


ISSN: 0973-7510
E-ISSN: 2581-690X
Exserohilum turcicum is an emerging pathogen of maize causing the northern leaf blight (NLB) disease with severe losses. The disease prevails all over the area under production but has varying severity. A temperature of 13°C to 33°C, along with 75-84% relative humidity, favours the pathogen development and growth. Thus, the present investigation was carried out to assess the prevalence of the disease in Bihar and the cultural, morphological, and pathogenic variability among the population through extensive field surveys conducted in maize-growing areas of seven major maize-producing districts of Bihar, India. A total of twenty-one isolates were identified as E. turcicum, which showed varying colony characteristics, colony margin, margin colour, pigmentation, growth pattern, sporulation, conidial shape, conidial size, and conidial septa. However, all the isolates produced characteristic symptoms of cigar-shaped necrotic lesions with variation in disease reaction on the leaves of susceptible maize inbred line CM-202. The isolates were categorized into three phylogenetic clusters with a coefficient of 0.42 based on their variations. A similar trend was observed, and three clusters with a coefficient of 0.52 were confirmed based on the disease reaction under field conditions. The isolates constituting Cluster I were deemed high virulent, followed by Cluster II as moderate virulent, and Cluster III as low virulent. All the isolates were further identified as Setosphaeria turcica, a teleomorph stage of E. turcicum, through molecular identification using ITS sequence analysis. Conclusively, a higher variability among the pathogen population was present, thus leading to the disease emergence, and their virulence assessment will aid in the identification of races and simultaneously accelerate the resistance breeding programmes in maize.
Maize, Northern Leaf Blight, Exserohilum turcicum, Pathogen Variability, 18S rDNA
Maize or corn is one of the staple foods for a global population of 4.5 billion people, comprising at least 30% of their diet.1 With climate change, there has been an increasing advent of disease outbreaks that have jeopardized maize production and, consequently, food security. Northern leaf blight (NLB) or Turcicum leaf blight (TLB) caused by the phytopathogenic fungus Exserohilum turcicum (Anamorph, formerly Helminthosporium turcicum) or Setosphaeria turcica (Teleomorph) has emerged as the major foliar disease of maize in recent times.2 The disease was first reported in 1878 from New Jersey, USA,3 and has been concluded to cause up to 50% loss in maize yield during severe incidence mainly by increased defoliation and decreased photosynthesis during the grain-filling period. The disease was first reported in Bihar by Butler in 1907 and simultaneously also found prevalent in Karnataka, Andhra Pradesh, Maharashtra, and Himachal Pradesh.4 The phytopathogen requires a moderately high temperature of 26°C, a high relative humidity of 80-90%, and cloudy weather with high rainfall.5 Hence, maize growing in tropical environments with hot and humid climatic conditions is more prone to this disease. India, being a sub-country, also suffers a loss varying from 25-90% in maize grain yield due to this disease.6
H. turcicum is a hemibiotrophic ascomycete fungus that spreads through conidia and causes infection in plants using the appressoria; thus, the disease incidence depends upon the successful conidial attachment and appressorium penetration into epidermal cells of maize leaves.7 Post-penetration, the fungi enters its biotrophic phase and suppresses or evades the defence response of maize plants while maintaining their viability.8,9 This is followed by the necrotic phase of fungi with higher production of secondary metabolites, cell wall degrading enzymes (CWDEs), and biomass that leads to necrosis.10,11 It also produces effector proteins that aid penetration and host colonization by modulating host defence responses.12,13 However, this effector-triggered reaction can be suppressed by elicitation of corresponding resistance gene (R-gene) response in maize by effector recognition.14 This is alternatively known as major gene resistance or qualitative resistance that is race-specific, which incites hypersensitive responses (HR) or localized cell death (LCD).15 This interrupts the supply of nutrients to the phytopathogen and results in their suppression of growth and development. To date, only a few genes have been identified to govern this resistance, namely Ht1, Ht2, Ht3, ht4, Htnl, Html, and NN.16 The maize breeders also utilize quantitative resistance, which utilizes a contribution of various genes with low additive effects for breeding schemes,17 as the major genes are vulnerable to resistance breakdown followed by the emergence of new races of the pathogen.18
A key strategy to combat E. turcicum in maize genotypes is to understand its mechanism of infection and the factors governing resistance. Since the phytopathogen is abundantly present in tropics and sub-tropics with high genetic variability,19 a thorough knowledge of the variations among and within the populations is imperative to develop appropriate breeding schemes.20 Various physiological races of E. turcicum have been determined based on their virulence to host genes, of which Race 0 is identified to be most prevalent among the maize-growing nations.21 However, despite significant efforts, the information related to variability in E. turcicum in Bihar is still absent. Hence, the present study was conducted to assess the cultural, morphological, and virulence variations among the isolates from seven major maize-producing regions of Bihar and identify the probable prevalence of races for accelerated resistance breeding programmes.
Sample collection and disease incidence
The diseased maize leaf samples showing characteristics symptoms of NLB were collected from seven major maize-producing districts of Bihar, India, namely Muzaffarpur, Bhagalpur, Saharsa, Madhepura, Purnea, Katihar, and Khagaria. Three blocks from each district were selected for sample collection, and disease incidence was recorded using a random observation method of 200 plants. Each sample was excised using sterilized scissors, collected in a sterilized polybag, and stored at 4°C for further processing. The disease incidence was calculated with the help of the following formulae:
Disease Incidence (%) = (Number of diseased plants / Total number of plants observed) x 100
Pathogen isolation, identification, and pure culture
The disease samples were surface sterilized with 1% sodium hypochlorite solution (NaOCl) for 1 minute, followed by rising thrice with sterilized distilled water. The lesions were then excised and incubated in a sterilized moist chamber for 3-5 days at 25°C to induce sporulation. The spores were then identified to the original description of E. turcicum using the microscope as per Geeta et al.22 The identified spores were then inoculated on potato dextrose agar (PDA) and incubated for 7 days at 25°C. The pure culture of each isolate was then obtained using the single-spore suspension method. The pure cultures were then incubated at room temperature for 14 days for further investigation. Further, they were also sub-cultured on PDA slants and preserved at 4°C.
Assessment of cultural and morphological variability
Each isolate was cultured on PDA at 25°C and observed for cultural characteristics at 2, 4, 6, and 8 days post-inoculation (DPI). The radial growth of the isolate was calculated using a scale from the centre to the margin in five angles for three replications. Sporulation of each isolate was calculated at 14DPI using the haemocytometer. The fungal colonies were also observed and recorded for shape, growing pattern, texture, and pigmentation at 14DPI. The conidial characteristics constituting shape, size, and septation were recorded using a compound microscope and micrometre.
Assessment of pathogenic variability and sporulation
The in vitro assessment of pathogenic variability was carried out by placing 5 mm discs of 14 days pure culture on surface-sterilized leaves of susceptible maize inbred line CM-202 followed by incubation at 25°C ± 2°C for 10 days. The disease development was observed daily, and days to the first symptom and lesion area were recorded for five replicates in a completely randomized design. The in vivo assessment of pathogenic variability was carried out by inoculating a 30-day plant of CM-202 with a conidial suspension of the isolate having 1 × 104 colony forming unit (CFU) ml-1 for each isolate. The maize plants were then observed after 14DPI for symptoms, lesion area, disease incidence, disease severity on a scale of 1-9,3 and the number of conidia cm-1 of lesion length. The disease incidence was calculated using the following formulae:
Disease Incidence (%) = [(Sum of numerical rating) / (Total number of plants observed ׳ Maximum rating)] × 100
The number of conidia cm-1 of lesion length was calculated by incubating a 1 cm diameter disc of leaf lesions for 4 days, followed by vortexing in a test tube with 5 ml of distilled sterilized water with Tween 20 and counting the conidia under a compound microscope using a haemocytometer.
Dendrogram development
The dendrogram of all the identified isolates of E. turcicum was developed to differentiate them into different clusters based on their cultural, morphological, and pathogenic variability on susceptible maize inbred line CM-202 using NTSYS software.
Molecular characterization and identification of the isolates
The genomic DNA of all the culturally identified isolates was harvested per Bankole et al.5 and their quality and quantity were checked using gel electrophoresis and spectrophotometer, respectively. The universal ITS1 (F:5’-TCCGTAGGTGAACCTGCGG-3’) and ITS4 (R:52 -TCCTCCGCTTATTGATATGC-32) were used for PCR amplification.23 The amplifications were checked for quality using gel electrophoresis and then sequenced using the Sanger sequencing method. The sequences were then aligned in the BLASTn program (www.ncbi.nlm.nih.gov/BLAST) to identify isolates, and the phylogenetic relationship analysis was carried out using MEGA X.24
Sample collection and disease incidence
A total of 21 blocks from 7 maize-producing districts of Bihar were surveyed, and the samples were collected along with the geographic location, weather conditions, disease incidence, and stage of the crop (Table 1). The disease incidence varied from 2 to 15%, with the highest observed in the Dholi (Moraul) block of the Muzaffarpur district.
Table (1):
List of sample collection sites, geographic location, weather conditions, disease incidence, and crop stage
| No. | Designated isolate | Location | Weather | DI (%) | Crop stage | ||||
|---|---|---|---|---|---|---|---|---|---|
| District | Block | GPS Coordinate | Temperature (℃) | RH (%) | |||||
| Min. | Max. | ||||||||
| 1 | Et1 | Muzaffarpur | Dholi (Moraul) | 26.025165/85.568179 | 14 | 28 | 84 | 15 | After Silking |
| 2 | Et2 | Muzaffarpur | Bandra | 26.067539/85.632066 | 15 | 30 | 76 | 13 | After Silking |
| 3 | Et3 | Muzaffarpur | Sakra | 25.965337/85.529875 | 14 | 26 | 75 | 7 | Grain Filling |
| 4 | Et4 | Bhagalpur | Sabour | 25.226653/87.043794 | 16 | 29 | 78 | 5 | Grain Filling |
| 5 | Et5 | Bhagalpur | Naugachhia | 25.367031/87.06027 | 13 | 28 | 77 | 10 | Before Silking |
| 6 | Et6 | Bhagalpur | Kahalgaon | 25.178475/87.117413 | 14 | 29 | 83 | 12 | Before Silking |
| 7 | Et7 | Saharsa | Sattarkataiya | 25.929987/86.560411 | 18 | 31 | 79 | 3 | Grain Filling |
| 8 | Et8 | Saharsa | Navhatta | 25.976439/86.529526 | 15 | 25 | 80 | 6 | After Silking |
| 9 | Et9 | Saharsa | Kahra | 25.823432/86.595469 | 19 | 30 | 78 | 4 | After Silking |
| 10 | Et10 | Madhepura | Ghelarh | 25.9798392/86.7270925 | 18 | 31 | 75 | 2 | Grain Filling |
| 11 | Et11 | Madhepura | Madhepura | 25.96701/6.749223 | 18 | 28 | 78 | 5 | Grain Filling |
| 12 | Et12 | Madhepura | Shankarpur | 26.009109/86.8186939 | 19 | 32 | 81 | 4 | After Silking |
| 13 | Et13 | Purnea | Kritiyanand Nagar | 25.848695/87.322638 | 16 | 29 | 76 | 3 | Silking |
| 14 | Et14 | Purnea | Banmankhi | 25.883885/87.178395 | 15 | 27 | 79 | 5 | Grain Filling |
| 15 | Et15 | Purnea | Dhamdaha | 25.755972/87.234309 | 16 | 30 | 82 | 8 | After Silking |
| 16 | Et16 | Katihar | Barsoi | 26.651351/87.921517 | 18 | 26 | 77 | 7 | Grain Filling |
| 17 | Et17 | Katihar | Azamnagar | 25.542218/87.847331 | 14 | 28 | 75 | 2 | After Silking |
| 18 | Et18 | Katihar | Pranpur | 25.47859/87.674578 | 15 | 27 | 79 | 3 | After Silking |
| 19 | Et19 | Khagaria | Chautham | 25.608672/86.590486 | 16 | 33 | 82 | 12 | After Silking |
| 20 | Et20 | Khagaria | Mansi | 25..506377/86.554478 | 14 | 29 | 78 | 9 | After Silking |
| 21 | Et21 | Khagaria | Khagaria | 25.505700/86.467200 | 18 | 31 | 76 | 6 | Grain Filling |
Cultural variability
A total of 21 pure cultures were obtained based on the description of E. turcicum. The isolates showed cottony growth with either a fluffy or flattened appearance. The margins were also either regular or irregular with white to grey to black colour (Figure 1 and Table 2). The radial growth varied significantly among the isolates and was categorized into four groups, namely rapid (60-70 mm) with five isolates (Et1, Et6, Et13, Et18, and Et19), fast (50-59mm) with nine isolates (Et2, Et3, Et4, Et5, Et8, Et9, Et16, Et20, and Et21), moderate (45-49 mm) with five isolates (Et7, Et10, Et11, Et12, and Et17), and poor (<45 mm) with two isolates (Et14 and Et16). The highest radial growth was observed in the Et6 isolate collected from Kahalgaon, with 67.8 mm, followed by the Et1 isolate collected from Dholi, whereas the lowest radial growth was observed in the Et16 isolate collected from Barsoi (Table 3). The isolates also showed a whitish-grey to greyish-black pigmentation at 14DPI. The highest sporulation of 9.80 × 105 spore ml-1 was observed in the Et1 isolate, followed by Et13 with 9.50 x 105 spore ml-1, whereas the lowest sporulation was observed in Et16 with 3.20 × 105 spore ml-1 at 14DPI (Table 3).
Table (2):
Cultural characteristics of the pure cultures isolated from 21 diseased plant samples on a PDA medium
Isolate |
Colony type |
Colony growth (8DPI) |
Margin |
Pigmentation (14DPI) |
Margin colour |
|---|---|---|---|---|---|
Et1 |
Slightly raised fluffy cottony |
++++ |
Irregular |
Greyish Black |
Black |
Et2 |
Slightly raised fluffy cottony |
+++ |
Regular |
Greyish Black |
Black |
Et3 |
Slightly raised fluffy |
+++ |
Regular |
Greyish Black |
Whitish |
Et4 |
Slightly raised fluffy |
+++ |
Irregular |
Greyish Black |
Whitish |
Et5 |
Flattened slightly raised |
+++ |
Irregular |
Greyish Black |
Grey |
Et6 |
Slightly raised cottony |
++++ |
Irregular |
Greyish Black |
Whitish Grey |
Et7 |
Fluffy raised |
++ |
Regular |
Whitish |
White |
Et8 |
Raised cottony |
+++ |
Irregular |
Greyish Black |
Grey |
Et9 |
Raised cottony |
+++ |
Regular |
Greyish Black |
Grey |
Et10 |
Raised cottony |
++ |
Regular |
Greyish Black |
Whitish Grey |
Et11 |
Raised cottony |
++ |
Regular |
Whitish |
White |
Et12 |
Raised cottony |
++ |
Regular |
Whitish Grey |
Grey |
Et13 |
Slightly raised white cottony |
++++ |
Regular |
Greyish Black |
Black |
Et14 |
Flattened cottony |
+ |
Regular |
Whitish Grey |
Whitish Grey |
Et15 |
Slightly raised cottony |
+++ |
Regular |
Whitish Grey |
Whitish Grey |
Et16 |
Flattened slightly raised |
+ |
Regular |
Whitish Grey |
Whitish Grey |
Et17 |
Fluffy raised cottony |
++ |
Regular |
Whitish Grey |
Whitish Grey |
Et18 |
Slightly raised white cottony |
++++ |
Irregular |
Grey |
Black |
Et19 |
Fluffy raised cottony |
++++ |
Regular |
Greyish Black |
Light Grey |
Et20 |
Raised cottony |
+++ |
Regular |
Whitish Grey |
Whitish Grey |
Et21 |
Slightly raised cottony |
+++ |
Irregular |
Greyish Black |
Black |
Colony growth: ++++ = Excellent (60-70mm), +++ = Good (50-59 mm), ++ = Moderate (45-49 mm), and + = Poor (<45 mm)
Table (3):
Radial growth and sporulation of the 21 pure cultures of E. turcicum on a PDA medium
| Isolate | Colony diameter (mm) | Sporulation (ml-1) at 14DPI | ||||
|---|---|---|---|---|---|---|
| 2DPI | 4DPI | 6DPI | 8DPI | Mean | ||
| Et1 | 36.5 | 64.0 | 76.5 | 90.0 | 66.8 | 9.80 × 105 |
| Et2 | 27.5 | 47.0 | 67.0 | 86.0 | 56.9 | 8.08 × 105 |
| Et3 | 28.0 | 46.0 | 66.5 | 84.0 | 56.1 | 8.12 × 105 |
| Et4 | 28.1 | 46.5 | 67.0 | 85.0 | 56.7 | 8.72 × 105 |
| Et5 | 28.5 | 49.5 | 68.5 | 87.0 | 58.4 | 8.85 × 105 |
| Et6 | 37.0 | 61.5 | 82.5 | 90.0 | 67.8 | 9.05 × 105 |
| Et7 | 21.0 | 42.5 | 61.0 | 73.0 | 49.4 | 5.20 × 105 |
| Et8 | 27.0 | 40.5 | 63.0 | 87.5 | 54.5 | 8.75 × 105 |
| Et9 | 25.5 | 45.0 | 68.0 | 81.0 | 54.9 | 8.40 × 105 |
| Et10 | 23.0 | 45.5 | 66.0 | 73.0 | 51.9 | 8.50 × 105 |
| Et11 | 21.2 | 41.5 | 58.5 | 69.0 | 47.5 | 5.45 × 105 |
| Et12 | 24.0 | 44.5 | 66.4 | 83.0 | 51.5 | 8.25 × 105 |
| Et13 | 34.0 | 62.0 | 83.5 | 90.0 | 67.4 | 9.50 × 105 |
| Et14 | 24.0 | 42.5 | 54.0 | 56.0 | 44.1 | 3.24 × 105 |
| Et15 | 27.5 | 47.0 | 66.5 | 81.0 | 55.5 | 8.30 × 105 |
| Et16 | 23.5 | 41.1 | 51.0 | 56.5 | 43.0 | 3.20 × 105 |
| Et17 | 22.0 | 43.0 | 61.5 | 73.0 | 49.9 | 5.50 × 105 |
| Et18 | 27.5 | 57.5 | 86.0 | 90.0 | 65.3 | 9.30 × 105 |
| Et19 | 31.0 | 59.0 | 85.5 | 90.0 | 66.4 | 9.08 × 105 |
| Et20 | 28.0 | 42.5 | 65.8 | 84.0 | 55.1 | 8.35 × 105 |
| Et21 | 27.5 | 42.3 | 64.8 | 82.0 | 54.2 | 8.87 × 105 |
DPI – Days post inoculation

Figure 1. Categorization of 21 E. turcicum isolates based on cultural and morphological variability of 21 E. turcicum isolates on PDA medium into Group-I, Group-II, and Group-III
Morphological variability
Varied morphological characteristics were observed among the E. turcicum isolates regarding spore colour, size, and septation. The colour of spores varied from brown to dark brown with 2 to 10 septations with a peculiar protruding hilum. The size of the spores also varied between 64.62 µm to 215.62 µm in length and 18.00 µm to 38.83 µm in width (Figure 1 and Table 4).
Table (4):
Morphological variability in conidia of the 21 pure cultures of E. turcicum on a PDA medium
| Isolate | Spore colour | Number of septa | Size of conidia | |
|---|---|---|---|---|
| Length (mm) | Width (mm) | |||
| Et1 | Dark Brownish | 3-10 | 215.62 | 36.05 |
| Et2 | Brownish | 3-5 | 123.69 | 31.04 |
| Et3 | Brownish | 3-8 | 146.67 | 32.00 |
| Et4 | Brownish | 3-6 | 113.22 | 24.41 |
| Et5 | Dark Brownish | 3-5 | 109.12 | 29.53 |
| Et6 | Dark Brownish | 3-9 | 195.48 | 35.44 |
| Et7 | Brownish | 2-6 | 64.62 | 29.73 |
| Et8 | Brownish | 3-9 | 169.16 | 33.52 |
| Et9 | Brownish | 3-8 | 142.62 | 34.17 |
| Et10 | Dark Brownish | 3-4 | 100.49 | 20.00 |
| Et11 | Dark Brownish | 3-7 | 74.96 | 28.28 |
| Et12 | Brownish | 3-7 | 126.82 | 31.24 |
| Et13 | Dark Brownish | 3-4 | 90.55 | 21.26 |
| Et14 | Brownish | 3-8 | 86.53 | 26.07 |
| Et15 | Brownish | 3-8 | 145.72 | 26.90 |
| Et16 | Brownish | 3-5 | 69.77 | 18.00 |
| Et17 | Brownish | 3-6 | 159.24 | 32.24 |
| E18 | Dark Brownish | 3-7 | 133.20 | 19.79 |
| Et19 | Dark Brownish | 3-10 | 207.20 | 38.83 |
| Et20 | Brownish | 3-5 | 84.02 | 22.00 |
| Et21 | Dark Brownish | 3-6 | 116.49 | 29.12 |
Pathogenic variability
The isolates produced significantly different disease reactions to the susceptible maize cultivar. In the in vitro study, five isolates were found to be highly virulent (Et1, Et6, Et13, Et18, and Et19) with 2DPI to the appearance of first symptom followed by eleven moderately virulent isolates (Et2, Et3, Et4, Et5, Et8, Et9, Et10, Et12, Et15, Et20, and Et21), and five least virulent isolates (Et7, Et11, Et17, Et14, and Et16) (Figure 2a). The lesion area was recorded to be about 250-400 mm2 at 10DPI for the highly virulent isolates, followed by 50-250 mm2 at 10DPI for moderately virulent isolates and 20-110 mm2 at 10DPI for the least virulent isolates (Figure 2b, 2c, and 2d). In the in vivo study, the initial symptoms from the isolates occurred as small elliptical to oval water-soaked greyish-green spots that enlarged along the leaf length with the progression of the disease. The lesions then became necrotic and spindle-shaped with straw-coloured centre and dark margins. The lesion appeared from the lower leaves to the upper leaves of the maize plant with a gradual increase in size (Figure 3). The highest lesion size was observed in Et6, Et13, Et18, and Et19 isolates with 35-40 mm × 9-13 mm. The lowest lesion size was observed in Et14 and Et16 isolates with 3 mm × 1 mm. The disease scoring scale also showed the highest ratings for Et1 and the lowest for Et14 and Et16. Similarly, the highest number of conidia cm-1 lesion length was observed in Et1, Et6, Et13, Et18, and Et19; the lowest was in Et14 and Et16 (Table 5).
Table (5):
Pathogenic variability in conidia of the 21 pure cultures of E. turcicum on a PDA medium
| Isolate | Lesion | Disease score (1-9 scale) cm-1 | No.of conidia lesion length | |
|---|---|---|---|---|
| Length (mm) | Width (mm) | |||
| Et1 | 40 | 13 | 8 | ++++ |
| Et2 | 14 | 4 | 4 | +++ |
| Et3 | 12 | 3 | 4 | +++ |
| Et4 | 10 | 3 | 3 | +++ |
| Et5 | 8 | 2 | 3 | +++ |
| Et6 | 35 | 9 | 7 | ++++ |
| Et7 | 12 | 4 | 5 | +++ |
| Et8 | 9 | 3 | 3 | +++ |
| ET9 | 13 | 4 | 4 | +++ |
| Et10 | 11 | 4 | 4 | +++ |
| Et11 | 8 | 2 | 2 | ++ |
| Et12 | 10 | 3 | 3 | +++ |
| Et13 | 38 | 12 | 7 | ++++ |
| Et14 | 3 | 1 | 1 | + |
| Et15 | 8 | 2 | 3 | +++ |
| Et16 | 3 | 1 | 1 | + |
| Et17 | 6 | 2 | 2 | ++ |
| Et18 | 36 | 10 | 7 | ++++ |
| Et19 | 37 | 12 | 7 | ++++ |
| Et20 | 10 | 3 | 3 | +++ |
| Et21 | 12 | 3 | 4 | +++ |
++++ = Excellent (50 – 60 × 103 conidia cm-1 lesion length), +++ = Good (38 – 49 × 103 conidia cm-1 lesion length), ++ = Fair (26 – 37 × 103 conidia cm-1 lesion length), and + = Poor (<26 × 103 conidia cm-1 lesion length).
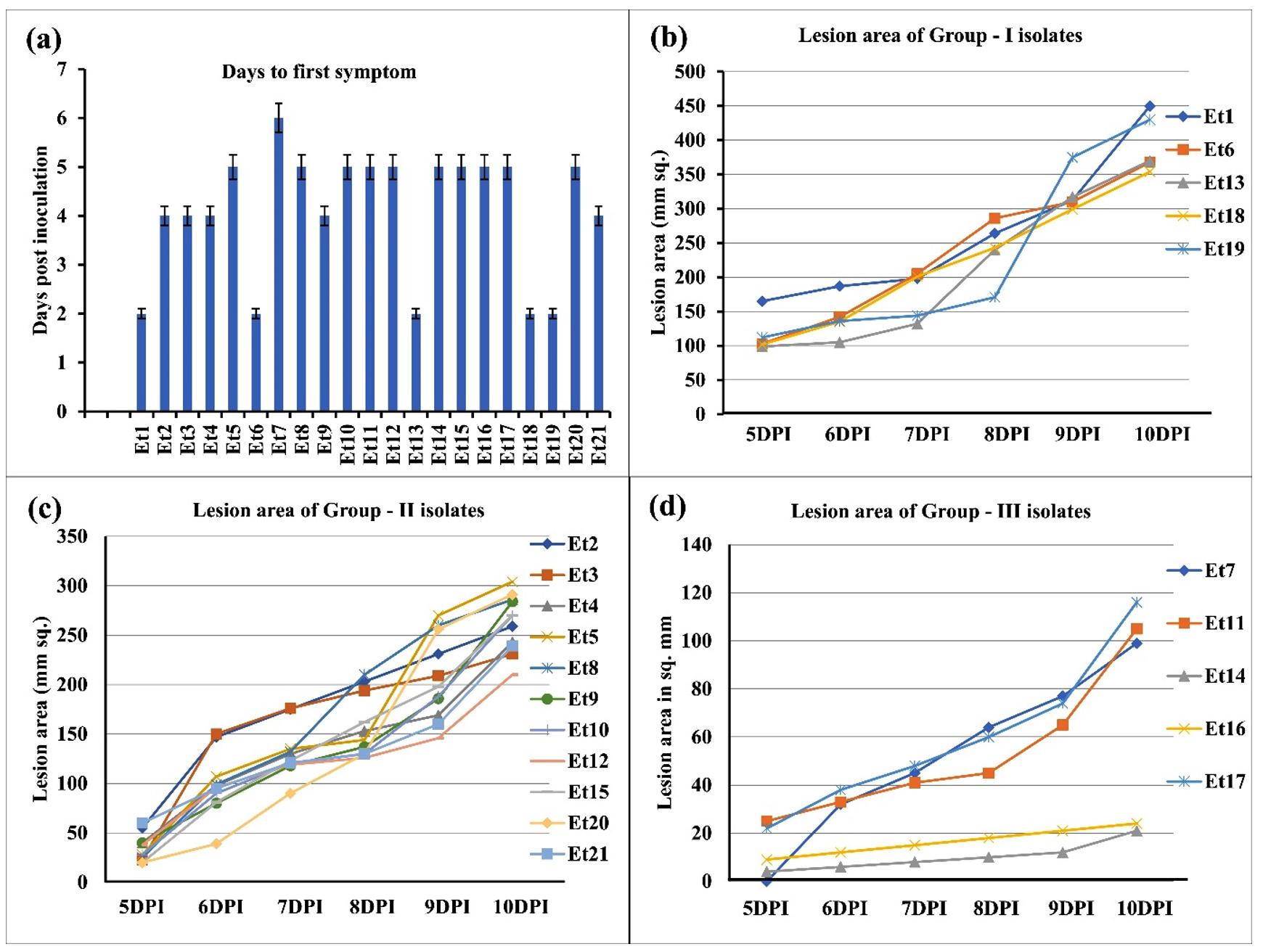
Figure 2. (a) Days to first symptom of disease under in vitro study for 21 E. turcicum isolates, (b) lesion area of group I isolates, (c) lesion area of group II isolates, and (d) lesion area of group III isolates

Figure 3. Symptom produced by E. turcicum isolates in in vivo study (a) initial symptom, (b) and (c) elliptical gray streaks on the surface of the leaves, (d) progression of symptoms, and (e) single, long and cigar shaped lesion
Dendrogram analyses
The dendrogram analyses based on cultural, morphological, and in vitro pathogenic variability of all the E. turcicum isolates on susceptible maize inbred line revealed 3 clusters with a similarity coefficient of 0.42 (Figure 4). Cluster I contained five isolates (Et1, Et6, Et13, Et18, and Et19) with a lesion length of 210-263 mm (mean of 10DPI) with somewhat similar cultural and morphological characters identified as highly virulent. Cluster II contained eleven isolates with a lesion length of 122-179 mm and had identical cultural and morphological characters identified as moderately virulent. Cluster III contained five isolates (Et7, Et11, Et14, Et16, and Et17) with lesion length of 10-60 mm identified as least virulent. Simultaneously, the dendrogram analyses based on in vivo pathogenic variability revealed 3 clusters of E. turcicum isolates with slight variation and a similarity coefficient of 0.52 (Figure 5). Cluster I contained six isolates (Et1, Et6, Et7, Et13, Et18, and Et19) with lesion length of 12-40 mm. Cluster II contained eleven isolates with lesion lengths of 8-14 mm, and Cluster III contained four isolates (Et11, Et14, Et16, and Et17) with 3-8 mm lesion lengths.
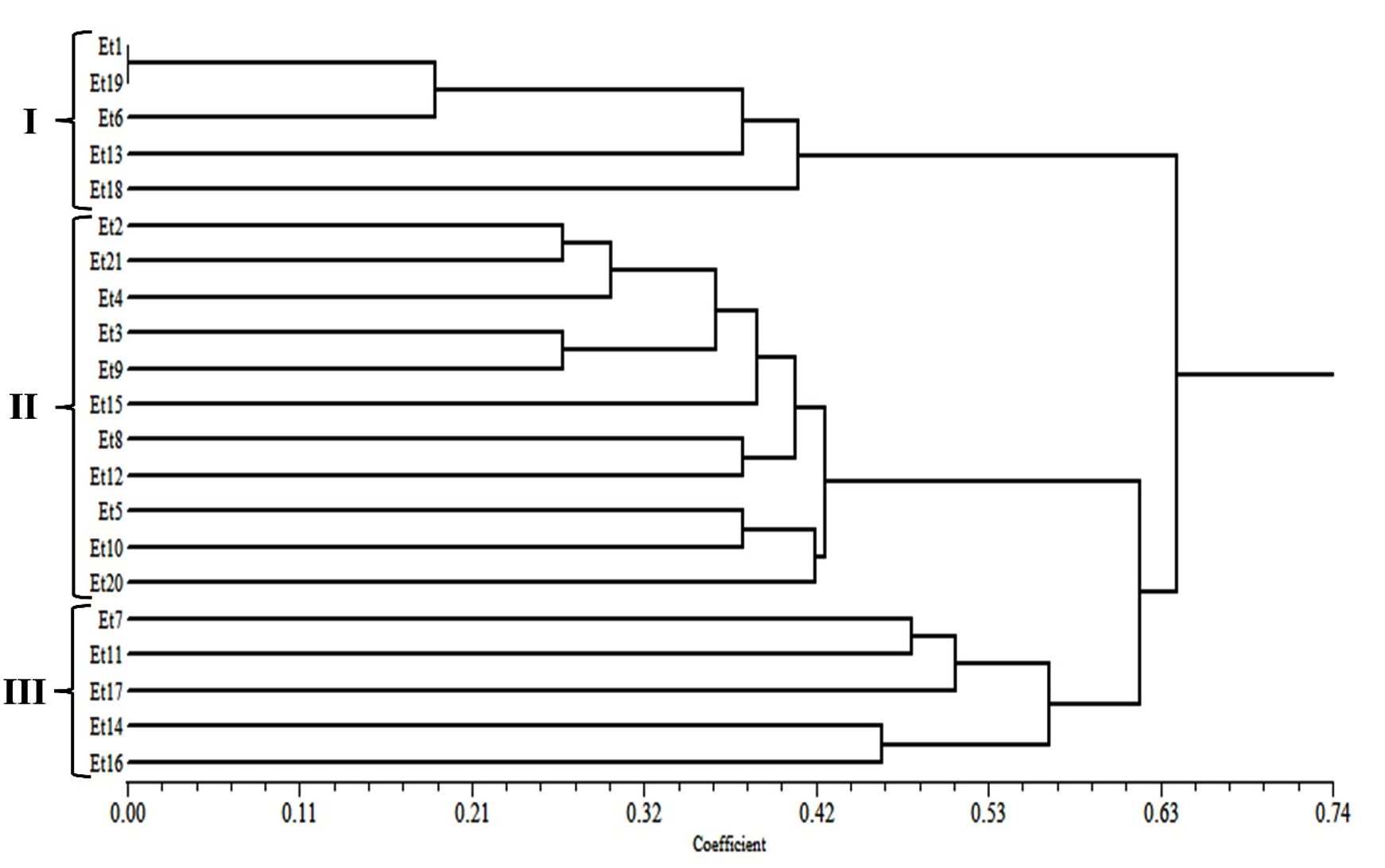
Figure 4. Dendrogram of 21 E. turcicum isolates based on cultural, morphological, and in vitro pathogenic variability
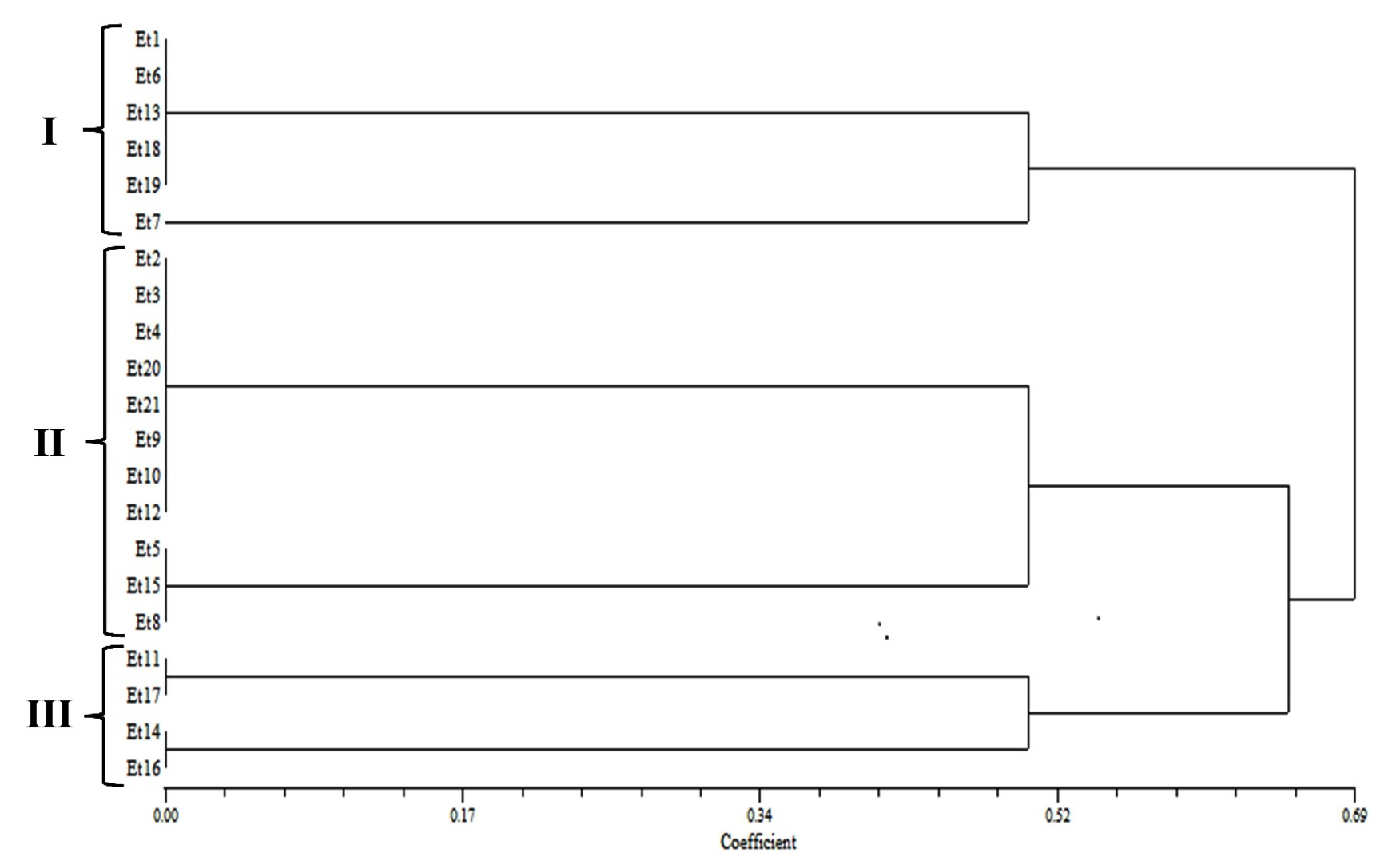
Figure 5. Dendrogram of 21 E. turcicum isolates based on in vivo pathogenic variability
Molecular characterization and phylogenetic relationship
All the isolates produced PCR amplification products to the universal primers (Figure 6a) and were identified as Setosphaeria turcica, the teleomorph stage of E. turcicum, from the BLASTn program. The phylogenetic analysis from the nucleotide sequence of isolates also revealed their relation with S. turcica strains and categorized them into three clades (Figure 6b). Clade I constituted Et7, Et16, Et11, and Et14, showing 100% identity to the reference isolate with Genbank accession number KJ956059.1. Clade II constituted Et1, Et17, Et6, Et13, Et19, and Et18 with 99% identity to the reference isolate with Genbank accession number KJ956053.1. Clade III constituted of Et2, Et3, Et4, Et5, Et9, Et10, Et12, Et15, Et20, and Et21 with about 46% identity to the reference isolate with Genbank accession numbers KJ956051.1, 956053.1, and 956056.1.
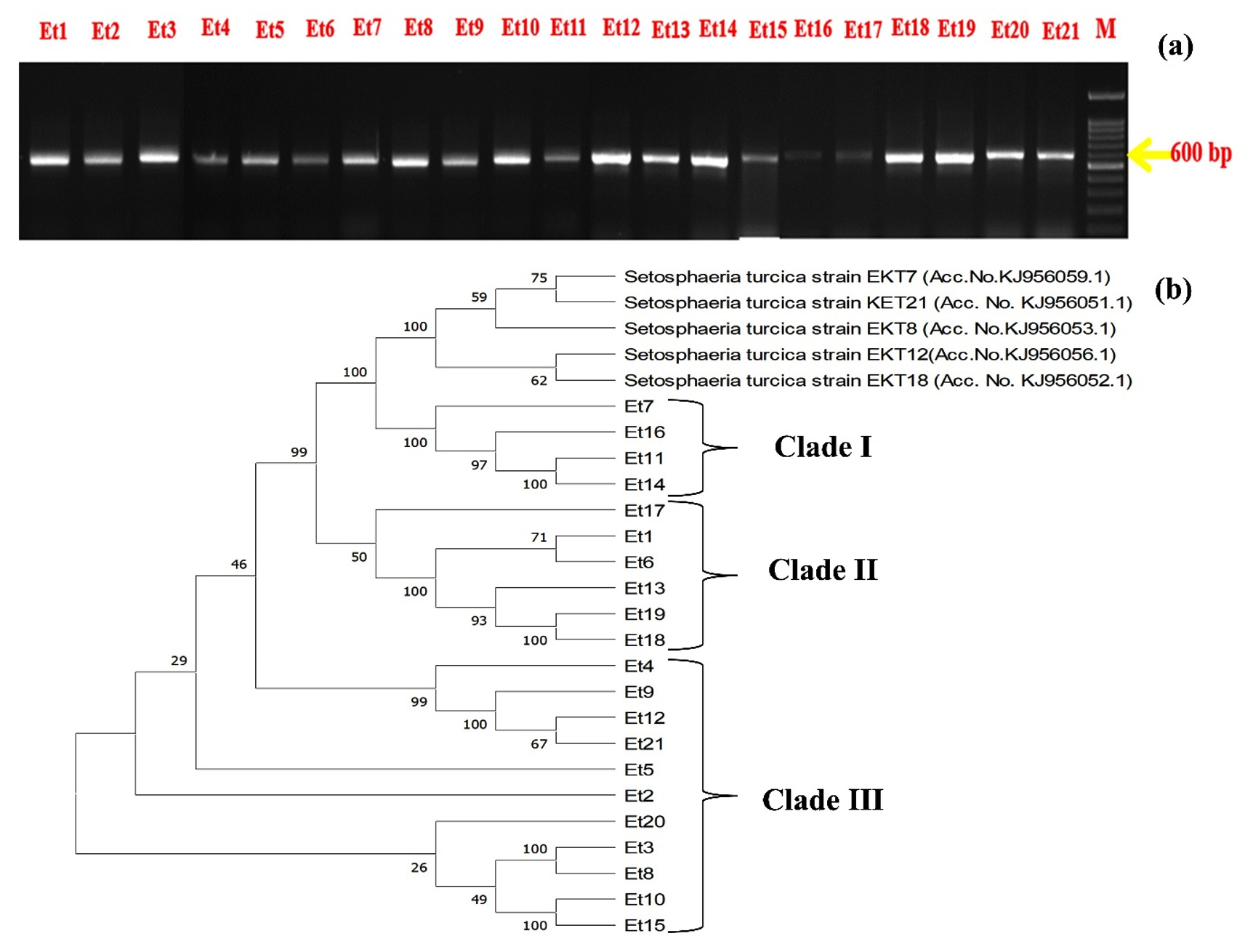
Figure 6. (a) PCR amplification product of ITS for isolates of E. turcicum on gel electrophoresis around 600 bp and (b) Phylogenetic relationship of 21 E. turcicum isolates
The present investigation is one of the first surveys to assess the variability in the population of E. turcicum associated with maize in Bihar, India. An evident variability in the population of the phytopathogen was present based on the cultural, morphological, and pathogenic characteristics. These variations were present both among the population and within the population of isolates collected from seven districts. A higher temperature and relative humidity in Dholi (Moraul), Bandra, Kahalgaon, and Chautham have resulted in higher disease incidence of 12-15% compared to lower temperatures and relative humidity at other locations.5,25 These climatic conditions favour the disease development process, contributing to higher disease incidence; thus, highly virulent isolates were obtained from these locations.
The isolates showed significant variations in cultural and morphological characteristics constituting radial growth, margin, pigmentation, sporulation, size and colour of conidia, and conidial septation. These variations could be due to the different environments in different areas and the phytopathogen’s survival strategy. Various studies have shown E. turcicum isolates to produce whitish to greyish-white colonies with white to greyish-black pigmentation on PDA medium.4,25,26 A similar range of shades was observed in the present investigation among the isolates. All the isolates produced conidia with a protruded hilum, a characteristic feature of the phytopathogen based on which it is named.27 The dimension and septation of the conidia among the isolated were also in conformation with previous research.26 The sporulation rate was higher in some of the isolates, which can be linked to the survival and spread of the phytopathogen.28 This can also be linked to higher virulence and disease incidence.25 As E. turcicum is a foliar phytopathogen, it spreads through rapid spore production and dissemination.6 Thus, a higher conidial production rate was observed in the isolates with higher virulence and vice-versa in the isolates with lower virulence. Additionally, this differential rate of radial growth and sporulation suggests that the phytopathogen is capable of rapid spread in the maize-growing regions under favourable conditions, which could present a stiff challenge in its management.
The assessment of pathogenic variability also showed a significant difference among the isolates. The disease caused stunted growth in infected maize plants and gradually progressed from lower to upper leaves. It reduced the photosynthetically active area of leaves and made them look burnt.6,19 The isolates with rapid radial growth and higher sporulation showed higher virulence and vice-versa in the in vivo study. This differential virulence is conditioned among and within the population due to the expression of effector genes5 in addition to the secondary metabolites, enzymes, and candidate genes that regulate pathogenicity in E. turcicum.11 As reported earlier in various studies, the phylogenetic analysis revealed genetic relatedness among the isolates from different districts. This is due to the high gene flow rate among the isolates of E. turcicum in addition to variations arising from sexual reproduction.29 Moreover, the phytopathogen also shows a high rate of sexual recombination in populations from topical regions,30 which adds to the genetic relatedness of isolates. This suggests that the phytopathogen can produce variants at a higher rate, so the breeding programmes should focus on quantitative resistance, avoiding the chances of resistance breakdown.
An evident variability among the E. turcicum population in Bihar, India, is responsible for causing NLB disease. All 21 isolates showed significant cultural, morphological, and cultural variability among and within the districts under investigation. The isolates were categorized into three clusters based on their aggressiveness: highly virulent, moderately virulent, and low virulent. Their virulence resulted from the growth rate and sporulation of the isolated coupled with environmental conditions. The variability among the population of phytopathogens would aid in developing management strategies and prospecting breeding programmes for NLB-resistant genotypes of maize to minimize yield loss. The study also concludes that the development of quantitative resistance would be more sustainable than qualitative resistance owing to the higher variability of the E. turcicum population.
ACKNOWLEDGMENTS
The authors are highly thankful to the Bihar Agricultural University, Sabour, India, for providing financial support, BAU communication No. 982/210602, and other necessities.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
MAA and RNS conceptualized the study. MAA, RNS, MMR and SA applied methodology. AK performed Wet-lab analyses. MI performed statistical analyses. MRA supervised the study. MAN wrote the manuscript. MMR reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Shiferaw B, Prasanna BM, Hellin J, Banziger M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011;3(3):307-327.
Crossref - Li P, Sohail A, Zeng F, Hao Z, Dong J. Setosphaeria turcica, the maize leaf blast pathogen: Current status and infection mechanisms. Agronomy. 2024;14(7):1488.
Crossref - Hooda KS, Khokhar MK, Shekhar M, et al. Turcicum leaf blight-Sustainable management of a re-emerging maize disease. J Plant Dis Protect. 2016;124(2):101-113.
Crossref - Sethy SK, Nongmaithem N, Sinha B, Singh KI, Singh NO, Priyadarshini M. Morphological and cultural variability among Exserohilum turcicum isolates causing Turcicum leaf blight disease of maize in Manipur. Int J Bio-Res Stress Manag. 2023;14:1052-1060.
Crossref - Bankole FA, Badu-Apraku B, Salami AO, Falade TDO, Bandyopadhyay R, Ortega-Beltran A. Variation in the morphology and effector profiles of Exserohilum turcicum isolates associated with the Northern Corn Leaf Blight of maize in Nigeria. BMC Plant Biol. 2023;23(1).
Crossref - Harlapur SI, Kulkarni MS, Yeshoda H, Kulkarni S. Variability in Exserohilum turcicum (Pass.) Leonard and Suggs, causal agent of Turcicum leaf blight of maize. Karnataka J. Agric. Sci. 2007;20:665-666.
- Zeng F, Meng Y, Hao Z, et al. Setosphaeria turcica ATR turns off appressorium mediated maize infection and triggers melanin involved self protection in response to genotoxic stress. Mol Plant Pathol. 2020;21(3):401-414.
Crossref - Ohm RA, Feau N, Henrissat B, et al. Diverse lifestyles and strategies of plant pathogenesis encoded in the genomes of eighteen Dothideomycetes fungi. PLoS Pathog. 2012;8(12):e1003037.
Crossref - Hurni S, Scheuermann D, Krattinger SG, et al. The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc Natl Acad Sci. 2015;112(28):8780-8785.
Crossref - Palma-Guerrero J, Ma X, Torriani SFF, et al. Comparative transcriptome analyses in Zymoseptoria tritici reveal significant differences in gene expression among strains during plant infection. Mol Plant Microbe Interact. 2017;30(3):231-244.
Crossref - Human MP, Berger DK, Crampton BG. Time-Course RNAseq reveals Exserohilum turcicum effectors and pathogenicity determinants. Front Microbiol. 2020;11.
Crossref - Lo Presti L, Lanver D, Schweizer G, et al. Fungal effectors and plant susceptibility. Annu Rev Plant Biol. 2015;66(1):513-545.
Crossref - Selin C, de Kievit TR, Belmonte MF, Fernando WGD. Elucidating the role of effectors in plant-fungal interactions: Progress and challenges. Front Microbiol. 2016;7:600.
Crossref - Flor HH. The complementary genic systems in flax and flax rust. Adv Genet. 1956;8:29-54.
Crossref - Galiano-Carneiro AL, Miedaner T. Genetics of resistance and pathogenicity in the maize/Setosphaeria turcica pathosystem and implications for breeding. Front Plant Sci. 2017;8:1490.
Crossref - Ndlovu N, Gowda M, Beyene Y, et al. A combination of joint linkage and genome-wide association study reveals putative candidate genes associated with resistance to northern corn leaf blight in tropical maize. Front Plant Sci. 2024;15.
Crossref - Yang Q, Balint-Kurti P, Xu M. Quantitative disease resistance: Dissection and adoption in maize. Mol Plant. 2017;10(3):402-413.
Crossref - Wende A, Shimelis H, Gwata ET. Genetic variability for resistance to leaf blight and diversity among selected maize inbred lines. In InTech eBooks. 2018.
Crossref - Muiru WM, Koopmann B, Tiedemann AV, Mutitu EW, Kimenju JW. Race typing and evaluation of aggressiveness of Exserohilum turcicum isolates of Kenya, German and Austrian origin. World J Agric Sci. 2010;6(3):277-284.
- Wiesner-Hanks T, Nelson R. Multiple disease resistance in plants. Annu Rev Phytopathol. 2016;54(1):229-252.
Crossref - Navarro BL, Hanekamp H, Koopmann B, von Tiedemann A. Diversity of expression types of Ht genes conferring resistance in maize to Exserohilum turcicum. Front Plant Sci. 2020;11:607850.
Crossref - Geeta, Aswathanarayana DS, Naik MK, Kenganal M, Kuchanur PH. Cultural, morphological and pathogenic variability of Exserohilum turcicum causing Turcicum leaf blight in maize. Int J Curr Microbiol Appl Sci. 2019;8(11):302-313.
Crossref - Kounosu A, Murase K, Yoshida A, Maruyama H, Kikuchi T. Improved 18S and 28S rDNA primer sets for NGS-based parasite detection. Sci Rep. 2019;9(1).
Crossref - Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35(6):1547-1549.
Crossref - Badu-Apraku B, Bankole FA, Ajayo BS, et al. Identification of early and extra-early maturing tropical maize inbred lines resistant to Exserohilum turcicum in sub-Saharan Africa. Crop Protect. 2020;139:105386.
Crossref - Anwer MA, Niwas R, Ranjan T, et al. Molecular and morphological characterization of Exserohilum turcicum (Passerini) Leonard and Suggs causing northern corn leaf blight of maize in Bihar. Bioengineering. 2022;9(8):403.
Crossref - Munkvold GP, White DG. Compendium of Corn Diseases. 4th Ed. American Phytopathological Society, St. Paul. 2016.
Crossref - Pusztahelyi T, Holb IJ, Pacsi I. Secondary metabolites in fungus-plant interactions. Front Plant Sci. 2015;6:573.
Crossref - McDonald BA, Linde C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu Rev Phytopathol. 2002;40(1):349-379.
Crossref - Borchardt DS, Welz HG, Geiger HH. Genetic structure of Setosphaeria turcica populations in tropical and temperate climates. Phytopathol. 1998;88(4):322-329.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.