


ISSN: 0973-7510
E-ISSN: 2581-690X
Bluetongue (BT) is an economically significant viral disease caused by bluetongue virus (BTV). BTV is spread by arthropods, which affects domestic and wild animals. In fetal ruminants, BTV exhibits neurotropism, leading to abortion and congenital defects in brain especially in cattle and sheep when infected in utero. Transplacental transmission (TPT) of wild Indian BTV-1 in late pregnancy has never been proven experimentally. This study is the first to demonstrate TPT of wild Indian BTV-1 during late pregnancy using an immunocompetent mouse model of IFNAR1 blockade. The present study examines sequential pathology, developmental anomalies, demonstration of BTV-1 antigen localization in the reproductive organs, haematological and biochemical alterations, and humoral immunity during the late stage of gestation. During late stage of gestation, the rate of TPT was notably higher at 80.00%. Clinical signs became increasingly noticeable in dams from 5 dpi onward, including symptoms such as abortion, anorexia, ocular discharge, and huddling tendency. Reduced growth rate, rough hair coat, and head tilting due to nervous signs were observed in 10-day-old fetuses. BTV-1 infection during the late stage of gestation caused abortion, maceration, necro-haemorrhagic lesions in reproductive organs, hemorrhages in foetal organs, and non-significantly decreased bone size (Alizarin red staining) in fetuses. Perivascular hemorrhages due to endothelial damage in lungs, interstitial pneumonia and meningitis were observed in 10-day-old fetuses. BTV-infected pregnant mice showed leukopenia with lymphopenia and seroconversion by 6 dpi (21 days of gestation). Further, significant alterations in serum biochemical values were observed when compared them to those of the uninfected control group. BTV-1 antigen was demonstrated in foetuses, placenta, uterus, and ovaries using immunohistochemistry and RT-PCR. This mouse model presents a suitable platform for studying the mechanisms underlying transplacental transmission.
Transplacental Transmission, Late-stage of Gestation, Bluetongue Virus, Mouse, Abortion, Type 1 IFN Receptors
Bluetongue (BT) is a non-contagious viral disease transmitted by arthropods, primarily biting midges of Culicoides spp., affecting domestic and wild ruminants. It is caused by the bluetongue virus (BTV), which belongs to the Orbivirus genus within the Reoviridae family.1 Being the most susceptible species, the prominent clinical manifestation of BT is typically observed in sheep; while, cattle and goats often show subclinical or mild signs. World Organization for Animal Health (WOAH) recognized BT as a notifiable disease and it leads to disruption of global economy due to the significant economic losses and barriers in trade of animals. The disease is manifested as a haemorrhagic syndrome, marked by fever, difficulty in breathing, oedema, haemorrhages in viscera, erosions of mucosa, ulcers, and coronitis.1 In pregnant animals, BTV infection can result in abortion, congenital abnormalities, brain malformations like hydranencephaly and porencephaly, leading to the birth of calves with viraemia.
Transplacental transmission (TPT) of BTV is causing huge economic losses in sheep farming especially low income group due to stillbirth, abortion, congenital foetal deformities, decreased birth weight in offspring, lower milk production and reduced fertility rates.2,3 TPT resulted in overwintering of BTV in the in utero infected foetuses during the period of absence of Culicoides activity.4 Lambs or calves born with viremia can remain infected for a long period of time and serve as potential carriers of introducing the disease into regions, which are free from BT during seasons without vectors.5 Transplacental transmission of BT in ruminants was caused by live attenuated vaccine strains (including BTV-1, -2, -9, -10, -11, -13, and -17), which was prior to the BTV-8 outbreak in Europe in 2006.2,3,5
Repeated passaging of BTV in suited cell lines resulted in the emergence of quasispecies with modified phenotypic characters including alterations in pathogenicity, tissue tropism, and the capacity to pass through the placental barrier.1,4 Various experimental studies and field epidemiological investigations documented the TPT of wild-type strains of BTV-8 in northern Europe in ewes, goats, and cows. The studies highlighted that field strains of BTV-1, -2 and -8 can readily cross the placenta around 70-75 days of pregnancy and causing encephalopathy in
ewes.6-11
The TPT in cattle had been observed with live attenuated vaccine viruses of four strains namely, BTV-10, -11, -13, and -17, which was not reported with wild-type strains until 2006.5 Further, incidence of TPT of wild BTV-8 was reported in Western Europe, which spread to northern Europe, causing severe BT infection in cattle and sheep.2,3 The rate of TPT varies depending on the species and stage of gestation, estimated at around 10% to 41.7% in cows12 and up to 69% in ewes.13 The transmission of BTV into disease-free areas as a result of the birth of calves, which were positive for the virus and the persistence of virus in heifers are the most significant effects of TPT. The isolation of BTV serotype-1 from aborted and non-viable goat fetuses in Sardarkrushinagar, Gujarat, India, in 2007 marked the first incidence of TPT of wild BTV.14 The isolation of BTV-1 from transplacentally infected foetuses indicated the initial confirmation of TPT of wild-type BTV, as no laboratory attenuated BTV strains had been used in the region and more than 50% of infected dams showed severe clinical manifestations, such as abortion or stillbirth.
Saminathan et al.15 conducted the first investigation into the TPT of wild-type Indian BTV-1 in IFNAR1-blocked pregnant mice at different stages of pregnancy and reported a higher transmission rate during the mid-stage (71.43%) than early-stage (57.14%) of gestation. Despite BTV infections in India dating back to 1964, the potential birth abnormalities in ruminants linked to TPT and the distribution of viral antigens in reproductive organs remain poorly understood. Using an IFNAR1-blocked immunocompetent mouse model, this study was the first to attempt to demonstrate the capability of transplacental transmission in wild-type Indian BTV at the late-stage of gestation and the pathogenesis of the disease in foetus and dam.
Animal experiments
Animals and animal shed
Required number of female Swiss albino mice (6-8 weeks of age), were obtained from Laboratory Animal Resources (LAR) Section, ICAR-Indian Veterinary Research Institute (ICAR-IVRI), Izatnagar. The animals were accommodated in Experimental Animal Shed with insect proof facility, CADRAD, ICAR-IVRI, Izatnagar. The studies were conducted as per the recommendations of the Institute Animal Ethics Committee (IAEC).
BTV serotype-1 infection in pregnant mice
Stage of estrous cycle determination
Young adult female mice between 6-8 weeks of age were used for breeding. Stage of the estrous cycle was identified based on the criteria reported by Byers et al.16 Proestrus stage in mice was identified by swollen, moist and pinkish vagina. The opening was wide and wrinkles or striations were observed along the ventral and dorsal edges. Estrus stage was identified by less pink, moist and swollen vaginal opening, and the striations are more pronounced (Figure 1).
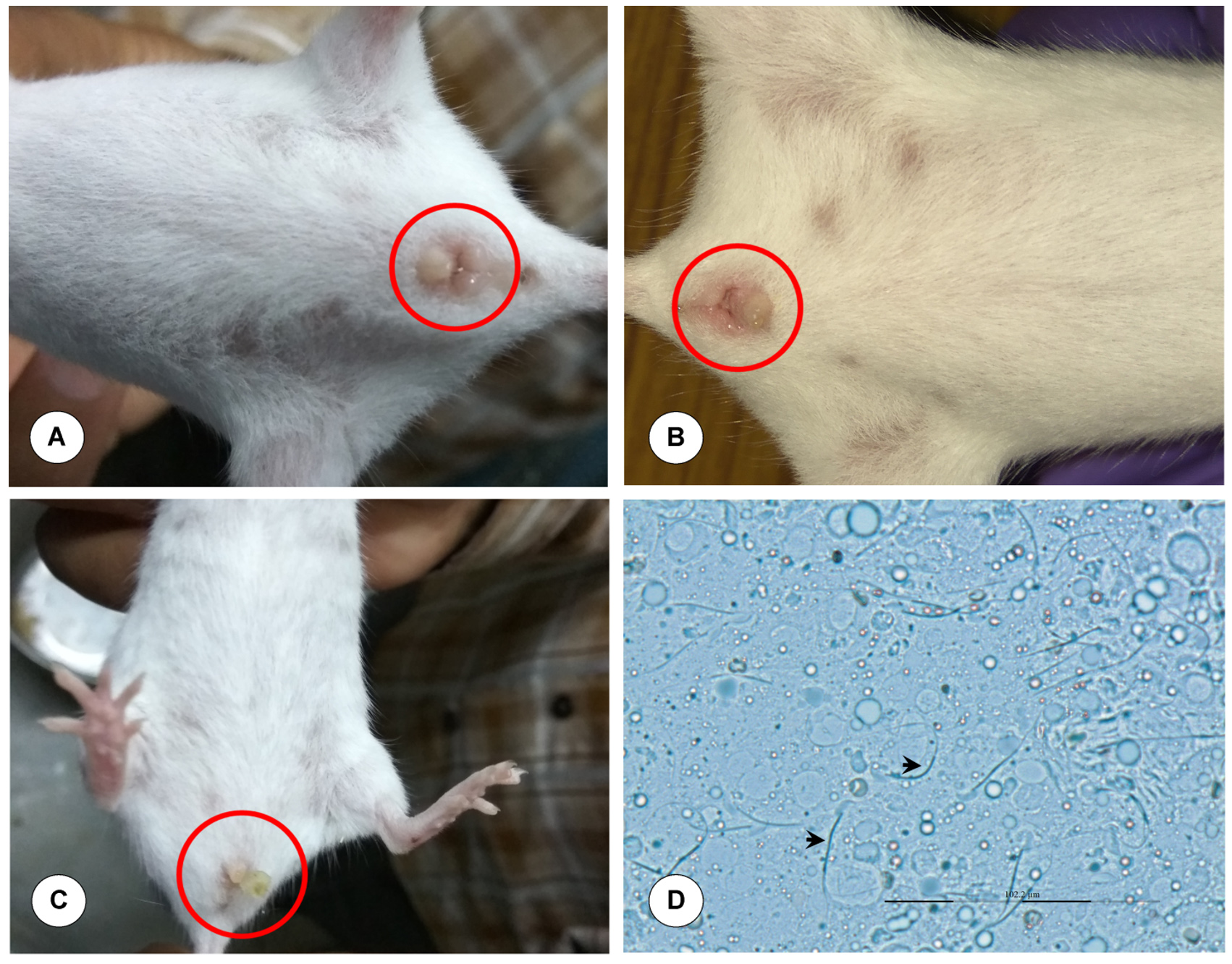
Figure 1. Breeding of mice. (A) Proestrus: Swollen and moist vagina. (B) Estrus: Moist and wide vaginal opening. (C) Copulatory plug: Coagulated secretions of seminal vesicles of male. (D) Confirmation of copulatory plug: Presence of hooked (curved head) spermatozoa. Magnification x400
Breeding of mice
The virgin mice in estrus and proestrus stages were grouped and permitted for mating for maximum of 15 nights in a manner that each cage were with fertile one adult male and two female mice of the same strain (2 females:1 male ratio). During the mating period, the copulatory plug in the vagina was identified and considered as first gestation day (GD).
Experimental design and blocking of type 1 IFN receptors
After confirmation of pregnancy, pregnant mice with late stage of gestation were randomly divided into two groups. Wild-type pregnant mice were administered with 1.5 mg/mouse of IFN-a/b monoclonal antibody intraperitoneally (I/P) one day prior to BTV infection and 0.5 mg/mouse one day after BTV infection to block IFNAR1 signaling.17 On 15 day of gestation, pregnant mice were infected with 50 µL volume containing 1 x 106 TCID50/mL of BTV-1 intravenously (I/V) and acted as BTV-infected group. On 15 gestation day, late stage pregnant mice were injected with MAR1-5A3 antibody I/P and uninfected tissue culture fluid I/V as described above and this group was designated as uninfected control group.
Clinical observations
All pregnant mice were daily monitored for clinical signs like anorexia, dullness, nasal and ocular discharges, respiratory distress, stillbirth or abortion and nervous signs. Three mice each from BTV-infected and uninfected groups were sacrificed at 1 (16 GD), 3 (18 GD), 5 (20 GD), and 6 (21 GD) dpi.
Sample collection
During sacrifice, around 1.5 ml of blood was collected in EDTA vacutainers from the heart of both BTV-infected and uninfected animals for haematological studies and RNA isolation from blood for detection of viremia by RT-PCR. Blood was also collected in clot activator tubes for separation of serum to detect BTV antibodies and estimation of biochemical parameters. Multiple sections of lymph nodes, spleen, brain, lungs, placenta, ovary, uterus, fetal organs, and entire fetuses were collected in 10% NBF and in RNAlater® (Ambion, USA).
Postmortem lesions
Systematic postmortem examination was carried out. In the pregnant animals, the uterus was examined thoroughly to determine the total number of fetuses, both live and dead fetuses, number of macerated and/or mummified fetuses.15 Gross pathological findings in ovary, fetuses, uterus, lymph nodes, spleen, lungs, brain, etc. were recorded. The sequelae of BTV-1 infection on late stage of pregnancy were compared with uninfected pregnant mice.
Effect of BTV-1 in postnatal life
Fetuses born from late stage of gestation were observed carefully for clinical signs like general health status, respiratory and neurological signs, and mortality until 10 days of postnatal life. At the end of an observation period, young ones from infected and uninfected animals were sacrificed and tissues of brain, spleen and lungs were collected in 10% NBF and duplicate were frozen at -80°C for virus distribution studies.
Transplacental transmission study of BTV-1
The pregnant dams were sacrificed at specific kinetics intervals after BTV inoculation to study the kinetics of vertical transmission of BTV in late stage of pregnancy. The placentas and fetuses were collected from infected dams to detect BTV-1 viremia using RT-PCR.
Study of developmental anomalies
The BTV-1 inoculated pregnant does were sacrificed at 21 day of gestation and fetuses were removed from uterus, viable fetuses were anesthetised and sacrificed. The foetuses were examined for the evidence of foetal abnormalities, and other macroscopic lesions. The deformities in the skeleton of fetuses were examined using Alizarin red staining. Briefly, the protocol includes removal of all the organs in the thoracic and abdominal cavities, and skinning the fetuses. The fetuses were fixed in 10% NBF for 24 hours. The fetuses were washed with distilled water (DW) and again fixed in 70% ethanol until future processing. The muscles of fetuses were digested with 1% potassium hydroxide (KOH) until the skeletal parts are readily visible. Fetal bones were stained with 0.2% alizarin red until the bones became deep purple in color and cartilage became unstained. Stained fetuses were washed with DW to remove the excess stain. The fetuses were cleared in clearing solution for 2 hours and stored in 100% glycerine.
Measurement of fetuses and different bones
The length of fetuses as a whole and the length of various bones in fore and hind-limbs, skull, and vertebral column length were measured using stereo zoom microscope.
Estimation of haematological parameters
At specified time intervals, blood samples from dam were examined for hematological parameters such as red blood cells (RBC), total leukocyte count (TLC), differential leucocyte count (DLC), packed cell volume (PCV), total platelet count (TPC), hemoglobin (Hb), mean corpuscular hemoglobin (MCH), mean corpuscular volume (MCV), and mean corpuscular hemoglobin concentration (MCHC).
Estimation of biochemical parameters
At specific intervals, serum samples from dam were analysed for biochemical parameters namely, serum glutamate oxalate transaminase (SGOT)/aspartate amino transferase (AST), serum glutamate pyruvate transaminase (SGPT)/alanine amino transferase (ALT), alkaline phosphatase (ALP), and creatine kinase (CK) were estimated by measuring optical density in serum samples (Coral Clinical Systems, India).
Assessment of humoral immunity
Detection of BTV antibodies by c-ELISA
The serum samples of infected pregnant mice collected at specified time intervals were processed for estimation of antibodies against BTV using Bluetongue Virus Antibody Test Kit, cELISA (Cat No. 287-5; VMRD Inc., USA) as per the manufacturer’s protocol.
Histopathology
After proper fixation of tissues in 10% NBF and tissues were processed by embedding in paraffin wax. Tissue sections of 4-5 µ thickness were prepared by microtomy and staining with haematoxylin and eosin (H&E) as per the standard procedure for microscopic examination of tissues.18
Immunohistochemistry for demonstration of BTV-1 antigen
Duplicate sections of the tissues were fixed on the APES (3-Aminopropyl triethoxysilane) coated glass slides to carry out IHC. BTV-1 antigen localization was confirmed in formalin fixed tissues of lymph nodes, trachea, spleen, lungs, brain, uterus, placenta, foetal tissues, and ovary of infected pregnant mice by following the protocol described by Saminathan et al.15 Briefly, the protocol includes slides were coated with 2% APES and 4-5 µ thickness tissue sections were taken on APES coated slides. Sections were rehydrated through serial changes in graded alcohols and rinsed with DW. Heat induced epitope retrieval was performed using 10 mM sodium citrate buffer (pH 6.0), in a microwave oven. Blocking of endogenous peroxidase activity was done using 3% H2O2 in 80% methanol for 30 min. Blocking of non-specific antigen was done with 5% normal goat serum. Tissue sections were incubated with hyper-immune serum raised against BTV-1 core antigen in rabbit for overnight in a humidified chamber at 4°C. Slides were incubated with peroxidase-conjugated goat anti-rabbit secondary antibody for 1 hour. Slides were incubated with ImmPACT™ DAB peroxidase substrate for 30 seconds and counter stained with Mayer’s haematoxylin for 2 min. Sections were covered with CC mount and examined under the microscope for positive signals.
BTV nucleic acid detection in different organs by RT-PCR
Detection of BTV nucleic acid in different organs at specified time intervals by RT-PCR was carried out in Eppendorf Thermocycler system, USA by using 2x DreamTaq green PCR master mix as per the manufacturer’s instruction.
Statistical analysis
The data were analyzed using GraphPad Prism and expressed as mean ± SEM. Unpaired
t-test was used to compare the effects of BTV-1 in late stage of gestation with uninfected control and P < 0.05 was considered as significant.
Clinical signs
The clinical signs in BTV-1 infected group were documented for 5/6 dpi (20/21 GD). The BTV-1 infected mice exhibited depression and dullness on 2 dpi. On 5 dpi onwards, clinical signs became more prominent in dams like weight loss, anorexia, conjunctivitis, ocular and nasal discharges, and huddling tendency. Some animals showed signs of abortion.
Gross lesions
Congestion or haematoma was noticed below the base of the ear in the fetus on 6 dpi/ 21 GD (Figure 2A). Maceration of the foetus was observed (Figure 2B) and congested, hemorrhagic and dead fetus was also noticed (Figure 2C). Reduced growth rate, rough hair coat, and head tilting due to nervous signs were observed in the 10-day-old fetuses born from BTV-1 infected dam during late stage of pregnancy (Figure 2D). Markedly enlarged and oedematous axillary (Figure 3A) and inguinal (Figure 3B) lymph nodes were observed in the dam. In the dam, both lung lobes were enlarged, oedematous, congested and rubbery consistency was observed on 6 dpi/21 GD (Figure 3C). Spleen was enlarged and diffusely oedematous on 6 dpi (Figure 3D).
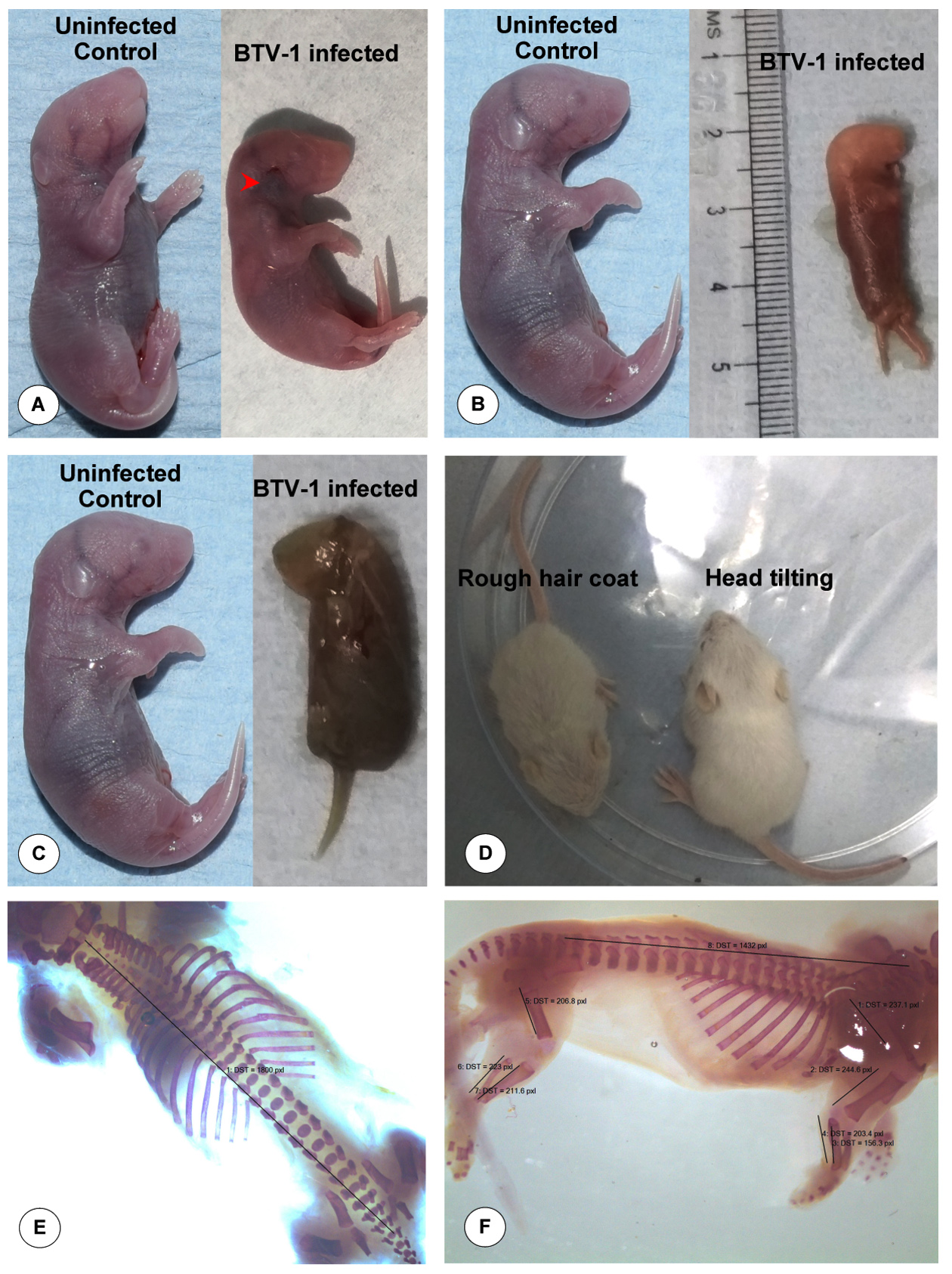
Figure 2. Gross pathological lesions in BTV-1 infected fetuses during late stage of gestation. (A) Congestion or haematoma below the base of the ear at 6 dpi (21 GD). (B) Congested dead fetus at 5 dpi (20 GD). (C) Maceration of dead fetus at 6 dpi (21 GD). (D) Reduced growth rate, rough hair coat, and head tilting due to nervous signs in fetuses at 10 days post-natal life. (E & F) Examination of skeletal deformities in BTV-1 infected fetus by Alizarin red staining
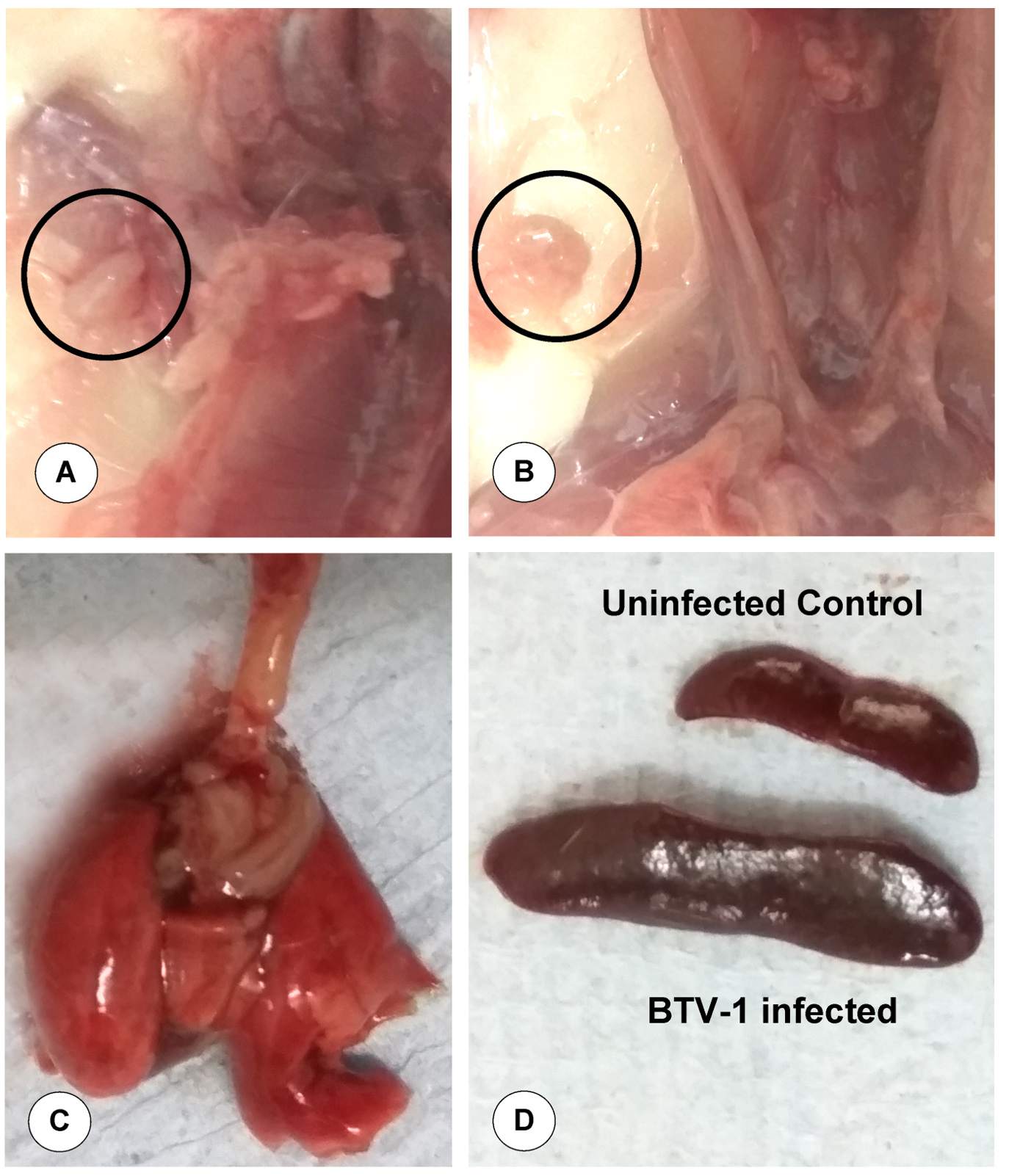
Figure 3. Gross pathological lesions in dam infected by BTV-1 during late stage of pregnancy. (A) Axillary lymph node showed marked enlargement and oedema on 6 dpi (21 GD). (B) Inguinal lymph node showed marked enlargement and oedema on 6 dpi. (C) Enlarged, oedematous, congested and rubbery consistency of lungs on 6 dpi. (D) Markedly enlarged and widespread oedematous spleen on 6 dpi
Histopathological lesions in fetuses
Severely congested alveolar capillaries, swollen endothelial cells (ECs), haemorrhages, and thickening of interalveolar septa were observed on fetal lungs on 3 dpi (18 GD). Perivascular hemorrhages due to endothelial damage and increased vascular permeability and hyalinization of tunica media was observed on 5 dpi/20 GD (Figure 4A). Pleuritis characterized by marked thickening of the pleura with infiltration of lymphocytes was observed on 6 dpi/21 GD (Figure 4B). Marked dilatation of epicardium with haemorrhages and severe congestion of blood vessels in myocardium were observed on 5 dpi/20 GD (Figure 4C). Lymph node showed lymphoid depletion and mild congestion in the cortex of 10-day-old fetuses (Figure 4D). Interstitial pneumonia with infiltration of mononuclear cells was noticed in 10-day-old foetuses. Severe meningitis characterized by meningeal thickening due to congestion (Figure 4E), swollen ECs and infiltration of lymphocytes was observed on 6 dpi. Hypertrophied ECs and perivascular edema were recorded in the 10-day-old fetuses. Focal proliferation, hypertrophy and accumulation of glial cells, and swollen ECs were recorded in cerebrum. Vasculitis with thickened wall and infiltration of MNCs were observed in cerebrum.
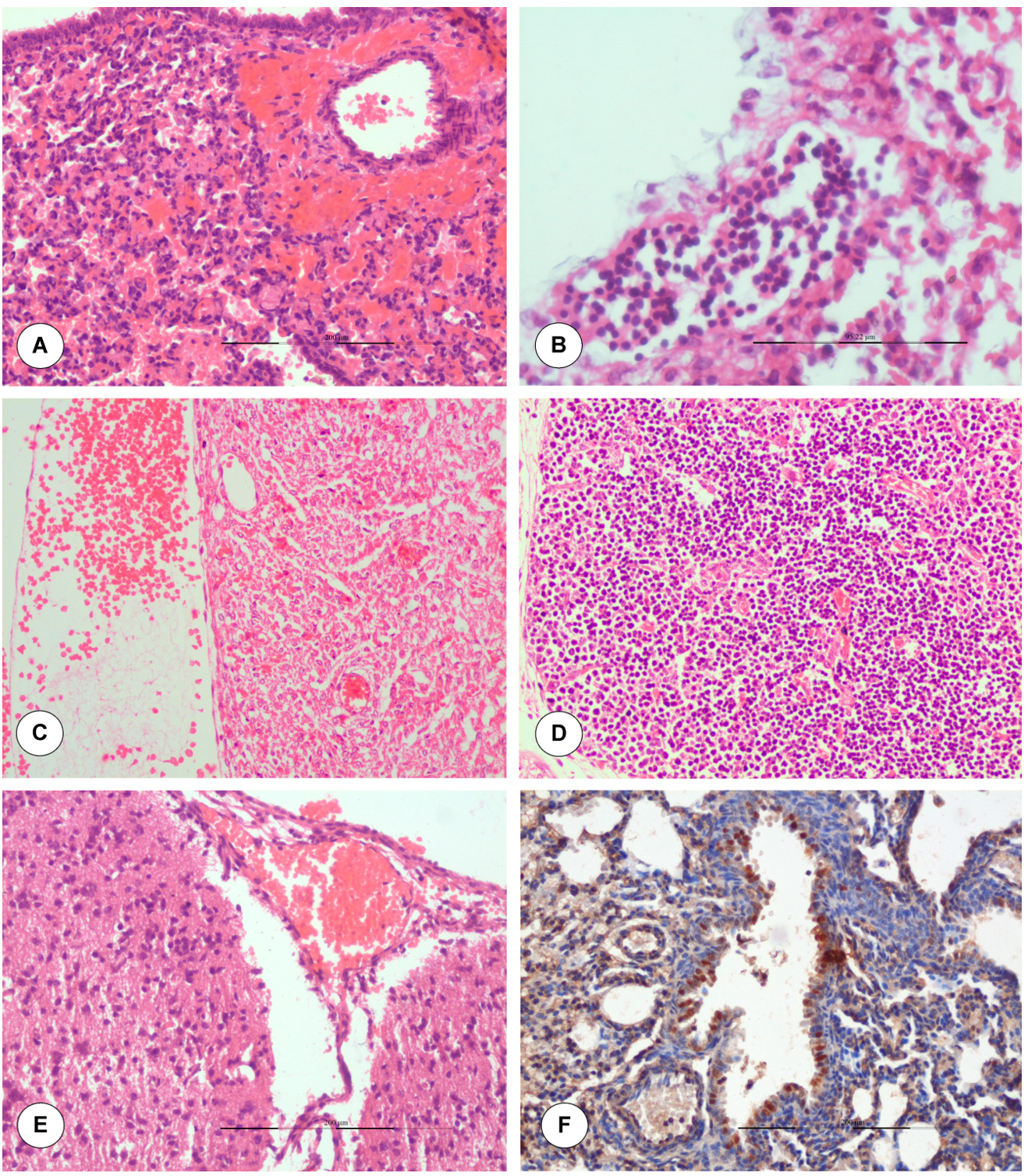
Figure 4. Histopathological lesions in BTV-1 infected fetuses during late stage of gestation. (A) Perivascular hemorrhages due to endothelial damage in fetal lungs on 5 dpi (20 GD). H&E x200. (B) Pleuritis: marked thickening of pleura with infiltration of lymphocytes in fetal lung on 6 dpi (21 GD). H&E x400. (C) Marked dilatation of epicardium with haemorrhages on 5 dpi (20 GD). H&E x200. (D) Moderate to severe lymphoid depletion in the cortex of lymph node H&E x200. (E) Marked congestion of meninges. H&E x200. (F). BTV-1 antigen showed positive immunolabelling in bronchiolar epithelium and alveolar macrophages. IP-DAB-MH x200
Histopathological lesions in dams
In the dam, necrosis and severe haemorrhages in the uterus were noticed on 6 dpi (Figure 5A). Multifocal necrotizing placentitis and infiltration of inflammatory cells were noticed in the labyrinth zone of placenta on 6 dpi (Figure 5B). Lymph node showed notable lymphoid depletion and congested capillaries with dilated endothelial cell were recorded on 5 dpi onwards. Germinal centre formation and lymphoid depletion were observed in white pulp of spleen on 6 dpi (Figure 5C).
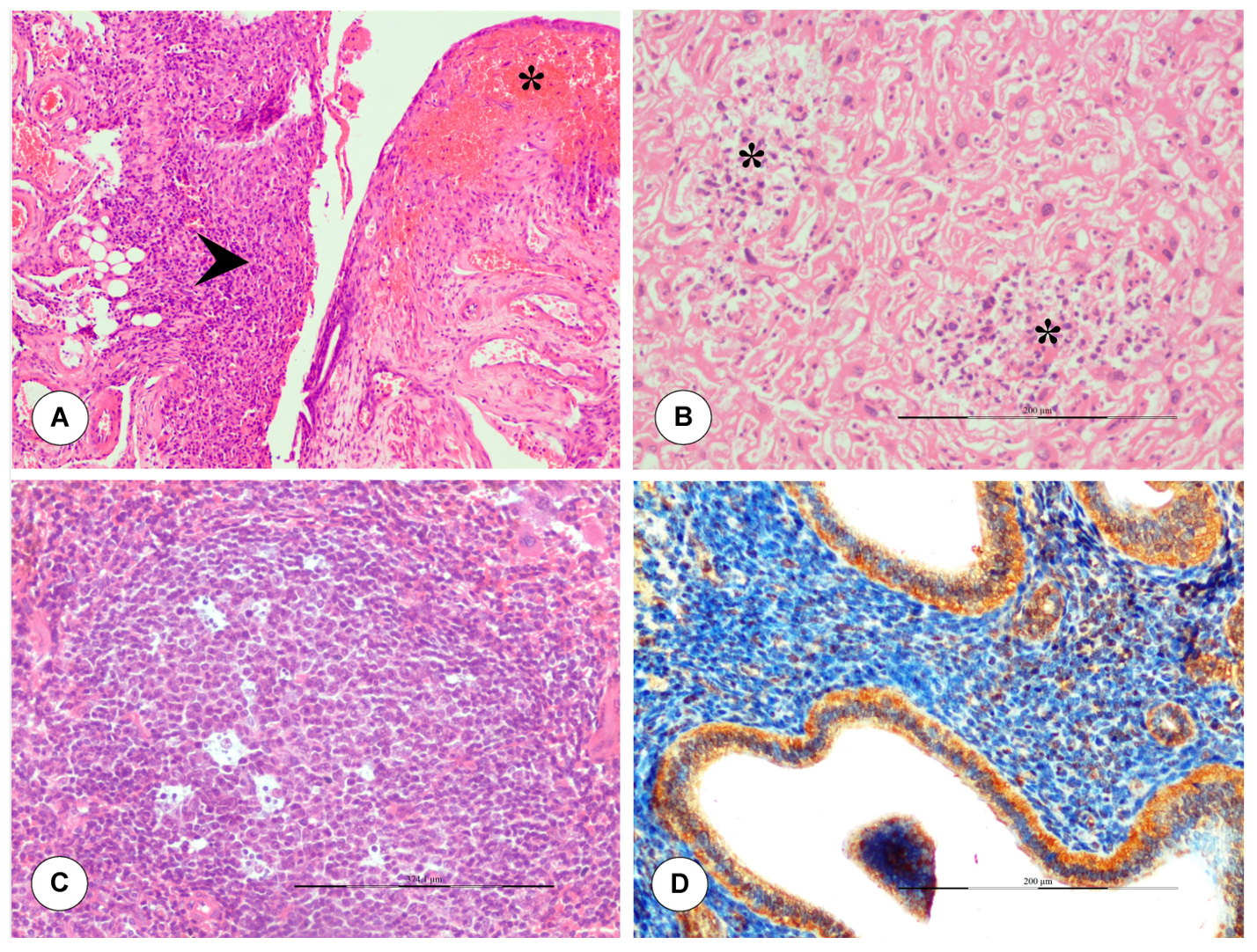
Figure 5. Histopathological lesions in dam infected with BTV during late stage of pregnancy. (A) Severe haemorrhages, necrosis (asterisk) and inflammatory cells infiltration (arrow) in uterus on 6 dpi. H&E x100. (B) Multifocal areas of necrosis and inflammatory cells infiltration were noticed in the placenta (labyrinth zone) on 6 dpi. H&E x200. (C) Spleen showed germinal centre formation with apoptosis of lymphocytes. H&E x200. (D) In uterus, BTV-1 antigen showed positive immunolabelling in endometrial glands, stromal cells, and luminal epithelium on 5 dpi. IP-DAB-MH x200
Localization of BTV-1 antigen by immuno-histochemistry
BTV-1 antigen was localized in representative tissue samples collected at different time intervals in brain, lungs, and spleen of fetuses and dam. In fetuses, BTV-1 positive signals were noticed in bronchiolar epithelium and alveolar macrophages of lungs (Figure 4F). In uterus, BTV-1 antigen showed positive immunolabelling in endometrial glands, stromal cells, and luminal epithelium on 5 dpi (Figure 5D). Localization of BTV-1 antigen was observed in the cytoplasm of endothelial cells, neurons, and microglial cells of brain in fetuses (Figure 6A). Brain from uninfected control fetuses showed no positive immunoreaction for BTV-1 antigen (Figure 6B). In spleen of fetuses, lymphocytes and histiocytes showed positivity (Figure 6C). In lymph node of dam, macrophages of medullary regions showed positivity (Figure 6D). Granulosa cells, theca interna and externa of follicles, and cortical stromal cells of ovary showed positive immunolabelling of BTV-1 antigen. Cortical stromal cells, small theca lutein cells and large granulosa lutein cells of corpus luteum of ovary showed positive immunolabelling of BTV-1 antigen (Figure 6E). The ovary of uninfected control mice did not show positive immunoreaction for BTV-1 antigen (Figure 6F).
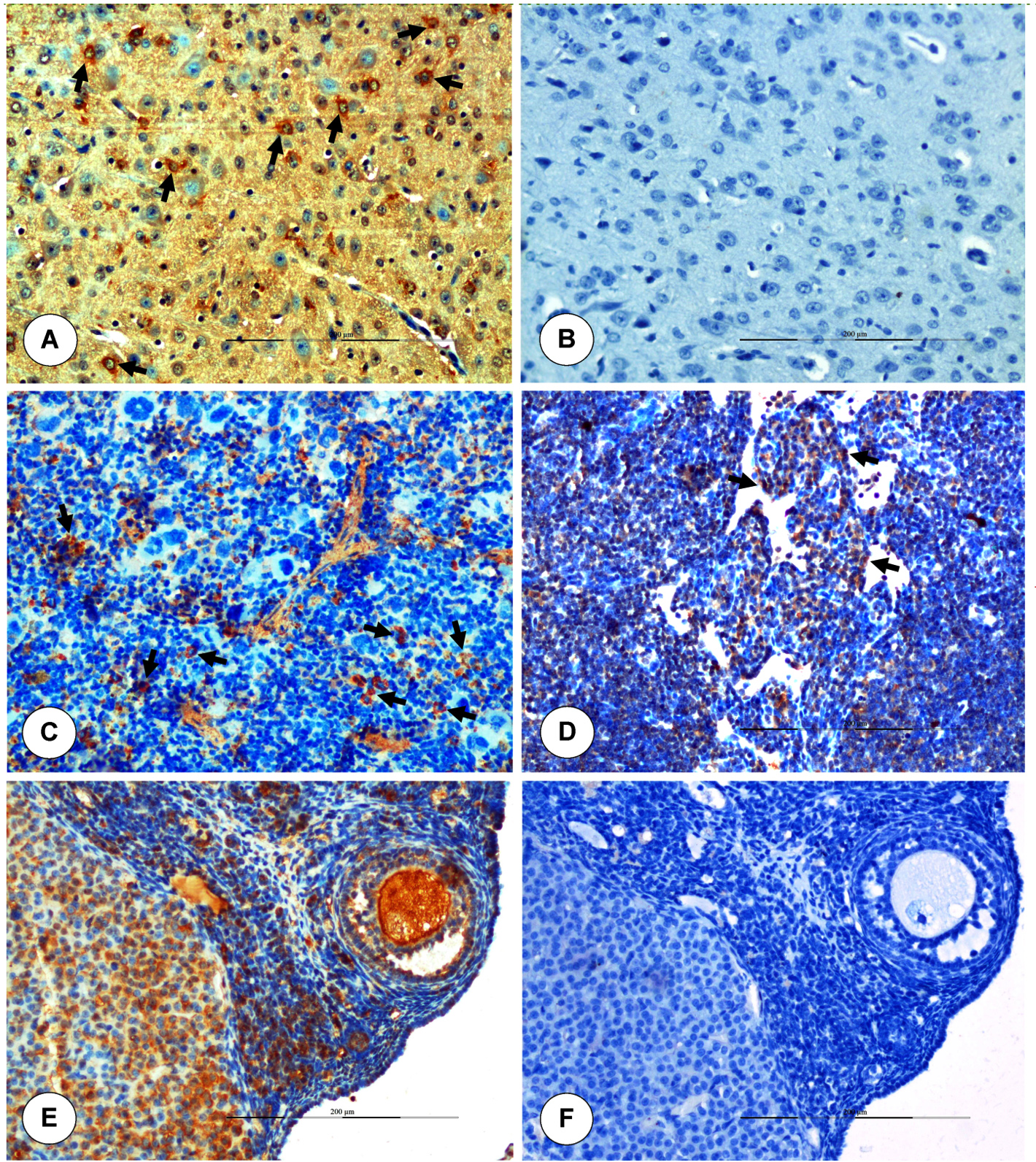
Figure 6. Immunohistochemical localization of BTV-1 antigen in internal organs of fetuses and dam. (A) Positive immunolabelling of BTV-1 antigen in the cytoplasm of neurons, endothelial cells and glial cells of brain. IP-DAB-MH x200. (B) Uninfected control: Brain showed no positive immunoreaction for BTV-1 antigen. IP-DAB-MH x200. (C) Positive immunolabelling of BTV-1 antigen in lymphocytes and histiocytes (arrow) of spleen. IP-DAB-MH x200. (D) Lymph node showed BTV-1 antigen in the medullary regions. IP-DAB-MH x200. (E) Positive immunolabelling of BTV-1 antigen in granulosa cells, theca interna and externa of follicles, lutein cells (small theca and large granulosa), and cortical stromal cells of corpus luteum of ovary. IP-DAB-MH x200. (F) Ovary did not show positive immunoreaction in uninfected control mice. IP-DAB-MH x200
Detection of BTV nucleic acid from blood and reproductive organs by RT-PCR
The BTV specific VP7 amplicons of 243 bp were detected from placenta, uterus, ovary and fetuses on 3 dpi (18 GD) onwards. The presence of viral nucleic acid was observed in 12 out of 15 (80.00%) placenta samples of BTV-infected dams collected during late gestation. The BTV nucleic acids were detected from 1 dpi (16 GD) in blood, lymph nodes and spleen on 3 dpi (18 GD), and lungs and brain on 5 dpi (20 GD) onwards in BTV-1 infected pregnant mice. The BTV was detected in fetuses born from the late stage of gestation till the experiment up to 10 days postnatal life.
Estimation of haematological parameters
The BTV-infected pregnant mice displayed a consistently decreased number of TLC up to 21 GD (Table 1). Markedly significant (P < 0.05) lower values of TLC were observed on 6 dpi (21 GD) when compared to uninfected control mice. The RBC and TPC levels revealed decreasing trend up to 21 GD in BTV-1 infected group than uninfected animals, and significantly (P < 0.05) low value was noticed on 21 GD. The Hb and PCV values in BTV-1 infected animals showed significantly (P < 0.05) decreased values from 1 dpi (16 GD) to 6 dpi (21 GD). Additionally, throughout the experiment, MCHC and MCV values of BTV-infected animals did not significantly differ from those of the control group (Table 1). The lymphocyte count showed decreasing trend from 1 to 6 dpi after BTV-1 infection than uninfected animals. The peak decrease in lymphocyte count was noticed on 6 dpi (21 GD); whereas, the neutrophils count in BTV-infected animals was significantly greater on 6 dpi in comparison with uninfected animals. Decrease in monocyte count was observed on 6 dpi in infected animals. Overall, in BTV infected animals, there was leukopenia (7.13 ± 0.85 to 9.15 ± 0.88 x103/µl), lymphopenia (61.35 ± 3.85 to 68.50 ± 3.25%) and neutrophilia (20.56 ± 1.75 to 28.85 ± 2.87%). Anaemia (decreased RBCs, Hb and PCV values) and thrombocytopenia (decreased TPC levels) were observed up to 6 dpi (21 GD) in BTV-1 infected group and statistically significant (P < 0.05) decrease was observed on 6 dpi when compared to uninfected control group. The MCV and MCHC values did not show any significant change in BTV-1 infected mice when compared to control group throughout experiment (Table 1).
No significant changes were observed in the haematological values of uninfected mice (Table 1).
Table (1):
Effect of BTV-1 infection on haematological values of late stage of pregnant mice at different time intervals (Mean ± SEM)
| Group | 1 dpi (16 GD) | 3 dpi (18 GD) | 5 dpi (20 GD) | 6 dpi (21 GD) |
|---|---|---|---|---|
| Red blood cells (x106/µl) | ||||
| BTV-1 | 5.50 ± 0.75 | 4.95 ± 0.52 | 4.55 ± 0.94 | 4.15 ± 0.88* |
| Control | 5.75 ± 0.78 | 5.24 ± 0.68 | 5.17 ± 0.69 | 5.88 ± 0.71 |
| Total leucocyte count (x103/µl) | ||||
| BTV-1 | 9.15 ± 0.88 | 8.75 ± 0.96* | 7.72 ± 0.78** | 7.13 ± 0.85** |
| Control | 10.86 ± 0.95 | 11.48 ± 1.22 | 12.25 ± 1.15 | 11.75 ± 1.13 |
| Total platelet count (x105/µl) | ||||
| BTV-1 | 9.85 ± 1.05 | 9.15 ± 1.15* | 8.75 ± 0.95* | 8.25 ± 1.25* |
| Control | 10.55 ± 1.09 | 11.25 ± 0.95 | 12.25 ± 1.15 | 11.85 ± 1.08 |
| Hemoglobin (g/dL) | ||||
| BTV-1 | 9.95 ± 0.59 | 9.25 ± 0.85 | 8.85 ± 0.95 | 8.17 ± 0.79* |
| Control | 10.25 ± 0.83 | 9.75 ± 0.74 | 9.15 ± 1.14 | 10.95 ± 1.05 |
| Hematocrit or packed cell volume (%) | ||||
| BTV-1 | 33.15 ± 2.29 | 32.35 ± 2.35 | 31.88 ± 2.15* | 31.24 ± 2.48* |
| Control | 34.75 ± 2.65 | 33.15 ± 2.76 | 34.85 ± 2.38 | 34.15 ± 2.55 |
| Lymphocytes (%) | ||||
| BTV-1 | 68.50 ± 3.25 | 65.35 ± 3.45* | 62.75 ± 3.16* | 61.35 ± 3.85* |
| Control | 70.35 ± 3.65 | 71.25 ± 3.08 | 68.85 ± 3.19 | 66.75 ± 3.54 |
| Neutrophils (%) | ||||
| BTV-1 | 20.56 ± 1.75 | 21.75 ± 1.48* | 25.58 ± 2.36* | 28.85 ± 2.87* |
| Control | 20.25 ± 2.05 | 18.55 ± 1.95 | 21.75 ± 2.58 | 21.55 ± 2.45 |
| Mean corpuscular volume (MCV; fL) | ||||
| BTV-1 | 44.95 ± 3.16 | 43.63 ± 3.05 | 42.79 ± 3.25 | 45.84 ± 2.85 |
| Control | 42.86 ± 2.75 | 44.25 ± 3.36 | 44.76 ± 2.56 | 43.73 ± 2.65 |
| Mean corpuscular hemoglobin concentration (MCHC; g/dL) | ||||
| BTV-1 | 37.85 ± 2.75 | 36.78 ± 2.83 | 35.54 ± 2.69 | 34.62 ± 2.93 |
| Control | 35.76 ± 2.69 | 35.86 ± 2.54 | 34.72 ± 2.96 | 33.65 ± 3.52 |
Values with superscript of asterisk (*) differs significantly in animals infected with BTV at specified time points when compared to uninfected animals (*P ≤ 0.05; ** P ≤ 0.01).
Estimation of serum biochemical parameters
The AST, ALT, CK, and ALP levels were significantly increased in BTV-1 infected animals in comparison with the uninfected control group (Table 2). The AST levels revealed significantly increased trend from 3 dpi (18 GD) to 6 dpi (21 GD), and the values varied between 120.32 ± 4.24 to 160.49 ± 5.82 IU/L. The significant (P < 0.05) increase in peak was observed on 6 dpi (160.49 ± 5.82 IU/L) in infected animals. The values of ALT significantly increased from 3 to 6 GD and the values varied between 85.16 ± 3.18 to 114.55 ± 4.62 IU/L when compared to uninfected animals. The values of ALT showed significant (P < 0.05) peak increase on 6 dpi (114.55 ± 4.62 IU/L) in infected animals (Table 2).
Table (2):
Effect of BTV-1 infection on serum biochemical values of late stage of pregnant mice at different time intervals (Mean ± SEM)
| Groups | 1 dpi (16 GD) | 3 dpi (18 GD) | 5 dpi (20 GD) | 6 dpi (21 GD) |
|---|---|---|---|---|
| Aspartate amino transferase (AST) or SGOT (IU/L) | ||||
| BTV-1 | 120.32 ± 4.24 | 131.45 ± 5.24* | 154.36 ± 4.41* | 160.49 ± 5.82** |
| Control | 122.70 ± 4.68 | 116.59 ± 4.52 | 124.16 ± 4.94 | 112.72 ± 4.60 |
| Alanine amino transferase (ALT) or SGPT (IU/L) | ||||
| BTV-1 | 85.16 ± 3.18 | 95.21 ± 3.37* | 105.39 ± 4.44* | 114.55 ± 4.62* |
| Control | 83.74 ± 3.66 | 84.79 ± 3.86 | 81.90 ± 4.30 | 83.52 ± 4.28 |
| Alkaline phosphatase (ALP) (IU/L) | ||||
| BTV-1 | 120.44 ± 4.36 | 128.27 ± 4.19 | 141.59 ± 4.64* | 146.87 ± 5.28* |
| Control | 116.51 ± 4.62 | 122.66 ± 4.39 | 120.88 ± 5.75 | 121.67 ± 4.42 |
| Creatine kinase (CK) (IU/L) | ||||
| BTV-1 | 140.54 ± 5.68 | 151.10 ± 5.25* | 174.32 ± 6.29** | 178.48 ± 5.75** |
| Control | 139.58 ± 4.84 | 141.84 ± 5.96 | 144.51 ± 5.82 | 142.62 ± 5.72 |
Values with superscript of asterisk (*) differs significantly in animals infected with BTV than uninfected animals at particular time points (*P ≤ 0.05; ** P ≤ 0.01).
The levels of ALP showed increasing trend on 5 and 6 dpi and significant (P < 0.05) increase in peak was recorded on 6 dpi (146.87 ± 5.28 IU/L) when compared to uninfected control group. The ALP values varied between 120.44 ± 4.36 to 146.87 ± 5.28 IU/L. The levels of CK showed an increasing trend in infected animals from 3 to 6 dpi. The significant (P < 0.05) increase in peak was observed on 6 dpi (178.48 ± 5.75 IU/L). The CK values varied between 140.54 ± 5.68 to 178.48 ± 5.75 IU/L (Table 2).
Assessment of humoral immune response
The BTV infected animals during late stage of pregnancy (15 GD) were seronegative up to 5 dpi (20 GD) and on 6 dpi (21 GD), the infected animals were showed positive (52.12%) for the BTV antibodies (Table 3).
Table (3):
c-ELISA OD values for detection of BTV specific antibodies during late stage of gestation at different intervals
| Group | Days post infection | Mean OD values | Percentage inhibition (PI) | Result |
|---|---|---|---|---|
| Mean of positive control | 0.155 | 88.65 | – | |
| Mean of negative control | 1.366 | – | – | |
| 50% of mean of negative control | 0.683 | – | – | |
| Late stage of pregnant mice | 1 dpi (16 GD) | 0.824 | 39.68 | Negative |
| 3 dpi (18 GD) | 0.749 | 45.17 | Negative | |
| 5 dpi (20 GD) | 0.705 | 48.39 | Negative | |
| 6 dpi (21 GD) | 0.654 | 52.12 | Positive | |
Study of developmental anomalies by Alizarin red staining
The skeletal deformities were characterized using Alizarin red staining in the fetuses born from the pregnant dam infected with BTV-1 during late stage gestation by comparing with uninfected control group. The length of fetuses and various bones in forelimbs, hind limbs, vertebral column length and skull showed a non-significant decrease when compared to uninfected control fetuses (Figures 2E & 2F, Table 4).
Table (4):
Effect of BTV-1 infection in the skeletal system of fetuses during late stage of gestation
| Groups | Length of fore limb (mm) | Length of hind limb (mm) | Length of vertebral column (mm; C7-S4) | Skull (mm) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Scapula | Humerus | Radius | Ulna | Femur | Tibia and fibula | Length | Diameter | ||
| Uninfected control | 1.82 ± 0.84 | 1.92 ± 0.73 | 1.97 ± 0.65 | 1.43 ± 0.58 | 1.67 ± 0.91 | 1.87 ± 0.76 | 13.45 ± 0.93 | 6.29 ± 0.76 | 3.25 ± 0.68 |
| Late stage of gestation (15-21) | 1.71 ± 0.65 | 1.82 ± 0.82 | 1.85 ± 0.86 | 1.32 ± 0.56 | 1.55 ± 0.76 | 1.76 ± 0.83 | 13.1 ± 0.72 | 5.85 ± 0.93 | 3.68 ± 0.78 |
The values differ non-significantly in animals infected with BTV than uninfected animals.
In the present study, transplacental transmission of field strain of Indian BTV in pregnant IFNAR1-blocked mice has been demonstrated experimentally for the first time. In addition, the study also demonstrated the localization of BTV in the uterus, placenta, foetuses, and ovary from the infected dams. Before 2006, transplacental infections by BTV serotypes were directly linked with the use of live attenuated vaccine viruses in sheep, cattle, and dogs, which had been a long-standing practice.19 Earlier, only four BTV serotypes or strains of modified live vaccine virus specifically, BTV-10, -11, -13, and -17 had been documented as causing TPT in ruminants.6,19
The BTV-1 serotype utilized in this study was isolated from the aborted fetuses, which provided concrete evidence of the TPT of the wild-type BTV-1 strain in goats.14 Furthermore, the emergence of BTV-8 infection in Europe in 2006 led to its rapid dissemination across northern Europe, resulting in a significant epizootic event. This outbreak caused a notable rise in abortion rates, stillbirths, the birth of weak or deformed offspring, and substantial economic losses in sheep, goats, and cattle, as documented by various studies.6,20 Moreover, van der Sluijs et al.11 have conclusively demonstrated the ability of BTV-1 to traverse the placental barrier and infect the fetus, leading to severe pathological consequences.
The TPT of natural or wild or field BTV serotypes in sheep, goats and cattle were reported in different experimental investigations13,21 and natural cases.7 Pregnant ewes infected with BTV-8 at 40-45 days of early gestation showed clinical signs (fever, rhinitis, conjunctivitis, and facial oedema at 6 to 9 dpi), gross pathological lesions like haemorrhages in different organs, abnormal content in the uterus and histopathologically, hepatitis, myocarditis and pneumonitis at 10 dpi. Necrotic suppurative bronchopneumonia, lymphocytic infiltration in portal triad, cholangio-hepatitis, microgranuloma in liver, and haemorrhages in various organs were noticed at 14 dpi. Transient fever, facial oedema, coronitis, oral ulcers, and anorexia were reported in ewes at 30 dpi.13 Lambs born from ewes infected with BTV-8 during early-gestation showed haemorrhages in the skin, and lymphocytic infiltration and haemorrhages in the placentomes at 10 dpi. Mummified fetuses, haemorrhage in various organs, lympho-histiocytic infiltration in meninges, and haemorrhages, necrosis and lymphocytic infiltration in placentome at 30 dpi.13
Pregnant ewes infected with BTV-8 at 70-75 days of mid gestation showed severe clinical signs (fever, facial oedema, loss of appetite, rhinitis, conjunctivitis, oral ulcers, salivation, and convulsions) and gross pathological lesions like necrotic haemorrhagic colitis, tubular necrosis and haemorrhages in the cortex of kidneys, haemorrhages in different tissues, and necrosis of placentome.13 Lambs born from ewes infected with BTV-8 during mid-gestation showed pathological lesions in the central nervous system like meningo-encephalitis with lympho-histiocytic infiltration in the cerebrum and cerebellum, vacuolation of the neuropil in the cerebrum and cyst in the cerebellar medulla, and spleen showed lymphoid depletion.13 The localization of BTV in the umbilical cord blood and foetal tissues like spleen, liver, thymus, and brain from ewes infected with wild-type BTV-8 serotype was reported.13 TPT occurred 10 days after the inoculation of ewes with BTV-8.
Belbis et al.21 reported for the first time, the TPT of the wild strain of BTV-8 in goats infected during the mid-stage of pregnancy (around 61 days of gestation) and virus was detected from the fetuses. The BTV-8 infected goats showed high rectal temperature, conjunctivitis, serous nasal discharge, and congestion of perineal skin and oral mucosa. Subcutaneous haemorrhage was reported in goats at 6 dpi. Pulmonary congestion and adhesions between the pleura and lungs were reported at 21 dpi. However, no significant gross and histopathological lesions were reported in the placenta of infected goats except recent haemorrhagic lesions (without haemosiderin) in placentomes.21
The current study reports the TPT of the wild-type Indian BTV-1 serotype after experimental infection in IFNAR1-blocked pregnant mice during the late stage of gestation. This phenomenon aligns with findings by Backx et al.,22 who reported the TPT of BTV-8 wild-type in the late stage of gestation in cattle under experimental conditions. Fetal infection during the late stage of gestation could lead to the birth of viremic calves up to six months. Studies have reported the TPT of BTV-8 in goats during the second trimester of gestation.23 In our study, we detected BTV in fetuses born during the late stage of gestation up to 10 days after birth and these findings are consistent with the observations of De Clercq et al.23, where they reported the viremic kids after delivery. Previous studies on the TPT of BTV-8 in cattle and sheep also demonstrated the viremic newborns by real-time PCR.6,23
Studies have reported that during the late stage of gestation in goats, mild intensity of clinical signs including fever, nasal discharge, conjunctivitis, and redness in the perineal skin were noticed.21,24 The effects of various BTV serotypes on the latter stages of pregnancy were initially investigated by direct intrauterine infection of the bovine fetuses.25 Serological investigations revealed that BTV infection during the early stage of pregnancy led to immunotolerance; whereas, infection during the late stage led to immunocompetence in the offspring.6,26-30
Saminathan et al.15 reported higher incidence of TPT in infected mice during the mid-stage (71.43%) when compared to the early stage (57.14%) of gestation. Abortion, reduced implantation sites, early embryonic death, and necrotic hemorrhagic lesions were documented during the early stage of gestation. Congenital defects and neurological abnormalities in fetuses, such as hemorrhage, diffuse cerebral edema, necrotizing encephalitis, and reduced bone dimensions were reported during the mid-stage of gestation. Saminathan et al.15 demonstrated the presence of BTV-1 antigen in various tissues including the mesometrium, embryo decidua, uterus, placenta, ovary, and foetal brain for the first time. In their study, BTV-infected mice showed seroconversion by 7 and 5 dpi onwards, and reached highest levels by 15 and 9 dpi during early and mid-stage of pregnancy, respectively.15
Drawing from literature review and knowledge, this study presents, for the first time, a comprehensive analysis of characterization of haematological and serum biochemical parameters, humoral immune responses of IFNAR1-blocked pregnant mice infected with BTV-1 during advanced stage of pregnancy.
The present study reported the higher rate of TPT during late-stage of pregnancy in experimental mouse model by Indian wild-type BTV-1 serotype. Common findings during late-stage of gestation in infected group were abortion in dam and neurological lesions in foetuses. BTV-1 antigen was demonstrated in different fetal organs and reproductive organs like placenta and uterus. Foetuses born from BTV-infected dams were viraemic, which points the role of either aborted or stillborn fetuses and the infected reproductive organs as the potential source of infection to other animals. Further researches are essential to explore the TPT potential of various Indian BTV serotypes. This IFNAR1-blocked immunocompetent mouse model can be used to test the efficacy of new vaccines for their potential to control the TPT of BTV.
ACKNOWLEDGMENTS
The authors are thankful to DBT-BBSRC, India and European Union, UK for providing research grant to this study. Authors are also thankful to the Director and Joint Director, CADRAD, ICAR-IVRI for providing all the facilities to carry out this research work.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
MS conceptualized the study, performed Investigation and visualization. SV applied methodology. SV, SP and AAPM performed formal analysis. SP, AAPM and SV performed data curation. SP, AAPM and KPS performed data validation. KPS supervised the study and project administration. MS wrote original draft. SKB and KD wrote, reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
The study was supported by research grant from DBT-BBSRC, India and European Union, UK (Grant No. BT/IN/Indo-UK/FADH/46/SM/2013, dated 8th Sep. 2014 and EU H20:20 PALE-Blu Grant).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study was approved by the Joint Directorate of Research, ICAR-Indian Veterinary Research Institute, Izatnagar, Uttar Pradesh, India, with approval no. F26-1/2015-16/JD(R).
- Saminathan M, Singh KP, Khorajiya JH, et al. An updated review on Bluetongue virus: Epidemiology, pathobiology, and advances in diagnosis and control with special reference to India. Vet Q. 2020a;40(1):258-321.
Crossref - MacLachlan NJ, Osburn BI. Induced brain lesions in calves infected with bluetongue virus. Vet Rec. 2008;162(15):490-491.
Crossref - Maclachlan NJ, Osburn BI. Teratogenic bluetongue and related orbivirus infections in pregnant ruminant livestock: timing and pathogen genetics are critical. Curr Opin Virol. 2017;27:31-35.
Crossref - Mayo CE, Mullens BA, Gibbs EPJ, MacLachlan NJ. Overwintering of Bluetongue virus in temperate zones. Vet Ital. 2016;52(3-4):243-246.
Crossref - van der Sluijs MTW, de Smit AJ, Moormann RJM. Vector independent transmission of the vector-borne bluetongue virus. Crit Rev Microbiol. 2016;42(1):57-64.
Crossref - Saegerman C, Bolkaerts B, Baricalla C, et al. The impact of naturally occurring, trans-placental bluetongue virus serotype-8 infection on reproductive performance in sheep. Vet J. 2011;187(1):72-80.
Crossref - Zanella G, Durand B, Sellal E, et al. Bluetongue serotype 8: abortion and transplacental transmission in cattle in the Burgundy region, France, 2008-2009. Theriogenology. 2012;77(1):65-72.
Crossref - Coetzee P, Stokstad M, Myrmel M, et al. Transplacental infection in goats experimentally infected with a European strain of bluetongue virus serotype 8. Vet J. 2013;197(2):335-341.
Crossref - Rasmussen LD, Savini G, Lorusso A, et al. Transplacental transmission of field and rescued strains of BTV-2 and BTV-8 in experimentally infected sheep. Vet Res. 2013;44(1):75.
Crossref - Shaw AE, Ratinier M, Nunes SF, et al. Reassortment between two serologically unrelated bluetongue virus strains is flexible and can involve any genome segment. J Virol. 2013;87(1):543-557.
Crossref - van der Sluijs MTW, Schroer-Joosten DPH, Fid-Fourkour A, et al. Transplacental Transmission of Bluetongue Virus Serotype 1 and Serotype 8 in Sheep: Virological and Pathological Findings. PLoS ONE. 2013;8(12):e81429.
Crossref - Batten CA, Maan S, Shaw AE, Maan NS, Mertens PPC. A European field strain of bluetongue virus derived from two parental vaccine strains by genome segment reassortment. Virus Res. 2008;137(1):56-63.
Crossref - van der Sluijs M, Timmermans M, Moulin V, et al. Transplacental transmission of Bluetongue virus serotype 8 in ewes in early and mid gestation. Vet Microbiol. 2011;149(1-2):113-125.
Crossref - Chauhan HC, Biswas SK, Chand K, et al. Isolation of bluetongue virus serotype 1 from aborted goat fetuses. Rev Sci Tech. 2014;33(3):803-812.
Crossref - Saminathan M, Singh KP, Vineetha S, et al. Virological, immunological and pathological findings of transplacentally transmitted bluetongue virus serotype 1 in IFNAR1-blocked mice during early and mid gestation. Sci Rep. 2020b;10(1):2164.
Crossref - Byers SL, Wiles MV, Dunn SL, Taft RA. Mouse estrous cycle identification tool and images. PLoS One. 2012;7(4):e35538.
Crossref - Saminathan M, Singh KP, Maity M, et al. Pathological and immunological characterization of bluetongue virus serotype 1 infection in type I interferons blocked immunocompetent adult mice. J Adv Res. 2021;31:137-153.
Crossref - Luna LG. Manual of histologic staining methods of the Armed Forces Institute of Pathology; 3rd ed., Blakiston Divsion;1968:xii-258.
- MacLachlan NJ, Conley AJ, Kennedy PC. Bluetongue and equine viral arteritis viruses as models of virus-induced fetal injury and abortion. Anim Reprod Sci. 2000;60-61:643-651.
Crossref - Wilson AJ, Mellor PS. Bluetongue in Europe: past, present and future. Philos Trans R Soc Lond B Biol Sci. 2009;364(1530):2669-2681.
Crossref - Belbis G, Breard E, Cordonnier N, et al. Evidence of transplacental transmission of bluetongue virus serotype 8 in goats. Vet Microbiol. 2013;166(3-4):394-404.
Crossref - Backx A, Heutink R, van Rooij E, van Rijn P. Transplacental and oral transmission of wild-type bluetongue virus serotype 8 in cattle after experimental infection. Vet Microbiol. 2009;138(3-4):235-243.
Crossref - De Clercq K, Vandenbussche F, Vandemeulebroucke E, et al. Transplacental bluetongue infection in cattle. Vet Rec. 2008;162(17):564.
Crossref - Dercksen D, Groot NN, Paauwe R, Backx A, van Rijn PA, Vellema P. First outbreak of bluetongue in goats in the Netherlands. Tijdschr Diergeneeskd. 2007;132(20):786-790.
- Waldvogel AS, Anderson CA, Phillips DL, Osburn BI. Infection of bovine fetuses at 120 days’ gestation with virulent and avirulent strains of bluetongue virus serotype 11. Comp Immunol Microbiol Infect Dis. 1992;15(1):53-63.
Crossref - Vercauteren G, Miry C, Vandenbussche F, et al. Bluetongue virus serotype 8-associated congenital hydranencephaly in calves. Transbound Emerg Dis. 2008;55(7):293-298.
Crossref - Saminathan M, Rana R, Ramakrishnan MA, Karthik K, Malik YS, Dhama K. Prevalence, diagnosis, management and control of important diseases of ruminants with special reference to Indian scenario. J Exp Biol Agric Sci. 2016;4(3S):338-367.
Crossref - Singh KP, Saminathan M, Dinesh M, et al. Epidemiology and pathology of bluetongue virus in India: A systematic review. Indian J Vet Pathol. 2021;45(3):151-174.
Crossref - Saminathan M, Singh KP, Vineetha S, et al. Factors determining the clinical outcome of bluetongue virus infection in adult mice. Indian J Vet Pathol. 2018;42(4):239-248.
Crossref - Saminathan M, Singh KP, Rajasekar R, et al. Role of type I interferons in the pathogenesis of bluetongue virus in mice and ruminants. J Exp Biol Agric Sci. 2019;7(6):513-520.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.