


ISSN: 0973-7510
E-ISSN: 2581-690X
Virulence-associated genes allow plant pathogenic bacteria to adapt effectively to their hosts. Among them, the function of type III effectors (T3Es) has been well documented. T3Es are introduced directly into host cells via the type III secretion system (T3SS). The presence of five Type III effector genes was tested in thirty-four X. citri pv. malvacearum isolates from major cotton-growing regions of India. Thirteen isolates carried the avrBs3 avirulence gene. Eight isolates were positive for the xopK gene, seven for the HopG1 gene, six for the xopAD gene, and five for the xopI gene. Defense gene expression was studied in cotton using PR-1 (chitinase), PR-5 (ribonuclease-like protein), and polyubiquitin as an internal control in a susceptible cotton cultivar (LRA5166) by quantitative real-time PCR. The differential gene expression profiles were studied in cotton leaves by pre-treatment of cotton seedlings with effective biocontrol agents such as Streptomyces mutabilis (RAL2) and Bacillus amyloliquefaciens (ETL2), along with salicylic acid (1000 ppm), before X. citri pv. malvacearum infection. Plants treated with S. mutabilis (RAL2) showed numerically maximum expression of the defense gene PR1 compared to those treated with B. amyloliquefaciens (ETL2). PR5 gene expression was numerically higher in plants treated with B. amyloliquefaciens (ETL2) than in those treated with S. mutabilis (RAL2).
Type III Effectors (T3Es), xopAD, avrBs3, HopG1, xopI and xopK, Xanthomonas citri pv. malvacearum, Cotton, Defense Genes (PR1 and PR5)
Cotton, often called white gold, is a major economic crop grown in India. It is affected by several diseases from the seedling to maturity. Among them, bacterial blight caused by Xanthomonas citri pv. malvacearum (Xcm) is a significant disease that causes crop yield losses in numerous cotton-growing regions worldwide,1,2 including India.3 Effectors are bacterial proteins injected into plant cells via the type III secretion system. They suppress plant defence signalling pathways, enabling the pathogen to evade the plant’s immunity. Type III effectors (T3E) in Xanthomonas citri pv. malvacearum (Xcm) are essential for its pathogenicity in cotton. Type III effectors are primary factors in Xcm-mediated disease development in cotton plants and in the pathogen’s escape from host resistance. Cotton Xcm injects various effector proteins into plant cells via the type III secretion system, aiding the pathogen’s nutrient absorption and disease development.4 The AvrBs3/PthA (TAL) family effectors activate host genes, while Xops effectors influence host defence pathways to enhance pathogen virulence or induce plant resistance. The variation among these effectors shapes the host range and virulence races of Xcm.
The role of type III effectors (T3Es) is among the best documented of virulence-related genes, helping explain how plant pathogenic bacteria adapt to their hosts. T3Es are inoculated into host cells via the type III secretion system (T3SS). It is a highly conserved protein secretion system encoded by a cluster of hrp (hypersensitive response and pathogenicity) genes.5 Avr proteins are a type of type III effectors that trigger disease resistance in hosts possessing specific resistance (R) genes, serving as determinants of race-cultivar compatibility specificity.6 Effector proteins are crucial for the virulence and host specificity of all bacterial species that possess the T3 secretion system.7 T3E repertoires vary both between species and among strains within a species, and are therefore considered key determinants of the range of host interactions.8 Over twenty (20) effector genes have been identified within the genus Xanthomonas.9 The effector genes determine the race of the pathogen and control the pathogen’s aggressiveness.10 Xanthomonads have evolved effectors that employ a variety of mechanisms to enhance virulence, including processes unique to them and to eukaryotes.11 X. citri pv. malvacearum isolate race 18 contained 3-5 more effectors than other strains.2 Different races of X. citri pv. malvacearum have various combinations of avirulence (avr) genes, which define races and influence cultivar specificity.12
PR-1, one of the many PR gene families, has often served as a marker for systemic acquired resistance in numerous plant species.13 Transgenic tobacco and Arabidopsis expressing the PR1 gene are reported to confer a high level of resistance to bacterial diseases.14 Patil et al.15 identified the genes that were up-regulated in cotton IM216, resistant to bacterial blight, after inoculation with X. citri pv. malvacearum, using the microarray analysis technique. They found 98% of genes were significantly up-regulated at one or more sampling times, and 63% matched plant genes involved in defense responses, including those related to disease, protein synthesis, secondary metabolism, signalling, stress, programmed cell death, or encoding pathogenesis-related or retrotransposon-like proteins. Chandrasekaran and Chun16 reported that B. subtilis induced PAL genes, thereby conferring soft-rot disease resistance. Yim et al.17 observed higher accumulation of PR proteins in tomato plants inoculated with Methylobacterium when challenged with Xanthomonas campestris pv. vesicatoria and Pseudomonas syringae pv. tomato.
This study was conducted to understand how pathogen virulence relates to effector diversity and host defence gene expression with the following objectives.
- Detection of various effector genes in the pathogen population and
- Defence gene expression analysis in cotton using effective bacterial biocontrol agents challenged with the pathogen.
Type III- effector gene diversity in X. citri pv. malvacearum
The presence of Type III effector genes in 34 X. citri pv. malvacearum (Xcm) isolates from India were assessed. Partial sequences of the effector genes, namely xopAD (954 bp), avrBs3 (374 bp), HopG1 (232 bp), xopI (573 bp), and xopK (467 bp), were amplified using PCR with gene-specific primers. The primers were designed for the avrBs3, xopI, and xopK genes by retrieving the sequences of the products of the avirulence protein AvrBs3 and T3SS effector proteins XopI and XopK of X. citri pv. malvacearum strain AR81009, complete genome (accession number: CP023155.1) from NCBI. The primer for the xopAD gene was designed using the sequence of the avirulence protein XopAD from the complete genome of the Xcm strain MS14003 (accession number: CP023159.1). The primer for the HopG1 gene was designed through the type III secretion system effector protein HopG1 of Xcm strain XcmN1003, complete genome (accession number: NZ_CP013006.1). The primers were designed using the Primer3 software available in NCBI (Table 1).
Table (1): Designed primer sequences used for PCR amplification
| gene | Primer sequences | |
|---|---|---|
| Forward | Reverse | |
| xopAD | GAGCTAACGGAGGATCAGGC | CAGCAGAAGAACTGACGGGT |
| avrBs3 | TCAACTCAAACGCCGGATCA | TCTTCGTTGAATGCCGGGAA |
| HopG1 | GCTATCGCAAGAGCAAACCG | TGCGTTGAGTTTGAAGTGCG |
| xopI | CACTCTCGGTGCTGTCCAAT | GCTTGACGAATGACCTGGGA |
| xopK | GCGATCGATACCAACGCCTA | ACGACAAAGCCCTTGTTCCA |
The PCR amplifications were performed with the following cycling conditions as described by Hajri et al.18 with slight modifications. The initial denaturation at 94 °C for 2 minutes was succeeded by 35 cycles of 94 °C for one minute (denaturation) and 60 – 63 °C (depending on gene: xopAD – 63 °C, avrBs3 – 60 °C, HopG1 – 61 °C, xopI – 62 °C and xopK – 61 °C) for one minute (annealing) and 72 °C for one minute (extension). The final extension was performed at 72 °C for 10 minutes. The 10 µL PCR reaction mixture consisted of 2 µL of template DNA (50 ng/mL), 1 µL of each forward and reverse primer, 5 µL of PCR master mix (Smart Prime 2X master mix-Red), and 1 µL of sterile water. PCR amplifications were performed using a thermocycler (Eppendorf Master cycler Nexus Gradient S, Eppendorf AG, Hamburg, Germany). The presence or absence of a band of the expected size was used to determine whether a specific effector gene was present in the isolates.
Defense gene expression analysis in cotton against X. citri pv. malvacearum using qRT-PCR
The present study was intended to investigate the differences in the mRNA abundance of defence genes in cotton, such as PR-1 (chitinase) and PR-5 (ribonuclease-like protein), using polyubiquitin19 as an internal control, which is a non-regulated reference gene in a susceptible cotton cultivar, LRA5166, following infection with X. citri pv. malvacearum (MNSu) isolate. The differential gene expression profiles were studied in cotton leaves by pre-treatment of cotton seedlings with effective biocontrol agents such as Streptomyces mutabilis (RAL2) and Bacillus amyloliquefaciens (ETL2), along with salicylic acid (1000 ppm), before X. citri pv. malvacearum infection. The primers were designed for the PR1 gene through retrieving the sequences of the product of pathogenesis-related protein 1 (PR1) gene in Gossypium hirsutum cultivar TM-1, chromosome 1 (accession number: NC_030074.1) and the PR5 gene using the product of pathogenesis-related protein 5 (PR5) gene in Gossypium hirsutum cultivar TM-1, chromosome 4 (accession number: NC_030077.1). Primer design was carried out using the Primer3 software available at NCBI (Table 2).
Table (2): Designed primer sequences used for qRT-PCR amplification
| gene | Primer sequences | |
|---|---|---|
| Forward | Reverse | |
| PR1 | CCATAAGGTCCACCGGAGTG | ACTACCTAGCCTTGGGAGGG |
| PR5 | TTCGGCTATGGAAACAGGCA | TGGCCTGCAATAATCACCGA |
| Polyubiquitin (internal control) | GAAGGCATCCCACCAGACCAG | AACAGGGCCAGACATCACAATCAT |
Defence gene expression after biocontrol agent application was confirmed by quantitative real-time PCR (qRT-PCR) using the Step One Plus detection system (Applied Biosystems, USA). Treatments were imposed on 20 day old cotton seedlings of the susceptible cultivar LRA5166 raised in pots. The biocontrol agents were applied as a foliar spray at 10 ml/plant, containing 108 CFU/ml. Salicylic acid (1000 ppm) was used as a chemical control. The highly virulent X. citri pv. malvacearum isolate, 24 hours old, was spray-inoculated onto cotton seedling leaves 24 hours post-treatment at a concentration of 2 x 106 CFU/ml. Control plants, including both inoculated and uninoculated, were also maintained.
Leaf samples were collected 6 days after inoculation with the pathogen for each treatment and immediately preserved in liquid nitrogen. The samples were ground to a fine powder using a sterile DEPC-treated pestle and mortar. The Spectrum Total Plant RNA Isolation Kit (Sigma-Aldrich, USA) was used to isolate total RNA according to the manufacturer’s instructions. The RNA was treated with 1 Unit of DNase (Thermo Scientific, USA) according to the manufacturer’s instructions, incubated at 37 °C for 30 minutes, and the reaction was halted by adding 1 µl of EDTA, then incubated at 65 °C for 10 minutes. Total RNA treated with DNase was reverse-transcribed into single-stranded DNA using the Transcriptor High Fidelity cDNA Synthesis Kit (Roche, Germany). The resulting cDNA was then used for qRT-PCR analysis.
Quantitative RT-PCR was performed on single-stranded DNA from all five samples using standard cycling conditions (10 min at 95 °C, 40 cycles of 95 °C for 15 sec and 60 °C for 60 sec), followed by melt curve analysis to assess amplification specificity. The reaction mixture includes 7.0 µl of SYBR Green Master Mix (Roche Diagnostics), 0.5 µl of each forward and reverse primer specific to the gene, and 2 µl of cDNA at 50 ng/µl. ‘No template controls’ (NTC) were kept to prevent cross-contamination of reagents and surfaces. The NTC contains all RT-PCR reagents except the cDNA template. Relative mRNA quantities were determined using the comparative Ct method. For each sample, three biological replicates and two technical replicates were maintained, with polyubiquitin serving as the endogenous reference gene for normalising Ct values. Relative fold-changes were determined using the 2-∆∆Ct method, as outlined by Livak and Schmittgen.20 Statistical analyses were performed using R software. Differences in gene expression among treatments were assessed by one-way analysis of variance (ANOVA), followed by Tukey’s honestly significant difference (HSD) test. Data were presented as fold expression level. Differences for gene expression to different treatments were statistically compared at P < 0.05.
Type III- effector gene diversity in X. citri pv. malvacearum
Thirteen X. citri pv. malvacearum isolates out of 34, viz., MNSu, MNSn, MND, MNN, MNAn, MNR, MAB, KDR, KDB, TCS, TCD, TWB, and PC-CICR, showed the presence of the avrBs3 avirulence protein gene with an amplicon size of 374 bp. Of 10 Maharashtra isolates, 7 (70%) were positive for the avrBs3 gene. The effector (type III secretion system) protein gene xopK (467 bp) was identified in eight isolates (MNSn, MAB, MRA, PFN, PFB, PFF, KDR, and KDB) (Table 3 and Figure 1).
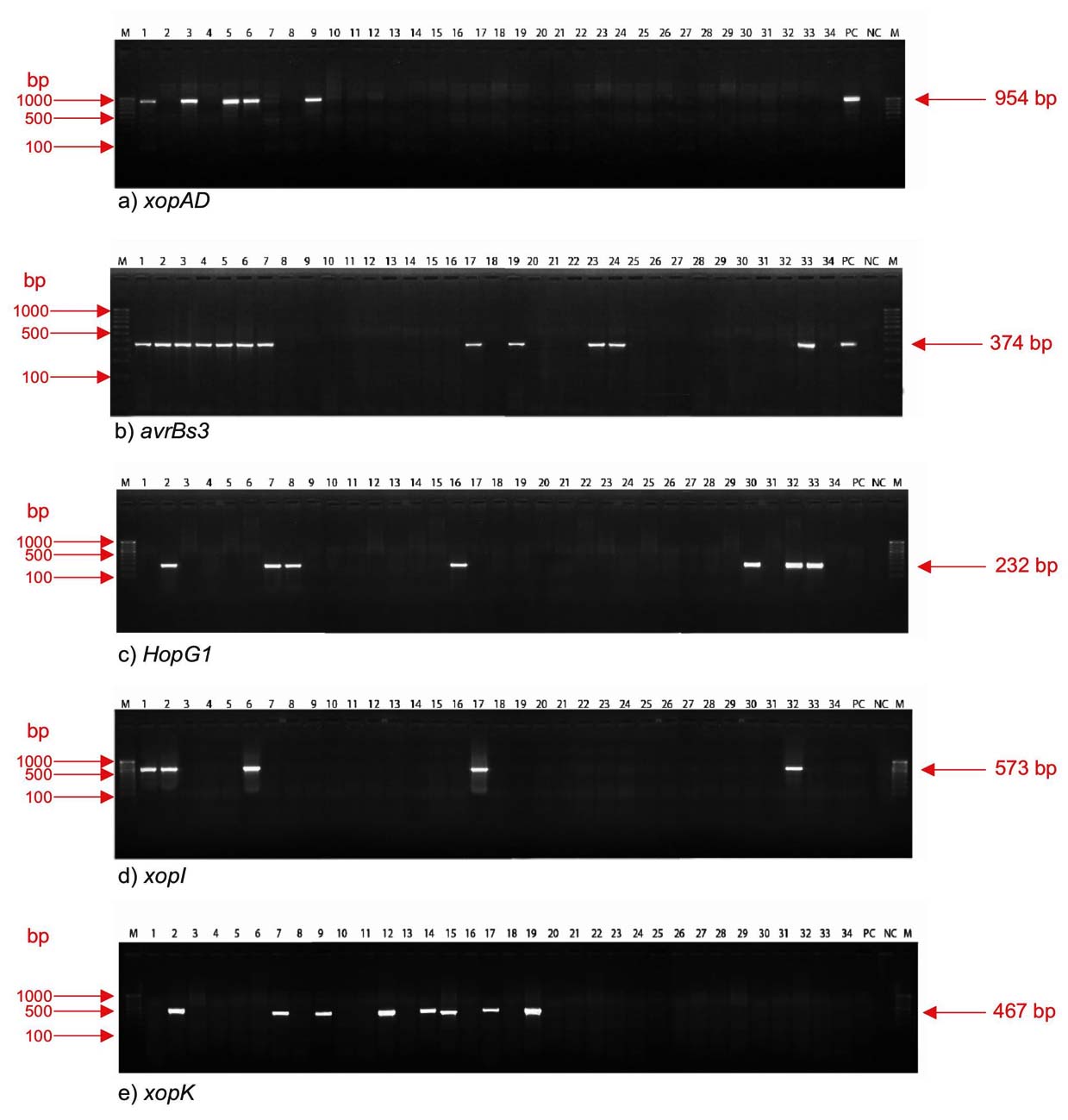
Figure 1. Molecular detection of effector genes of X. citri pv. malvacearum isolates using genes specific primers.
a) xopAD, b) avrBs3, c) Hop G1, d) хорl, е) хорK. М: 100 bp ladder, Lanes 1 to 34: MNSu, MNSn, MND, MNR, MNN. MNAn, MAB, MAA, MRA, MRN, PBH, PFN, PFA, PFB, PFF, KDS, KDR, KDG, KDB, KDSP, TCSrb, TCT, TCD, TCS, TCB, TCL, TDR, TDJ, ANR, AGD, AGR, TWR, TWB and THV, respectively. Lane 35: positive control (PC-CICR)
Table (3): Molecular detection of effector genes of X. citri pv. malvacearum isolates using gene-specific primers
| No. | Isolate Code | Effector genes | ||||
|---|---|---|---|---|---|---|
| xopAD | avrBs3 | HopG1 | xopI | xopK | ||
| 1 | MNSu | + | + | – | + | – |
| 2 | MNSn | – | + | + | + | + |
| 3 | MND | + | + | – | – | – |
| 4 | MNR | – | + | – | – | – |
| 5 | MNN | + | + | – | – | – |
| 6 | MNAn | + | + | – | + | – |
| 7 | MAB | – | + | + | – | + |
| 8 | MAA | – | – | + | – | – |
| 9 | MRA | + | – | – | – | + |
| 10 | MRN | – | – | – | – | – |
| 11 | PBH | – | – | – | – | – |
| 12 | PFN | – | – | – | – | + |
| 13 | PFA | – | – | – | – | – |
| 14 | PFB | – | – | – | – | + |
| 15 | PFF | – | – | – | – | + |
| 16 | KDS | – | – | + | – | – |
| 17 | KDR | – | + | – | + | + |
| 18 | KDG | – | – | – | – | – |
| 19 | KDB | – | + | – | – | + |
| 20 | KDSp | – | – | – | – | – |
| 21 | TCSrb | – | – | – | – | – |
| 22 | TCT | – | – | – | – | – |
| 23 | TCD | – | + | – | – | – |
| 24 | TCS | – | + | – | – | – |
| 25 | TCB | – | – | – | – | – |
| 26 | TCL | – | – | – | – | – |
| 27 | TDR | – | – | – | – | – |
| 28 | TDJ | – | – | – | – | – |
| 29 | ANR | – | – | – | – | – |
| 30 | AGD | – | – | + | – | – |
| 31 | AGR | – | – | – | – | – |
| 32 | TWR | – | – | + | + | – |
| 33 | TWB | – | + | + | – | – |
| 34 | THV | – | – | – | – | – |
| 35 | PC-CICR | + | + | – | – | – |
(+) – Presence of effector gene; (-) – Absence of effector gene
Seven isolates (MNSn, MAB, MAA, KDS, AGD, TWR, and TWB) showed positive amplification of the type III secretion system effector protein gene HopG1 (232 bp). Avirulence protein gene xopAD (954 bp) was recorded in six isolates (MNSu, MND, MNN, MNAn, MRA and PC-CICR). The other effector protein gene, xopI (573 bp), also involved in the type III secretion system, was detected in five isolates, including MNSu, MNSn, MNAn, KDR and TWR (Table 3 and Figure 1).
Defense gene expression analysis in cotton against X. citri pv. malvacearum using qRT-PCR
Plants treated with S. mutabilis (RAL2) showed maximum expression of the defence gene PR1 compared to those treated with B. amyloliquefaciens (ETL2). The PR5 gene expression was higher in B. amyloliquefaciens (ETL2) treated plants, followed by S. mutabilis (RAL2). However, the salicylic acid (1000 ppm)- treated plants showed much higher expression of both genes than the biocontrol agents. The pathogen-inoculated and uninoculated controls expressed low levels of PR1 and PR5 genes (Figures 2 and 3).
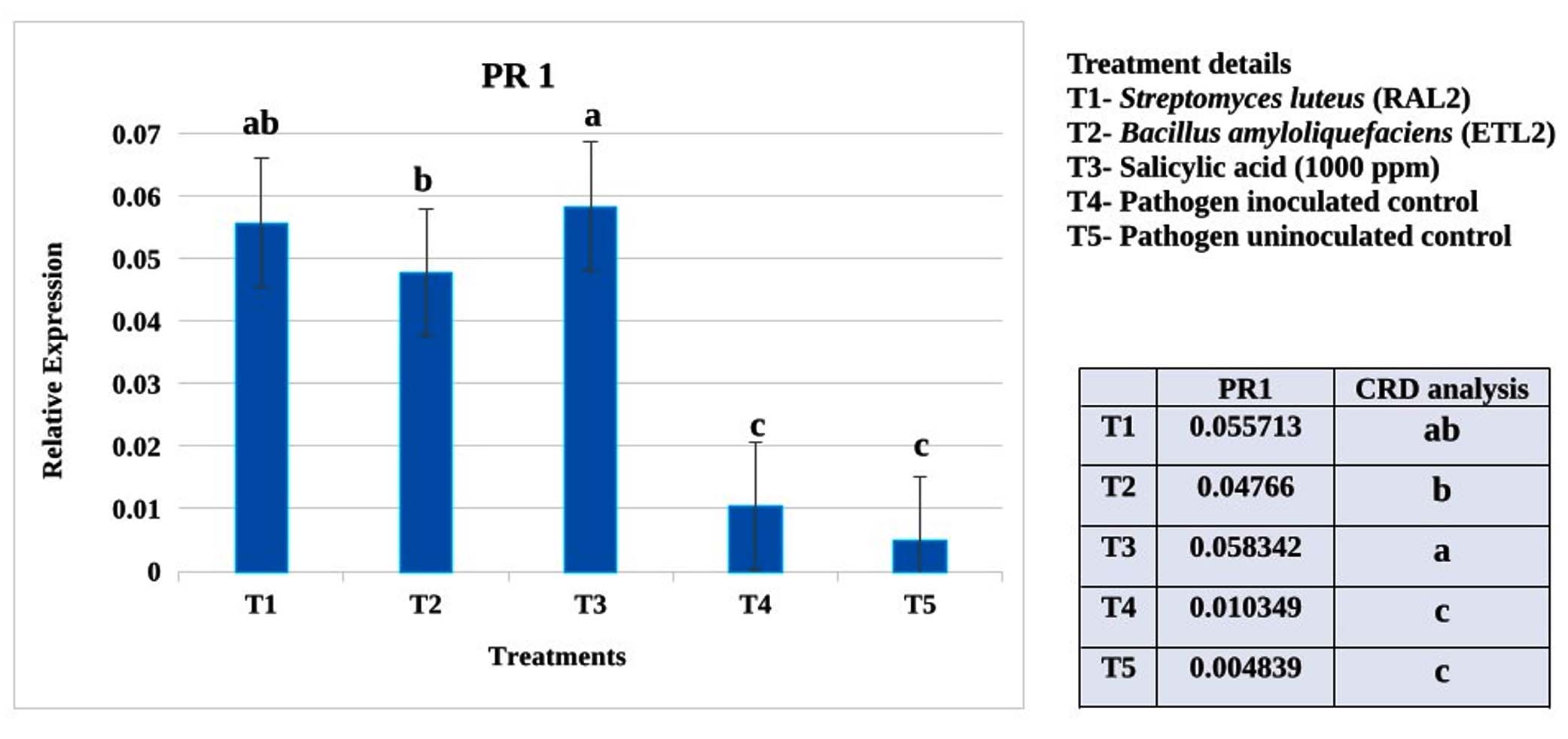
Figure 2. Expression analysis of defense gene PR1 in cotton seedlings (LRA5166) treated with biocontrol agents and chemical inducer challenge inoculated with X. citri pv. malvacearum (MNSu) using qRT-PCR. Polyubiquitin gene was used as internal control. Statistical analyses were performed using R software. Differences in gene expression among treatments were assessed by one-way analysis of variance (ANOVA), followed by Tukey’s honestly significant difference (HSD) test. Data is presented as fold expression level. Error bars indicated standard error obtained from three replicates per treatment. Means followed by different letters are significantly different for gene expression at P < 0.05
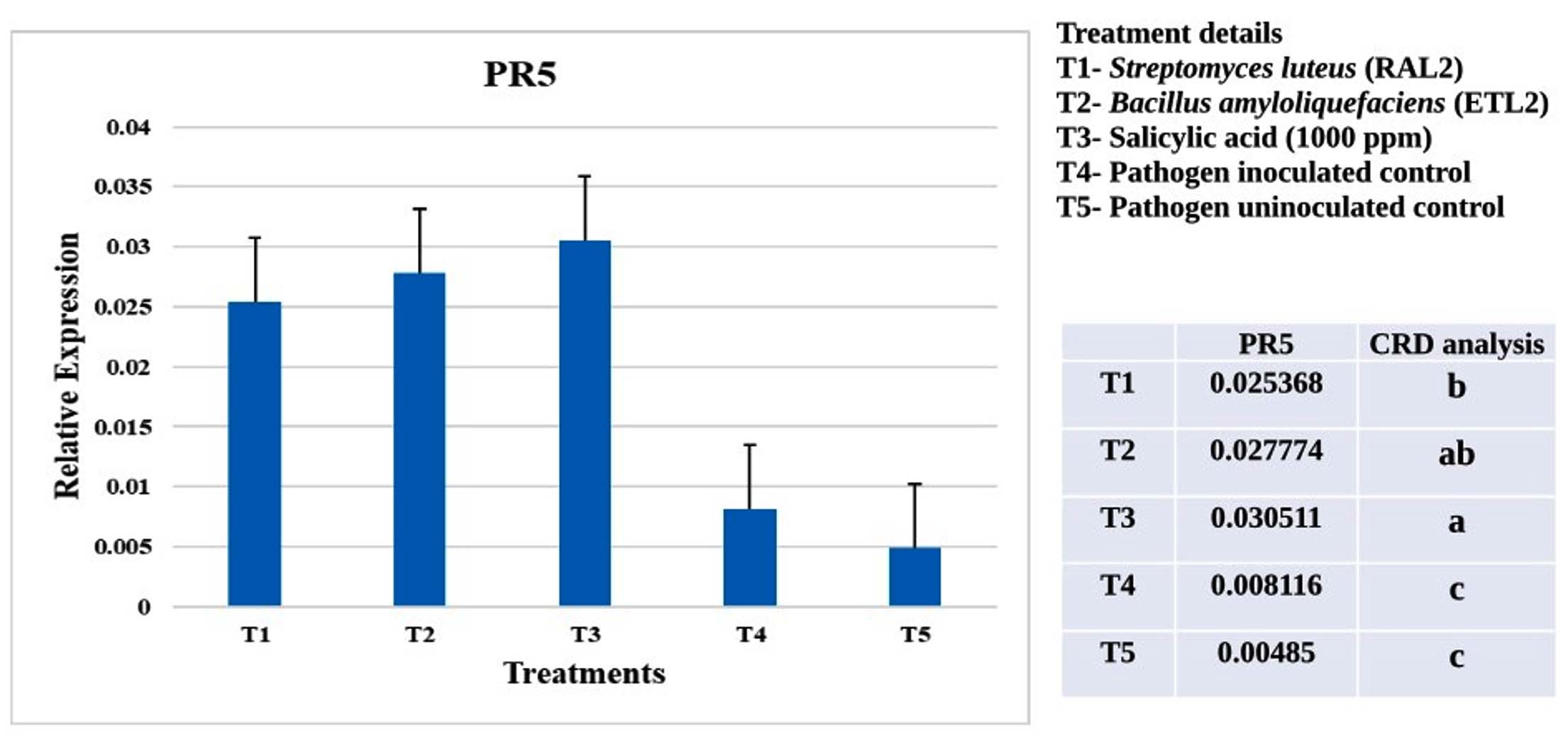
Figure 3. Expression analysis of defense gene PR5 in cotton seedlings (LRA5166) treated with biocontrol agents and chemical inducer challenge inoculated with X. citri pv. malvacearum (MNSu) using qRT-PCR. Polyubiquitin gene was used as internal control. Statistical analyses were performed using R software. Differences in gene expression among treatments were assessed by one-way analysis of variance (ANOVA), followed by Tukey’s honestly significant difference (HSD) test. Data is presented as fold expression level. Error bars indicated standard error obtained from three replicates per treatment. Means followed by different letters are significantly different for gene expression at P < 0.05
This study aimed to identify five different effector genes in 34 Xcm isolates collected from key cotton-growing regions across India. Previous researchers also studied the diversity of effectors in the cotton bacterial blight pathogen. Zhai et al.21 identified 181 and 178 potential virulence-related effectors through whole-genome sequencing of the highly virulent strain GSPB2388 of X. citri pv. malvacearum from Sudan and race 18 of GSPB1386 from Nicaragua, including type I to IV secretion systems. Type III secretion system effectors, called Xanthomonas outer proteins (Xops), are essential for bacterial growth and colonisation in various eukaryotic hosts.22 Genes responsible for producing effector proteins in plant pathogens are functionally identified as avirulence (avr) genes.23 In the present study, a maximum of thirteen isolates, including the positive control (PC-CICR), recorded positive for the avrBs3 gene. Of 10 Maharashtra isolates, 7 (70%) were positive for the avrBs3 gene. The result was consistent with Park et al.,23 who studied the diversity of the avrBs3 avirulence gene in 155 strains of X. axonopodis pv. glycines. They found that avrBs3 was the primary pathogenic factor in soybean. The avrBs3 mutant strain, or strains lacking avrBs3, lost pathogenicity or induced only a few pustules on highly susceptible cultivars. The avrBs3 gene belongs to the transcription activator-like (TAL) effector family, also called the AvrBs3/PthA family, which is found in most Xanthomonas strain genomes.18 Mokryakova et al.24 also studied effector gene diversity in 53 strains of the genus Xanthomonas, including eight completely sequenced genomes representing four different species, using 19 effector gene-specific primers. They found that xopD was the most frequent gene (85% of strains) and indicated its importance for pathogenesis. 30% of the strains carried the avrBs3 group. They also examined how the content and number of effector genes relate to different disease symptom types and the main pathogen race.
The Type III secretion system effector protein gene xopK was detected in eight isolates, and the HopG1 gene in seven isolates. Likewise, Schwartz et al.25 identified XopE2 in all field strains of Xanthomonas perforans and considered it a commonly shared effector among Xanthomonas euvesicatoria and Xanthomonas gardneri strains. They also detected AvrBsT in most X. perforans field strains and an identical version of X. euvesicatoria XopJ1 in X. gardneri field strains. This suggests a more broadly defined YopJ family within the shared effector list. Xanthomonas axonopodis pv. punicae contains six Xop-effectors, including XopC2, XopE1, XopL, XopN, XopQ and XopZ.22 The current study results showed that the genes xopAD and xopI recorded the minimum number of isolates, namely six and five, respectively. Generally, the functions of each effector differ and play a crucial role in symptom expression in cotton plants. The AvrBs3 activates host susceptibility genes, thereby inducing hypertrophy and water-soaking symptoms in plants. XopK effector prevents stomatal closure through suppressing ABA/MeJA signal suppression. The variable effector XopAD may be involved in host identification. XopI aids the pathogen’s virulence, while HopG1 suppresses plant defence responses.
The highly virulent and race 18 MNSn isolate had the most effector genes, with a maximum of 4, followed by MAB and KDR, each with 3. The positive control (PC-CICR) recorded with two genes, namely avrBs3 and xopAD. Nine of 10 isolates from Maharashtra carried at least one effector gene. Most of the isolates from the central zone were highly virulent and belonged to race 18. The south zone isolates recorded fewer effector genes, which are moderately to less virulent. The analysis of draft genomes of thirteen X. citri pv. malvacearum isolates, along with four previously published genomes, revealed the presence of 24 conserved and 9 variable type three effectors.2 They also found that race 18 contains 3-5 more effectors than other strains. Ochiai et al.6 noted that a variety of effector genes and mobile elements contribute to the significant race differentiation observed in Xanthomonas strains. The AvrBs3 effector is used as a marker for the identification of Xanthomonas strains. Further study of effectors will help understand how Xanthomonas manipulates host cells and develop new pathogen-resistance genes in plants and disease management strategies.
In the present study, the effective biocontrol agents, namely Streptomyces mutabilis (RAL2) and Bacillus amyloliquefaciens (ETL2), treated plants showed increased expression of defence genes, such as PR1 and PR5, in cotton seedlings challenged with X. citri pv. malvacearum in cotton compared to the control. Both the biocontrol agents did not show any statistically different gene expression for both the genes. PR1 and PR5 genes are the key pathogenesis-related genes in cotton, signalling the salicylic acid (SA) pathway during plant defence against pathogens and stress. Several workers have observed the activation of defence genes in crop plants when using biocontrol agents and chemicals to protect against bacterial and fungal pathogens. Gupta et al.26 described how the B. subtilis strain FZB-G activates defence-related compounds in tomato plants. In the present study, salicylic acid-treated plants showed higher expression of defence genes, indicating the effectiveness of chemical inducers. In pot and field experiments, the biocontrol agents significantly reduced the incidence of bacterial blight and promoted plant growth and yield. This shows that chemical inducers can rapidly activate the plant defence system. Biocontrol agents have the added advantage of promoting plant growth compared to chemical inducers. Similarly, Marmey et al.27 found that the hypersensitive reaction (HR) of cotton in response to inoculation of X. citri pv. malvacearum race 18 was associated with 9S-lipoxygenase activity (LOX) responsible for lipid peroxidation. GhLOX1 was highly expressed during pathogen-induced HR. Sequence analysis showed that GhLOX1 is a putative 9-LOX, and the GhLOX1 promoter contains SA- and JA-responsive elements, suggesting that GhLOX1 encodes 9-LOX activity and is involved in cell death during cotton HR. Zambounis et al.19 examined the relative expression levels of two pathogenesis-related (PR) genes (PR-3 and PR-10) and a detoxification gene (GST18) in fully susceptible (Lacta) and partially field-resistant (Emerald) cotton cultivars challenged with an Australian isolate of Fusarium oxysporum f. sp. oxysporum. Chemical inducers, such as BION® (a chemical analogue of salicylic acid) and methyl jasmonate (MeJA), were applied as a pretreatment before pathogen inoculation. They found that all PR genes were overexpressed in both hypocotyls and roots after pathogen inoculation in the Emerald cultivar, but were not affected in cultivar Lacta. Likewise, Khan et al.28 investigated how G. hirsutum expresses various pathogenesis-related protein genes, including PR1, PR4, PR5, β-1,3-glucanase, and chitinase, in response to cotton leaf curl virus disease. They found a significant interaction between the virus coat protein gene and the PR1, PR5, and Chitinase genes in plants. They concluded that the expression of PR1, PR5, and Chitinase genes can be exploited to confer resistance to leaf curl virus in cotton.
Of thirty-four X. citri pv. malvacearum isolates, 13 harboured the avrBs3 avirulence gene, as evidenced by 374 bp amplicons. The effector protein gene xopK (467 bp) was identified in eight isolates. Seven isolates showed amplification of the type III secretion system effector protein gene HopG1 (232 bp). Avirulence protein gene xopAD (954 bp) was recorded in six isolates. The gene xopI, coding for the type III secretion system effector protein (573 bp), was found in five isolates.
Plants treated with S. luteus (RAL2) showed numerically maximum expression of the defence gene PR1 compared to those treated with B. amyloliquefaciens (ETL2). The PR5 gene expression was numerically higher in B. amyloliquefaciens (ETL2) treated plants, followed by S. mutabilis (RAL2).
ACKNOWLEDGMENTS
The authors would like to thank the Director, ICAR-Central Institute for Cotton Research, Nagpur; the Professor and Head, Department of Plant Pathology; and the Dean (SPGS), Tamil Nadu Agricultural University, Coimbatore, for their support during this study. The authors also sincerely thank DST (FIST) and UGC (SAP) for providing laboratory facilities.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Jalloul A, Sayegh M, Champion A, Nicole M. Bacterial blight of cotton. Phytopathol Mediterr. 2015;54(1):3-20.
Crossref - Phillips AZ, Berry JC, Wilson MC, Vijayaraghavan A, Burke J. Genomics-enabled analysis of the emergent disease cotton bacterial blight. PLoS Genetics. 2017;13(9):e1007003.
Crossref - Sampath Kumar A, Eraivan AAK, Nakkeeran S, Manickam S. Current status and prevalence of cotton bacterial blight disease in India. J. Cotton Res. Dev. 2019;33:131-135.
- Buttner D, Bonas U. Regulation and Secretion of Xanthomonas Virulence Factors. FEMS Microbiol Rev. 2010;34(2):107-133.
Crossref - Alfano JR, Collmer A. The type III (Hrp) secretion pathway of plant pathogenic bacteria: trafficking harpins, Avr proteins, and death. J Bacteriol. 1997;179(18):5655-5662.
Crossref - Ochiai H, Inoue Y, Takeya M, Sasaki A, Kaku H. Genome sequence of Xanthomonas oryzae pv. oryzae suggests that large numbers of effector genes and insertion sequences contribute to its race diversity. Jpn Agric Res Q. 2005;39(4):275-287.
Crossref - Grant SR, Fisher EJ, Chang JH, Mole BM, Dangl JL. Subterfuge and manipulation:type III effector proteins of phytopathogenic bacteria. Annu Rev Microbiol. 2006;60(1):425-449.
Crossref - Sarkar SF, Gordon JS, Martin GB, Guttman DS. Comparative genomics of host-specific virulence in Pseudomonas syringae. Genetics. 2006;174(2):1041–1056.
Crossref - Thieme F, Koebnik R, Bekel T, et al. Insights into genome plasticity and pathogenicity of the plant pathogenic bacterium Xanthomonas campestris pv. vesicularia revealed by the complete genome sequence. J. Bacteriol. 2005;187(21):7254-7266.
Crossref - Kjemtrup S, Nimchuk Z, Dangl JL. Effector proteins of phytopathogenic bacteria:bifunctional signals in virulence and host recognition. Curr Opin Microbiol. 2000;3(1):73-78.
Crossref - Kay S, Bonas U. How Xanthomonas type III effectors manipulate the host plant. Curr Opin Microbiol. 2009;12(1):37-43.
Crossref - Chakrabarty PK, Duan YP, Gabriel DW. Cloning and Characterization of a Member of the Xanthomonas avr/pth Gene Family That Evades All Commercially Utilized Cotton R Genes in the United States. Phytopathol. 1997;87(11):1160-1167.
Crossref - Lee HJ, Park YJ, Seo PJ, et al. Systemic immunity requires SnRK2.8-mediated nuclear import of NPR1 in Arabidopsis. Plant Cell. 2015;27(12):3425-3438.
Crossref - Li ZT, Dhekney SA, Gray DJ. PR-1 gene family of grapevine: a uniquely duplicated PR-1 gene from a Vitis interspecific hybrid confers high-level resistance to bacterial disease in transgenic tobacco. Plant Cell Rep. 2011;30(1):1-11.
Crossref - Patil MA, Pierce ML, Phillips AL, Venters BJ, Essenberg M. Identification of genes unregulated in bacterial blight resistant upland cotton in response to inoculation with Xanthomonas campestris pv. malvacearum. Physiol Mol Plant Pathol. 2005;67(6):319-335.
Crossref - Chandrasekaran M, Chun SC. Expression of PR protein genes and induction of defense-related enzymes by Bacillus subtilis CBR05 in tomato (Solanum lycopersicum) plants challenged with Erwinia carotovora subsp. carotovora. Biosci Biotechnol Biochem. 2016;80(11):2277-2283.
Crossref - Yim WJ, Kim KY, Lee YW, Sundaram SP, Lee Y, Sa TM. Real time expression of ACC oxidase and PR-protein genes mediated by Methylobacterium spp. in tomato plants challenged with Xanthomonas campestrispv. vesicatoria. J Plant Physiol. 2014;171(12):1064-1075.
Crossref - Hajri A, Pothier JF, Fischer-Le Saux M, et al. Type three effector gene distribution and sequence analysis provide new insights into the pathogenicity of plant-pathogenic Xanthomonas arboricola. Appl Environ Microbiol. 2012;78(2):371-384.
Crossref - Zambounis AG, Kalamaki MS, Tani EE, Paplomatas EJ, Tsaftaris AS. Expression analysis of defense-related genes in cotton (Gossypium hirsutum) after Fusarium oxysporum f. sp. vasinfectum infection and following chemical elicitation using a salicylic acid analog and methyl jasmonate. Plant Mol Biol Rep. 2012;30(1):225–234.
Crossref - Livak KJ, Schmittgen TD. Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2−ΔΔCTMethod. Methods. 2001;25(4):402-408.
Crossref - Zhai J, Xia Z, Liu W, Jiang X, Huang X. Genomic sequencing globally identifies functional genes and potential virulence-related effectors of Xanthomonas axonopodis pv. malvacearum. Eur J. Plant Pathol. 2013;136(4):657-663.
Crossref - Kumar R, Mondal KK. XopN-T3SS effector modulates in planta growth of Xanthomonas axonopodis pv. punicae and cell-wall-associated immune response to induce bacterial blight in pomegranate. Physiol Mol Plant Pathol. 2013;84(1):36-43.
Crossref - Park HJ, Han SW, Oh C, et al. Avirulence gene diversity of Xanthomonas axonopodis pv. glycines isolated in Korea. J Microbiol Biotechnol. 2008;18(9):1500-1509.
- Mokryakova MV, Abdeeva IA, Piruzyan ES, Schaad NW, Ignatov AN. Diversity of effector genes in plant pathogenic bacteria of genus Xanthomonas. Microbiol. 2010;79(1):58-65.
Crossref - Schwartz AR, Potnis N, Timilsina S, et al. Phylogenomics of Xanthomonas field strains infecting pepper and tomato reveals diversity in effector repertoires and identifies determinants of host specificity. [Commentary]. Front Microbiol. 2015;6:535.
Crossref - Gupta VP, Bochow H, Dolej S, Fischer I. Plant growth-promoting Bacillus subtilis strain as potential inducer of systemic resistance in tomato against Fusarium wilt. J Plant Dis Prot. 2000;107(2):145-154.
- Marmey P, Jalloul A, Alhamdia M, et al. The 9-lipoxygenase GhLOX1 gene is associated with the hypersensitive reaction of cotton Gossypium hirsutum to Xanthomonas campestris pv malvacearum. Plant Physiol. Biochem. 2007;45(8):596-606.
Crossref - Khan MF, Naqvi SAH, Iqbal A, et al. Quantitative analysis of pathogenesis-related protein expression in Gossypium hirsutum L. to elicitor-induced resistance against cotton leaf curl disease and predicted in-silico protein-protein interactions. Physiol Mol Plant Pathol. 2025;137:102611.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.