


ISSN: 0973-7510
E-ISSN: 2581-690X
The filamentous fungi Trichoderma is used asmost important biocontrol agent. Because of the absence of sexual reproduction in this fungus, other methods of genetic improvement have been developed to enhance its bicontrol potential. In the present investigation protoplast fusion of four different Trichoderma spp. was evaluated for their antagonistic activity against the soil borne plant pathogen like Fusarium spp. which causes wilting diseases in many crops. In protoplast fusion technique, protoplast isolation by lysis enzyme at the concentration of 10 mg/ml, protoplasts were fused intra and inter specifically, with 30% (w/v) PEG solution and those fusants regenerated on osmatic stabilizer amended PDA media. Eight self fusants and eighteen fusants were selected for assessment of antagonastic activity against fusarium spp. in dual culture technique. The results revealed that among eight intra fusants, fusant Tvdsf-2 (93.70%) was recorded highest per cent inhibition against F. o. f. sp. udum and Tvssf-1 (86.70%) against the F. solani and inter specific fusants, the maximum per cent inhibition mycelia growth was observed in Tvs×Tas-1 (94.07%) and Tvd×Tas-1 (94.07%) against F. o. f. sp. udumand Tvd×Tas-1 (89.63%) against the F. solani. These outcomes indicated that protoplast fusion technique was successful tools to enhance the antagonistic effects of Trichoderma species against several fungal plant pathogens.
Trichoderma spp., protoplast fusion, regeneration and antagonism.
Soil borne pathogens have a broad host range and persist for longer periods in soil by resistant resting structures. Chemical control of soil borne pathogens provides certain degree of control but at the same time have adverse effects on environment affecting the beneficial soil microorganisms. Therefore, biological control of plant pathogens has been considered as a potential control strategy in recent years and search for these biological agents is increasing. Trichoderma is the most commonly used fungal biological control agent and have long been known as effective antagonists against plant pathogenic fungi (Papavizas, 1985).
Soil borne diseases caused Fusarium spp. which was causes economical loss in many crops. Pesticide treatment is the most usual method for controlling these diseases. However, these pesticides are expensive and are harmful on human health. Therefore, using biocontrol agents are more effective, less expensive and safer for human health. Genus Trichoderma is one of the most important filamentous fungi used as a biocontrol agent. Many species under this genus has been used against diseases in a wide variety of economically important crops. Because of the absence of sexual reproduction, other methods of genetic improvement have been developed in these fungi such as transformation and proptoplast fusion (Balasubramanian and Lalithakumari, 2008).
The present work aimed to apply protoplast fusion of four Trichoderma spp. to enhancement their bio-control abilities against some of important plant fungal pathogens (Fusarium spp.), which cause wilting diseases that attacking different crops.
The mutation was carried out by using four Trichoderma species viz., Trichodermaharzianum, Trichodermavirens and Trichodermaviride isolates were collected from NBAIR (National Bureau of Agricultural Insects Resources) Bengaluru and Trichodermaasperellum from IIOR (Indian Institute of Oil Research) Hyderabad andFusariumoxysporum f. sp. udum and Fusariumsolani, the causal agent of wilt diseases in pigeon pea and acid lime were isolated from diseased plants. Those bio-agent and pathogenic fungi was sub cultured on PDA slants and allowed to grow at 28±1°C for ten days and such slants were preserved in a refrigerator. The procedure or protocol followed for the isolation of protoplast, fusion of protoplasts is given below.
Isolation of protoplasts from Trichoderma species
The parental strains of Trichoderma species like Trichoderma harzianum, Trichoderma virens, Trichoderma viride and Trichoderma asperellum were grown separately in 50 ml of PDB in 150 ml Erlenmeyer flasks by inoculating 1 ml conidial suspension of respective culture at H”5´106 conidia/ml. The flasks were incubated on a rotary shaker with a speed of 120 rpm at room temperature for 15 hours. Then cultures were harvested and the young mycelia were separated by filtration using Whitman No. 1 filter paper. About 100 mg of fresh mycelium was washed with sterile distilled water followed by 0.1 M phosphate buffer (pH 5.8) and osmatic stabilizer (0.7 M KCl) then incubated with Lysing Enzymes (Sigma Chemicals Co.) at 10 mg/ml concentration prepared in phosphate buffer containing 0.7 M KCl as osmotic stabilizer. The mycelia–enzyme mixtures were incubated on a rotatory shaker with a speed of 75 rpm at room temperature and the lysis of fungal cell wall and the release of protoplasts were monitored at 30 min interval under a light microscope. After 4hr, the enzyme-protoplast mixture was filtered through a sterile cotton swab and centrifuged at 3000 rpm for 5 min at 4°C. The supernatant was discarded and the protoplast sediment was suspended immediately in buffer and osmotic stabilizer solution. These protoplasts were used for further fusion process.
Intra and inter-specific protoplast fusion
Intra and Inter-specific protoplast fusion was carried out by the method given by Prabavathy et al. (2006). Intra specific fusion was done within the species, such as, Trichoderma harzianum (Th×Th), T. virens, (Tvs×Tvs), T. viride (Tvd×Tvd) and T. asperellum (Tas×Tas). In case of inter specific fusion, on different combination of Trichoderma was fused with the another species of Trichoderma (Th×Tvs, Th×Tvd, Th×Tas, Tvs×Tvd, Tvs×Tas and Tvd×Tas).
Table List of regenerated Intra-specific fused protoplasts
Sl. No. |
Trichoderma spp. |
Name of regenerated isolates |
|---|---|---|
1 |
Trichoderma harzianum |
Thsf-1 & Thsf-2 |
2 |
Trichoderma viride |
Tvssf-1 & Tvssf-2 |
3 |
Trichoderma virens |
Tvdsf-1 & Tvdsf-2 |
4 |
Trichoderma asperellum |
Tassf-1 & Tassf-2 |
Note: Thsf– Trichoderma harzianum self fusant Tvssf– Trichoderma virens self fusant
Tvdsf– Trichoderma viride self fusantTassf-Trichoderma asperellum self fusant
Table List of regenerated Intra-specific fused protoplasts
Sl. No. |
Trichoderma spp. |
Name of regenerated isolates |
|---|---|---|
1 |
Trichoderma harzianum x Trichoderma viride |
Th×Tvs-1, Th×Tvs-2, Th×Tvs-3 |
2 |
Trichoderma harzianum x Trichoderma virens |
Th×Tvd-1, Th×Tvd-2, Th×Tvd-3 |
3 |
Trichoderma harzianum x Trichoderma asperellum |
Th×Tas-1, Th×Tas-2, Th×Tas-3 |
4 |
Trichoderma virens x Trichoderma viride |
Tvs×Tvd-1, Tvs×Tvd-2, Tvs×Tvd-3 |
5 |
Trichoderma virens x Trichoderma asperellum |
Tvs×Tas-1, Tvs×Tas-2, Tvs×Tas-3 |
6 |
TrichodermaviridexTrichodermaasperellum |
Tvd×Tas-1, Tvd×Tas-2, Tvd×Tas-3 |
Polyethylene glycol (PEG), (MW 6000, Himedia company) prepared in STC buffer contained 0.6 M Sorbitol, 10 mM Tris-HCl and 10 mM Calcium chloride (pH 7.5) was used as fusogen. One ml of protoplasts suspension (1×106 protoplasts/ml) was mixed with an equal volume of 30% PEG solution and the fusion mixture was incubated at room temperature. After 10 min, the mixture was diluted with 1 ml of STC buffer. The protoplast fusion was observed under microscope (Axiozeiss Company) and microphotographs were taken.
The fusion mixture was serially diluted with the osmotic stabilizer for regeneration and plated on regeneration medium (PDA amended with osmotic stabilizer) and controls were maintained separately for non-fusions. Observations were made every day on regeneration of fusion protoplasts on agar media. After regeneration of protoplasts on media, all the growing fusants were selected based on the fast growth comparing with the parent isolates as check and grown separately on the PDA plates, Selected regenerated intra and inter specific fused protoplasts were enlisted as below.
The antagonistic effects of each intra and inter specific protoplast fusions of different Trichoderm spp. and their parental Trichoderma spp. isolates were evaluated against Fusarium oxysporum f. sp. udum and Fusarium solani were evaluated in dual culture plate technique. The percent inhibition of the growth of the pathogen was calculated by using the formula suggested by Vincent (1947)
Percent inhibition in mycelial growth/conidial germination = C – T × 100
C
Where, C = Mycelial growth/conidial germination in control
T = Mycelial growth/conidial germination in treatment
Protoplast fusion has rapidly become acceptable tools for the fungal genetics with the development of new techniques of gene transfer such as protoplast fusion and enhancement of antagonistic ability of Trichoderma spp. byprotoplast fusion. These experiments included protoplast isolation, regeneration and intra and inter specific fusions. In addition, evaluation of obtained fusants by dual culture experiment.
Protoplast isolation fusion and regeneration
Isolation of protoplast, first completely lysed the young mycelium and release of protoplasts were observed after 3 hour (Fig.1 to 4). Initially released protoplasts were smaller in size, but later enlarged and formed spherical structures. The protoplasts yield significantly affected by the concentrations of lysing enzymes and specific conditions including mycelium age, temperature, medium, etc.. The conditions for releasing the protoplasts were similar as reported by EL-Bondkly (2006).
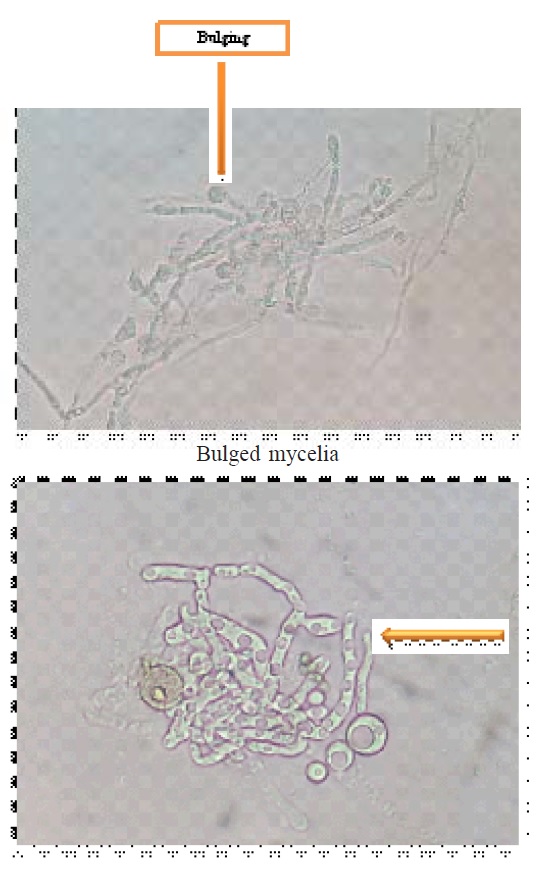 Fig. 1. Protoplast releasing from young mycelia
Fig. 1. Protoplast releasing from young mycelia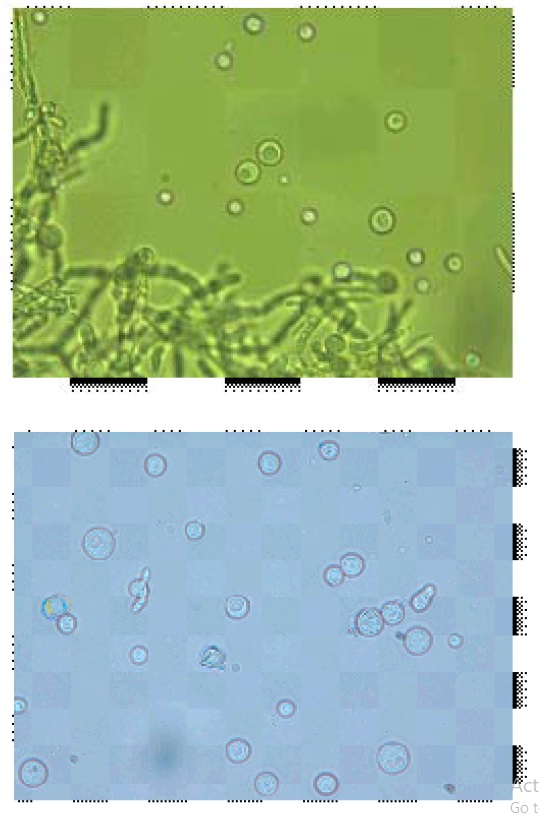 Fig. 2. Released protoplast free from mycelia
Fig. 2. Released protoplast free from mycelia Fig. 3. Protoplast fusion
Fig. 3. Protoplast fusion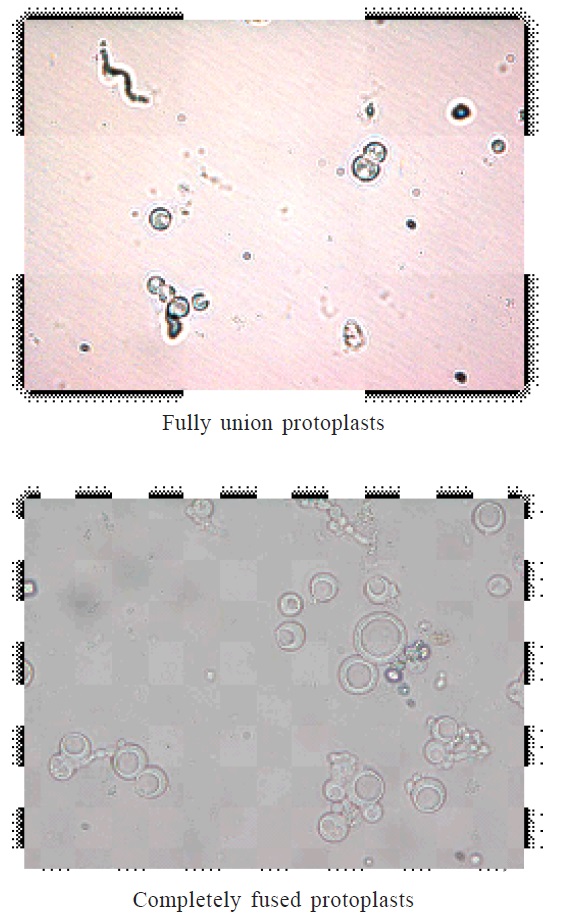 Fig. 4. Protoplats fusion
Fig. 4. Protoplats fusionThe lysis of fungal mycelium was confined only to the tip portion, 10 mg/ml concentrations of lysing enzymes with 0.7 M KCl as osmotic stabilizer was optimal for the release of protoplasts. Peåer and Chet (1990) obtained highest protoplasts yield from T. harzianum using Novozyme 234 at 10 mg/ml with 0.6 M KCl. The isolated protoplasts were subjected to intra and inter specific protoplast fusion with the use of 30% PEG 6000. The protoplasts were attracted to each other’s and pairs of protoplasts were observed when PEG solution was added to the protoplasts. Later, the plasma membrane at the place of contact dissolved and protoplasmic contents fused together that may half union and fully fusion (fig 9 and 10), then followed by nuclear fusion in most cases.
The fused protoplasts became single, larger and round or oval-shaped structure representing in fig 10. As similar concentration of PEG was reported as optimum for inter-specific fusion of protoplasts between T. harzianum and T. reesei, as well as intergeneric protoplast fusion between T. harzianum and A. niger (ELBondkly, 2002; EL-Bondkly, 2006). However, Peåer and Chet (1990) used 33 % PEG for Protoplast fusion in T. harzianum.
Finally regeneration the fused protoplasts on PDA medium supplemented with osmatic stabilizer was done. Out of number of regenerated colony forming units on PDA media, based on fast growth, eight intra specific and 18 inter specific protoplast fusions were selected. They were subjected to their antagonistic activity evaluation along with the parent isolates against the soil borne pathogens like F. oxysporum f. sp.udum and F. solani.
Antagonistic activity of intra and inter fusants in dual culture
Evaluaton of antagonistic activity in intra specific fusions was observed against F. oxysporum f. sp. udum and F. solani, as compared to their parental isolates in below following tables. Among eight fusants Tvdsf-2 (93.70%) recorded the highest per cent inhibition (fig 6), followed by Thsf-2 (91.85%), Tvssf-2 (90.74%), Tvssf-1(90.00%) and Tvdsf-1 (89.63 %) against F. oxysporum f. sp. udum. The least inhibition was noticed in Tassf-1 (88.15 %) and Thsf-1 (88.52 %). Tvdsf-2 and Thsf-2 recorded more per cent inhibition against F. oxysporum f. sp. udum than the parent isolates (Table.1). Highest per cent inhibition against Fusarium solani observed in fusantTvssf-1 recorded 86.70 per cent (fig. 5), followed by Thsf-1 (85.9%), Tvssf-2 (85.6%), Thsf-2 (85.2%) and Tvdsf-2 (84.4%) against Fusarium solani. The least inhibition (82.2%) was observed in Tvdsf-1 and Tassf-2 against Fusarium solani (Table. 2).These results are in agreement with findings of Prabavathy et al. (2006) noticed the improvement of antagonistic activity in self-fusions. Maximum mycelial growth of Rhizoctonia solani inhibited self-fusions (SFTh2, SFTh8, SFTh10, SFTh12 and SFTh13) as compared to the parent strain (67.6%).
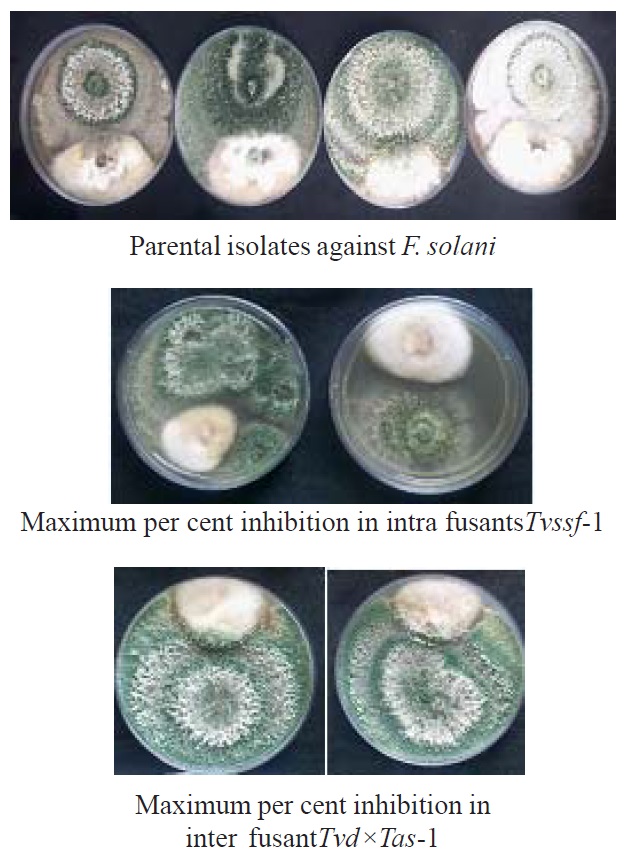 Fig. 5. Dual culture plates of parents and fusants against F. solani
Fig. 5. Dual culture plates of parents and fusants against F. solaniTable (1):
In vitro evaluation of intra specific fusions against Fusarium oxysporum f. sp. udum.
Sl. No. |
Parents and Fusants |
Mean per cent inhibition |
|---|---|---|
1 |
PTh |
85.56*(67.65) |
2 |
PTvs |
85.55(67.64) |
3 |
PTvd |
91.11(72.79) |
4 |
PTas |
86.67(68.57) |
5 |
Thsf-1 |
88.52(70.20) |
6 |
Thsf-2 |
91.85(73.50) |
7 |
Tvssf-1 |
90.00(71.55) |
8 |
Tvssf-2 |
90.74(72.35) |
9 |
Tvdsf-1 |
89.63(71.29) |
10 |
Tvdsf-2 |
93.70(75.48) |
11 |
Tassf-1 |
88.15(69.87) |
12 |
Tassf-2 |
89.26(70.85) |
S. Em± |
1.04 |
|
C.D. @1% |
4.12 |
*Arc sine values
Table (2):
In vitro evaluation of intra specific fusions against Fusarium solani.
Sl. No. |
Parents and Fusants |
Mean per cent inhibition |
|---|---|---|
1 |
PTh |
78.5*(62.38) |
2 |
PTvs |
82.2(65.04) |
3 |
PTvd |
85.2(67.36) |
4 |
PTas |
78.9(62.63) |
5 |
Thsf-1 |
85.9(67.98) |
6 |
Thsf-2 |
85.2(67.36) |
7 |
Tvssf-1 |
86.7(68.57) |
8 |
Tvssf-2 |
85.6(67.71) |
9 |
Tvdsf-1 |
82.2(65.04) |
10 |
Tvdsf-2 |
84.4(66.75) |
11 |
Tassf-1 |
82.6(65.34) |
12 |
Tassf-2 |
82.2(65.04) |
S. Em± |
0.78 |
|
C.D. @1% |
3.09 |
*Arc sine values
In case of inter specific fusions, the maximum mycelial growth was inhibited by fusant Tvs×Tas-1 (94.07%) and Tvd×Tas-1 (94.07%) represented in figure 6, followed by fusantTvs×Tvd-2 (93.70%) which was on par with other fusants Th×Tas-1, Th×Tas-2, Th×Tas-3, Tvs×Tvd-1, Tvs×Tvd-3, Tvd×Tas-2 and Tvd×Tas-3. Fusants were found better to their parental strain, in inhibiting the maximum mycelial growth of F. oxysporum f. sp. udum, except in parent T. viride- PTvd (91.11%) recorded maximum growth inhibition more than some of the fusants (Table. 3).
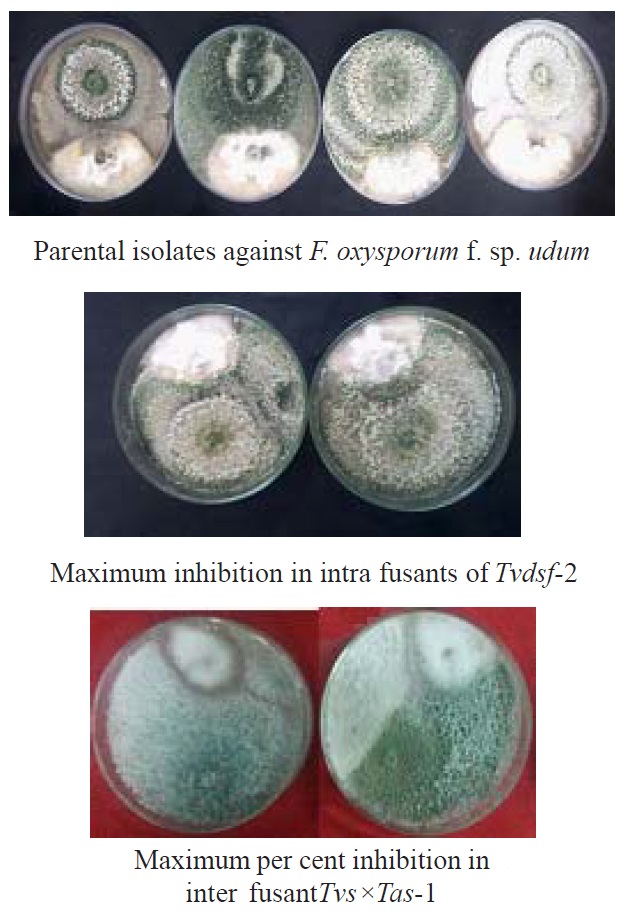 Fig. 6. Dual culture plates of parents and fusants against F. o. f. sp. udum
Fig. 6. Dual culture plates of parents and fusants against F. o. f. sp. udum
Table (3):
In vitro evaluation of inter specific fusions against F. o. f. sp. udum.
Sl. No. |
Parents and Fusants |
Mean per cent inhibition |
|---|---|---|
1 |
PTh |
85.56*(67.65) |
2 |
PTvs |
85.55(67.64) |
3 |
PTvd |
91.11(72.79) |
4 |
PTas |
86.67(68.57) |
5 |
Th×Tvs-1 |
90.00(71.55) |
6 |
Th×Tvs-2 |
90.00(71.67) |
7 |
Th×Tvs-3 |
87.41(69.21) |
8 |
Th×Tvd-1 |
87.78(69.52) |
9 |
Th×Tvd-2 |
87.04(69.02) |
10 |
Th×Tvd-3 |
88.52(70.23) |
11 |
Th×Tas-1 |
92.59(74.37) |
12 |
Th×Tas-2 |
90.74(72.35) |
13 |
Th×Tas-3 |
91.11(72.71) |
14 |
Tvs×Tvd-1 |
92.59(74.37) |
15 |
Tvs×Tvd-2 |
93.70(75.48) |
16 |
Tvs×Tvd-3 |
90.00(71.75) |
17 |
Tvs×Tas-1 |
94.07(75.97) |
18 |
Tvs×Tas-2 |
87.78(69.52) |
19 |
Tvs×Tas-3 |
88.52(70.25) |
20 |
Tvd×Tas-1 |
94.07(76.34) |
21 |
Tvd×Tas-2 |
91.48(73.05) |
22 |
Tvd×Tas-3 |
91.85(73.50) |
S. Em± |
1.39 |
|
C.D. @1% |
5.31 |
*Arc sine values
Similarly against F. solani, the maximum per cent inhibition of mycelial growth in F. Solani was observed in the fusantTvd×Tas-1 (89.63%) represented in figure 5, followed by Tvs×Tvd-2(89.26%), Tvs×Tvd-3 (87.78%), Tvs×Tvd-1 (87.41%), Tvs×Tas-1 (87.04%), Th×Tvd-2 (86.30%) and Th×Tas-1 (86.30%) they were on par with fusantTvd×Tas-1. But many fusants exhibited better results than that of the parental isolate (Table.4).
Table (4):
In vitro evaluation of inter specific fusions against Fusarium solani.
Sl. No. |
Parents and Fusants |
Mean per cent inhibition |
|---|---|---|
1 |
PTh |
78.52*(62.38) |
2 |
PTvs |
82.22(65.04) |
3 |
PTvd |
85.18(67.36) |
4 |
PTas |
78.89(62.63) |
5 |
Th×Tvs-1 |
80.74(63.95) |
6 |
Th×Tvs-2 |
83.33(65.95) |
7 |
Th×Tvs-3 |
82.96(65.64) |
8 |
Th×Tvd-1 |
85.18(67.36) |
9 |
Th×Tvd-2 |
86.30(68.27) |
10 |
Th×Tvd-3 |
84.44(66.75) |
11 |
Th×Tas-1 |
86.30(68.27) |
12 |
Th×Tas-2 |
83.33(65.88) |
13 |
Th×Tas-3 |
81.11(64.22) |
14 |
Tvs×Tvd-1 |
87.41(69.19) |
15 |
Tvs×Tvd-2 |
89.26(71.16) |
16 |
Tvs×Tvd-3 |
87.78(69.56) |
17 |
Tvs×Tas-1 |
87.04(68.90) |
18 |
Tvs×Tas-2 |
82.22(65.04) |
19 |
Tvs×Tas-3 |
82.96(65.61) |
20 |
Tvd×Tas-1 |
89.63(71.21) |
21 |
Tvd×Tas-2 |
85.55(67.64) |
22 |
Tvd×Tas-3 |
82.59(65.33) |
S. Em± |
0.94 |
|
C.D. @1% |
3.57 |
*Arc sine values
These results are in line with the findings of Hassan Abdel-Latif and Haggag (2010) who described that evaluation of inter specific fusion of Trichoderma koningii and Trichoderma reesei, against the four fungal pathogens like Fusarium oxysporum, Pythium ultimum, Sclerotia rolfsii and Sclerotinia sclerotiorum indicated the effectiveness in suppressing the mycelial growth more than the parental isolates. Similar kind of results have also revealed by Migheliet al. (2008) who noticed that the protoplast fusants have higher bio-control potential against Botrytis cinerea, Sclerotinia sclerotiorum and Pythium ultimum.
In summary, the intra and inter-fusion of protoplasts in Trichoderma spp. resulted in appreciable increase of bio-control activity in most of the fusants. Results revealed the scope and significance of the protoplast fusion technique, which could successfully be used to develop superior hybrid strains in filamentous fungi that lack sexual reproduction.
- Balasubramanian, N. and Lalithakumari, D., Characteristics of protoplast inter, intra-fusant and regeneration of antagonistic fungi Trichoderma harzianum and Trichoderm aviride. African J. Biotechnol., 2008; 7(18):3235-3243.
- EL-Bondkly, A. M. Genetic transformation in Trichodermareesei for the improvement of cellulase production.Ph.D., Thesis,Kafr EL-Sheikh, TantaUniversity 2002.
- EL-Bondkly, A. M. Gene transfer between different Trichoderma species and Aspergillus niger through intergeneric protoplast fusion to convert ground rice straw to citric acid and cellulases. Appl. Biochem. Biotechnol., 2006; 135(2): 117-132.
- El-Bondkly, A. M., Gene transfer between different Trichoderma species and Aspergillus niger through intergeneric protoplast fusionto convert ground rice strawto citric acid and cellulases.Appl.Biochem. Biotechnol., 2006; 135: 117-132.
- Hassan Abdel-Latif A. and Haggag, W. M., Mutagenesis and inter-specific protoplast fusion between Trichoderma koningii and Trichoderma reesei for bio control improvement. American J. Scientific Indust.Res, 2010; pp.504-515.
- Migheli, Q., Whipps, J. M., Budge, S. and Lynch, J., Production of inter and intra hybrids of Trichoderma spp. by protoplast fusion and evaluation of their bio control activity against soil-borne and foliar pathogens. J. Phytopathol., 2008; 143: 91-97.
- Papavizas, G. C., Trichoderma and Gliocladium biology, ecology and potential for biocontrol. Ann. Rev. Physiopathol., 1985; 23:23-54.
- Peer, S. and Chet, I., Trichoderma protoplast fusion: a tool for improving bio control agents. Canadian J. Microbiol., 1990; 36:6-9.
- Prabavathy, V. R., Mathivanan, N., Sagadevan, E., Murugesan, K. and Lalithakumari, D., Intra-strain protoplast fusion enhances carboxy methyl cellulase activity in Trichoderma reesei. Enz.Microb. Technol., 2006; 38: 719–723.
- Vincent, J. M., Distortion of fungal hyphae in the presence of certain inhibitors. Nature, 1947; 159: 850-851.
© The Author(s) 2017. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.