


ISSN: 0973-7510
E-ISSN: 2581-690X
Colibacillosis is an infectious disease in animals, which is caused by Escherichia coli, leading to economic losses. The current investigation was conducted to evaluate the molecular typing of colibacillosis-associated Escherichia coli for determination of O157:H7 serotype and virulence factors. Overall, 100 faecal samples were screened between November 2023 to February 2024 from in and around Jaipur district of Rajasthan. Sample screening was done for isolating E. coli in which 77 isolates were recovered from 100 diarrhoeic cattle and its serological identification indicated presence of the most frequent serotype, encompassing O76 (5), O3 (4), O7 (4), O16 (4), O50 (4), O111 (3), O113 (2), O125 (2), O135 (2), O2 (1), O4 (1), O9 (1), O19 (1), O27 (1), O66 (1), O85 (1), O109 (1), O114 (1) and O156 (1). Finally, Molecular characterization of the recovered isolates was conducted using PCR for detecting presence of fliCH7, rfbO157, stx1, stx2, EhlyA along with eaeA genes associated with E. coli. Among 77 E. coli isolates, none were recognized as O157:H7 serotypes. Among total E. coli isolates, 40 comprised Shiga toxin-producing E. coli (STEC) genes. The stx2 gene was the most prevalent (32.5%), followed by stx1 (17.5%), while combinations involving stx1, stx2 and EhlyA were detected in 10%-12.5% of isolates. The eaeA gene was rare, occurring in only 2.5% of the virulence-positive isolates. Results suggest that conventional culture methods, along with serological and molecular assays, are key techniques for isolating and characterizing STEC from fecal samples of diarrheic cattle.
Colibacillosis, Escherichia coli, STEC, Serotype, Virulotype
Colibacillosis is a significant neonatal disease in livestock that leads to substantial economic losses and is caused by Escherichia coli.1 In calves affected by colibacillosis, morbidity rates can range between 30% and 75%, while mortality rates may vary from 10%-50%, depending on the clinical care.2 Diarrheagenic E. coli (DEC) is classified into 6 primary groups, but only STEC is most significant in terms of zoonotic transmission.3 STEC is frequently found at high rates in cattle feces, particularly in newborn calves.4,5
Based on the modified Kauffman scheme, E. coli serotyping is performed by identifying their surface antigens: O (somatic), H (flagellar) and K (capsular).6,7 Over 400 O:H serotypes of STEC have been identified in cattle as well as humans, with more than 100 serotypes associated with human illnesses. “World Health Organization (WHO)” recognizes E. coli O157:H7 along with six other serogroups O45, O121, O26, O145, O111 and O103 as most prevalent STEC strains linked to human infections globally.8 E. coli O157 is transmitted via fecal-oral pathway, mainly through interactions between animals and farm visitors.9 Rise of STEC exhibits significant challenge to food safety along with global cattle industry.10
Production of stx1, stx2, intimin, as well as plasmid-encoded enterohemolysin is the primary cause of virulence of STEC. Shiga toxins usually inhibit synthesis of proteins, leading to host cell death along with conditions like “hemolytic uremic syndrome (HUS) and hemorrhagic colitis (HC), while intimin causes formation of attaching and effacing (A/E) lesions on intestinal” lining.11 Enterohemolysin is regarded as an important epidemiological marker for identifying STEC strains.12 STEC along with other E. coli strains can obtain virulence genes through horizontal gene transfer (HGT), leading to development of new E. coli pathotypes that represent a significant public health risk.13
Sample collection
For the present study, 100 rectal swab samples from diarrheic cattle (irrespective of age, sex and breed) in different places were collected from in and around locations of Jaipur district of Rajasthan. The samples included 25 from Bassi (C-1 to C-25), 15 from Sanganer (C-26 to C-40), 10 from Chaksu (C-41 to C-50), 15 from Chomu (C-51 to C-65), 20 from Jamwa Ramgarh (C-66 to C-85), and 15 from Jhotwara (C-86 to C-100). Samples were promptly collected, properly labelled and transported to the laboratory in an ice box for bacteriological analysis.
Isolation and characterization of Escherichia coli
Fecal samples were “enriched in nutrient broth for facilitating identification of E. coli. Enriched cultures were then streaked on MacConkey agar and then incubated for 24 hrs. Colonies showing a bright pink to red coloration were chosen and then subcultured on Eosin Methylene Blue (EMB) agar”. Following overnight incubation, colonies displaying typical metallic green sheen, suggestive of E. coli were further analysed using biochemical tests provided in a commercially available identification “kit (KB010 Hi E. coli Identification Kit, HiMedia)”.
DNA extraction
Genomic DNA was isolated by employing HiPurA® Multi-Sample DNA Purification Kit (MB554), following protocol provided by the manufacturer. Positive control used in this study were procured from HiMedia (MBT102) and the CDSRZ Laboratory.
Screening for virulence-associated genes
Polymerase chain reaction (PCR) was carried out by employing previously published oligonucleotide primers targeting virulence-associated genes, including stx1, stx2, eaeA, EhlyA, fliCH7 and rfbO157 as mentioned in Table 1.14-16 PCR mixture (10 µl total volume) comprised 2.0 µl of genomic DNA template, 1.0 µl of dNTP mix, 1.0 µl of 10× PCR buffer, 1.0 µl of each primer, 0.4 µl of Taq DNA polymerase, along with 4.6 µl of nuclease-free water. To confirm PCR amplification, electrophoresis on a 1.5% agarose gel at 100 V for 1 hr was performed. Amplified DNA fragments were visualized under UV illumination using the Fusion Solo S gel documentation system (Vilber, France), appearing as distinct bands of the expected size.
Table 1. Oligonucleotide Sequences and Thermal Cycling condition
Name of primer |
Specific genomic region |
Primer sequence |
Amplification conditions |
Size of amplicon |
|---|---|---|---|---|
stx1 |
Shiga toxin-1 |
F: TTCGCTCTGCAATAGGTA R: TTCCCCAGTTCAATGTAAGAT |
95 °C-5 min, 95 °C-30 sec, 54 °C-1 min, 72 °C-1.30 min, 72 °C-10 min, repeat of 35 cycles |
555 bp |
stx2 |
Shiga toxin-2 |
F: GTGCCTGTTACTGGGTTTTTCTTC R: AGGGGTCGATATCTCTGTCC |
95 °C-5 min, 95 °C-30 sec, 51°C-1 min, 72 °C-1.00 min, 72 °C-10 min, repeat of 40 cycles |
118 bp |
eaeA |
Intimin |
F: ATATCCGTTTTAATGGCTATCT R: AATCTTCTGCGTACTGTGTTCA |
95 °C-5 min, 95 °C-30 sec, 52 °C-1 min, 72 °C-1.30 min, 72 °C-10 min, repeat of 35 cycles |
425 bp |
EhlyA |
Entero-hemolysin |
F: GCATCATCAAGCGTACGTTCC R: AATGAGCCAAGCTGGTTAAGCT |
95 °C-5 min, 95 °C-45 sec, 59 °C-45 sec, 72 °C-45 sec, 72 °C-7 min, repeat of 35 cycles |
534 bp |
rfbO157 |
E. coli O157 |
F: CGTGATGATGTTGAGTTG R: AGATTGGTTGGCATTACTG |
94 °C-1 min, 94 °C-1 min, 51 °C-1 min, 72 °C-1 min, 72 °C-7 min, repeat of 35 cycles |
420 bp |
fliCH7 |
E. coli H 7 |
F: GCGCTGTCGAGTTCTATCGAGC R: CCACGGTGACTTTATCGCCATTCC |
94 °C-1 min, 94 °C-1 min, 59 °C-1:30 min, 72 °C-1 min, 72 °C-10 min, repeat of 35 cycles |
625 bp |
Serological Identification of E. coli Isolates
E. coli isolates obtained during the study were sent to National Salmonella and Escherichia Centre, Central Research Institute (CRI), Kasauli (Himachal Pradesh), for serotyping analysis.
Hemolysin activity
Phenotypic detection of enterohemolysin (EhlyA) was carried out on 5% sheep blood agar plates.17
Isolation and Identification of E. coli
Among 100 diarrhoeic fecal samples analyzed, 77 E. coli isolates were tested positive which indicated overall incidence rate of 77%. The isolates were cultured on selective media and subjected to Gram staining, which revealed Gram-negative, medium-sized rod-shaped bacteria. Biochemical characterization of the isolates demonstrated positive results for the Indole, β-Glucuronidase, Nitrate Reduction, Methyl Red, Lysine Decarboxylase, ONPG, and sugar fermentation tests (Lactose, Glucose, and Sucrose). The isolates tested negative for Citrate utilization, Voges-Proskauer, as well as Sorbitol fermentation tests. Additionally, all isolates were oxidase-negative as well as catalase-positive, consistent with typical biochemical profile of E. coli.
Detection of virulence genes in E. coli isolates
All 77 E. coli isolates were assessed for prevalence of virulence-associated genes through PCR. Genes targeted included those encoding intimin (eaeA), Shiga toxins (stx1, stx2), as well as enterohemolysin (EhlyA). Out of the total, 40 isolates exhibited positive results for one or more virulence genes. The Distribution of virulence genes was as follows: stx1 was detected in 7 isolates (17.5%) as shown in Figure 1, stx2 in 13 isolates (32.5%) (Figure 2), and both stx1 and stx2 in 5 isolates (12.5%). As shown in Figure 3, the eaeA gene was detected in only one isolate (2.5%) and was found in combination with stx2. Additionally, combinations involving the EhlyA gene were observed as illustrated in Figure 4, with 5 isolates (12.5%) positive for stx1 + EhlyA, 5 isolates (12.5%) for stx2 + EhlyA, and 4 isolates (10%) for stx1 + stx2 + EhlyA.
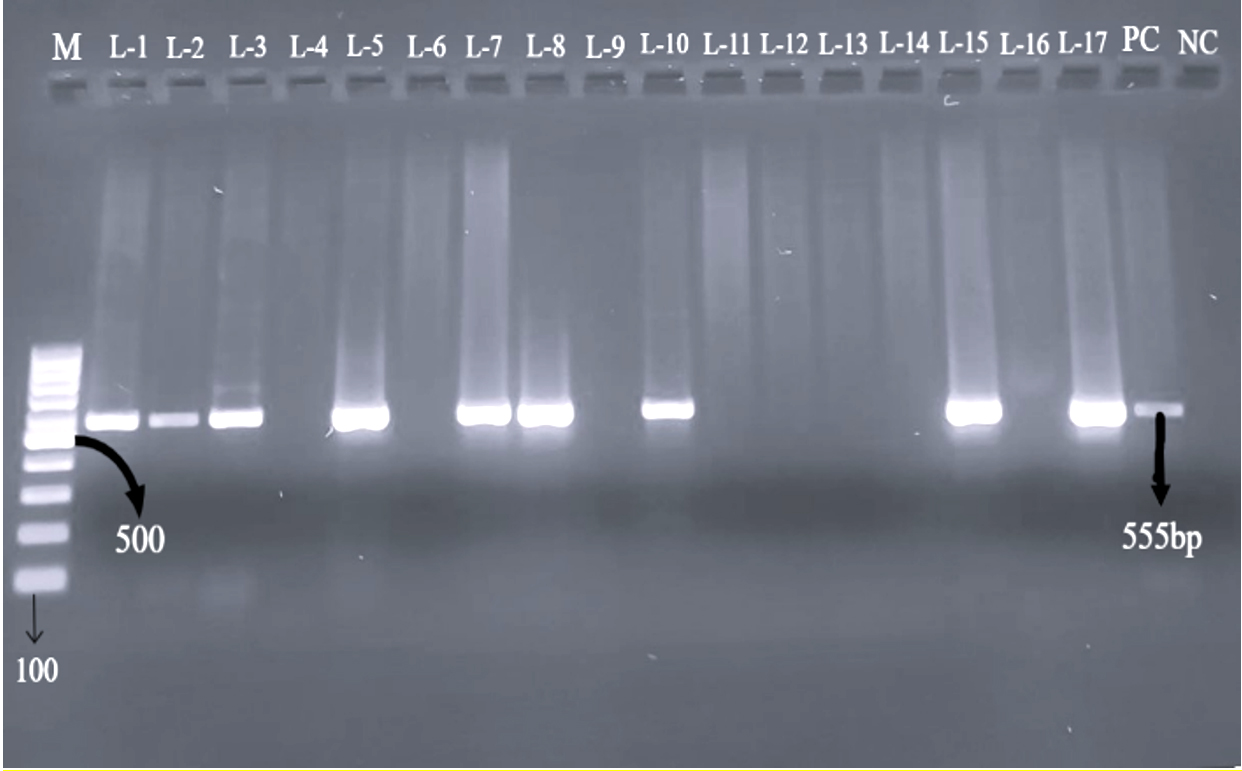
Figure 1. Agarose gel electrophoresis showing PCR products of 555 bp corresponding to the stx1 gene in tested isolates. Lane M: 100 bp DNA ladder; Lanes L1 (C-2), L2 (C-7), L3 (C-8), L5 (C-19), L7 (C-21), L8 (C-29), L10 (C-47), L15 (C-48) and L17 (C-88) represent positive samples; PC: positive control; NC: negative control
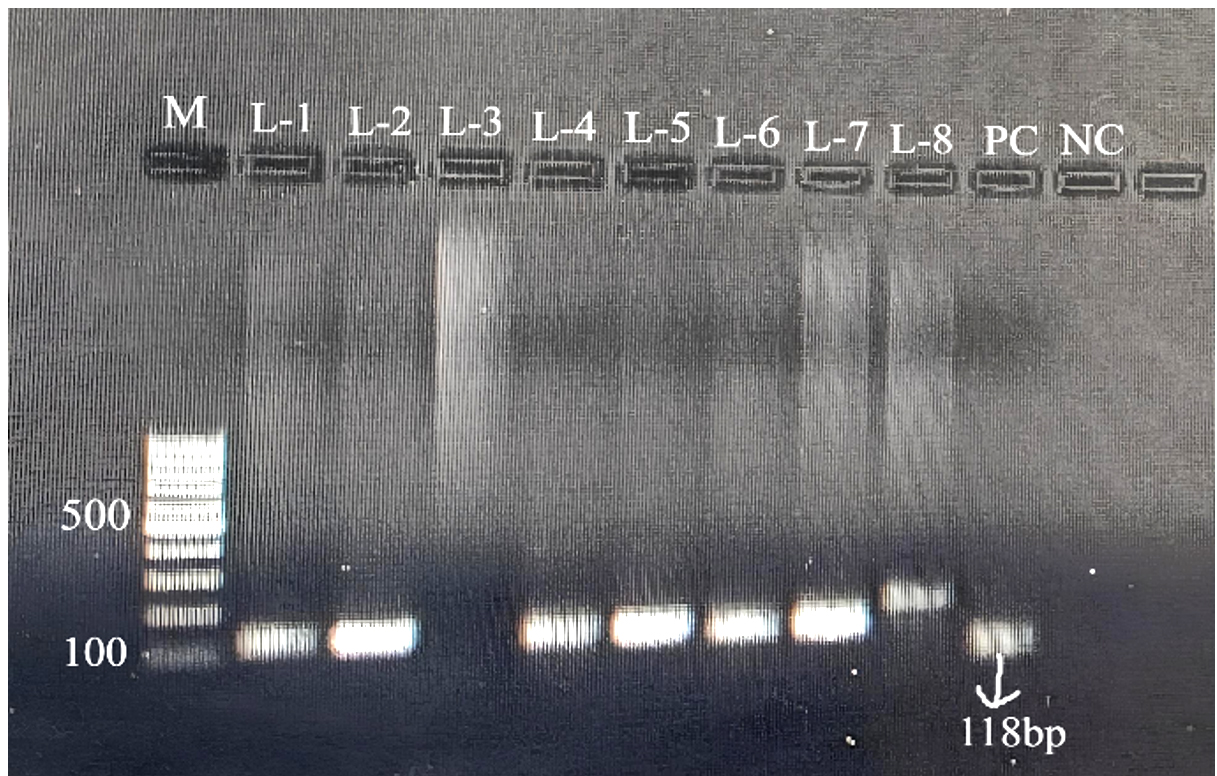
Figure 2. Agarose gel electrophoresis showing PCR amplification of the stx2 gene with a product size of 118 bp in isolates. Lane M: 100 bp DNA ladder; lanes L1 (C-2), L2 (C-3), L4 (C-8), L5 (C-27), L6 (C-48) and L7 (C-98) show positive samples; PC: positive control; NC: negative control
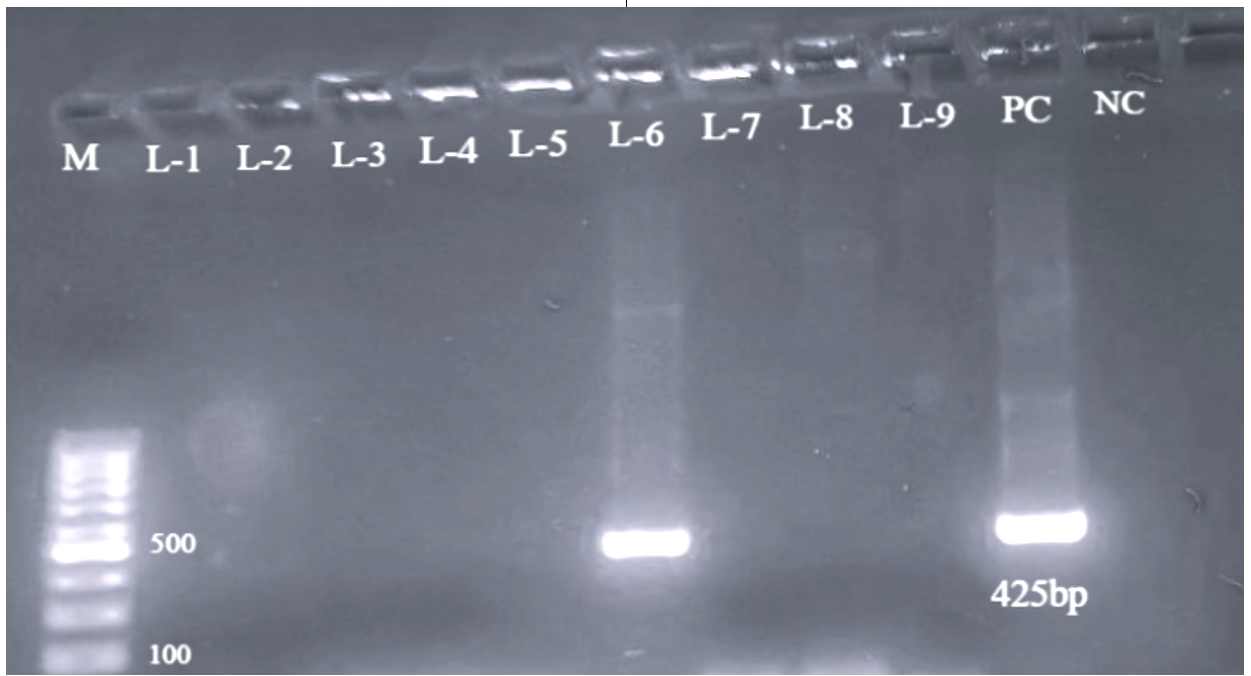
Figure 3. Agarose gel electrophoresis showing PCR amplification of the eaeA gene with a product size of 425 bp in the isolate C-34. Lane M: 100 bp DNA ladder; PC: positive control; NC: negative control
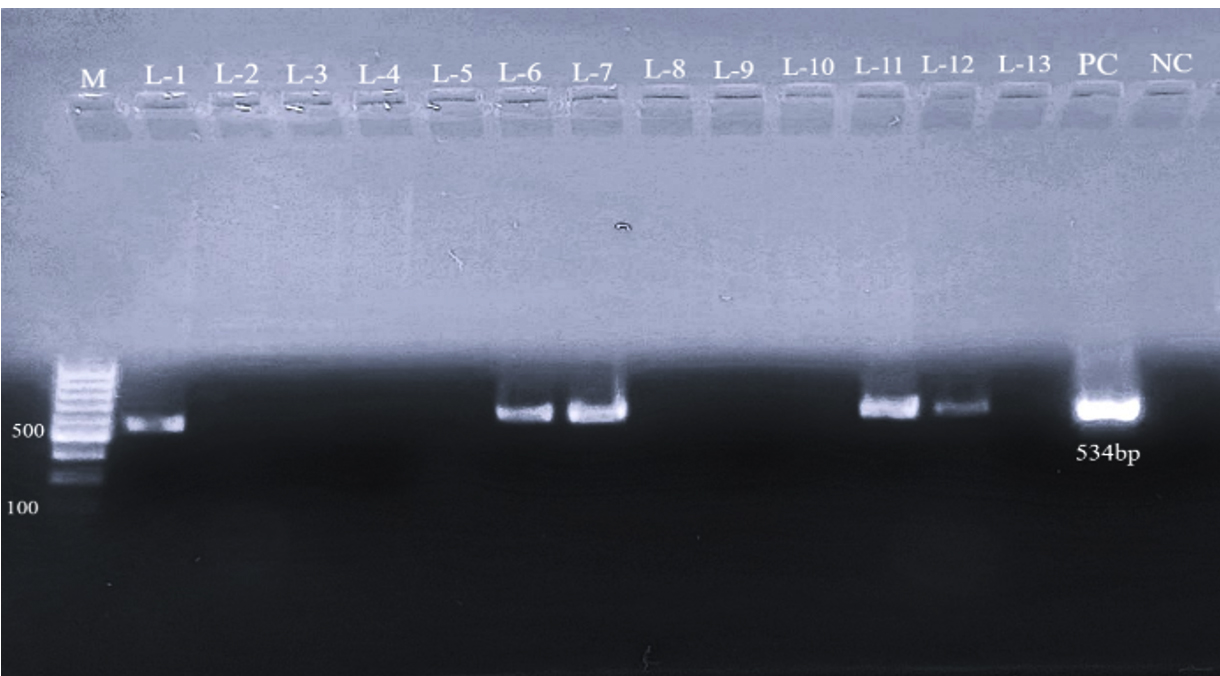
Figure 4. Agarose gel electrophoresis showing PCR products of 534 bp corresponding to the EhlyA gene in various isolates. Lane M: 100 bp DNA ladder; positive samples are shown in lanes L1 (C-2), L6 (C-7), L7 (C-10), L11 (C-21) and L12 (C-45); PC: positive control; NC: negative control
Among 77 E. coli isolates tested, stx1 gene was identified in 21 isolates (27.27%), while stx2 gene was present in 28 isolates (36.36%). The eaeA gene was identified in a single isolate (1.28%), and the EhlyA gene was found in 14 isolates (18.18%). None of the isolates showed amplification for the rfbO157 or fliCH7 genes, indicating absence of E. coli O157:H7 serotype within sampled population.
Virulence Gene Profiling of E. coli isolates
Virulotyping was conducted on E. coli isolates on basis of virulence genes (stx1, stx2, EhlyA and eaeA) presence. Among 77 isolates, 40 (designated as STEC) carried one or more stx genes, either individually or in combination. These were classified into seven distinct virulotypes (V1 to V7) as shown in Table 2. Of these, virulotype V2, characterized by the presence of stx2 alone, was the most prevalent with 13 positive isolates, while virulotype V7 (stx2 and eaeA combined) was the least common, represented by only one isolate.
Table 2. Virulotyping of E. coli isolates
Virulotype |
Virulence factors |
Distribution in Sample (n = 40) |
Sample Id |
|---|---|---|---|
V-1 |
stx1 alone |
7 |
C-47, C-54, C-57, C-65, C-82, C-88, C-97 |
V-2 |
stx2 alone |
13 |
C-12, C-16, C-36, C-37, C-38, C-39, C-42, C-43, C-89, C-90, C-95, C-96, C-98 |
V-3 |
stx1 & stx2 |
5 |
C-24, C-27, C-29, C-48, C-83 |
V-4 |
stx1 & EhylA |
5 |
C-7, C-10, C-21, C-45, C-58 |
V-5 |
stx2 & EhylA |
5 |
C-1, C-3, C-5, C-6 C-87 |
V-6 |
stx1, stx2 & EhylA |
4 |
C-2, C-4, C-8, C-19 |
V-7 |
stx2 & eaeA |
1 |
C-34 |
Determination of serotypes in E. coli isolates
In the study, out of 77 isolates, there were 19 different serotypes were observed in 70 randomly selected studied isolates. Out of which 40 were serotyped for “O” antigen whereas 2 were rough and the remaining 28 were untypable isolates. The most frequent serotype was reported O76 (5) followed by O3 (4), O7 (4), O16 (4), O50 (4), O111 (3), O113 (2), O125 (2), O135 (2), O2 (1), O4 (1), O9 (1), O19 (1), O27 (1), O66 (1), O85 (1), O109 (1), O114 (1) and O156 (1) as mentioned in Table 3.
Table 3. Serotyping of the E. coli isolates
| No. | Identified Serotypes | Isolate ID E. coli | Total no. of serogroups isolates | Prevalence of (%) |
|---|---|---|---|---|
| 1. | O2 | C-66 | 1 | 1.428% |
| 2. | O3 | C-25, C-81, C-93, C-100 | 4 | 5.71% |
| 3. | O4 | C-92 | 1 | 1.428% |
| 4. | O7 | C-36, C-38, C-54, C-72 | 4 | 5.71% |
| 5. | O9 | C-98 | 1 | 1.428% |
| 6. | O16 | C-9, C-15, C-23, C-57 | 4 | 5.71% |
| 7. | O19 | C-51 | 1 | 1.428% |
| 8. | O27 | C-19 | 1 | 1.428% |
| 9. | O50 | C-1, C-3, C-16, C-53 | 4 | 5.71% |
| 10. | O66 | C-27 | 1 | 1.428% |
| 11. | O76 | C-4, C-18, C-24, C-84, C-99 | 5 | 7.14% |
| 12. | O85 | C-6 | 1 | 1.428% |
| 13. | O109 | C-58 | 1 | 1.428% |
| 14. | O111 | C-37, C-65, C-83 | 3 | 4.28% |
| 15. | O113 | C-34, C-45 | 2 | 2.85% |
| 16. | O114 | C-71 | 1 | 1.428% |
| 17. | O125 | C-2, C-10 | 2 | 2.85% |
| 18. | O135 | C-62, C-87, | 2 | 2.85% |
| 19. | O156 | C-12 | 1 | 1.428% |
| 20. | Untypable (U.T.) | C-5, C-7, C-8, C-11, C-13, C-17, C-21, C-22, C-28, C-29, C-31, C-32, C-35, C-41, C-42, C-43, C-46, C-47, C-48, C-50, C-63, C-82, C-86, C-89, C-90, C-95, C-96, C-97 | 28 | 40% |
| 21. | Rough | C-52, C-70 | 2 | 2.85% |
| Total | 70 | |||
*Serial numbers 20 and 21 do not represent serotypes
Among the 40 STEC isolates, serogrouping revealed association of virulence profiles with several O-serogroups. The predominant virulotype V2 (stx2 alone) was distributed among serogroups O7, O9, O50, O111, O156, and several untypable isolates. Serogroup O111 exhibited considerable virulence diversity, harboring V1, V2, and V3 virulotypes. The only eaeA-positive isolate (V7) belonged to serogroup O113. Untypable isolates displayed the greatest heterogeneity and were represented in all major virulotypes (V1-V6), indicating substantial genetic diversity among STEC strains. Serogroups O76, O111, O113 and O125 were associated with multiple virulence gene combinations, suggesting varying pathogenic potential within these serogroups. The correlation between virulotypes and serogroups of STEC isolates, as well as the distribution of serogroups and their associated virulotypes, are presented sequentially in Tables 4 and 5, respectively.
Table 4. Correlation between Virulotypes and Serogroups of STEC Isolates
Isolate ID |
Sero-group |
Virulo-type |
Virulence Gene Profile |
|---|---|---|---|
C-1 |
O50 |
V5 |
stx2 + EhlyA |
C-2 |
O125 |
V6 |
stx1 + stx2 + EhlyA |
C-3 |
O50 |
V5 |
stx2 + EhlyA |
C-4 |
O76 |
V6 |
stx1 + stx2 + EhlyA |
C-5 |
UT |
V5 |
stx2 + EhlyA |
C-6 |
O85 |
V5 |
stx2 + EhlyA |
C-7 |
UT |
V4 |
stx1 + EhlyA |
C-8 |
UT |
V6 |
stx1 + stx2 + EhlyA |
C-10 |
O125 |
V4 |
stx1 + EhlyA |
C-12 |
O156 |
V2 |
stx2 |
C-16 |
O50 |
V2 |
stx2 |
C-19 |
O27 |
V6 |
stx1 + stx2 + EhlyA |
C-21 |
UT |
V4 |
stx1 + EhlyA |
C-24 |
O76 |
V3 |
stx1 + stx2 |
C-27 |
O66 |
V3 |
stx1 + stx2 |
C-29 |
UT |
V3 |
stx1 + stx2 |
C-34 |
O113 |
V7 |
stx2 + eaeA |
C-36 |
O7 |
V2 |
stx2 |
C-37 |
O111 |
V2 |
stx2 |
C-38 |
O7 |
V2 |
stx2 |
C-39 |
Not serotyped/ UT |
V2 |
stx2 |
C-42 |
UT |
V2 |
stx2 |
C-43 |
UT |
V2 |
stx2 |
C-45 |
O113 |
V4 |
stx1 + EhlyA |
C-47 |
UT |
V1 |
stx1 |
C-48 |
UT |
V3 |
stx1 + stx2 |
C-54 |
O7 |
V1 |
stx1 |
C-57 |
O16 |
V1 |
stx1 |
C-58 |
O109 |
V4 |
stx1 + EhlyA |
C-65 |
O111 |
V1 |
stx1 |
C-82 |
UT |
V1 |
stx1 |
C-83 |
O111 |
V3 |
stx1 + stx2 |
C-87 |
O135 |
V5 |
stx2 + EhlyA |
C-88 |
Not serotyped |
V1 |
stx1 |
C-89 |
UT |
V2 |
stx2 |
C-90 |
UT |
V2 |
stx2 |
C-95 |
UT |
V2 |
stx2 |
C-96 |
UT |
V2 |
stx2 |
C-97 |
UT |
V1 |
stx1 |
C-98 |
O9 |
V2 |
stx2 |
C-90 |
UT |
V2 |
stx2 |
C-95 |
UT |
V2 |
stx2 |
C-96 |
UT |
V2 |
stx2 |
C-97 |
UT |
V1 |
stx1 |
C-98 |
O9 |
V2 |
stx2 |
C-90 |
UT |
V2 |
stx2 |
C-95 |
UT |
V2 |
stx2 |
C-96 |
UT |
V2 |
stx2 |
Table 5. Distribution of Serogroups and Their Associated Virulotypes among STEC Isolates
Serogroup |
Virulotype(s) Observed |
|---|---|
O7 |
V1, V2 |
O9 |
V2 |
O16 |
V1 |
O27 |
V6 |
O50 |
V2, V5 |
O66 |
V3 |
O76 |
V3, V6 |
O85 |
V5 |
O109 |
V4 |
O111 |
V1, V2, V3 |
O113 |
V4, V7 |
O125 |
V4, V6 |
O135 |
V5 |
O156 |
V2 |
Untypable (UT) |
V1, V2, V3, V4, V5, V6 |
Hemolysin activity
Isolates exhibiting presence of EhlyA gene by PCR were evaluated for hemolytic activity by employing 5% sheep blood agar. Findings demonstrated that all these isolates produced clear hemolysis as shown in Figure 5 on the agar plates.
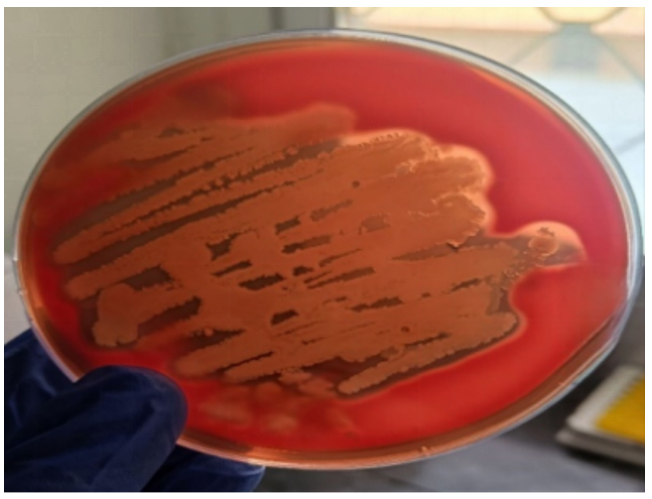
Figure 5. Enterohemolysin (Ehly) expression on 5% sheep blood agar plate
The present investigation demonstrated a considerable occurrence of Shiga toxin-producing Escherichia coli (STEC) among diarrhoeic cattle, with 40 of the 77 E. coli isolates harbouring one or more virulence-associated genes. The overall prevalence of STEC in the 100 samples examined was 40%, which is comparable with earlier reports from India and other countries, including Shinde et al.18 (39%), Leung et al.19 (41.5%), Cooley et al.20 (37.9%), and Ghoneim et al.21 (40%). These findings indicate that cattle serve as an important reservoir of STEC and may contribute to environmental contamination and transmission of pathogenic strains.
A higher prevalence of the stx2 gene compared to stx1 was observed in the present study. This is of epidemiological significance, as stx2-positive STEC strains are more frequently associated with severe human diseases such as haemorrhagic colitis and haemolytic uremic syndrome. Similar observations have been reported by Kuhnert et al.,22 Arya et al.,23 and Parul et al.,24 In contrast, some studies have reported a predominance of stx1 (Cerqueira et al.,25 and Buvens et al.,26) suggesting geographical variation in gene distribution. The eaeA gene was detected in only one isolate, indicating that most STEC strains may utilize alternative adherence mechanisms, as also reported in non-O157 bovine isolates.
Serotyping revealed substantial diversity with nineteen different serogroups. Although the O157 serogroup was not detected, several non-O157 serogroups of public health importance were identified. Among these, O111 and O113 are particularly noteworthy due to their established association with human infections and outbreaks.27,28 Several STEC serotypes, including O113:H21, O26:H11, O26, as well as O111, are recognized for their “zoonotic” potential.29 Several STEC serotypes, including O113:H21, O26:H11, O26, as well as O111, are recognized for their “zoonotic” potential.29 Furthermore, Capps et al.30 reported O2 and O109 as non-top STEC serogroups, with isolates carrying the stx2 gene, particularly subtype 2a, highlighting their possible role in human infections. Similar outcomes were demonstrated by Blanco et al.,31 who identified serotypes such as O113, O26, O91, and O8, and by Shahrani et al.32 who documented a diverse range of non-O157 STEC serotypes, including O128, O121, and O113, among diarrheic calves in Iran.
Virulotyping identified seven distinct virulotypes, indicating marked genetic variability among isolates. The predominance of the stx2-only profile suggests its wide distribution in the study population. The presence of combinations of stx1, stx2 and EhlyA genes indicates circulation of strains with enhanced virulence potential, which may be of greater public health concern.
Overall, the study highlights the epidemiological importance of non-O157 STEC in cattle. Although O157 was not detected, the presence of several zoonotically significant non-O157 serogroups underscores their potential public health risk. Continuous surveillance is essential to monitor their distribution and evolution.
The study is limited by its relatively small sample size, single geographical coverage, and analysis of a limited set of virulence genes. Further studies involving larger multi-regional sampling, Shiga toxin subtyping, antimicrobial resistance profiling, and whole-genome sequencing are recommended for a more comprehensive understanding of STEC epidemiology.
The present study demonstrated the occurrence of diverse STEC-associated Escherichia coli strains in diarrhoeic cattle from Jaipur district, Rajasthan. Among the 77 E. coli isolates recovered, a considerable proportion carried one or more virulence genes, confirming the presence of potentially pathogenic STEC in the study population. The predominance of the stx2 gene suggests the circulation of strains with increased virulence potential, while the low detection of eaeA indicates that alternative pathogenic mechanisms may be involved in these isolates.
Although the O157:H7 serotype was not detected, several non-O157 serogroups, including O111, O113 and O125, were identified. These serogroups have been associated with human infections and therefore highlight the zoonotic relevance of cattle-derived STEC. The diversity of serogroups and virulence profiles observed in this study emphasizes the importance of non-O157 STEC as an emerging public health concern.
The findings confirm that diarrhoeic cattle can serve as reservoirs of potentially zoonotic STEC and reinforce the need for continuous surveillance of non-O157 serogroups. Further studies involving larger sample populations and advanced molecular characterization are required to better understand the epidemiology, virulence attributes and public health significance of STEC circulating in cattle.
ACKNOWLEDGMENTS
The authors sincerely acknowledge the Centre for Diagnosis, Surveillance, and Response to Zoonotic Diseases (CDSRZ), Department of Veterinary Medicine, Post Graduate Institute of Veterinary Education and Research (PGIVER), Jaipur, for providing facilities and support for this study. Authors are also grateful to the Principal Investigator, CDSRZ, and the Dean, PGIVER, Jaipur, for their valuable guidance, encouragement, and support.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
NKJ and SKS conceptualized and visualized the study. WC conducted the investigation. WC, AB, SM, AC, JB and A performed data curation and developed the methodology. WC wrote the manuscript. NKJ, SKS and JB reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study was approved by the Institutional Animal Ethical Committee, Post Graduate Institute of Veterinary Education and Research (PGIVER), Jaipur, India (Regd. No. 1971/GO/Re/SL/17/CPCSEA; Dated 16 June 2017).
- Sa’Ayinzat FE, Shaibu SJ, Tekdek LB. The Earliest Occurrence of Escherichia coli in Calves in Zaria, Nigeria. Int J Curr Microbiol Appl Sci. 2015;4(6):218-223
- Radostits OM, Gay CC, Hincheliff KW, Cos D. Verinary Medicine. A text book of the diseases of cattle, sheep, pigs, goats and horses, 10th Edition, New York, W.B. Saunders Company Ltd. 2000;77:847-896.
- Fadel HM, Afifi R, Al-Qabili DM. Characterization and zoonotic impact of Shiga toxin producing Escherichia coli in some wild bird species. Vet World. 2017;10(9):1118.
Crossref - Dastmalchi SH, Ayremlou N. Characterization of Shiga toxin producing Escherichia coli (STEC) in faeces of healthy and diarrhoeic calves in Urmia region, Iran. Iran J Microbiol. 2012;4(2):63
- Islam MZ, Musekiwa A, Islam K, et al. Regional variation in the prevalence of E. coli O157 in cattle: A Meta-Analysis and Meta-Regression. PLoS One. 2014;9(4):93299.
Crossref - Edwards R, Ewing WN. Identification of Enterobacteriaceae. 3rd edition., Burgess Publishing Co., Minnesota. 1972.
- Lior H. Classification of Escherichia coli. In: Carlton L. Gyles, (eds). Escherichia coli in Domestic Animals and Humans. Wallingford, United Kingdom: CAB International; 1994: 31-72.
- Panel EB, Koutsoumanis K, Allende A, et al. Pathogenicity assessment of Shiga toxin producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020;18(1):5967.
Crossref - Stein RA, Chirila M. Routes of transmission in the food chain. In: Dodd CER, Aldsworth T, Stein RA, Cliver DO, Riemann HP, ed. Foodborne diseases. Academic Press. 2017:65-103.
Crossref - Callaway TR, Edrington TS, Loneragan GH, Carr MA, Nisbet DJ. Shiga toxin producing Escherichia coli (STEC) ecology in cattle and management based options for reducing faecal shedding. Agric Food Anal Bacteriol. 2013;3(1):39-69.
- Dean-Nystrom EA, Bosworth BT, Cray Jr WC, Moon HW. Pathogenicity of Escherichia coli O157: H7 in the intestines of neonatal calves. Infect Immun. 1997;65(5):1842-1848.
Crossref - Farrokh C, Jordan K, Auvray F, et al. Review of Shiga-toxin-producing Escherichia coli (STEC) and their significance in dairy production. Int J Food Microbiol. 2013;162(2):190-212.
Crossref - Muller D, Greune L, Heusipp G, et al. Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl Environ Microbiol. 2007;73(10):3380-3390.
Crossref - Franck SM, Bosworth BT, Moon HW. Multiplex PCR for enterotoxigenic, attaching and effacing and Shiga toxin-producing Escherichia coli strains from calves. J Clin Microbiol. 1998;36(6):1795-1797.
Crossref - Seker E, Kus FS. The prevalence, virulence factors and antibiotic resistance of Escherichia coli O157 in faeces of adult ruminants 19 slaughtered in three provinces of Turkey. Veterinarski Arhiv. 2019;89(1):107- 121.
Crossref - Maurer JJ, Schmidt D, Petrosco P, et al. Development of primers to O-antigen biosynthesis genes for specific detection of Escherichia coli O157 by PCR. Appl Environ Microbiol. 1999;65(7):2954-2960.
Crossref - Beutin L, Prada J, Zimmermann S, Stephan R, Ørskov I, Ørskov F. Enterohemolysin, a new type of hemolysin produced by some strains of enteropathogenic E. coli (EPEC). Zentralblatt Für Bakteriologie Mikrobiologie Und Hygiene Series a Medical Microbiology Infectious Diseases Virology Parasitology. 1988;267(4):576-588.
Crossref - Shinde DB, Singhvi S, Koratkar SS, Saroj SD. Isolation and characterization of Escherichia coli serotype O157: H7 and other Verotoxin-producing E. coli in healthy Indian cattle. Vet World. 2020;13(10):2269.
Crossref - Leung PHM, Yam WC, Ng WWS, Peiris JSM. The prevalence and characterization of Verotoxin-producing Escherichia coli isolated from cattle and pigs in an abattoir in Hong Kong. Epidemiol Infect. 2001;126(2):173-179.
Crossref - Cooley MB, Jay-Russell M, Atwill ER, et al. Development of a robust method for isolation of Shiga toxin-positive Escherichia coli (STEC) from fecal, plant, soil and water samples from a leafy greens production region in California. PLoS One. 2013;8(6):65716.
Crossref - Ghoneim NH, Abdel-Moein KA, Mohamed MA. Are non-O157 Shiga toxin-producing Escherichia coli imposing their predominance over O157 in farm animals and human? Global Veterinaria. 2014;12(5):636-642.
Crossref - Kuhnert P, Dubosson CR, Roesch M, Homfeld E, Doherr MG, Blum JW. Prevalence and risk-factor analysis of Shiga toxigenic Escherichia coli in faecal samples of organically and conventionally farmed dairy cattle. Vet Microbiol. 2015;109(1-2):37-45.
Crossref - Arya G, Roy A, Choudhary V, Yadav MM, Joshi CG. Serogroups, atypical biochemical characters, colicinogeny and antibiotic resistance pattern of Shiga toxin producing Escherichia coli isolated from diarrhoeic calves in Gujarat, India. Zoonoses Public Health. 2008;55(2):89-98.
Crossref - Parul S, Bist B, Sharma B, Jain U, Yadav JK. A study on association of virulence determinants of Verotoxic Escherichia coli isolated from cattle calves. Vet World. 2016; 9(8):915.
Crossref - Cerqueira AMF, Guth BEC, Joaquim RM, Andrade JRC. High occurrence of Shiga toxin-producing Escherichia coli (STEC) in healthy cattle in Rio de Janeiro State, Brazil. Vet Microbiol. 1999;70(1-2):111-121.
Crossref - Buvens G, De Gheldre Y, Dediste A, et al. Incidence and virulence determinants of verocytotoxin-producing Escherichia coli infections in the Brussels-Capital Region, Belgium, in 2008–2010. J Clin Microbiol. 2012;50(4):1336-1345.
Crossref - Abd El-Tawab AA, Nasef SA, El-Hofy FI, Ibrahim OA. Prevalence of eaeA and qacEΔ1 genes in Escherichia coli isolated from omphalitis in baby chicks. Benha Vet Med J. 2017;32(1):184-192.
Crossref - Feng PC, Delannoy S, Lacher DW, et al. Shiga toxin-producing serogroup O91 Escherichia coli strains isolated from food and environmental samples. Appl Environ Microbiol. 2017;83(18):01231-17.
Crossref - Sandhu KS, Gyles CL. Pathogenic Shiga toxin-producing Escherichia coli in the intestine of calves. Can J Vet Res. 2002;66(2):65
- Capps KM, Ludwig JB, Shridhar PB, et al. Identification, Shiga toxin subtypes and prevalence of minor serogroups of Shiga toxin-producing Escherichia coli in feedlot cattle feces. Sci Rep. 2021;11(1):8601.
Crossref - Blanco M, Blanco JE, Mora A, et al. Serotypes, virulence genes, and intimin types of Shiga toxin (Verotoxin)-producing Escherichia coli isolates from cattle in Spain and identification of a new intimin variant gene (eae-ξ). J Clin Microbiol. 2004;42(2):645-651.
Crossref - Shahrani M, Dehkordi FS, Momtaz H. Characterization of Escherichia coli virulence genes, pathotypes and antibiotic resistance properties in diarrheic calves in Iran. Biol Res. 2014;47(1):28-40.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.