


ISSN: 0973-7510
E-ISSN: 2581-690X
Trichoderma viride strains, which are filamentous fungi commonly found in soil, possess the ability to parasitize various fungi harmful to plants. In this study, ten strains of T. viride were isolated from different locations in Uttar Pradesh, India, and examined for their cultural, physiological, morphological, and antagonistic characteristics against soil-borne pathogens. The T. viride isolates were assessed at different pH levels for their growth, sporulation, and antagonistic efficacy under in vitro conditions using the dual culture technique against five major soil-borne pathogens that cause significant diseases in cereal and pulse crops. Among the ten isolates tested, the 49CP isolate from Sultanpur exhibited the maximum growth and sporulation at a pH of 6.5 as well as the highest inhibition percentages of mycelial growth in the pathogens: 63.23% against Fusarium oxysporum f. sp. ciceri, 65.85% against Sclerotium rolfsii, 53.33% against Sclerotinia sclerotiorum, 53.84% against Pythium sp., and 48.00% against Rhizoctonia bataticola. Trichoderma viride is also recognized for its effectiveness as a biocontrol agent against soil-borne pathogens, which are responsible for significant crop losses.
Antagonistic, pH, Trichoderma, Pathogens, Soil Borne
Trichoderma species are filamentous fungi that are commonly found in soil and root ecosystems, where they actively interact with root, soil, and leaf environments. These fungi produce secondary metabolites that can trigger localized or systemic plant defense responses. When Trichoderma strains colonize roots, they often promote root growth, enhance crop yield, increase resilience to environmental stress, and improve nutrient absorption and utilization. The highest protein content, measured 0.41% was observed in chickpea seedlings, bioprimed with a Trichoderma viride formulation (T. viride @ 5 g/kg seeds), and increased germination, root and shoot length of chickpea crops.1 Trichoderma species are also recognized for their effectiveness as antagonists against various soil-borne diseases, which have led to their exploration as bio-control agents for plant diseases. The motivation for this study stems from the benefits of using bio-control agents, which are considered safer and have a lower environmental impact than synthetic pesticides, as highlighted.2,3 Among the commonly used species, Trichoderma harzianum and Trichoderma viride have been developed into several commercial biological control products to inhibit the growth and development of various soil-borne pathogenic fungi. T. viride is marketed in Europe, while Trichoderma harzianum is marketed in India, both targeting different soil-borne pathogens in crop fields, greenhouses, and vegetable crops.4
Many Trichoderma species exhibit characteristics of opportunistic, nonpathogenic plant symbionts, forming mutualistic endophytic associations with numerous plant species. These species act as beneficial partners to plants, serving as natural bio control agents against various harmful phytopathogenic fungi. Trichoderma is one of the most effective mycoparasites, making it a promising biocontrol agent against a broad range of soil-borne phytopathogens. The efficacy of T. viride in inhibiting pathogens such as Rhizoctonia solani, S. rolfsii, and Fusarium spp. across different crops.5 The alternative methods such as protoplast fusion have been developed to enhance its antagonistic properties for bio control potential of Trichoderma species as noted.6 Its widespread use as a fungal biocontrol agent is largely due to its notable antagonistic activity against soil-borne plant pathogens.
The present study investigated the effects of various pH levels on the growth, sporulation, and antagonistic effectiveness of T. viride isolates, a fungus known for its ability to combat plant pathogens. The experiments were conducted at the Biocontrol Laboratory in the Department of Plant Pathology at C.S. Azad University of Agriculture and Technology, Kanpur, Uttar Pradesh, India.
Collection of soil samples and isolation and purification of T. viride
Fifty soil samples were collected from the rhizosphere of chickpea fields across various districts of Uttar Pradesh, each taken at a depth of approximately 5 to 6 cm. The samples were carefully placed in polythene bags, with each bag clearly labeled to indicate the location and date of collection. The samples were then transported to the laboratory for further analysis. Trichoderma species were isolated using the Trichoderma selective medium (TSM), which consisted of the following components: MgSO₄.7H₂O (0.2 g), K₂HPO₄ (0.9 g), KCl (0.15 g), NH₄NO₃ (1.0 g), glucose (3.0 g), chloramphenicol (0.20 g), Apron 35SD (0.01 g), Captan (0.2 g), Rose Bengal (0.15 g), agaragar (20 g), and distilled water (1 liter).7 The soil samples were processed using the serial dilution technique,8 followed by incubation of the plates at 28°C ± 1°C for 5 days. The appearance of colonies was noted and recorded between the third and fifth days. Individual colonies were selected and maintained in pure culture for further examination. T. viride isolates were purified using the single spore isolation method, with subculturing performed from the growing tip of each new colony. A small quantity of spores was aseptically taken and streaked onto potato dextrose agar (PDA) plates using a sterilized inoculating needle.
Morphological characterization of Trichoderma sp
The Trichoderma genus can be identified when grown on TSM by observing specific colony characteristics, such as growth pattern, growth rate, odor, and color.9
Microscopic features
Identifying Trichoderma species typically involves examining these characteristics under a light microscope, as described.10-12 Key morphological features used for identification include the branching pattern of conidiophores, the elongation and shape of conidiophore tips (whether coiled, straight, or undulating), the structure, size, and shape of phialides, and the morphology of conidia.
Coding of T. viride isolates
Chickpea rhizospheric soil samples were collected from various districts of Uttar Pradesh, including Etawah, Kaushambi, Faizabad, Sultanpur, Mirzapur, and Bhadohi. These samples were coded as 02CP, 10CP, 13CP, 21CP, 25CP, 45CP, 49CP, 64CP, 70CP, and 117CP, respectively.
Reconfirmation of cultures by ITCC, New Delhi
The Trichoderma pure culture was obtained using the single spore technique. Microscopic observation of mounted slides of the cultures was performed, and the cultures were sent to the ITCC in New Delhi, India, for accurate identification and to obtain accession numbers for the pure culture.
Collection of disease samples, isolation, and purification of pathogen
PDA medium was prepared for isolating the pathogen, following the method outlined.13 The medium consisted of 200 g of peeled potato, 20 g of dextrose, 20 g of agar, and 1000 mL of distilled water. Pathogen identification involved comparing the cultural and morphological characteristics of the fungus with those described14 for Fusarium oxysporum f. sp. ciceri. The color nomenclature was based on the standards provided.15 The growth habit, cultural traits, and morphological features of the fungus were examined on the PDA medium.
Effect of various pH levels on growth and sporulation of T. viride isolates
The potato dextrose broth (PDB) medium was prepared with different pH levels (4.5, 5.5, 6.5, and 7.5) by adding the appropriate amount of citrate phosphate buffer and adjusting the pH accordingly. Each 250 mL of conical flask was filled with 100 mL of PDB. The flasks were tightly sealed with nonabsorbent cotton plugs and covered with aluminum foil secured by rubber bands. The medium was sterilized by autoclaving at 15 psi (121.6°C) for 20 minutes. Under aseptic conditions in a laminar airflow, the PDB medium in the flasks was inoculated with 5 mm discs from 7-day-old cultures of T. viride isolates, with three replicates for each treatment. The inoculated flasks were then incubated in a biological oxygen demand (BOD) chamber at 28°C ± 2°C for 7 days. After incubation, the intensity of growth and sporulation of the different isolates of T. viride was assessed. The mycelial mats were harvested by filtering the culture through sterilized Whatman Filter Paper No. 4 and then dried in a hot air oven at 35°C for 48 h to obtain their dry weight. The biomass was calculated using the methods described.16,17
Antagonistic efficacy of T. viride isolates against soil borne pathogens by dual culture assay
The effectiveness of T. viride isolates was evaluated against Fusarium oxysporum f. sp. ciceri, Sclerotium rolfsii, Sclerotinia sclerotiorum, Rhizoctonia bataticola, and Pythium sp. using the dual culture technique. Both the target pathogens and the fungal antagonist (T. viride) were cultured on the PDA medium. Five-millimeter-diameter bits were cut from the edges of five-day-old cultures of both the pathogens and the antagonist, and these were placed diametrically opposite each other on the Petri dish, 5 mm from the edge. A control Petri dish was inoculated with each target pathogen alone without an antagonist. Each treatment was replicated three times and incubated at 28°C ± 2°C. After 7 days, the presence of inhibition zones between T. viride and the pathogens was observed. The radial growth of the pathogens on both the dual culture and control plates was measured, and the inhibition percentage of the pathogens was calculated using the methodology outlined.18
Percent inhibition (I) = C-T/C x 100
Where,
C represent the mycelial growth of pathogens in control.
T is the mycelial growth of pathogens in the dual culture plate.
Isolation and identification of T. viride isolates
Fifty soil samples were collected from the rhizosphere of chickpea fields across different districts of Uttar Pradesh. The bioagents were isolated and purified using the serial dilution technique, as described in the Materials and Methods section. These isolates were morphologically identified under microscopic observation, and out of the 50 isolates, 10 were confirmed as T. viride by the Indian Type Culture Collection (ITCC), Division of Plant Pathology, IARI (New Delhi). These 10 isolates were assigned accession numbers ITCC-9825 to ITCC-9834 (Table 1).
Table (1):
Trichoderma viride isolates with their accession numbers
| Isolates Code | Accession No. | Place of Collection | Fungus identified |
|---|---|---|---|
| 02CP | ITCC 9825 | Etawah | Trichoderma viride |
| 10CP | ITCC 9826 | Kaushambi | |
| 13CP | ITCC 9827 | Kaushambi | |
| 21CP | ITCC 9828 | Kaushambi | |
| 25CP | ITCC 9829 | Kaushambi | |
| 45CP | ITCC 9830 | Faizabad | |
| 49CP | ITCC 9831 | Sultanpur | |
| 64CP | ITCC 9832 | Mirzapur | |
| 70CP | ITCC 9833 | Mirzapur | |
| 117CP | ITCC 9834 | Bhadohi |
Effect of various pH levels on growth and sporulation of T. viride
All 10 confirmed T. viride isolates were tested at different pH levels to assess their impact on the growth and sporulation of the fungi. The pH levels of 4.5, 5.5, 6.5, and 7.5 were maintained across three replicate samples in PDB medium to determine the optimal conditions for the growth and biomass production of T. viride. The pH adjustments were made by adding an appropriate amount of citrate phosphate buffer. After a 7 day incubation period in the PDB medium with varying pH levels, the mycelial mats were harvested, and their average dry weights (measured in milligrams) were recorded and presented in Table 2. A review of the data (Table 2) revealed notable disparities in biomass production across all isolates. The highest average dry weight, 278.67 mg, was consistently recorded at a pH of 6.5, whereas the lowest, 227.67 mg, was observed at a pH of 4.5. In addition, isolate 49CP consistently exhibited the highest dry weight production at all pH levels, whereas isolate 117CP consistently demonstrated the lowest (Figure 1).
Table (2):
Effect of various pH levels on growth and sporulation of T. viride isolates
| T. viride isolates | Dry weight of mycelium (milligram) | |||
|---|---|---|---|---|
| pH 4.5 | pH 5.5 | pH 6.5 | pH 7.5 | |
| 02CP | 238.667 | 257.333 | 269.333 | 262.667 |
| 10CP | 242.000 | 262.667 | 271.667 | 263.667 |
| 13CP | 235.000 | 255.000 | 265.667 | 261.000 |
| 21CP | 240.333 | 258.000 | 266.000 | 262.000 |
| 25CP | 244.667 | 263.000 | 272.667 | 269.000 |
| 45CP | 250.000 | 266.000 | 275.333 | 273.000 |
| 49CP | 252.667 | 269.667 | 278.667 | 275.000 |
| 64CP | 232.667 | 243.000 | 263.667 | 256.667 |
| 70CP | 230.667 | 240.000 | 257.667 | 250.000 |
| 117CP | 227.667 | 239.000 | 251.667 | 245.000 |
| CD @ 5% | 7.100 | 13.516 | 7.709 | 5.505 |
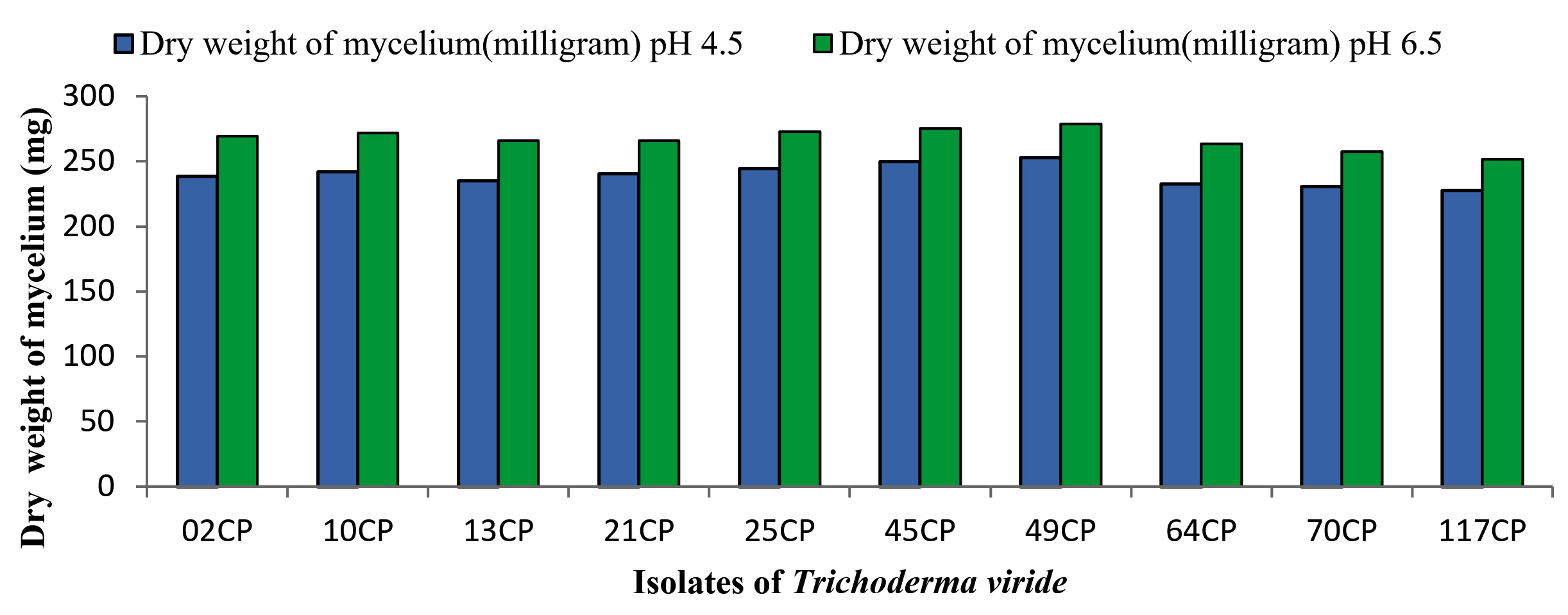
Figure 1. Effect of various pH levels on growth and sporulation of Trichoderma viride isolates
Antagonistic evaluation of T. viride isolates against soil borne pathogens
The antagonistic effectiveness of 10 T. viride isolates against five phytopathogens, isolated from chickpea rhizospheric soil, was assessed in vitro using the dual culture technique. The data summarized in Table 3 clearly indicate that all the isolates significantly impeded the radial growth of the tested pathogens. Among them, isolate 49CP exhibited the highest inhibition of mycelial growth, with percentages of 65.85%, 63.23%, 53.84%, 53.33%, and 48.00% against S. rolfsii, F. oxysporum f. sp. ciceri, Pythium sp., S. sclerotiorum, and R. bataticola, respectively. Conversely, the lowest inhibition rates were observed with isolates 64CP (51.21%), 70CP (48.50%), 45CP (40.00%), 13CP (39.25%), and 10CP (28.80%) against the same pathogens. The dual culture technique was applied to assess the antagonistic activity of T. viride against soil-borne pathogens under laboratory conditions, with the goal of identifying the most effective isolate among the ten tested. The results are shown in Figure 2 to Figure 6.
Table (3):
Antagonistic activity of T. viride isolates against soil borne pathogens
| S. No. | Foc | S. rolfsii | S. sclerotiorum | Pythium sp. | R. bataticola | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| T. viride isolates | Growth (mm) | *I.P. | Growth (mm) | I.P. | Growth (mm) | I.P. | Growth (mm) | I.P. | Growth (mm) | I.P. |
| 02CP | 28.0 | 58.82 | 38.0 | 53.65 | 33.0 | 51.11 | 36.0 | 44.61 | 39.5 | 36.80 |
| 10CP | 26.0 | 61.76 | 34.0 | 58.53 | 35.0 | 48.14 | 38.0 | 41.53 | 44.5 | 28.80 |
| 13CP | 32.0 | 52.94 | 36.0 | 56.09 | 41.0 | 39.25 | 32.0 | 50.76 | 39.5 | 36.80 |
| 21CP | 27.0 | 60.29 | 39.0 | 54.43 | 37.0 | 45.18 | 32.0 | 50.76 | 41.0 | 34.40 |
| 25CP | 30.0 | 55.88 | 37.0 | 54.87 | 34.5 | 48.88 | 33.0 | 49.92 | 40.5 | 35.20 |
| 45CP | 29.0 | 57.35 | 32.0 | 60.97 | 36.0 | 46.66 | 39.0 | 40.00 | 35.5 | 43.20 |
| 49CP | 25.0 | 63.23 | 28.0 | 65.85 | 31.5 | 53.33 | 30.0 | 53.84 | 32.5 | 48.00 |
| 64CP | 27.0 | 60.29 | 40.0 | 51.21 | 36.0 | 46.66 | 37.0 | 43.07 | 39.0 | 37.60 |
| 70CP | 35.0 | 48.52 | 34.0 | 58.53 | 32.5 | 51.58 | 38.0 | 41.53 | 37.0 | 40.80 |
| 117CP | 30.0 | 55.88 | 40.0 | 51.21 | 33.5 | 50.37 | 36.0 | 44.61 | 37.0 | 40.80 |
| Control | 68.0 | 00 | 82.0 | 00 | 67.5 | 00 | 65.0 | 00 | 62.5 | 00 |
| CD @ 5% | 2.861 | 4.013 | 4.274 | 4.268 | 6.062 | |||||
*I.P. = Inhibition Percentage
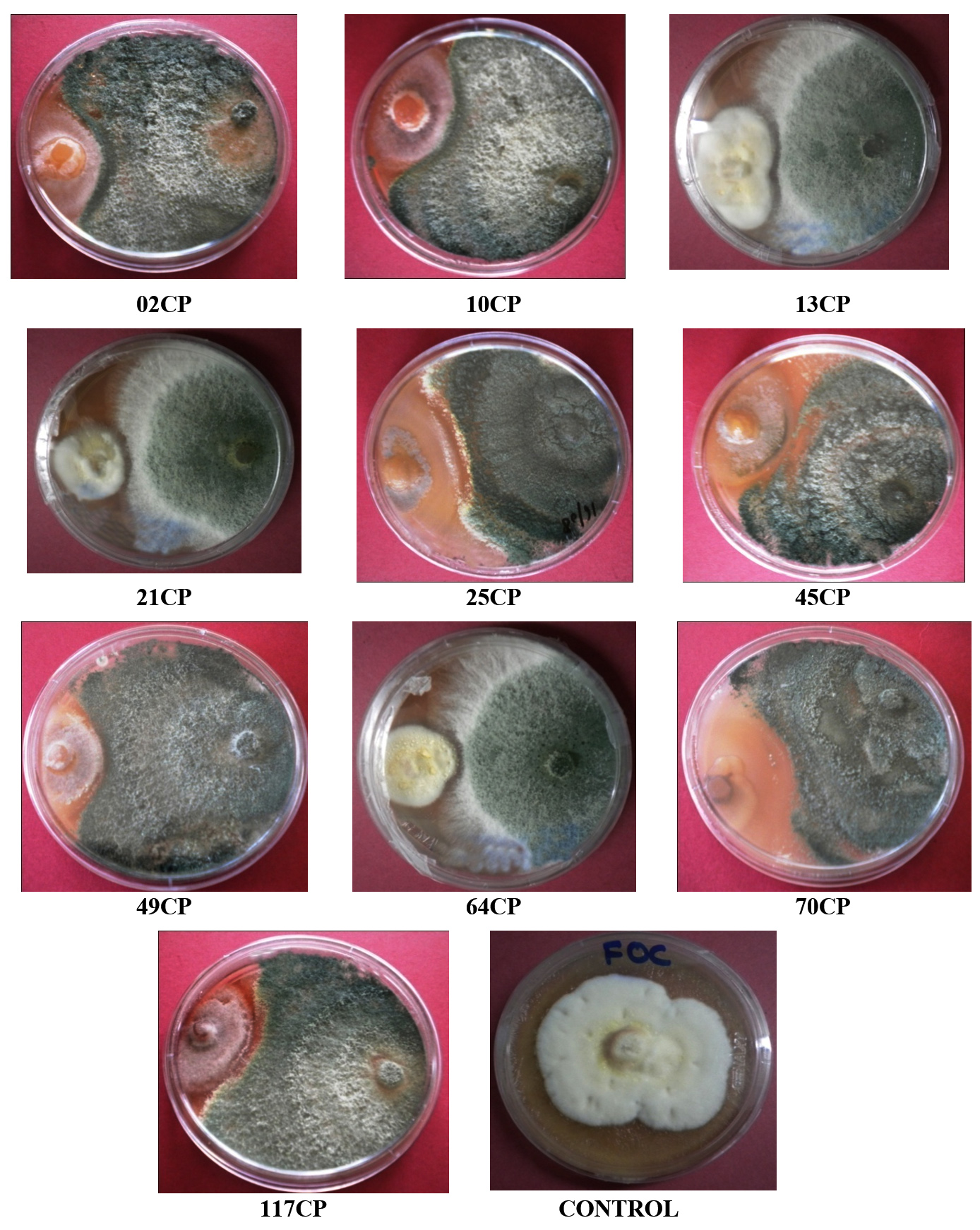
Figure 2. Antagonistic activity of T. viride isolates against Fusarium oxysporum f. sp. ciceri
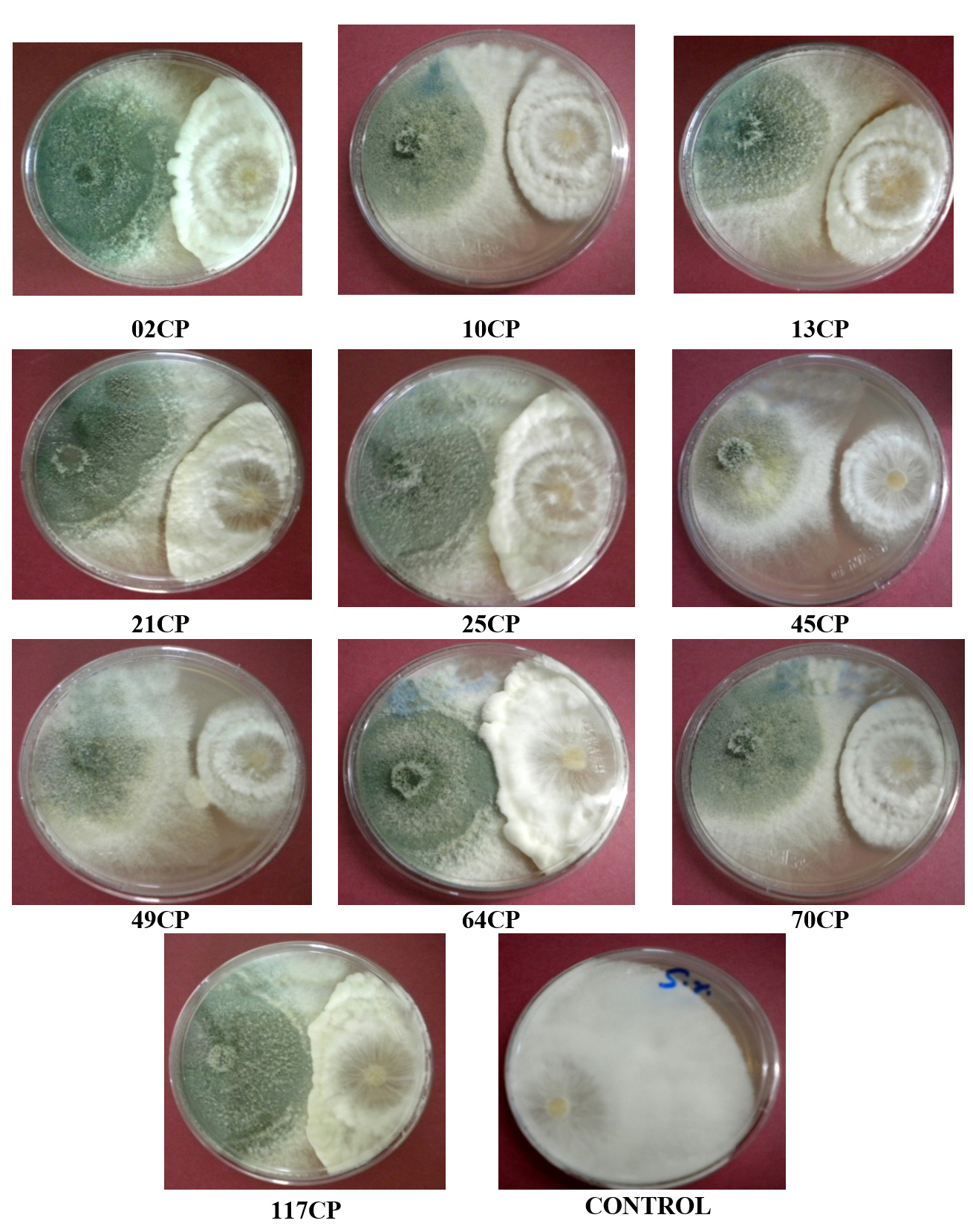
Figure 3. Antagonistic activity of T. viride isolates against Sclerotium rolfsii
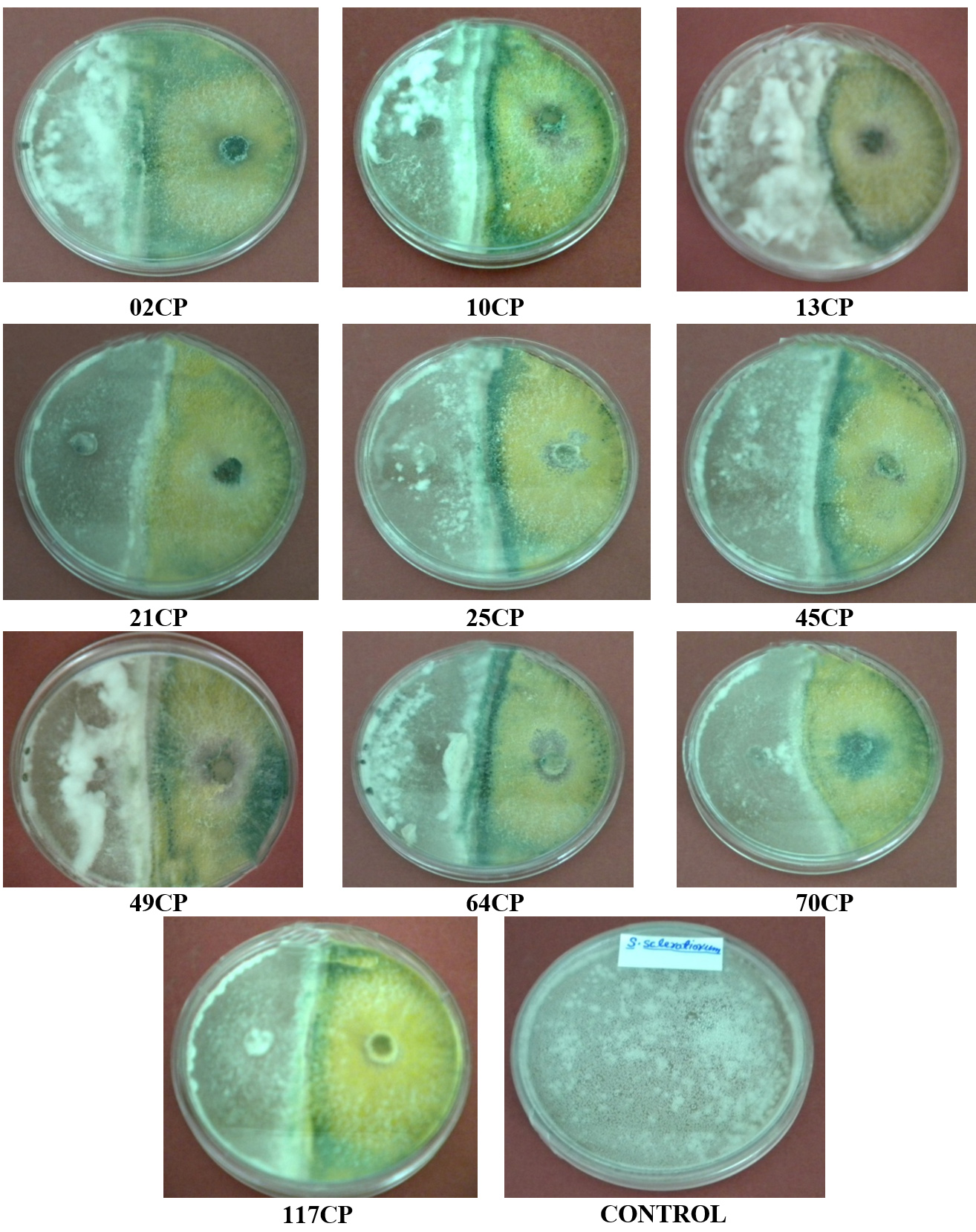
Figure 4. Antagonistic activity of T. viride isolates against Sclerotinia sclerotiorum
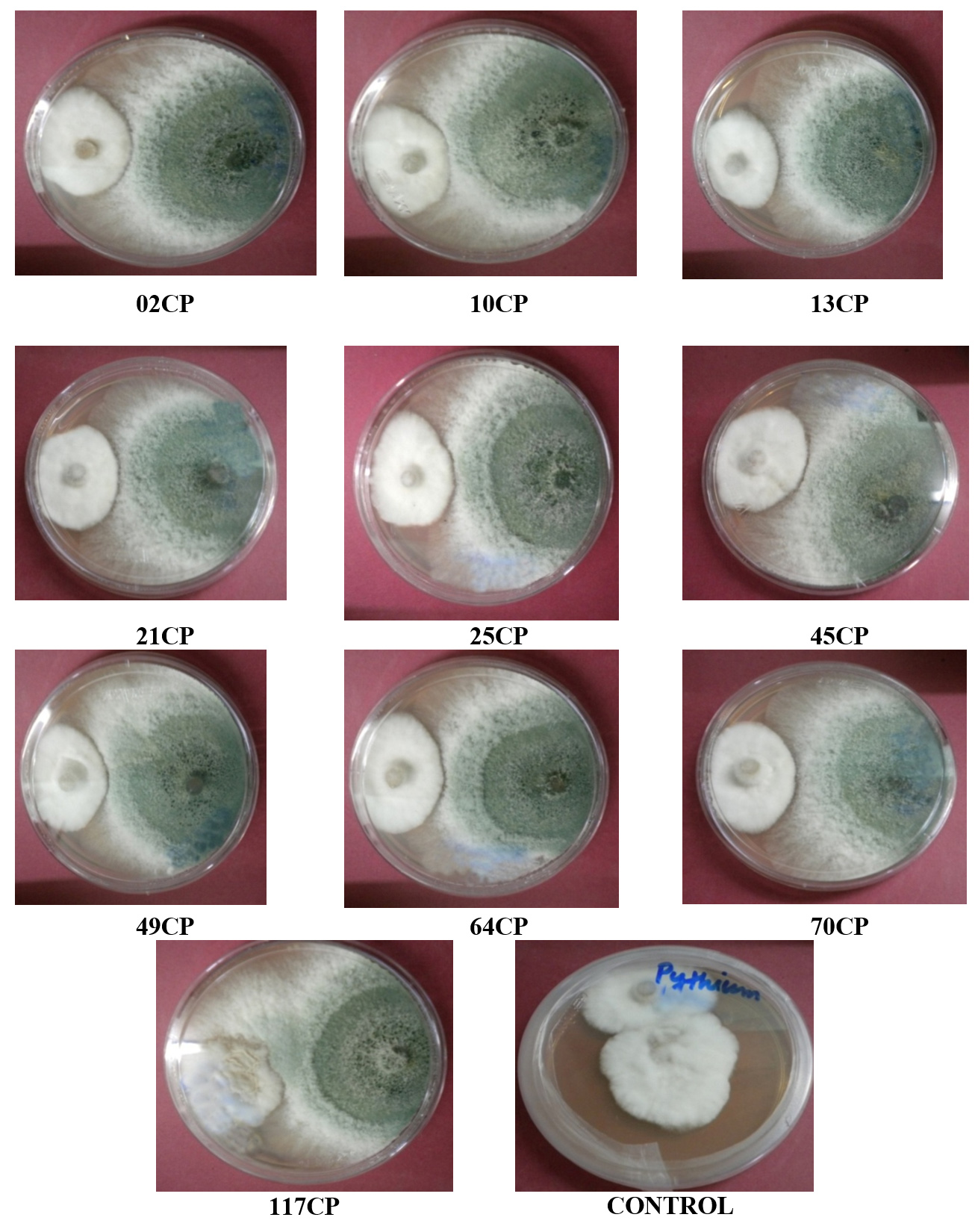
Figure 5. Antagonistic activity of T. viride isolates against Pythium spp.
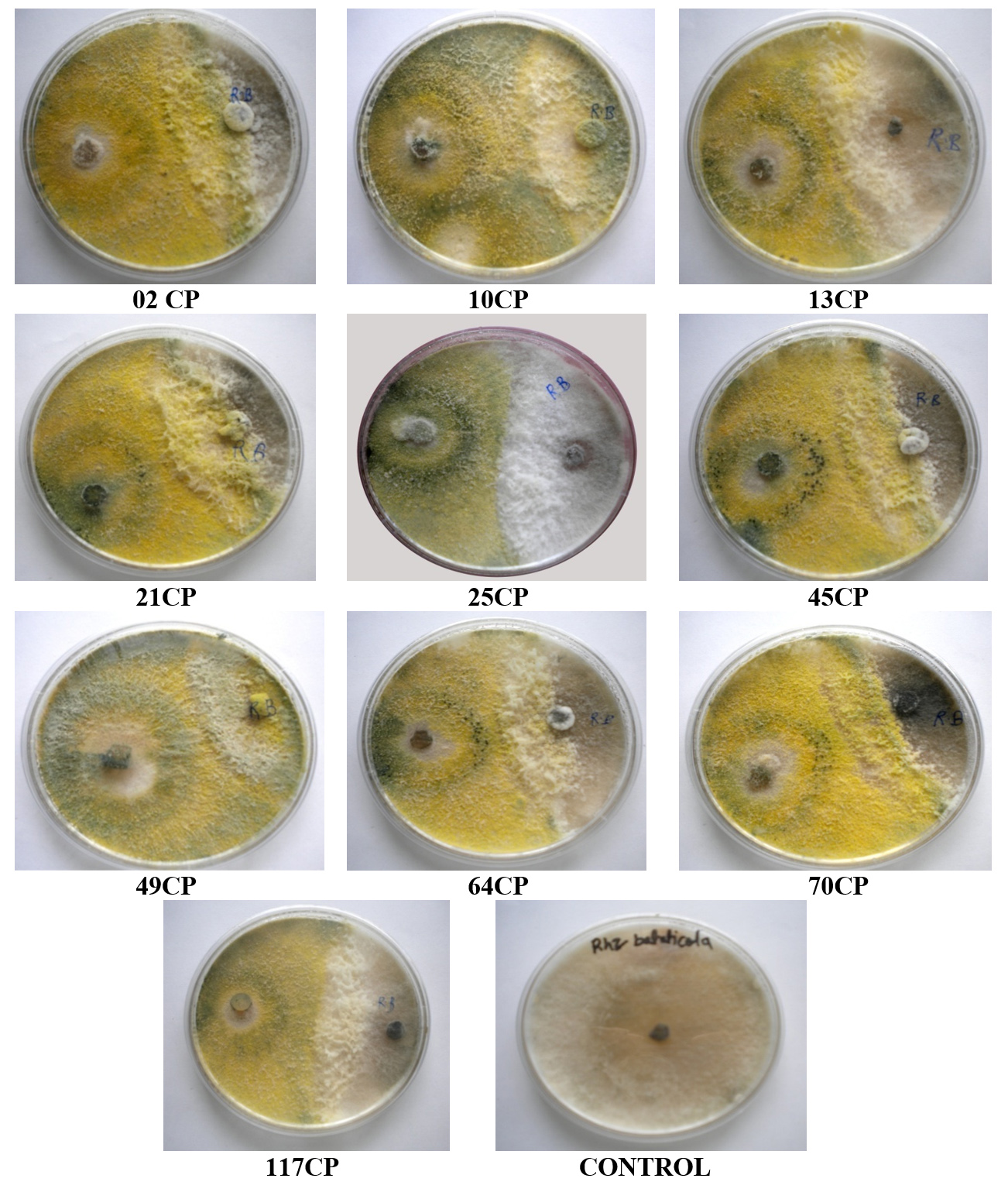
Figure 6. Antagonistic activity of T. viride isolates against Rhizoctonia bataticola
Fluctuations in pH significantly affected the growth and biomass production (sporulation) of T. viride isolates. Statistical analysis revealed that isolate 49CP exhibited superior growth compared with the other isolates, whereas the others performed similarly to each other. An optimal pH of 6.5 was identified for biomass production, yielding 278.67 mg of dry weight. This finding aligns with the results,16 who conducted experiments to determine the optimal pH conditions for biomass production of Trichoderma spp., exploring various pH levels and temperatures. They observed significant differences in biomass production across pH levels ranging from 4.0 to 8.0, with the most favorable pH range being between 5.5 and 7.5, where mycelium dry weight varied from 1.41 to 1.35 g. Similarly,17 investigated the physiological characteristics of Trichoderma sp. across different liquid media, temperatures, and pH levels. They found the pH range of 6.5 to 7.5 to be the most favorable, with mycelium dry weight ranging from 144.8 to 142.4 mg.19 Studied the effects of temperature (10°C, 15°C, 25°C, and 35°C) and pH levels (5.5, 7.0, and 8.5) on the growth and development of 26 native strains of Trichoderma spp. Most of these strains exhibited optimal growth and development at
25°C and pH 5.5.
The antagonistic evaluation of T. viride isolates against soil-borne pathogens in laboratory conditions aimed to identify the most efficient isolate. Similar findings were reported,20 who tested six strains of Trichoderma spp. to assess their ability to inhibit soil-borne pathogens, including Rhizoctonia solani, S. rolfsii, and S. sclerotiorum. In their study, co-culturing these pathogens with Trichoderma spp. revealed that T. viride (Tv-2) significantly reduced the mycelial growth of R. solani by 71.41% compared with the control. Similarly, T. viride (Tv-1) was the most effective isolate against S. rolfsii and S. sclerotiorum, exhibiting inhibition rates of 67.91% and 66.21%, respectively, over the control,21 evaluated the antagonistic capabilities of different Trichoderma species against F. oxysporum f. sp. ciceri, F. oxysporum f. sp. udum, and P. aphanidermatum. The highest inhibition of mycelial growth (65.00%) was observed against P. aphanidermatum, followed by 63.66% against Fusarium oxysporum f. sp. udum and 62.00% against Fusarium oxysporum f. sp. ciceri. However, T. virens (T.vi (CSAU) demonstrated the least effectiveness against the tested pathogens.22 explored the antagonistic effects of T. viride on mycelial proliferation of Fusarium oxysporum f. sp. ciceri, reporting a maximum inhibition of mycelial growth at 59.25% and the lowest average colony growth recorded at 11 mm by the 01PP isolate.23 Investigated eight Trichoderma species against prevalent phytopathogens, finding that Trichoderma viride (01PP) exhibited inhibition percentages of 50.00%, 70.42%, 78.88%, and 76.00% against S. rolfsii, R. solani, P. aphanidermatum, and F. oxysporum f. sp. ciceri, respectively. Similarly,24 evaluated five Trichoderma species-T. aggressivum, T. citrinoviride, T. erinaceum, T. harzianum, and T. koningiopsis isolated from various regions in India, for their antagonistic activity against Fusarium oxysporum f. sp. ciceri and Rhizoctonia bataticola, which cause wilt complex in chickpea. T. koningiopsis stood out with high tolerance, achieving the highest mycelial inhibition rates of 55.0% against F. oxysporum f. sp. ciceri and 36.3% against R. bataticola.25 Assessed the antagonistic efficacy of T. viride and T. harzianum strains against Fusarium proliferatum and Fusarium verticillioides using a dual culture assay. T. viride exhibited strong antagonistic activity, with inhibition rates of 80.17% against F. proliferatum and 70.46% against F. verticillioides. The silmiar study supported with Cyriac et al,26 investigated the antagonistic properties of Trichoderma isolates TRKR1, TRPN3, TRPN7, TRPN10, and TRPN18 against soil-borne pathogens Pythium aphanidermatum and Rhizoctonia solani, with TRPN7, TRPN15, and TRKR2 showing the highest mycelial inhibition (89.71%) against both pathogens. Similarly,27 evaluated nine native Trichoderma isolates for bio-efficacy against Sclerotium rolfsii, with isolate ARS K-21 achieving the highest inhibition (89.26%), followed by ARS K-11 at 83.70% in a dual culture assay.
A pH level of 6.5 (slightly acidic) was found to be the most conducive for the growth and biomass (sporulation) production of T. viride isolates. Among the ten T. viride isolates investigated in this study, the 49CP isolate demonstrated superior performance across all parameters, particularly in terms of antagonistic activity. Therefore, the 49CP isolate of T. viride is recommended for further analysis, including the preparation of bioformulations and both in vitro and in vivo trial experiments for managing soil-borne pathogens. T. viride-based bioformulations could serve as an effective alternative to chemical treatments for the management of plant diseases.
ACKNOWLEDGMENTS
The authors are highly grateful for the financial support granted by the Department of Science and Technology (DST) under the Inspire Fellowship. Authors are also thankful to Biocontrol Lab, Department of Plant Pathology, C.S.A. University of Agriculture & Technology Kanpur, Uttar Pradesh, for providing necessary facilities.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Maurya MK, Srivastva M, Pandey S, Shahid M, Ratan V. Protein profiling and biochemical analysis of chickpea seedling treated with T. viride (49CP). J Pure Appl Microbiol. 2016;10(4):3241-3247.
Crossref - Cotxarrera L, Trillas-Gay MI, Steinberg C. Alabouvette.Use of sewage sludge compost and Trichoderma asperellum isolates to suppress Fusarium wilt of tomato. Soil Biol Biochem. 2002;34(4):467-476.
Crossref - Brimner TA, GJ Boland. A review of the non-target effect of fungi used to biologically control plant diseases. Agriculture, Ecosystem and Environment. 2003;100:3-16
Crossref - Koch E. Evaluation of commercial products for microbial control of soil borne plant disease. Crop Prod. 1999;18(2):119-125.
Crossref - Gaigole AH, Wagh GN, Khadse AC. Antifungal activity of Trichoderma species against soil borne pathogen. Asia J Biotech Res. 2011;2:461-465.
- Chandrappa BP, Basavarajappa MP. Enhancement of antagonism through protoplast fusion in Trichoderma spp. J Pure Appl Microbiol. 2017;11(1):277-284.
Crossref - Elad Y, Chet I, Henis Y. A selective medium for improving qualitative isolation of Trichoderma sp. from soil. Phytoparasitica;1981;(9) 59-67.
- Johnson LF, Crul EA. Methods of research on ecology of soil borne pathogens. Minneapolis U.S. Burgess Pub. Soil Sci Soc Am J. 1972; 36(4):247.
- Gams W, Bissett J. Morphology and identification of Trichoderma. In: Harmann, G. E., Kubicek, C. P. (eds.) Trichoderma and Gliocladium. Taylor and Francis, 1998; 3-34
- Bisset J. A revision of the genus Trichoderma. II. Infrageneric classification. Canadian Journal of Botany, 1991; (69) 2357-2372.
- Rifai MA. A revision of the genus Trichoderma. Mycol. Paper. 1969;116:1-56.
- Sammuls G J. Trichoderma: a review of biology and systematic of the genus. Mycol. Res. 1996; (100): 923-935.
- Johnston A, Booth C. Plant Pathologists Pocketbook, Second ed., Commonwealth Mycological Institute, Kew. 1983:439.
- Booth C. The genus Fusarium. CMI, Kew, Surrey, England. 1971:231-237.
- Ridway R. Colour standard and colour nomenclature. Washington D.C., USA. Press of Boen and Co. Baltimore, M. D. 1912.
- Singh A, Shahid M, Srivastava M, Pandey S, Sharma A. Optimal Physical Parameters for Growth of Trichoderma Species at Varying pH, Temperature and Agitation. Virol Mycol. 2014;3(1):127.
- Srivastava M, Singh V, Shahid M, Singh A, Kumar V. Determination of biochemical and physiological aspects of a biocontrol agent Trichoderma harzianum Th azad. Int J Adv Res. 2014;2(3):841-849.
- Vincent JM, Budge SP. Screening for sclerotial mycoparasites of Sclerotinia sclerotiorum. Mycol Res. 1990;94(5):607-612.
Crossref - Andres PA, Alejandra PM, Benedicto MC, Nahuel RI, Clara BM. A comparative study of different strains of Trichoderma under different conditions of temperature and ph for the control of Rhizoctonia solani. Agric Sci. 2022;13(6):702-714.
Crossref - Amin F, Razdan VK, Mohiddin FA, Bhat KA, Banday S. Potential of Trichoderma species as biocontrol agents of soil borne fungal propagules. J Phytol. 2010;2(10):38-41.
- Srivastava M, Singh A, Shahid M. In vitro growth performance of Trichoderma species and antagonistic activity against soil borne pathogens. Int J Sci Res. 2012;3(7):672-675.
- Singh A, Shahid M, Srivastava M, Kumar V, Bansal A. Antagonistic activity of Trichoderma viride isolates against different Fusarium oxysporum isolated from legume crop of U.P. Society for Sci. Dev. in Agric. and Tech. (Progressive Research). 1912;8(1):47-50.
- Kumar V, Shahid M, Srivastava M, Singh A, Pandey S, Maurya MK. Screening of Trichoderma species for virulence efficacy on seven most predominant phytopathogens. Afr J Microbiol Res. 2015;9(11):793-799.
Crossref - Trivedi S, Srivastava M, Singh A, et al. Trichoderma koningiopsis a new and strong antagonist against soil borne pathogens of chickpea. J Pure Appl Microbiol. 2015;9(4):3265-3275.
- Yassin MT, Mostafa AAF, Al-Aksar AA, Sayed SRM, Rady AM. Antagonistic activity of T. harzianum and T. viride strains against some fusarial pathogens causing stalk rot disease of maize, in vitro. J King Saud Univer Sci. 2021;33(3):101363.
Crossref - Cyriac A, Sible GV, Johnson JM, Radhika NS, Krishnan AG. Antagonistic efficacy of Trichoderma isolates against soil-borne plant pathogens, Pythium aphanidermatum and Rhizoctonia solani. Journal of Biological Control. 2021;35(2):48-56.
Crossref - Meena N, Yadav DL, Gautam C, Yadav VK, Yadav SL, Meena CB. Trichoderma isolates against abiotic stresses and management of collar rot of lentil (lens culinaris L.) caused by sclerotium rolfsii. Indian J Microbiol. 2024;64(3):1366-1375.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.