly131, Lys83, Lys192, Lys193 | Tyr84 |
| Astilbic acid | -7.7 | Ser35, Ser36, Thr221, Asp32, Asp86 | Ile82, Ala133, Leu216, Tyr84 | Gly34, Gly85, Asp218, Lys83, Lys192, Lys193 | Tyr84 |
| Beta-Sitosterol | -7.6 | Thr13, Thr221, Thr222, Asp86 | Ile305, Ile30, Ile12, Ile123, Ile223 | Gly220, Gly85, Asp303, Glu302, Ser301, Ala119 | Tyr225 |
| Bergenin | -7.4 | Ser35, Thr221, Thr222, Asp32 | Ile123, Ile305, Ile12, Ile223, Leu216 | Gly34, Gly220, Gly84, Asp86, Asp218 | Tyr84, Tyr225 |
| Fluconazole (Control) | -7.0 | Asp32, Thr221, Thr222, | Ile30, Ile123, | Gly34, Gly85, Gly220, | Tyr84 |
| Asp86 | Leu216, Ile305 | Asp218, Asp303, Lys193 | |||
| Caffeic acid | -6.4 | Ser35, Ser88, Asp32, | Ile123, Ile30, | Gly34, Gly220, Gly85, | Tyr84 |
| Asp86 | Leu216, Tyr84 | Ala119 |
Molecular interaction analysis
The visualization of docking conformations through 2D and 3D interaction maps further confirmed the strong ligand engagement within the SAP5 catalytic domain. Ligand overlay analyses indicated that the A. rivularis compounds occupied conserved substrate-binding pockets, mimicking natural inhibitory motifs. Hydrogen bonds were predominantly formed between hydroxyl or carbonyl groups of the ligands and the backbone residues of Asp218 and Ser278, while hydrophobic stabilization occurred through aromatic ring interactions with Tyr222 and Thr221 (Figure 3). The combination of these bonding interactions accounts for the substantial binding energies observed, providing a mechanistic basis for their antifungal potential.
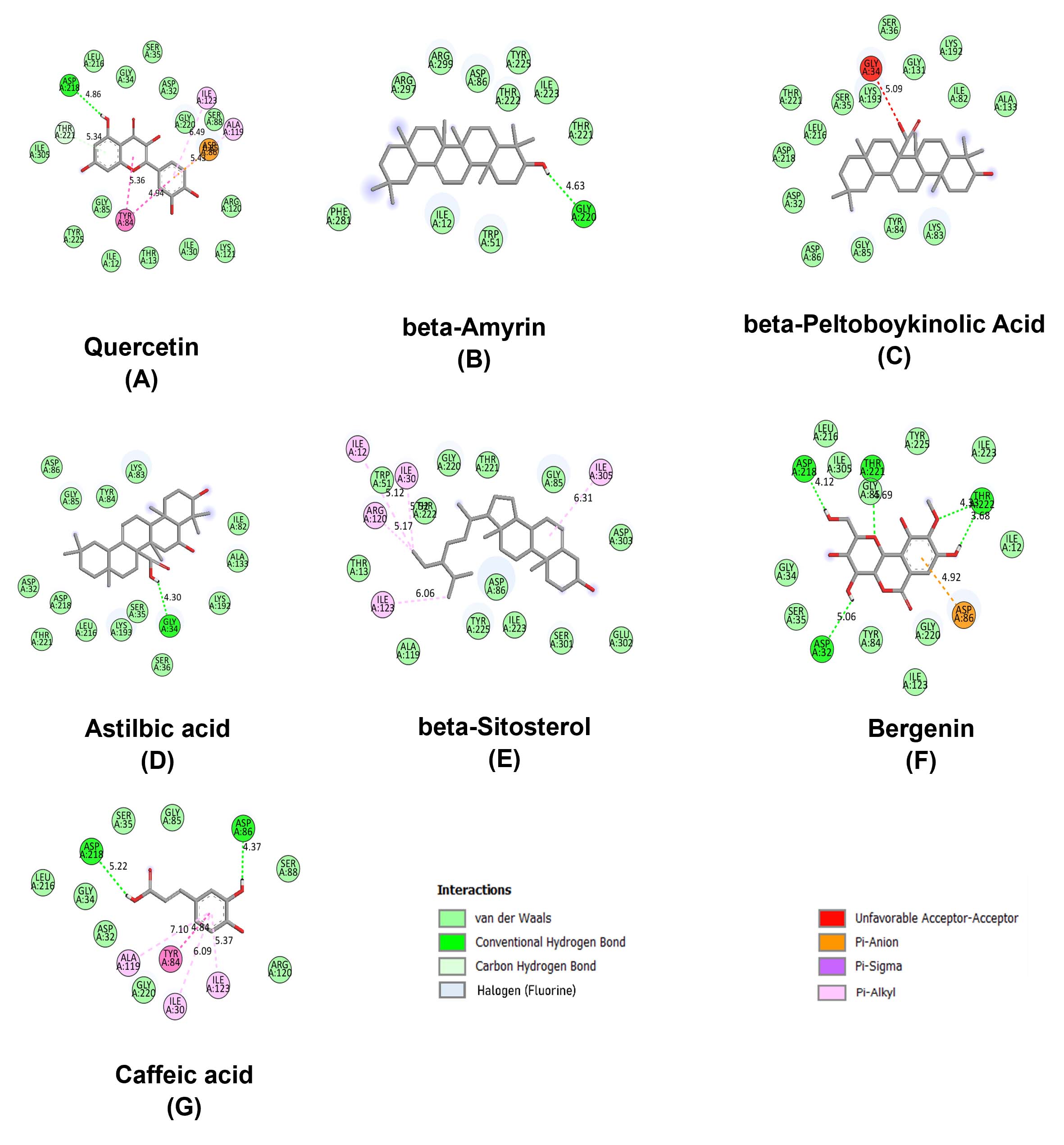
Figure 3. Binding orientations of the top phytometabolites of A. rivularis derived ligands within the SAP5 active site, illustrating the strong binding affinity through favourable molecular interactions
Pharmacokinetic radar profiling
The pharmacokinetic radar charts generated from the SwissADME tool revealed that all seven A. rivularis metabolites complied with Lipinski’s Rule of Five, suggesting their suitability as orally active drugs. Each metabolite exhibited ideal lipophilicity (Log P < 5), molecular weight (<500 Da), and balanced hydrogen bond donor and acceptor counts. The radar plots displayed optimized regions across six physicochemical parameters-lipophilicity, size, polarity, solubility, flexibility, and saturation-indicating strong absorption potential and good membrane permeability (Figure 4). Notably, none of the phytometabolites violated major pharmacological constraints, reinforcing their potential as lead antifungal scaffolds. When compared to the standard antifungal drug fluconazole, the A. rivularis derived compounds demonstrated comparable or superior physicochemical balance, particularly in parameters related to lipophilicity and polarity, suggesting improved bioavailability and reduced risk of efflux-mediated resistance. Importantly, none of the phytometabolites violated major pharmacological constraints or exhibited any PAINS or Brenk alerts, reinforcing their potential as lead antifungal scaffolds. Collectively, these ADME radar profiles validate that A. rivularis phytoconstituents possess desirable pharmacokinetic and drug-likeness properties, warranting further experimental validation as natural alternatives or adjuncts to fluconazole in the management of C. albicans biofilm-associated infections.
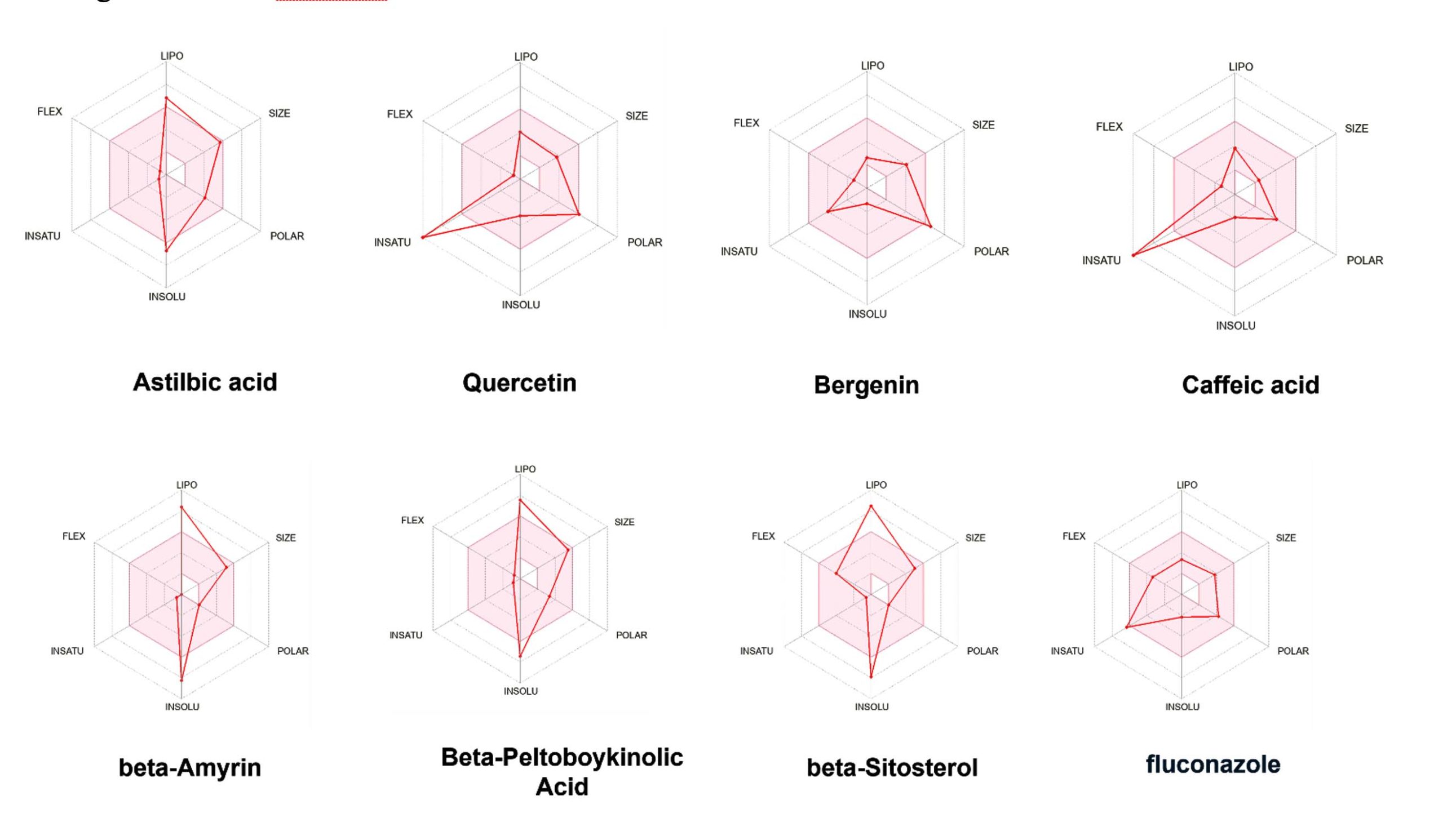
Figure 4. Three-dimensional radar profiling of A. rivularis derived ligands within the SAP5 active site, illustrating their interaction characteristics and binding performance
Drug-likeness and ADME profiling
The drug-likeness evaluation of the seven phytometabolites derived from A. rivularis revealed that all molecules satisfied the major criteria outlined in Lipinski’s Rule of Five, confirming their potential as orally active and bioavailable compounds (Figure 5A). Metabolites such as β-amyrin, β-sitosterol, bergenin, caffeic acid, quercetin, β-peltoboykinolic acid, and astilbic acid were found to fully comply with the limits for molecular weight (<500 Da), Log P (<5), hydrogen bond donors (≤5), and hydrogen bond acceptors (≤10). This indicates that the bioactive molecules possess favourable physicochemical attributes supporting good absorption and permeability profiles.
All metabolites exhibited optimal ADME characteristics, with high gastrointestinal absorption and no violation of major pharmacokinetic constraints. The strong connectivity of the metabolites in the network demonstrates their compatibility with favourable pharmacokinetic behaviour. When compared to the standard antifungal control fluconazole, the A. rivularis phytometabolites showed equivalent or improved profiles in lipophilicity, polarity, and flexibility, suggesting enhanced bioavailability with minimal toxicity risk (Figure 5B). Collectively, the findings confirm that these metabolites exhibit excellent drug-likeness and pharmacokinetic properties, reinforcing their potential as safe, natural antifungal candidates targeting Candida albicans biofilm protein SAP5.
Figure 5. (A) Drug-likeliness evaluation of Astilbe rivularis phytometabolites based on physicochemical parameters and established pharmacokinetic rules; (B) ADMET analysis of A. rivularis phytometabolites predicting absorption, distribution, metabolism, excretion and toxicity profiles
The current study explored the antifungal potential of bioactive phytometabolites from A. rivularis against the C. albicans virulence enzyme SAP5 through an integrated molecular docking and pharmacokinetic screening approach. The computational findings revealed that these plant-derived metabolites exhibit strong affinity toward the active site of SAP5, enhance their potential to interfere with biofilm-associated proteolytic mechanisms. The protein interaction analysis further emphasized SAP5 as a key regulatory enzyme within the Candida virulence network, consolidating its role as a strategic molecular target for antifungal intervention. These insights collectively suggest that A. rivularis derived metabolites may function as natural inhibitors capable of modulating critical pathways involved in fungal adhesion, invasion, and biofilm development.
Previous investigations have predominantly concentrated on elucidating the biochemical characteristics and substrate specificities of C. albicans SAP1-SAP6, emphasizing their affinity for hydrophobic residues and their involvement in proteolysis and host tissue invasion.23,24 Among these, SAP1 and SAP2 have been extensively characterized and shown to differ in both kinetic behavior and pathogenic significance. In particular, SAP2 exhibits a higher association rate constant (kₒₙ) and stronger affinity toward acetyl-pepstatin than SAP1, reflecting its greater efficiency in inhibitor binding and proteolytic activity.25 Despite these detailed insights into SAP1-SAP6, the structural and inhibitory mechanisms of SAP5 have not been adequately explored. Given SAP5’s well-established association with biofilm formation, hyphal transition, and antifungal resistance, the present study seeks to bridge this gap by conducting computational docking and pharmacokinetic analyses of A. rivularis derived phytometabolites against SAP5. This investigation provides the first in silico evidence highlighting SAP5 as a viable molecular target for natural antifungal agents and extends current understanding beyond the traditionally studied SAP isoforms.
Several plant-derived metabolites, including apigenin, baicalein, curcumin, quercetin, resveratrol, silymarin, and taxifolin, have been previously reported to inhibit C. albicans biofilm formation by interfering with adhesion, hyphal development, and virulence pathways.26,27 Extracts from plants such as Schinus terebinthifolius, Croton urucurana, and Syzygium aromaticum have also demonstrated potent antibiofilm activity due to their rich content of alkaloids, terpenoids, and phenolic compounds.28,29 A recent computational study identified homovanillic acid as a promising SAP5 inhibitor in Candida albicans biofilms, demonstrating high drug-likeness, low toxicity, and stable protein-ligand interactions through molecular docking, MM/GBSA analysis, and 100 ns molecular dynamics simulations, reflecting the value of in silico approaches in antifungal drug discovery and AMR biofilm management.30 In contrast to such earlier reports, the present study explores a unique medicinal plant, Astilbe rivularis, which has not been previously evaluated against C. albicans. The computational analysis of A. rivularis phytometabolites against the biofilm-associated enzyme SAP5 introduces a novel plant source and highlights its potential as a natural reservoir of SAP5 inhibitors for disrupting Candida biofilm virulence mechanisms.
Several of the phytometabolites represented in our set (e.g., quercetin, β-sitosterol, bergenin-like phenolics) have independent literature support for anti-Candida or anti-biofilm actions. Quercetin has repeatedly been shown to reduce biofilm formation, hyphal development, and the secretion of hydrolases, and it can sensitize fluconazole-resistant strains-mechanistic themes consistent with targeting virulence pathways, such as SAPs.31 Likewise, β-sitosterol has shown antibiofilm activity, with molecular docking and mechanistic work implicating ergosterol pathway interference and broader virulence attenuation.32 These reports enhance the translational plausibility of our in silico hits.
Secreted aspartyl proteases are well-recognized drivers of tissue invasion, immune evasion, and biofilm maturation in C. albicans.33,34 Clinically, C. albicans biofilms display significant tolerance to triazole antifungals such as fluconazole, primarily due to matrix sequestration, altered sterol regulation, and efflux pump activation. Plant-derived phytochemicals that target virulence mechanisms such as SAP-mediated proteolysis, hyphal transition, and adhesion offer a complementary approach by disrupting biofilm integrity rather than merely inhibiting fungal growth.35,36 Previous studies demonstrating quercetin-fluconazole synergy in biofilm inhibition further support the potential of combining natural metabolites with conventional antifungals.37 Proteolysis within biofilms has been proposed as both a marker and a therapeutic entry point, and SAP5 is preferentially expressed during hyphal/biofilm phases. By selecting SAP5 as the molecular target, the present study aligns with emerging research that focuses on virulence attenuation as a resistance-sparing therapeutic strategy, emphasizing SAP enzymes as key regulators of pathogenicity rather than direct fungicidal targets.
Limitations of the study
While the present in silico investigation provides strong preliminary evidence for the antifungal potential of A. rivularis phytometabolites against C. albicans SAP5, it is not without limitations. The docking analysis, though predictive, does not account for dynamic conformational changes in the protein-ligand complex under physiological conditions. The lack of molecular dynamics (MD) simulations restricts a deeper understanding of complex stability, solvent interactions, and time-dependent flexibility. Moreover, the in silico ADME and toxicity predictions, while useful may differ from actual pharmacokinetic behaviour in biological systems. The absence of experimental validation such as enzyme inhibition assays, biofilm quantification, and cytotoxicity studies limits the immediate translational applicability of these findings. Additionally, potential synergistic or antagonistic interactions among multiple phytometabolites within the plant extract were not assessed, which may influence overall efficacy in vivo.
Future scope
Future work should focus on validating the present in silico findings through molecular dynamics simulations to assess the stability of the SAP5 ligand complexes under physiological conditions, followed by in vitro enzyme inhibition and biofilm assays to confirm antifungal efficacy. Comparative evaluation with standard drugs such as fluconazole can determine synergistic potential, while cytotoxicity and hemolytic studies will help establish safety profiles. Incorporating A. rivularis phytometabolites into nanocarrier-based delivery systems or green-synthesized nanoparticles may further enhance bioavailability and therapeutic effectiveness. These approaches will collectively strengthen the translational potential of A. rivularis-derived compounds as sustainable antifungal agents targeting Candida albicans biofilm-associated virulence.
The present study highlights the antifungal potential of bioactive phytometabolites derived from A. rivularis against the C. albicans virulence enzyme SAP5, a key determinant in biofilm formation and pathogenicity. Molecular docking revealed strong and stable binding affinities, supported by favourable ADME and drug-likeness profiles, suggesting that these metabolites could effectively inhibit SAP5-mediated proteolytic activity. The interaction network and physicochemical evaluations further emphasized SAP5 as a promising antifungal target and positioned A. rivularis metabolites as viable natural inhibitors. Overall, this study provides a computational foundation for developing plant-based antifungal therapeutics and encourages further in vitro and in vivo investigations to validate their efficacy and safety for combating Candida albicans biofilm-associated infections.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
DSS conceptualized the study. SPS collected resources. DSS and NK applied methodology. SPS performed supervision, validation and project administration. DSS performed data curation, formal analysis, and visualization. NK performed investigation and data interpretation. DSS wrote the original draft. SPS and NK wrote and reviewed the manuscript. SPS, DP and NK revised the manuscript. DP approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Lopes JP, Lionakis MS. Pathogenesis and virulence of Candida albicans. Virulence. 2022;13(1):89-121.
Crossref - Schille TB, Sprague JL, Naglik JR, Brunke S, Hube B. Commensalism and pathogenesis of Candida albicans at the mucosal interface. Nat Rev Microbiol. 2025;23(8):525-540.
Crossref - Pereira R, Dos Santos Fontenelle RO, de Brito EHS, de Morais SM. Biofilm of Candida albicans: formation, regulation and resistance. J Appl Microbiol. 2021;131(1):11-22.
Crossref - Dohlman AB, Klug J, Mesko M, et al. A pan-cancer mycobiome analysis reveals fungal involvement in gastrointestinal and lung tumors. Cell. 2022;185(20):3807-3822.e12.
Crossref - Fang W, Wu J, Cheng M, et al. Diagnosis of invasive fungal infections: challenges and recent developments. J Biomed Sci. 2023;30(1):42.
Crossref - Bongomin F, Gago S, Oladele RO, Denning DW. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J Fungi. 2017;3(4):57.
Crossref - Lass-Florl C, Kanj SS, Govender NP, Thompson GR 3rd, Ostrosky-Zeichner L, Govrins MA. Invasive candidiasis. Nat Rev Dis Primers. 2024;10(1):20.
Crossref - Ivanov M, Ciric A, Stojkovic D. Emerging Antifungal Targets and Strategies. Int J Mol Sci. 2022;23(5):2756.
Crossref - Roe K. Treatment alternatives for multidrug-resistant fungal pathogens. Drug Discov Today. 2023;28(6):103596.
Crossref - Szczygiel T, Kozirog A, Otlewska A. Synthetic and Natural Antifungal Substances in Cereal Grain Protection: A Review of Bright and Dark Sides. Molecules. 2024;29(16):3780.
Crossref - Bouhtit F, Najar M, Rahmani S, et al. Bioscreening and pre-clinical evaluation of the impact of bioactive molecules from Ptychotis verticillata on the multilineage potential of mesenchymal stromal cells towards immune- and inflammation-mediated diseases. Inflamm Res. 2022;71(7-8):887-898.
Crossref - Szymanski M, Chmielewska S, Czyżewska U, Malinowska M, Tylicki A. Echinocandins – structure, mechanism of action and use in antifungal therapy. J Enzyme Inhib Med Chem. 2022;37(1):876-894.
Crossref - Hori K, Wada M, Yahara S, Watanabe T, Devkota HP. Antioxidant phenolic compounds from the rhizomes of Astilbe rivularis. Natural Prod Res. 2018;32(4):453-456.
Crossref - Rai V, Pogu SV, Bhatnagar R, et al. Biological evaluation of a natural steroid ester, Stigmasta-5 (6), 22 (23)-dien-3-beta-yl acetate isolated from the Himalayan herb Astilbe rivularis as potential antitumor agent. Chem Biol Interact. 2022; 360:109935.
Crossref - Tiwary BK, Ghosh R, Moktan S, et al. Prospective bacterial quorum sensing inhibitors from Indian medicinal plant extracts. Lett Appl Microbiol. 2017;65(1):2-10.
Crossref - Kulshrestha A, Gupta P. Secreted aspartyl proteases family: a perspective review on the regulation of fungal pathogenesis. Future Microbiol. 2023;18(5):295-309.
Crossref - Dhanasekaran S, Selvadoss PP, Manoharan SS. Anti-fungal potential of structurally diverse FDA-approved therapeutics targeting secreted aspartyl proteinase (SAP) of Candida albicans: an in silico drug repurposing approach. Appl Biochem Biotechnol. 2022;195(3):1983-1998.
Crossref - Vivek-Ananth RP, Mohanraj K, Sahoo AK, Samal A. IMPPAT 2.0: An Enhanced and Expanded Phytochemical Atlas of Indian Medicinal Plants. ACS Omega. 2023;8(9):8827-8845.
Crossref - Berman HM, Westbrook J, Feng Z, et al. The Protein Data Bank. Nucleic Acids Res. 2000;28(1):235-242.
Crossref - Rao VS, Srinivas K, Sujini GN, Kumar GN. Protein-protein interaction detection: methods and analysis. Int J Proteomics. 2014;2014:147648.
Crossref - Daina A, Michielin O, Zoete V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci Rep. 2017;7(1):42717.
Crossref - Karami TK, Hailu S, Feng S, Graham R, Gukasyan HJ. Eyes on Lipinski’s Rule of Five: A New “Rule of Thumb” for Physicochemical Design Space of Ophthalmic Drugs. J Ocul Pharmacol Ther. 2022;38(1):43-55.
Crossref - Koelsch G, Tang J, Loy JA, et al. Enzymic characteristics of secreted aspartic proteases of Candida albicans. Biochim Biophys Acta. 2000;1480(1-2):117-131.
Crossref - Kozik A, Gogol M, Bochenska O, et al. Kinin release from human kininogen by 10 aspartic proteases produced by pathogenic yeast Candida albicans. BMC Microbiol. 2015;15(1):60.
Crossref - Backman D, Danielson UH. Kinetic and mechanistic analysis of the association and dissociation of inhibitors interacting with secreted aspartic acid proteases 1 and 2 from Candida albicans. Biochim Biophys Acta. 2003;1646(1-2):184-195.
Crossref - Al Aboody MS, Mickymaray S. Anti-Fungal Efficacy and Mechanisms of Flavonoids. Antibiotics. 2020;9(2):45.
Crossref - Lee JH, Kim YG, Park I, Lee J. Antifungal and antibiofilm activities of flavonoids against Candida albicans: Focus on 3,2’-dihydroxyflavone as a potential therapeutic agent. Biofilm. 2024;8:100218.
Crossref - Barbieri DSV, Tonial F, Lopez PV, et al. Antiadherent activity of Schinus terebinthifolius and Croton urucurana extracts on in vitro biofilm formation of Candida albicans and Streptococcus mutans. Arch Oral Biol. 2014;59(9):887-96.
Crossref - El-Saber Batiha G, Alkazmi LM, Wasef LG, Beshbishy AM, Nadwa EH, Rashwan EK. Syzygium aromaticum L. (Myrtaceae): Traditional Uses, Bioactive Chemical Constituents, Pharmacological and Toxicological Activities. Biomolecules. 2020;10(2):202.
Crossref - Kulshrestha A, Gupta P. Multi-computational screening identifies homovanillic acid as a potential SAP5 inhibitor against Candida albicans biofilms. Comput Biol Chem. 2025;118:108453.
Crossref - Singh BN, Upreti DK, Singh BR, et al. Quercetin sensitizes fluconazole-resistant Candida albicans to induce apoptotic cell death by modulating quorum sensing. Antimicrob Agents Chemother. 2015;59(4):2153-2168.
Crossref - Mohana P, Singh A, Rashid F, et al. Inhibition of Virulence Associated Traits by β-Sitosterol Isolated from Hibiscus rosa-sinensis Flowers Against Candida albicans: Mechanistic Insight and Molecular Docking Studies. J Microbiol. 2024;62(12):1165-1175.
Crossref - Mitthun ANK, Smiline GAS, Vijayashree PJ, Gunasekaran S, Kannan KP. Detection of sap in Candida albicans and Targeting with Bioactive Compounds from Aegle marmelos: An in vitro and in silico Analysis. J Mycol Infect. 2025;30(1):25-34.
Crossref - Padmanabhan S, Palanimuthu UM, Raghavan M, Kumar P, Mohanram K. Detection of Secreted Aspartic Proteases (SAP) enzyme in the clinical isolates of Candida by Modified Stab Method. IP Int J Med Microbiol Trop Dis. 2024;10(2):169-173.
Crossref - Sanjana, Priyadharshini R, Rajeshkumar S, Anandan J. Evaluation of Antimicrobial and Cytotoxic Effects of Eclipta prostrata-mediated Calcium Oxide Nanoparticle-incorporated Mouth Rinse. J Environ Nanotechnol. 2025;14(1):63-68.
Crossref - Priyadharshini SS, Ragavendran C, Sherwood A, Piriyanga R. Antimicrobial efficacy of carbonated hydroxyapatite against Streptococcus mutans, Enterococcus faecalis, and Candida albicans: an in vitro study. Avicenna J Dent Res. 2024;16(2):90-96.
Crossref - Janeczko M, Gmur D, Kochanowicz E, Gorka K, Skrzypek T. Inhibitory effect of a combination of baicalein and quercetin flavonoids against Candida albicansstrains isolated from the female reproductive system. Fungal Biol. 2022;126(6-7):407-420.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.