


ISSN: 0973-7510
E-ISSN: 2581-690X
The Almanagil province located in Gezira scheme, Gezira state, Sudan, represents a suitable environment for the breeding of malaria-carrying mosquitoes. An estimated 5.9% of Sudanese people suffer from malaria, with 87.6% of cases caused by Plasmodium falciparum and 12.4% by Plasmodium vivax. Clinical manifestation of malaria cases range from mild uncomplicated to severe and fatal complications and the genetic variants and multiplicity of falciparum infection can worsen the manifestations of malaria. The objective of this work is to determine the degree of genetic variation in P. falciparum infection in a high-transmission region of central Sudan by analyzing merozoite surface protein-1 (msp1) and merozoite surface protein-2 (msp2) variations. During the rainy season of 2022, Eighty-nine children with confirmed severe falciparum malaria whom admitted to Almanagil Pediatric Hospital were included in this study. Dry blood spots were used to extract the DNA and amplification of three msp1 and two of msp2 allelic subfamilies, namely K1, RO33 and MAD20 and FC27 and IC/3D7, respectively. The data was analyzed by using SPSS computer program (v 23.0). The three genetic subfamilies of msp1 (K1, RO33 and MAD20) and the two alleles of msp2 (FC27 and IC/3D7) were identified. Msp1 variants represent K1 (64/89, 71.9%), RO33 (56/89, 62.9%) and MAD20 (72/89, 80.9%), while msp2 diversity represents ICI/3D7 (52/89, 58.4%), FC27 (62/89, 69.6%) and ICI/3D7/FC27(33/89, 37.1%). The MAD20 and FC27 showed high genetic diversity among both genes respectively. RO33 allele shows a strong association with severity of falciparum malaria (OR 2.572, P 0.045 ), while the K1 was the lowest risk factor for malaria severity. The allele subfamily K1 and MAD20 of msp1 were associated with hypoglycemia (OR 4.21 and 2.91) respectively. Our study revealed high genetic polymorphisms of msp1 and msp2. Among Central Sudanese children with high MOI of P. falciparum isolates, there was a significant frequency of msp1, a strong association between the K1 allele and hypoglycemia, and a substantial association between the RO33 and MAD20 alleles with the severity of the infection. These findings could help develop malaria control strategies.
Plasmodium falciparum, Malaria, MSP-1, MSP2, Children, Sudan
The human malaria parasite Plasmodium falciparum is a single-celled eukaryotic organism. Clinical signs of Falciparum malaria range from asymptomatic parasitemia to fever, headache, exhaustion, abdominal discomfort, vomiting, and diarrhea in the case of uncomplicated disease to coma and death in the case of complicated disease. Malaria burden is high, in some countries, worldwide, which causes high morbidity and mortality, approximately two hundred forty nine million cases and about 93.6% of cases and 95.4% of deaths globally; 78.1% of all deaths in African region were among children aged under 5 years in 2022 according to the WHO.1
The overall malaria prevalence in Sudan is 5.9%, with 87.6% attributable to P. falciparum and 5% attributable to a mixed infection with P. vivax.2 A knowledge of the genetic structure of the malaria parasite population is necessary for determining its dynamics, diversity, distribution, and epidemiology3,4 the identification of genetic diversity can help in controlling and elimination of malaria. Factors in the host, the parasite, and the environment all contribute to the severity of a malaria infection, the outcome of malaria treatment and recovery may affected by the variations in parasites like merozoite surface protein polymorphism (MSP). In many geographical contexts, P. falciparum displays highly variable genetic diversity.5 The virulence and complex diversity of P. falciparum may interpret its ability to exhibit with a variety of clinical symptoms and signs. Malaria is made worse by the production of several chemokines and lymphokines, which may be affected by the multiplicity of infections.6 At the surface of the Plasmodium merozoite, the MSP1 is the most important protein.5,7 For Plasmodium blood stages, MSP-1 is necessary8 and has an important role in erythrocyte invasion,9 particularly at the merozoite’s first contact with RBCs,10,11 In addition to red blood cell rupture and parasite egress.12 In vitro investigations have shown that antibodies to MSP-1 may reduce parasite development,9,13 and other epidemiological studies have linked these antibodies to protection against malaria.14,15,16 The diversity of protein on the merozoite surface P. falciparum may affect the transmission and immune evasion.17 Many merozoite surface proteins of P. falciparum linked to virulence and parasite infection severity have been identified.18,19 Vaccines have been developed targeting the proteins that are produced by the P. parasite throughout its pre-erthrocytic blood stages. The MSP-1 gene spans 17 sequence blocks on chromosome 9; of these, block 2 is the most polymorphic, splitting into the MAD20, K1, and RO3320 allelic groups. The gene for the msp2 glycoprotein is located on chromosome 2. The protein has five distinct regions, the most pleomorphic of which is the middle region (block 3), and it has a molecular weight of about 30 kDa. There are two groups of msp2 alleles: 3D7/IC1 and FC27. The dynamics of malaria transmission and its virulence were predicted using the msp2 subfamily.17
The multiplicity of P. falciparum infection (MOI) is a challenge in treating the malaria infection extremely in area with high transmission season, which can aggravate resistant strain to antimalarial chemotherapy.21,22 Microscopic analysis of thick and thin blood films is the “gold standard” method for parasite diagnosis and therapy recommendations.23 There is a lack of information on the msp1 and msp2 genetic variants in P. falciparum that circulate in areas like Gezira state in central Sudan, even though malaria is very prevalent in that country. This study details the frequency and variety of the msp1 and msp2 genes in P. falciparum parasites found in P. falciparum positive patients in Almanagil, Sudan. It also examines the relationship between these alleles and the severity of sickness and the number of infections. These findings could help in treating, elimination and controlling malaria in our study area.
Study area and population
This was a hospital based cross-sectional study conducted in Almanagil, Gezira State, Central Sudan, between September and December 2022. The city located south of Khartoum about 156 kilometers in Gezira Agricultural Scheme’s irrigated region, where a highly variable mesoendemic to hyperendemic malaria transmission pattern exists. Malaria is endemic in this region all year long, with P. falciparum being the most common kind and P. vivax being the second. Children who were treated at Almanagil Pediatric Teaching Hospital for malaria and meeting the inclusion and exclusion criteria were recruited for this study. Blood films (thin and thick) were stained with 10% Giemsa’s at pH 7.2 and then examined microscopically by expert technician for the presence of malaria parasites; 200 fields under 100 x magnification was examined from the thick film before the slide was considered negative. The parasitemia was determined by counting only the a sexual stages against 200 white blood cells (WBC) and the number of parasites multiplied by 8000 then divided by 200, assuming the average of total WBC count of individuals equal to 8000 cells.12 When microscopic examination revealed P. falciparum mono-infection in any child younger than 18 years old, regardless of gender, were included in this study. After explaining the objectives of the study, how to collect the sample, and the minor harm that could occur, the verbal consent was taken from one of the parents, and then the patients were included in the study. The demographic, clinical and laboratory results were collected by using a well-structured questionnaire.
Sample collection
Under aseptic conditions, containers containing EDTA were used to collect 3 ml of venous blood from each participant, for 89 samples. Prior to storage in appropriate sealed containers with desiccant, blood samples were spotted onto filter paper (Whatmann® No. 3, Sigma-Aldrich, Germany), allowed to air dry, and then preserved.24 Blood films were stained by Giemsa and parasite density was quantified according to Monica-cheesbrough as mentioned above.
DNA extraction
Small pieces of filter paper with dried blood spots (DBSs) on them were sniped off using scissors and placed into 1.5 mL microtubes. After adding about 500 µL of dH2O to the test tubes, they were allowed to sit at room temperature for a time of 5 minutes. We threw out the dH2O and did it twice more. Five hundred µL of extraction buffer (PBS or 10 mM Tris-EDTA (TE)) was added to filter paper. The filter sheets were treated with 100 µL of red blood cell lysis solution, then 10 µL of the same buffer was mixed with 1 mg/mL of proteinase K, and the combination was left to incubate at 37°C overnight. A clean pipette tip was used to repeatedly push the filter papers against the tube bottom. Cell lysates were heated to 95 degrees Celsius for 15 minutes to maximize extraction. The filter sheets were discarded after a short centrifugation (only 2-3 seconds) and the DNA-rich supernatant was frozen at 80°C for further analysis.25,26
Electrophoresis procedure for msp1 gene subfamilies
The primers sequences as shown in Table 1, were used for the amplification of msp1 of P. falciparum and subfamilies (Table 2 and 3).
Table (1):
primers sequences using to amplify Pfmsp1
| PCR cycle | The Primer | The sequence |
|---|---|---|
| First PCR msp1 Second PCR | M1-OF | F:5- -CTAGAAGCTTTAGAAGATGCAGTATTG-3- |
| M1-OR | R:5- -CTTAAATAGTATTCTAATTCAAGTGGATCA-3- | |
| M1-KF | 5- -AAATGAAGAAGAAATTACTACAAAAGGTGC-3- | |
| M1-KR | 5- -GCTTGCATCAGCTGGAGGGCTTGCACCAGA-3- | |
| M1-MF | 5- -AAATGAAGGAACAAGTGGAACAGCTGTTAC-3- | |
| M1-MR | 5- -ATCTGAAGGATTTGTACGTCTTGAATTACC-3- | |
| M1-RF | 5- -TAAAGGATGGAGCAAATACTCAAGTTGTTG-3- | |
| M1-RR | 5- -CATCTGAAGGATTTGCAGCACCTGGAGATC-3- |
The PCR cycle condition performed as described by Ananias et al. The product size was 1200-1400 bp27
Table (2):
The PCR conditions used to amplify msp1 of P. falciparum
| msp1 outer | X1 | Temperature | |
|---|---|---|---|
| H2O | 9.0 | 95°C/5 min | 35 Cycles |
| Mix | 4.0 | 94°C /30 sec | |
| msp1-O1 | 1.0 | 55°C/30 sec | |
| msp1-O2 | 1.0 | 72°C/1.0 min | |
| DNA | 5.0 | 72°C/5 min |
Table (3):
Amplification of Pfmsp1 gene subfamily
| msp1 nested | X1 | nested Pfmsp1 (K1, RO33, and MAD 20) | |
|---|---|---|---|
| H20 | 12.0 | 95°C/5 min. | 35 Cycles |
| Mix | 4.0 | 94°C /30 sec. | |
| MSP1-O1 | 1.0 | 56°C/30 sec. | |
| MSP1-O2 | 1.0 | 72°C/1.5 min. | |
| DNA | 2.0 | 72°C/5 min. |
The amplified products were mixed thoroughly with loading buffer, which included Bromophenol blue 0.25%, ficoll 25%, 10 mM Tres, and 1 mM EDTA. Five microliters of each product were then added to a 2% agarose gel. The gel was electrophoresed using a buffer containing Tris-Borate-EDTA, which contained 0.09M boric acid, 0.09M Tris, and 0.002M EDTA (Figure 1). The DNA fragments were stained with ethidium bromide. The marker of DNA (100 pb ladder) was used to identify the fragments size (Boehringer Mannheim, U.K). The condition of electrophoresis gel as follows: run for 10-120 minutes in buffer and 120 volts. The products size was visualized by the photo documentation system. R fragment sizes were estimated according to migration (Amersham Pharmacia Biotech).
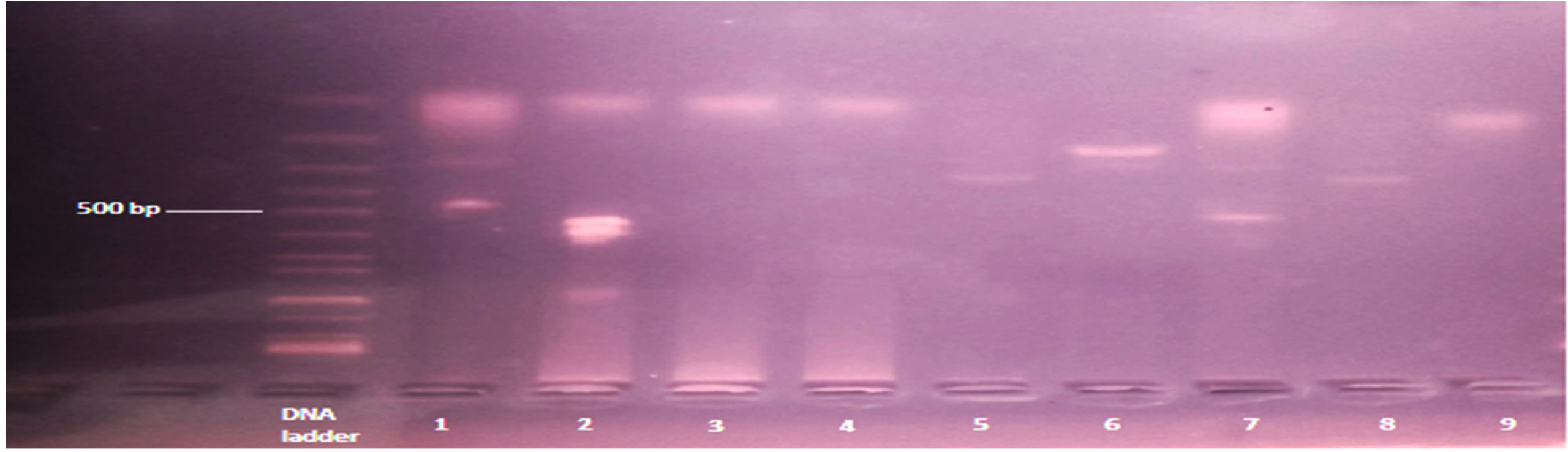
Figure 1. Shows RO33 allele of msp1: The band size ranged between 100-500 bp. numbers 1 (size 100/250 bp), 2 (size 100/500 bp) and 7 (size 100/500 bp) are positive with multiple clones, while the remaining was monoclonal RO33 allele
msp2 amplification and genotyping
The genus and species-specific primers were used to identify the species of Plasmodium by using 18S rRNA gene based nested PCR as described by Atroosh et al.28 The nested PCR was used to amplify the polymorphic area of Pfmsp2 (block 3) and subfamily of alleles (FC27 and IC1/3D7). The PCR conditions and reactions were as follows: 1. Included in the 25 µl reaction volume are 2 µl of falciparum DNA, 0.2 mM of dNTPs, 2 mM of MgCl2, 1 µl of each primer, and 1 unit of Taq Polymerase (Vivantis, Selangor Darul Ehsan, Malaysia). Second, the outer PCR needs the following conditions: three minutes of denaturation at 94°C, thirty seconds of annealing at 55°C, and two minutes of extension at 72°C. In a thermal cycler (SensoQuest, Gottingen, Germany), the most recent cycle extension was at 72°C for 10 minutes (Figure 2 and 3).
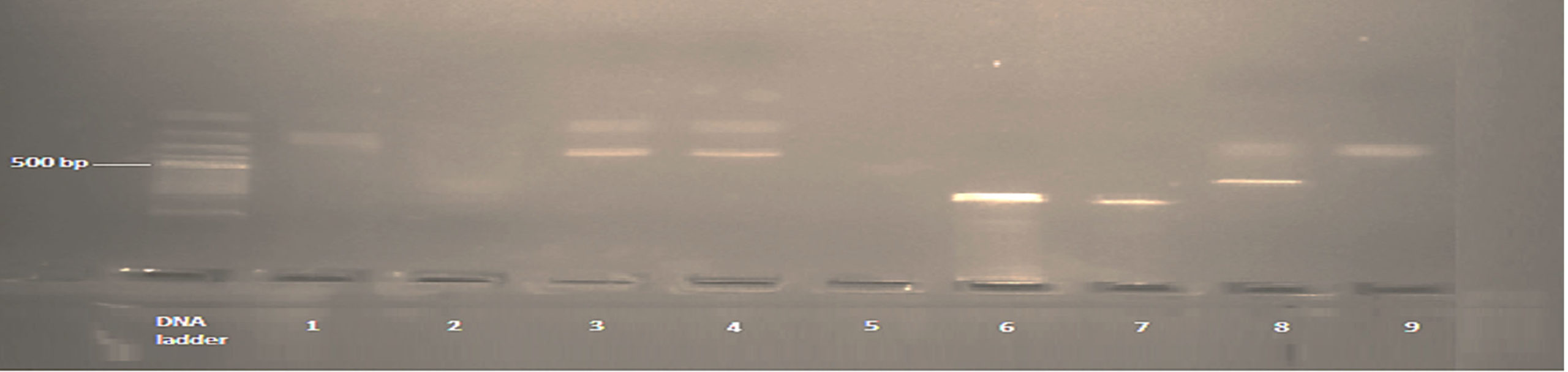
Figure 2. Shows K1 allele of msp1. The band size ranged between 100-650 bp. Numbers 3 (fragment size 300/500 bp), 4 (fragment size 300/500 bp) and 8 (fragment size 600/500 bp) are positive K1 allele with multiple clones, while the remaining were positive
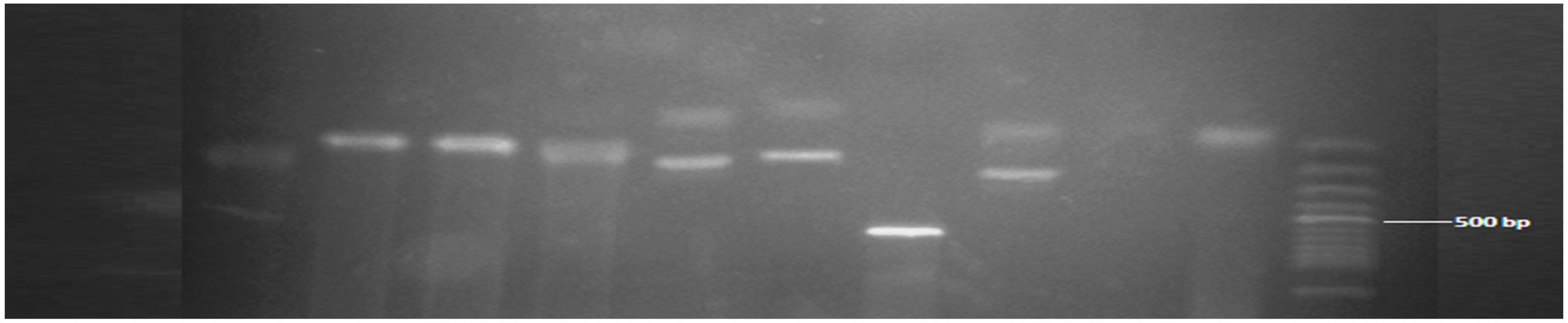
Figure 3. Showed MAD20 allele of msp1. The band size ranged between 90-600 bp. Numbers 5 (fragment size 90/100 bp), 6 (fragment size 90/100 bp), 8 (fragment size 100/300 bp) were positive with multiple clones, while the remaining were positive monoclonal MAD20 all
Detection of FC27and IC1/3D7 alleles
As mentioned before, in the electrophoresis technique for the msp1 gene subfamily, the amplified PCR products were subjected to electrical analysis. The method for determining the multiplicity of infection (MOI) was outlined by Abukari et al.29
Data analysis
The data was analyzed by using SPSS program (version 20.0). Allelic distribution of both genes was counted as described by Abukari et al. 29 There was an application of the 95% confidence interval and degree of certainty.
Ethical statement
In Gezira State, Sudan, our research was authorized by the Scientific and Research Ethics Committee. The patient’s parent or legal guardian gave their informed permission. Throughout, we made sure that patient records remained secret.
Out of 89 children with falciparum malaria participated in this study. (48/89, 53.9%) were male and (41/89, 46.1%) were female. The common age group was less than 5 years representative (47/89, 52.8%) (Table 4).
Table (4):
Characteristics of study participants
Frequency |
Percent (%) |
||
|---|---|---|---|
Age ranges |
47 |
52.8 |
|
≥5 to ≤12 years |
34 |
38.2 |
|
>12 to ≤18 years |
8 |
8.9 |
|
Total |
89 |
100 |
|
Gender |
Male |
48 |
53.9 |
Female |
41 |
46.1 |
|
Total |
89 |
100 |
The results of genetic diversity among the isolates of falciparum malaria showed the MAD20 was (72/89, 80.9%), K1 (64/89, 71.9%) and RO33 (56/89, 62.9%) of msp1 and FC27 (62/89, 69.97%) and ICI/3D7 52/89, 58.4%) of msp2. The MAD20 and FC27 showed high genetic diversity among both msp1 and msp2 alleles, respectively, as shown in Table 5.
Table (5):
The frequency of the msp1 and msp2 gene alleles among study subjects
| Alleles | Status | Frequency | Percent (%) |
|---|---|---|---|
| K1 | + | 64 | 71.9 |
| – | 25 | 28.1 | |
| MAD20 | + | 72 | 80.9 |
| – | 17 | 19.1 | |
| RO33 | + | 56 | 62.9 |
| – | 33 | 37.1 | |
| FC27 | + | 62 | 69.7 |
| – | 27 | 30.3 | |
| ICI/3D7 | + | 52 | 58.4 |
| – | 37 | 41.6 |
Our results revealed that the multiplicity of K1+RO33, K1+MAD20 and MAD20 of msp1 were 7.8%, 6.7% and 5.6% respectively, while the multiplicity of ICI/3D7+FC27 was 37.1% of msp2 as shown in Table 6.
Table (6):
The Pfmsp1 and Pfmsp2 multiplicity of P. falciparum infection
89i |
Frequency |
Percent |
|---|---|---|
K1+RO33 |
7 |
7.8 |
K1+MAD20 |
6 |
6.7 |
MAD20+RO33 |
5 |
5.6 |
MOI of msp1 |
1.35 |
|
IC1/3D7/FC27 |
33 |
37.1 |
Multiclonal isolates of IC1/3D7/FC27 |
14 |
15.7 |
Mix infections IC1/3D7/FC27 |
35 |
39.3 |
MOI of msp2 |
1.48 |
The result showed that the percentage of msp1 alleles among patients with severe malaria were (24/29, 83%), (22/29, 76%), and (21/29, 72%) for MAD20, RO33 and K1, respectively, while the correlation between the presence of the MAD20 followed by RO33 alleles and the severity of illness P = 0.037, Odd ratio 2.900 and P=0.045; Odds ratio = 2.572 respectively. Also, the percentage of msp2 alleles among severe malaria cases was 72% for FC27 and 59% for 3D7, when compared between both types of malaria infection Table 7.
Table (7):
Shows the association between msp1 and msp2 allelic subfamilies and severity of falciparum infection
| Allelic subfamilies of msp1 gene | Severity of infection | Total | P. value | Odd ratio | ||||
|---|---|---|---|---|---|---|---|---|
| Severe | Mild | |||||||
| Frequency | % | Frequency | % | |||||
| RO33 | + | 22 | 76 | 33 | 55 | 55 | 0.045 | 2.572 |
| – | 7 | 24 | 27 | 45 | 34 | |||
| Total | 29 | 100 | 60 | 100 | 89 | |||
| K1 | + | 21 | 72 | 44 | 73 | 65 | 0.559 | 0.955 |
| – | 8 | 28 | 16 | 27 | 24 | |||
| Total | 29 | 100 | 60 | 100 | 89 | |||
| MAD20 | + | 24 | 83 | 48 | 80 | 72 | 0.037 | 2.900 |
| – | 5 | 17 | 12 | 20 | 17 | |||
| Total | 29 | 100 | 60 | 100 | 89 | |||
| FC27 | + | 21 | 72 | 41 | 68 | 62 | 0.835 | 0.977 |
| – | 8 | 28 | 19 | 32 | 27 | |||
| Total | 29 | 100 | 60 | 100 | 89 | |||
| ICI/3D7 | + | 17 | 59 | 35 | 58 | 52 | 0.642 | 0.819 |
| – | 12 | 41 | 25 | 41 | 37 | |||
| Total | 29 | 100 | 60 | 100 | 89 | |||
Patients with severe malaria showed high risk for hypoglycemia with (Odd ratio 4.2 and 2.91) for K1 and MAD20 alleles of msp1, respectively, while severe anemia was slightly liked to msp1-K1 allele as shown in Table 8.
Table (8):
Shows the association between msp1 allelic subfamilies and severe anemia and hypoglycemia
| Complications | Present | Absent | P. Value | Odd ratio | |||
|---|---|---|---|---|---|---|---|
| Count | % | Count | % | ||||
| K1 allele | |||||||
| Severe Anemia | Yes | 17 | 26.2 | 5 | 20.8 | 0.414 | 1.40 |
| No | 48 | 73.8 | 19 | 79.2 | |||
| Total | 65 | 100.0 | 24 | 100.0 | |||
| Hypoglycemia | Yes | 10 | 15.4 | 1 | 4.2 | 0.142 | 4.21 |
| No | 55 | 84.6 | 23 | 95.8 | |||
| Total | 65 | 100.0 | 24 | 100.0 | |||
| MAD20 allele | |||||||
| Severe Anemia | Yes | 4 | 24 | 18 | 25 | 0.586 | 0.923 |
| No | 13 | 76 | 54 | 75 | |||
| Total | 17 | 100 | 72 | 100 | |||
| Hypoglycemia | Yes | 4 | 24 | 7 | 10 | 0.128 | 2.91 |
| No | 13 | 76 | 65 | 90 | |||
| Total | 17 | 100 | 72 | 100 | |||
| RO33 allele | |||||||
| Severe Anemia | Yes | 12 | 22 | 10 | 29 | 0.228 | 0.670 |
| No | 43 | 78 | 24 | 71 | |||
| Total | 55 | 100 | 34 | 100 | |||
| Hypoglycemia | Yes | 5 | 9 | 6 | 18 | 0.194 | 0.467 |
| No | 50 | 91 | 28 | 82 | |||
| Total | 55 | 100 | 34 | 100 | |||
Among patients carrying FC27 and ICI/3D7 alleles of msp2, showed no association between genetic diversity of msp2 and severity of malaria infection (Odd ratio 0.777 and 0.572). Also, there was no association between FC27 and ICI/3D7 alleles of msp2 and hypoglycemia (Odd ratio 0.761 and 0.465), respectively, as shown in Table 9.
Table (9):
The association between msp2 allelic subfamilies and severe anemia and hypoglycemia
| Complications | Present | Absent | P. Value | Odd ratio | |||
|---|---|---|---|---|---|---|---|
| Count | % | Count | % | ||||
| FC27 allele | |||||||
| Severe Anemia | Yes | 14 | 22.5 | 6 | 22.2 | 0.575 | 0.777 |
| No | 48 | 77.5 | 21 | 77.8 | |||
| Total | 62 | 100.0 | 27 | 100.0 | |||
| Hypoglycemia | Yes | 7 | 11.3 | 5 | 18.5 | 0.139 | 0.761 |
| No | 55 | 88.7 | 22 | 81.5 | |||
| Total | 62 | 100.0 | 27 | 100.0 | |||
| 3D7 allele | |||||||
| Severe Anemia | Yes | 11 | 21.2 | 4 | 10.8 | 0.438 | 0.572 |
| No | 41 | 78.8 | 33 | 89.2 | |||
| Total | 52 | 100 | 37 | 100 | |||
| Hypoglycemia | Yes | 4 | 9 | 6 | 18 | 0.394 | 0.465 |
| No | 48 | 91 | 28 | 82 | |||
| Total | 52 | 100 | 37 | 100 | |||
Human erythrocyte invasion by P. falciparum is likely facilitated by the merozoite surface protein 1 and 2 (MSP1 and MSP2). Examining the polymorphism and the fluctuating variety of this crucial gene in malaria-endemic locations could shed light on the disease’s toxicity, epidemiology, and diagnosis.30
The MSP1 gene is located on chromosome 9 and encodes the major surface protein 1 (MSP1) of P. falciparum. This protein is composed of 17 blocks of sequences that are separated by conserved regions.17 It is often targeted by the immune system and plays a crucial role in erythrocyte invasion.17,31 The MSP2 gene is located on chromosome 2, and it encodes merozoite surface protein 2, the second most abundant Glycophosphatidylinositol (GPI) anchored merozoite surface protein. The gene is composed of five blocks, with the greatest polymorphism occurring in the middle block, block 3.32 It is present in two allelic subfamilies namely (3D7-like and FC27-like).33
In our research region, we found the allelic subfamilies of msp1 (K1, RO33 and MAD20) and msp2 (FC27 and 3D7). Some falciparum malaria isolates have shown a tendency toward predominance for msp1 and msp2, respectively; they include MAD20 and K1 of msp1 and FC27 of msp2. This conclusion was consistent with research conducted in the Republic of the Congo (Mayengue et al), but not with research conducted in Tanzania,17,34 Thailand35 and Central Africa in Sudan.31
The study region yielded variants of Pfmsp2 alleles, including FC27 (62/89, 69.7%) and ICI/3D7 (52/89, 58.4%), with 33 out of 89 samples (37.1%) testing positive for both FC27 and IC1/3D7. Our study area had a higher prevalence of the FC27 allelic family compared to the IC1/3D7 allelic family. This finding aligns with previous research in White Nile State in Central Sudan, Gublak, Northwest Ethiopia, and Jazan, Saudi Arabia.36-38 On the other hand, studies in Northwest Ethiopia, Boset and Badewacho Districts, Southern Ethiopia, and Southwestern Nigeria39,40 have shown different results. These discrepancies could be attributed to differences in transmission dynamics, sample size, or study area.
The severity of malaria is considered as defined by WHO (WHO 2015), the result showed a significant correlation between the presence of the MAD20 followed by RO33 alleles and the severity of illness P = 0.037, Odd ratio 2.900 and P=0.045; Odds ratio = 2.572 respectively, when compared between severe and mild malaria cases. Researchers have shown that individuals with the RO33 allele had higher levels of tumor necrosis factor alpha, which is linked to the development of severe malaria (Robert et al 1996). The link between allelic backgrounds and illness severity was investigated. Samples positive for the RO33 allelic family of MSP1 among patients with mild malaria infection were represented (33/60, 55%) and severe malaria illness were (22/29, 76%). Positive samples for the K1 allelic family were found in (21/29, 72% and 44/60, 73%) with severe and mild malaria, respectively. The positivity of MAD20 allelic family is detected (24/29, 83%) of patients with severe malaria and (48/60, 80%) of patients with mild malaria. This contradicts recent research in Senegal that linked the K1 allelic family to severe malaria.38,41 The current study’s modest size, especially in the most severe cases of malaria, may explain this discrepancy. Consistent with previous research in Bobo-Dioulasso.42 Severe anemia and hypoglycemia are among the most common complications of malaria in our study area. The study revealed no link between MSP1 alleles and severe anemia in individuals carrying the K1 allele (p=0.414), MAD20 (P=0.586) and RO33 (p=0.228). Our report showed no association between hypoglycemia among patients carrying K1, MAD20 or RO33 alleles p. value 0.142, 0.128 and 0.194, respectively. K1 allele is both strongly associated with hypoglycemia and severe anemia, with odds ratios of 4.21, and 1.4, respectively. The result exhibited that the subjects carrying the K1 allele of msp1 had a greater risk of hypoglycemia than severe anemia. The RO33 allele was not associated with hypoglycemia (odd ratio 0.467) or severe anemia (odd ratio 0.670). This confirms what previous studies in Indonesia have shown.20 Our study found that 15.7% of samples had multi-clonal isolates, 39.3% had mixed infections, and the MOI was 1.48. This could be because of the high rates of malaria transmission and the frequency of multiple infections; people living in areas that typically harbor multiple parasite strains are likely to be affected. Consistent with previous research it indicates that P. falciparum infection diversity may vary with respect to geographic location, transmission intensity, and sample population.43
Our study revealed high genetic polymorphisms of msp1 and msp2. Among children in Central Sudan with high MOI of P. falciparum isolates, there was a substantial connection between the RO33 and MAD20 alleles and the severity of the illness, and a high frequency of msp1, K1 allele was related with hypoglycemia. Our findings highlight the need for constant monitoring of P. falciparum polymorphism in the study area and the findings could help in treating, eliminating and controlling malaria in our study area.
ACKNOWLEDGMENTS
The authors would like to thank the Almanagil Pediatric Teaching Hospital Laboratory team and all the malaria patients who consented to be investigated.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
This study was supported by the Ministry of Education and King Abdulaziz University, DSR, Jeddah, Saudi Arabia, under grant number GPIP: 1422-140-2024.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study was approved by the Institutional Ethics Committee of Planning and Development, Ministry of Health, Gezira State, Sudan (Reference No K1/T/44).
INFORMED CONSENT
Written informed consent was obtained from the participants before enrolling in the study.
- Letcher B, Maciuca S, Iqbal Z. Gene Conversion Drives Allelic Dimorphism in Two Paralogous Surface Antigens of the Malaria Parasite P. falciparum. bioRxiv. 2023;2023:02.27.530215.
Crossref - Mohamedahmed KA, Ahmed ZA, Nour BYM, Abakar AD, Babker AMa. Impact of Sever Plasmodium Falciparum Infection on Platelets Parameters among Sudanese Children Living in Al-Jazira State. Int J Clin Biomed Res. 2020;6:5-9.
Crossref - Arnott A, Barry AE, Reeder JC. Understanding the Population Genetics of Plasmodium Vivax Is Essential for Malaria Control and Elimination. Malar J. 2012;11:14.
Crossref - Talha AA, Pirahmadi S, Mehrizi AA, Djadid ND, Nour BYM, Zakeri S. Molecular Genetic Analysis of Plasmodium Vivax Isolates from Eastern and Central Sudan Using Pvcsp and Pvmsp-3a Genes as Molecular Markers. Infect Genet Evol. 2015;32:12-22.
Crossref - Le, HG, Kang JM, Jun H, et al. Changing Pattern of the Genetic Diversities of Plasmodium Falciparum Merozoite Surface Protein-1 and Merozoite Surface Protein-2 in Myanmar Isolates. Malar J. 2019;18(1):241.
Crossref - Hamid MMA, Elamin AF, Albsheer MMA, et al. Multiplicity of Infection and Genetic Diversity of Plasmodium Falciparum Isolates from Patients with Uncomplicated and Severe Malaria in Gezira State, Sudan. Parasit Vectors. 2016;9(1):362.
Crossref - Holder AA, Freeman RR. Immunization against Blood-Stage Rodent Malaria Using Purified Parasite Antigens. Nature. 1981;294(5839):361-364.
Crossref - O’Donnell RA, Saul A, Cowman AF, Crabb BS. Functional Conservation of the Malaria Vaccine Antigen MSP-119 across Distantly Related Plasmodium Species. Nat Med. 2000;6(1):91-95.
Crossref - Blackman MJ, Heidrich HG, Donachie S, McBride JS, Holder AA. A Single Fragment of a Malaria Merozoite Surface Protein Remains on the Parasite during Red Cell Invasion and Is the Target of Invasion-Inhibiting Antibodies. J Exp Med. 1990;172(1):379-382.
Crossref - Baldwin MR, Li X, Hanada T, Liu SC, Chishti AH. Merozoite Surface Protein 1 Recognition of Host Glycophorin a Mediates Malaria Parasite Invasion of Red Blood Cells. Blood. 2015;125(17):2704-2711.
Crossref - Boyle MJ, Richards JS, Gilson PR, Chai W, Beeson JG. Interactions with Heparin-like Molecules during Erythrocyte Invasion by Plasmodium Falciparum Merozoites. Blood. 2010;115(22):4559-4568.
Crossref - Chesbrough Monia. District Laboratory Practice in Tropical Countries Part 1, Second Edition United States of America by Cambridge University Press, New York. 2005.
- Woehlbier U, Epp C, Kauth CW, et al. Analysis of Antibodies Directed against Merozoite Surface Protein 1 of the Human Malaria Parasite Plasmodium Falciparum. Infect Immun. 2006;74(2):1313.
Crossref - Tolle R, Fruh K, Doumbo O, et al. A Prospective Study of the Association between the Human Humoral Immune Response to Plasmodium Falciparum Blood Stage Antigen Gp190 and Control of Malarial Infections. Infect Immun. 1993;61(1):40-47.
Crossref - Conway DJ, Cavanagh DR, Tanabe K, et al. A Principal Target of Human Immunity to Malaria Identified by Molecular Population Genetic and Im-munological Analyses. Nat Med. 2000;6(6):689-692.
Crossref - Lin CS, Uboldi AD, Epp C,et al. Multiple Plasmodium Falciparum Merozoite Surface Protein 1 Complexes Mediate Merozoite Binding to Human Erythrocytes. J Biol Chem. 2016;291(14):7703-7715.
Crossref - Marwa KJ, Lyimo E, Konje ET, Kapesa A, Kamugisha E, Swedberg G. Plasmodium Falciparum Merozoite Surface Proteins Polymorphisms and Treatment Outcomes among Patients with Uncomplicated Malaria in Mwanza, Tanzania. J Trop Med. 2022;2022:5089143.
Crossref - Jaschke A, Coulibaly B, Remarque EJ, Bujard H, Epp C. Merozoite Surface Protein 1 from Plasmodium Falciparum Is a Major Target of Opsonizing Antibodies in Individuals with Acquired Immunity against Malaria. Clin Vaccine Immunol. 2017;24(11):e00155-17.
Crossref - Hang JW, Tukijan F, Lee EQH, Abdeen SR, Aniweh Y, Malleret, B. Zoonotic Malaria: Non- Laverania Plasmodium Biology and Invasion Mechanisms. Pathogens. 2021;10(7):889.
Crossref - Jamil KF, Pratama NR, Marantina SS, et al. Allelic Diversity of Merozoite Surface Protein Genes (Msp1 and Msp2) and Clinical Manifestations of Plasmodium Falciparum Malaria Cases in Aceh, Indonesia. Malar J. 2021;20(1):182.
Crossref - Transmission Efficiency of Plasmodium Falciparum Malaria Resistant Strains in the African Mosquito Vectors | Request PDF Available online: https://www.researchgate.net/publication/355092444_Transmission_Efficiency_of_Plasmodium_falciparum_Malaria_Resistant_Strains_in_the_African_Mosquito_Vectors (accessed on 2 May 2024).
- Usman-Yamman H, Innocent-Omalu CJ, Abubakar A, Abolarinwa SO, Sameul ES, Otuu CA. Genetic Diversity of Plasmodium Falciparum Isolates in Minna, North Central Nigeria Inferred by PCR Genotyping of Merozoite Surface Protein 1 and 2. Infect Genet Evol. 2021;96:105143.
Crossref - Mbanefo A, Kumar N. Evaluation of Malaria Diagnostic Methods as a Key for Successful Control and Elimination Programs. Trop Med Infect Dis. 2020;5(5):102.
Crossref - Moyeh MN, Ali IM, Njimoh DL, et al. Comparison of the Accuracy of Four Malaria Diagnostic Methods in a High Transmission Setting in Coastal Cameroon. J Parasitol Res. 2019;2019:1417967.
Crossref - Kang JM, Cho PY, Moe M, Lee J, et al. Comparison of the Diagnostic Performance of Microscopic Examination with Nested Polymerase Chain Reaction for Optimum Malaria Diagnosis in Upper Myanmar. Malar J. 2017;16(1):119.
Crossref - Choi EH, Lee SK, Ihm C, Sohn YH. Rapid DNA Extraction from Dried Blood Spots on Filter Paper: Potential Applications in Biobanking. Osong Public Health Res Perspect. 2014;5(6):351-357.
Crossref - Ananias D, Rainho JP, Ferreira A, et al. Synthesis and Characterization of Er(III) and Y(III) Sodium Silicates: Na3ErSi3O9, a New Infrared Emitter. Chem Mater. 2002;14(4):1767-1772.
Crossref - Atroosh WM, Lau YL, Snounou G, Azzani M, Al-Mekhlafi HM. Plasmodium Falciparum Histidine Rich Protein 2 (Pfhrp2): An Additional Genetic Marker Suitable for Anti-Malarial Drug Efficacy Trials. Malar J. 2022;21(1):2.
Crossref - Abukari Z, Okonu R, Nyarko SB, et al. The Diversity, Mul-tiplicity of Infection and Population Structure of P. falciparum Parasites Circulating in Asymptomatic Carriers Living in High and Low Malaria Transmission Settings of Ghana. Genes. 2019;10(6):44.
Crossref - Kiwuwa MS, Ribacke U, Moll K, et al. Genetic diversity of Plasmodium falciparum infections in mild and severe malaria of children from Kampala, Uganda. Parasitol Res. 2013;112:1691-1700.
Crossref - Mohammed H, Mindaye T, Belayneh M, et al. Genetic Diversity of Plasmodium Falciparum Isolates Based on MSP-1 and MSP-2 Genes from Kolla-Shele Area, Arbaminch Zuria District, Southwest Ethiopia. Malar J. 2015;14.
Crossref - Gilson PR, Nebl T, Vukcevic D, et al. Identification and Stoi-chiometry of Glycosylphosphatidylinositol-Anchored Membrane Proteins of the Human Malaria Parasite Plasmodium Falciparum. Mol Cell Proteomics. 2006;5(7):1286-1299.
Crossref - Fenton B, Clark JT, Khan CMA, et al. Structural and Antigenic Polymorphism of the 35- to 48-Kilodalton Merozoite Surface Antigen (MSA-2) of the Malaria Parasite Plasmodium Falciparum. Mol Cell Biol. 1991;11(2):963-971.
Crossref - Yavo W, Konate A, Mawili-Mboumba DP, et al. Genetic Polymorphism of Msp1 and Msp2 in Plasmodium Falciparum Isolates from Cote d’Ivoire versus Gabon. J Parasitol Res. 2016;2016:3074803.
Crossref - Congpuong K, Sukaram R, Prompan Y, Dornae A. Genetic Diversity of the Msp-1, Msp-2, and Glurp Genes of Plasmo-dium Falciparum Isolates along the Thai-Myanmar Borders. Asian Pac J Trop Biomed. 2014;4(8):598.
Crossref - Hamid MMA, Mohammed SB, El Hassan I.M. Genetic Diversity of Plasmodium Falciparum Field Isolates in Central Sudan Inferred by PCR Genotyping of Merozoite Surface Protein 1 and 2. N Am J Med Sci. 2013;5(2):95-101.
Crossref - Dajem SMB. Genetic Diversity in the Msp-1 and Msp-2 Alleles among Plasmodium Falciparum Field Isolates from Jazan, Saudi Arabia. J Egypt Soc Parasitol. 2015;45(1):199-206.
Crossref - Mohammed H, Hassen K, Assefa A, et al. Genetic Diversity of Plasmodium Falciparum Isolates from Patients with Uncomplicated and Severe Malaria Based on Msp-1 and Msp-2 Genes in Gublak, North West Ethiopia. Malar J. 2019;18(1):413.
Crossref - Mohammed H, Kassa M, Assefa A, Tadesse M, Kebede A. Genetic Polymorphism of Merozoite Surface Protein-2 (MSP-2) in Plasmodium Falciparum Isolates from Pawe District, North West Ethiopia. PLoS One. 2017;12(5):e0177559.
Crossref - Chekol T, Alemayehu GS, Tafesse W, et al. Genetic Diversity of Merozoite Surface Protein-1 and -2 Genes in Plasmodium Falciparum Isolates among Asymptomatic Population in Boset and Badewacho Districts, Southern Ethiopia. J Parasitol Res. 2022;2022:7728975.
Crossref - Eldh M, Hammar U, Arnot D, Beck HP, et al. Multiplicity of Asymptomatic Plasmodium Falciparum Infections and Risk of Clinical Malaria: A Systematic Review and Pooled Analysis of Individual Participant Data. J Infect Dis. 2020;221(5):775-785.
Crossref - Some AF, Bazie T, Zongo I, et al. Plasmodium Falciparum Msp1 and Msp2 Genetic Diversity and Allele Frequencies in Parasites Isolated from Symptomatic Malaria Patients in Bobo-Dioulasso, Burkina Faso. Parasit Vectors. 2018;11(1):323.
Crossref - Mustafa SO, Hamid MMA, Aboud MA, et al. Genetic Diversity and Multiplicity of Plasmodium Falciparum Merozoite Surface Protein 2 in Field Isolates from Sudan. F1000Res. 2017;6:1790.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.