


ISSN: 0973-7510
E-ISSN: 2581-690X
This work was aimed at immobilization, characterization, and utilization of chitinase from Kurthia gibsonii Mb126. Immobilization of Kurthia gibsonii Mb126 chitinase on glutaraldehyde treated chitosan was carried out with immobilization yield of 106%. The optimal factors of the immobilization technique such as concentration of glutaraldehyde, chitinase concentration, and immobilization time were evaluated. After optimizing process parameters of immobilization (Glutaraldehyde concentration 4%, chitinase conc. 60mg, immobilization time 30min.), the specific activity of immobilized chitinase improved to 4.3-fold compared to the free form of chitinase. Temperature and pH optima of the immobilized chitinase and free enzyme were same i.e., 7.5 and 40°C respectively. The relative activity of immobilized chitinase remained 90% at 40°C, at 50°C, and at 60°C for 120 min. In the pH range from 5.5 to 8, the immobilized chitinase retained 100% activity. The results confirmed that the pH stability and thermal stability of chitinase increased by immobilizing chitinase on chitosan. The immobilized enzyme system maintained 90% of its efficiency even after 16 successive reaction cycles. The immobilized chitinase maintained 78% of its activity even after 20 months. Fermentation of prawn shell waste with immobilized chitinase indicated a high level of deproteinization. Deproteinization experiments were carried out with 5mL (0.4 mg/mL ) of immobilized and free chitinase on 300 mg/mL of prawn shell waste for 20 days without any additional supplements at 40°C and 6.5 pH. Protein content was reduced from 38.4 to 0.8% with immobilized chitinase. Results suggests the possibility of using immobilized enzymes to remove the prawn shell waste from the environment. To the best of our knowledge there was no such study about the deproteinization of prawn shell waste using immobilized chitinase till the date.
Kurthia gibsonii Mb126, Chitinase, Deproteinization, immobilization, chitosan
India alone generates 1,00,000 tonnes of prawn wastes every year by the shrimp processing industry.1 Rattanakit et al.,2 reported that, these chitinous wastes are disposed of through ocean dumping, incineration, and landfilling, leading to resource depletion and creating environmental pollution.2 Deproteinization process helps to extract protein content which leads to the transformation of chitinous waste into chitin-derived end products. In chemical methods of deproteinization concentrated acids and bases are using, and this results in structural changes of the chitin such as removal of acetyl group and depolymerisation, which leads to undesired end products and it also creates waste disposal problems.3 Eco-friendly deproteinization processes by enzymatic treatments have been evolving as a preferred treatment process, which is more economic and allows easy recovery of end products.
Enzymes are potential biocatalysts on account of their distinctive physicochemical characteristics.4 They are remarkably excellent because of their versatile role in oxidation/reduction reactions, hydrolysis, transfer of functional group, cleavage of various bonds, isomerisation, joining new bonds between two molecules and translocation of ions or molecules.5 With the advancement in biotechnology, mainly in the area of molecular biology, proteomics and genomics there are exciting possibilities for the commercial uses of enzymes in various fields such as production of bio diesel,6 various dairy products,7 pharmaceuticals,8 and agriculturally important products.9 Enzymes are very efficient biological catalyst, therefore its production and utilization may be a better alternative of chemical catalysts.
Chitinase are the enzymes which degrades chitin, which is a cell wall component of insects, molluscs, crustaceans, fungi, algae, and marine invertebrates. Chitinase enzymes are widely using in various processes i.e., control of phytopathogens,10 isolation of fungal protoplasts,11 mosquito control by degrading cuticle of insects,12 and bioconversions of prawn shell waste to single-cell protein and ethanol.13 Chitinase hydrolyse chitin to chitooligomers, have functional properties that have made them possible in agricultural, industrial, and biomedical fields.14 Chitinase can be categorized into 2 major classes: endochitinases and exochitinases. Endochitinases cleave randomly at internal bonds, generating multimers of N acetylglucosamine (NAG), while exochitinase catalyzes the release of monomers of NAG from non-reducing end of chitin monofibril. In this work, a natural chitinolytic bacteria, K. gibsonii Mb126, isolated from coastal areas of “Ernakulum, Kerala, India” was selected for chitinase production. K. gibsonii Mb126 was cultured under solid substrate fermentation process in which dried shrimp shell powder was used as the only carbon and energy source. Kurthia gibsonii Mb126 was accepted as chitinase producer because of feasibility of mass culture of this bacteria by utilizing readily available low cost raw materials. An extracellular chitinase secreted by K. gibsonii Mb126 was purified to homogeneity by blending ammonium sulphate precipitation, DEAE cellulose ion exchange chromatography, affinity adsorption column chromatography and Sephadex G-100 gel filtration chromatography.
Commercial applications of chitinases are restricted by its cost of production and stability. The process of purification of chitinase was expensive, labour intensive, complicated and time-consuming. Enzyme Immobilization act as an excellent technique to overcome constraints such as shelf-life stability, high-cost of production, extraction and purification of enzymes.15 Immobilization reduces the enzyme cost by allowing the repeated use of enzymes for many times, it enhances catalytic activity, specificity and stability of the enzyme, it protect the enzyme from inhibitor and it offer highly pure product recovery and the successful separation of the enzyme from the reaction mixture.16-19
Mainly four protocols are employing for enzyme immobilization, they are physical entrapment, non-covalent adsorption, bio-conjugation and covalent attachment. Covalent bonds are forming between functional groups of protein and leads to stable attachment. Covalent technique is the most preferred method for immobilization of enzymes because it prevents enzyme diffusion.20 Chitosan is an ideal support material for immobilization because of its features such as mechanical strength, low cost, and protecting enzymes from the action of metal ions, and chemicals.21,22 Chitosan is, biodegradable, hydrophilic, nontoxic, inert, and low cost material for immobilization of enzyme.23,24 Glutaraldehyde is the preferred functional cross-linking agent because it is soluble in water, efficient, and can enhance stability of immobilized enzyme by multipoint attachment with functional groups of enzymes, such as amine, imidazole, thiol, and phenol. High concentration of aldehyde groups on the surface of chitosan result the formation of multipoint inter-and intra-subunit covalent bonds between the enzyme and the matrix, so the enzyme immobilized on glutaraldehyde-activated chitosan will be more stable or unaffected to any changes in the conformation than randomly immobilized enzymes or their free enzyme.25 By introducing flexible spacer arms such as glutaraldehyde between enzyme and chitosan, reduce the steric hindrance, thus it allows to access the substrate more easily.26
The first objective of this work was to analyse covalent immobilization of chitinase onto chitosan which was previously activated by glutaraldehyde. The optimal parameters affecting immobilization such as concentration of glutaraldehyde, amount of chitinase, and immobilization time were determined. After that, pH and temperature profile of immobilized chitinase and free chitinase were analyzed. Then, the reusability and shelf life of immobilized chitinase were determined. Finally, the effectiveness of immobilized chitinase of K. gibsonii Mb126 for deproteinization of prawn shell waste was evaluated.
A protocol of covalent immobilization of chitinase on glutaraldehyde-activated chitosan was developed. The specific activity and stability towards changes in pH and temperature of immobilized chitinase were improved. After optimizing process parameters of immobilization (Glutaraldehyde concentration 4%, chitinase conc. 60mg, immobilization time 30min.), the specific activity of immobilized chitinase improved to 4.3-fold compared to the free form of chitinase. Temperature and pH optima of the immobilized chitinase and free enzyme were same i.e., 7.5 and 40°C respectively. The relative activity of immobilized chitinase remained 90% at 40°C, at 50°C, and at 60°C for 120 min. In the pH range from 5.5 to 8, the immobilized chitinase retained 100% activity. The results confirmed that the pH stability and thermal stability of chitinase increased by immobilizing chitinase on chitosan. The immobilized enzyme system maintained 90% of its efficiency even after 16 successive reaction cycles. The immobilized chitinase maintained 78% of its activity even after 20 months. Fermentation of prawn shell waste with immobilized chitinase indicated a high level of deproteinization. Deproteinization experiments were carried out with 5mL (0.4 mg/mL) of immobilized and free chitinase on 300 mg/mL of prawn shell waste for 20 days without any additional supplements at 40°C and 6.5 pH. Protein content was reduced from 38.4 to 0.8% with immobilized chitinase. Metal ions like Ca2+, and Mg2+ improved the enzymatic activity of immobilized chitinase, whereas Ag+, Fe2+, Cu2+ and Zn2+ inhibited enzyme activity. Triton X 100 and EDTA at a concentration of 10 mM decreased the relative activity of the immobilized enzyme by about 29±0.65% and 23±0.21%, respectively. SDS decreased the activity of immobilized chitinase to 47±0.32%. The immobilized chitinase could retain 75±1.34% of its activity in the presence of CTAB. Free chitinase was completely inhibited by DMSO, n-hexane, acetone, methanol, ethanol and n-propanol, however, these organic solvents have no remarkable influence on the activity of immobilized chitinase. Kinetic constants revealed that the immobilised chitinase was more sensitive to the substrate and two times faster than its free form. Fermentation of prawn shell waste with immobilized chitinase indicated a high level of deproteinization showed an excellent potential of Kurthia gibsonii Mb126 chitinase on chitosan for bioprocessing of prawn shell waste.
Purified chitinase extracted from Kurthia gibsonii Mb126, which was previously isolated from coastal areas of “Ernakulam, Kerala, India”, was used for immobilization experiments. Colloidal chitin was prepared from prawn shell powder.27 Analytical grade reagents were used in all experiments.
Kurthia gibsonii Mb126 was inoculated into medium containing powdered, dried prawn shell. Solid substrate fermentation was carried for 3 days with a moisture content of 75%. The temperature was set at 45°C and the pH of the moistening solution was 8. The exo chitinase released in the culture supernatant was purified and used for further experiments.
For chitinase activity analysis, colloidal chitin was used as the substrate. N-acetyl D-glucosamine, the hydrolytic end product of chitinase was estimated according to the method of Miller.28 The unit of the chitinase was defined as amount of enzyme required for liberating one mmole of N-acetyl D-glucosamine per minute under standard assay conditions. The culture supernatant was also analysed for protein concentration.29
Immobilization of chitinase by covalent methods Covalent linkage by chitosan30
One g of chitosan was mixed with 5 ml of 0.1M HCl and 2% glutaraldehyde solution (~ 0.2M), stirred gently, for 2 hr. at 4°C followed by overnight incubation. Chitosan was precipitated with 1 ml of 0.1M NaOH and was collected by filtration. The glutaraldehyde bounded chitosan was washed in water to remove the unbounded glutaraldehyde, then mixed with 40mg of purified chitinase (Specific activity- 10.31 U/mg) at 4°C overnight for enzyme immobilization. Immobilized enzyme activity of chitosan glutaraldehyde – chitinase conjugates was determined after precipitation and washing. Free chitinase activity and protein concentration were determined in each wash solution which contained unbound free chitinase.
The effectiveness of immobilization was determined:
Immobilization yield (%) = [Immobilized chitinase activity / Free chitinase ] × 100
Where,
Free chitinase = Chitinase added (U) – Unbound free chitinase (U)]
Optimization of Glutaraldehyde concentration for immobilization
5ml of 0.1M HCl containing various concentrations of glutaraldehyde (2 to 5%) was shaken with 1 g of chitosan for 2 h at 4°C. Immobilized chitinase activities were determined. Protein concentration and Free chitinase activity of each wash solution were determined.
Optimisation of Chitinase concentration for immobilization
To study the optimal concentration of chitinase on immobilization, different concentrations of (10mg to 70mg) enzyme immobilized onto chitosan which was pre-treated with 4% glutaraldehyde was used.
Optimization of immobilization time
For calculating the time required for effective immobilisation, 60mg chitinase was incubated with 4% glutaraldehyde treated chitosan for different time intervals beginning from 15 min. to 2h. After immobilization time, chitosan was washed thoroughly with acetate buffer and the assay has been conducted.
Production of chitooligosaccharides using the prepared immobilized chitinase
0.5 mL of immobilised chitinase (0.6%) was shaken with 0.9 ml (1%) soluble chitosan in 1.0 ml 0.05 M acetate buffer for 1.0 h at 40°C and at pH 5. The amount of chitooligosaccharides produced in the supernatant was estimated by dinitrosalicylic method.28
Degree of hydrolysis (D.H.) % = Chitooligosaccharide concentration(mg) / Chitosan concentration(mg)
Characterization of Free and Immobilized Chitinase
Characterization of Free and Immobilized Chitinase were analysed by determining the relative enzyme activity by the equation:
Relative activity (%) = A1/A0× 100
A0 is the maximum absorbance of the free/ immobilized chitinase, whereas A1 is the absorbance of the free/immobilized chitinase under different experimental conditions.
Effects of pH, and temperature on immobilized chitinase
pH profile of free and immobilized chitinase were analysed at pH range 5.0-8.5 by using 0.05 M acetate buffer of pH 5, 0.1M phosphate buffers having pH 5.5 to 8.0, and 0.1M sodium carbonate buffer of 8.5. The optimal temperature of free and immobilized chitinase were analysed at different temperature (20 to 80°C) at a pH of 6.5.
Thermostability of free and immobilized chitinase
Thermostability of free and immobilized chitinase was analysed at 40°C, 50°C, and 60°C for 2h and the enzyme assay was conducted in 20 min. time interval.
Reusability of immobilized chitinase
Chitinase-immobilized chitosan beads were removed and washed with distilled water after each use, and the reusability of immobilized chitinase was verified 16 times by estimating the activity.
Storage Stability of Immobilized Chitinase
After storing of immobilized chitinase–chitosan beads for different time interval up to 20 months at 4°C, the relative activity of immobilized chitinase was determined.
Kinetic properties of immobilised chitinase
Various concentrations of colloidal chitin were incubated with 0.5 mL of immobilised chitinase (0.6%) for 1 h. at pH 6.5 and temperature (40°C). The Km and Vmax were calculated from Line Weaver-Burk’s plot.
Effect of metal ions on immobilised chitinase
The immobilized chitinase 0.5 mL of immobilised chitinase (0.6%) was incubated with 10 mM concentration of various metal ions in 0.1 M phosphate buffer for 1 h. at pH 6.5 and temperature 40°C and specific activity were estimated.
Effects of ethylenediaminetetraacetic acid, detergents, and organic solvents
0.5 mL of immobilised chitinase (0.6%) was incubated with chelating agent ethylene diamine tetra acetic acid (EDTA), detergents such as cetyl trimethyl ammonium bromide (CTAB), Triton X-100 and SDS (10 mM) at 40°C. After 1 hour the activity of the immobilized chitinase were evaluated. The immobilized chitinase was incubated with 10% v/v of organic solvents including DMSO, n-hexane, acetone, methanol, ethanol and n-propanol in 0.1 M phosphate buffer for 1 h. at pH 6.5 and temperature 40°C and then relative activity of immobilized enzymes was determined.
Utilization of prawn shell waste
Shrimp shell waste powdered into uniform sized particles and used for further experiments. 300 mg prawn shell waste was mixed with 100 mL water (pH 6.5), was placed in 250-mL flasks. After sterilisation, 5 mL (0.4 mg/mL) of immobilized chitinase and free enzyme were transferred to different flask and carried out fermentation for different periods at 40°C on a shaking incubator. After specific time interval, fermented residues, were collected, oven-dried, and analyzed for deproteinization. Efficiency of deproteination (% DP) was measured 31 by the following equation:
% DP= ( PO . O) – (PR .R) x100/( PO . O)
PO and PR protein mass fractions in g/g before and after fermentation respectively.
O is the mass (g) of the original sample and R is the mass (g) of the residue after fermentation.
When Kurthia gibsonii Mb126 chitinase was immobilized on chitosan beads (2%) led to a 2.2-fold increase in the specific activity. Such a notable enhancement in the specific activity of immobilized chitinase revealed that the immobilization of chitinase on chitosan beads increased chitinase activity, and stability. Hence, this immobilization protocol used in our study successfully created a potent immobilized chitinase which could overcome diffusional limitation of substrate to the enzyme. Immobilization may result in stabilization improvement of the active centre of the chitinase through a bond between the amino group present in the chitosan and the enzyme.32 This may be due to the favourable and more protective environment for chitinase activity may have provided by chitosan beads. Chitosan was utilised in the immobilization of chitinases of Streptomyces griseus and Paenibacillus illinoisensis.33 Bushan34 also used chitosan beads for the immobilization of Bacillus sp. BG11 chitinase, in that case a 93.5% increase in the specific activity was reported.
Immobilization efficiency of chitinase with chitosan by covalent binding at various concentrations of glutaraldehyde (GA) as shown in Fig. 1. The immobilization efficiency of the chitinase immobilized with chitosan increased with the enhanced concentration of glutaraldehyde. The optimal concentration of glutaraldehyde for immobilization of chitinase with chitosan was 4%. Amino groups which are free on the chitosan were activated and become cross-linked with glutaraldehyde and consequently, more glutaraldehyde will be needed for immobilization and the immobilized amount of the enzyme also increased. In contrast Bilal et al.,35 and Spagna et al.,36 carried out enzyme immobilization with 2% and 0.5% glutaraldehyde concentration.
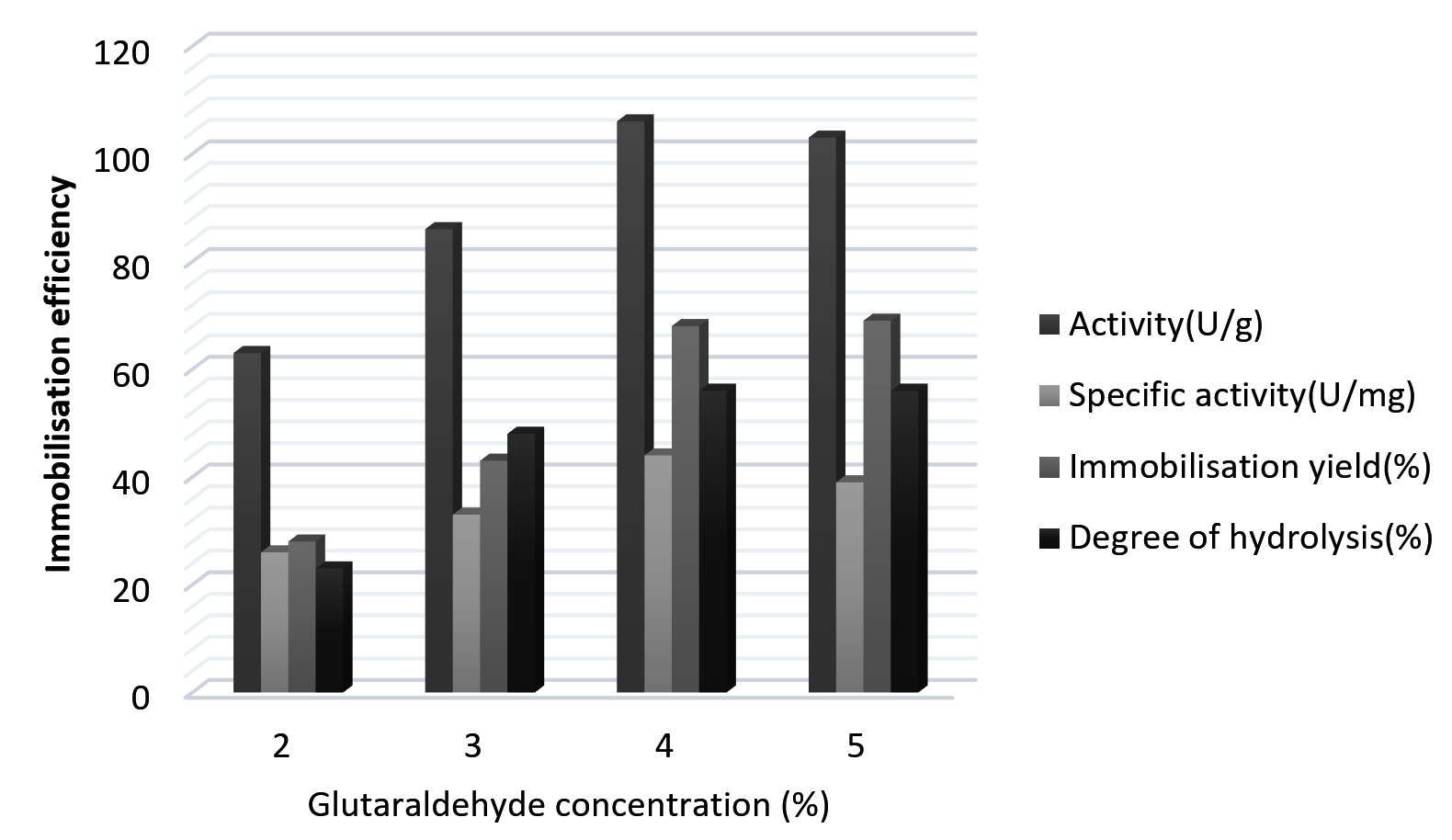
Fig. 1. Immobilization of Kurthia gibsonii Mb126 chitinase with chitosan by covalent binding at different concentrations of glutaraldehyde (GA).
Immobilization rate depends on the concentration and the specific activity of enzymes during immobilization. The affordability of the by-products depends on the cost of enzyme, so the optimization of the concentration of enzyme is important in immobilization study. Effect of chitinase concentration on Immobilization of Kurthia gibsonii Mb126 chitinase was shown in Fig. 2, as the concentration of chitinase increased, corresponding increase in the relative activity of the immobilized chitinase was noted. Relative activity was maximum (100%) with 60 chitinase /g of chitosan. The relative activity of the immobilized chitinase slightly decreased or remained constant when the enzyme loading increased to 70 mg. Too much enzyme loading leads to protein-protein interaction which reduces flexible stretching and causes steric hindrance and so the relative activity of the enzyme decreases.

Fig. 2. Effect of Chitinase Concentration on Immobilization of Kurthia gibsonii Mb126 chitinase.
Production of maximum enzyme activity in a short interval was the objective behind the optimal loading time determination. Fig. 3 shows relative activity during different time periods from 15 min. to 2h. with 60mg chitinase. The relative activity reached approximately 56% after 15 min. and 100% at 30 min and thereafter activity decreased. When immobilizing chitinase on chitosan the relative activity was approximately 56% after 15 min., shows that immobilization process happened rapidly. After 30 minutes chitinase activity decreased. This may be due to the formation of schiff’s bases and multiple bonds between the enzyme and chitosan beads and it may inactivate its active site and reduce the activity of the immobilized enzyme.37
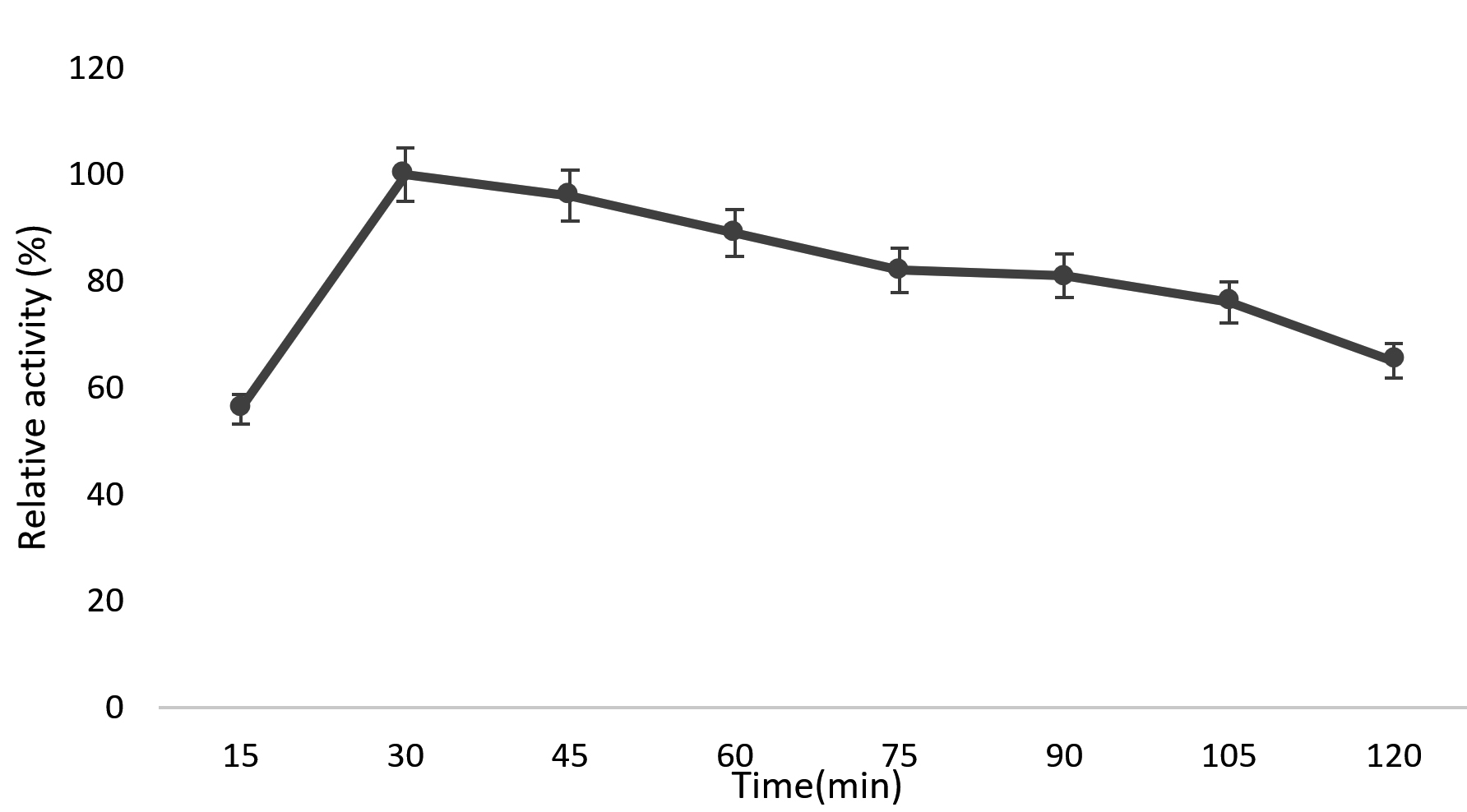
Fig. 3. Effect of time on Immobilization of Kurthia gibsonii Mb126 chitinase.
After optimizing process parameters of immobilization (Glutaraldehyde concentration 4%, chitinase conc. 60mg, immobilization time 30min.), the specific activity of immobilized chitinase improved to 4.3-fold compared to the free form of chitinase. After the optimisation of process variables, the immobilisation yield was increased to 95.8%.
Characterization of Free and Immobilized Chitinase
pH profile
Effect of different pH conditions on the stability of free and immobilized chitinase was shown in Fig. 4. At pH 6.5, free-chitinase and immobilized chitinase showed maximum activity. This may be due to too acid or too alkaline will cause enzyme denaturation, which leads to the reduction in enzyme activity. Free chitinase showed the 100% activity also at the same pH 6.5; however, above 6.5 the relative activity decreased rapidly. Reduction of relative activity of free chitinase may be due to the autolysis as well as aggregation of protein within the isoelectric point of free chitinase.38 However, immobilized chitinase may be resistant to these changes. In the pH range from 5 to 8, the immobilized chitinase was stable, immobilization appreciably enhanced pH stability. Immobilized chitinase of Pseudomonas aeruginosa K-187 was best at pH 8,39 and in the case of Aspergillus awamori EM66 the optimum pH was at 5.40
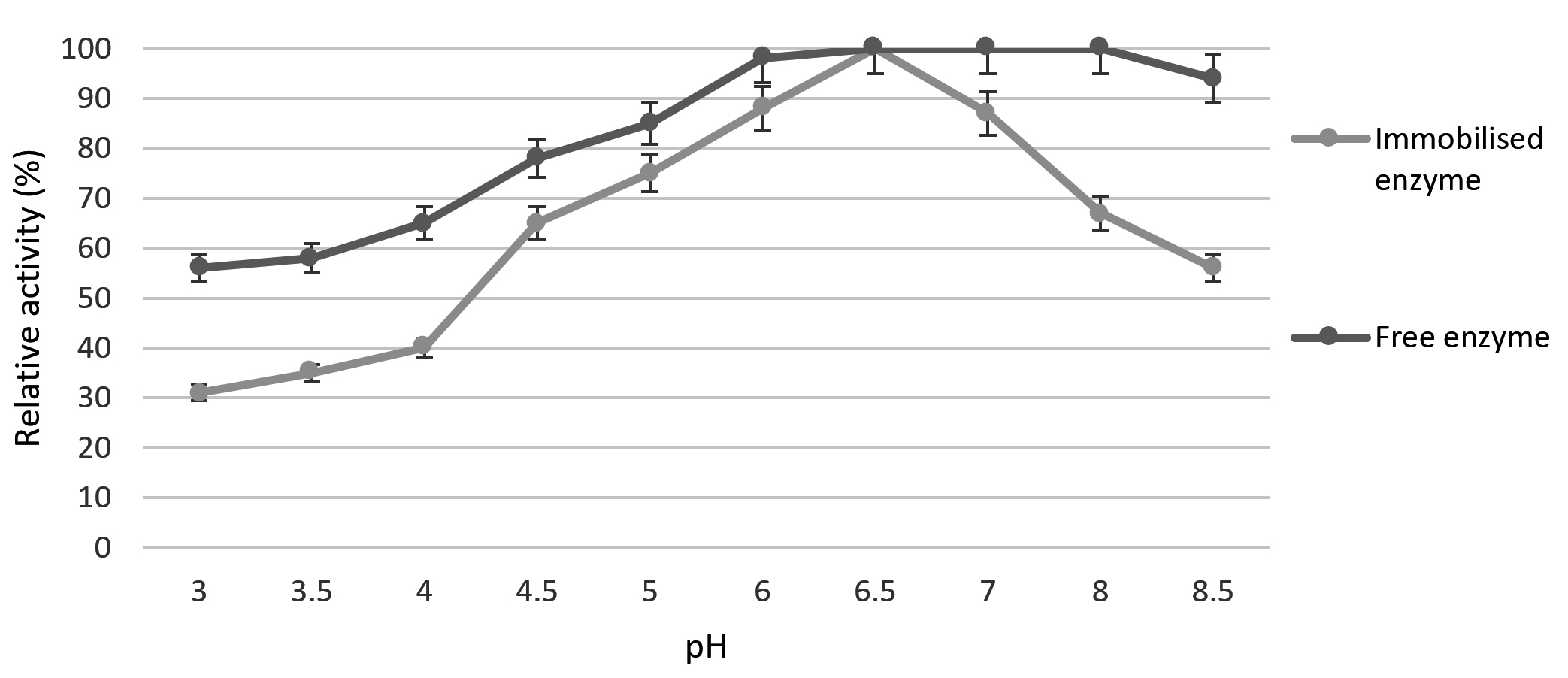
Fig. 4. pH profile of free and immobilized Kurthia gibsonii Mb126 chitinase.
Optimal temperature for chitinase immobilization
The relative activity as well as the denaturation rate of enzyme increase with increasing temperature. At medium temperature denaturation rate is low or negligible, however, the relative activity of enzyme increases with temperature. At higher temperatures the rate of enzyme denaturation increases. When the enzymatic reaction happens at an optimum temperature, equilibrium between relative activity and enzyme denaturation will established. The optimum temperature for soluble chitinase was at 40°C and the immobilized chitinase showed maximum activity between 40°C to 55°C (Fig. 5). The immobilized chitinase could withstand higher temperature ranges compared to free chitinase may be due to the multiple covalent bonds of immobilized enzyme. Similarly, the immobilized chitinase of Nocardia orientalis IFO 12806 was best at 40-50°C.41
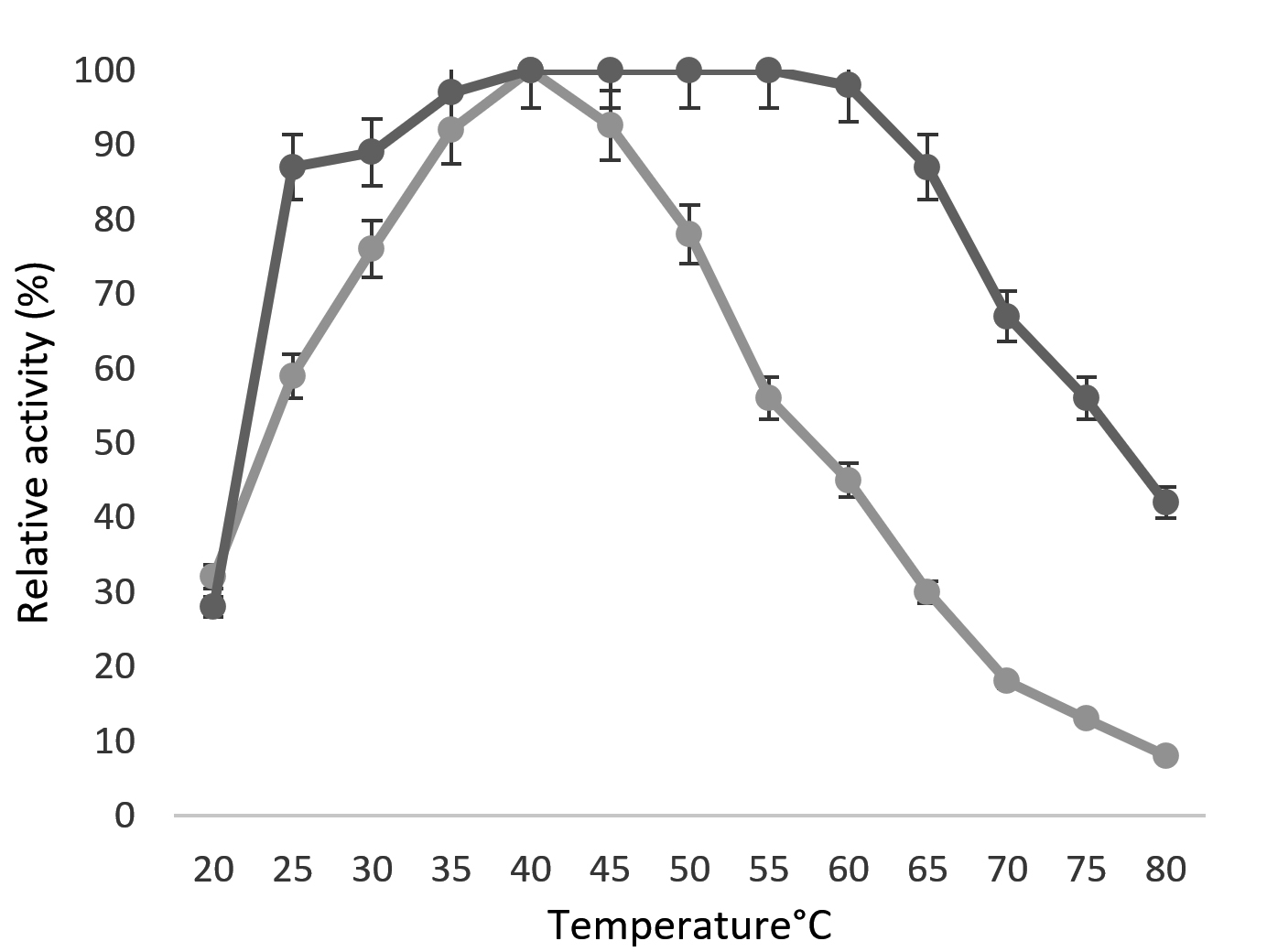
Fig. 5. The temperature profile of immobilized Kurthia gibsonii Mb126 chitinase.
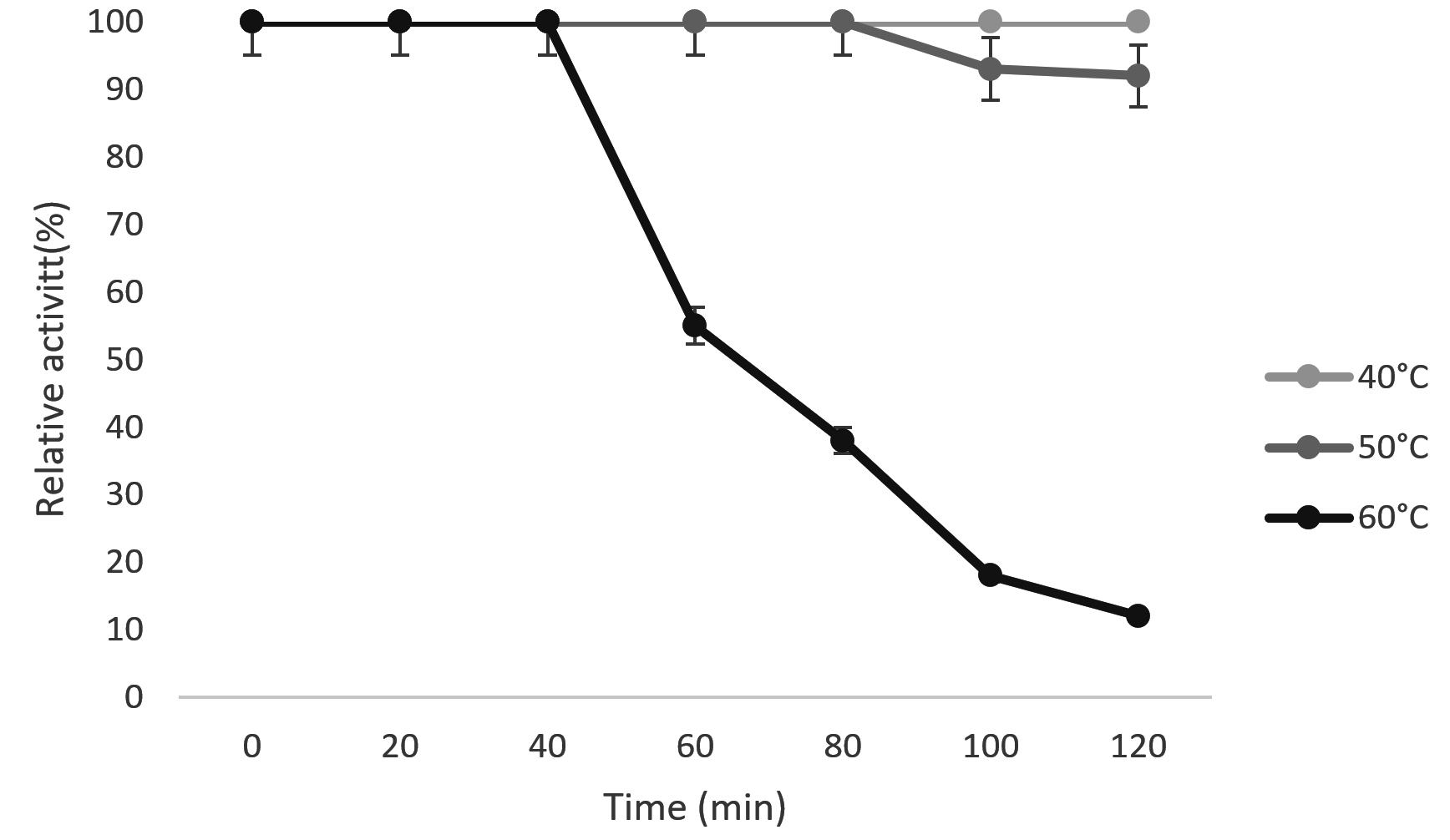
Fig. 6. Thermal stability of free chitinase.
Thermal stability
Thermal stability is one of the most important goal behind immobilization study. Fig. 6 and 7 shows the thermal stability of free and immobilized chitinase at 40°C, 50°C and 60°C. The purified chitinase enzyme was almost stable for 120 min. at 40°C and 50°C, however at 60°C the enzyme was stable for up to 40 min., then the enzyme activity decreased. Whereas, the relative activity of immobilized chitinase (Fig. 7) remained more than 90% at 40°C, at 50°C, and at 60°C for 120 min. Relative activity of free chitinase was decreased to12% after 2-hour incubation at 60°C, in contrast immobilized chitinase retained 92% activity. The results confirmed that the immobilized chitinase could withstand thermal unfolding compared to free chitinase. This increased thermal stability may be due to the cross-linking of chitinase enzymes with chitosan at multipoints.42 This results in induced rigidity in structure of immobilized enzyme so that denaturation and structural changes by temperature were limited.43,44 The enhanced thermal stability helpful in scale up process and high temperature reduce the chance of microbial contamination.45 A similar result was reported by Prasad and Palanivelu immobilized chitinase of Thermomyces lanuginosus showed remarkable stability at 50°C by keeping 45% of the activity for 6.46 Kidibule et al.,47 reported that the immobilized chitinase from Trichoderma harzianum CECT2413 retained 50% of its activity at 50°C.

Fig.7. Thermal stability of immobilized Kurthia gibsonii Mb126 chitinase.
Kinetic properties of immobilized Kurthia gibsonii Mb126 chitinase
The kinetic properties of immobilized chitinase were determined from Lineweaver–Burk plot (Fig. 8). Km and Vmax values of the free chitinase were 11.1 mg/ mL, 11.12 µmoles/µg h, respectively.48 Km and Vmax of immobilized chitinase showed 1mg/ mL, 20 µmoles/µg h, respectively. On comparing Km and the Vmax values of the free and the immobilized chitinase, we can conclude that the immobilised chitinase was more sensitive to the substrate than the free enzyme and the immobilized chitinase is two times faster than its free form. Kinetic parameters revealed that diffusion limitations were absent or negligible. Km and Vmax values of immobilized chitinase from Thermomyces lanuginosu were 1.3 mM and 4.5 mol/min/mg of protein, respectively.46
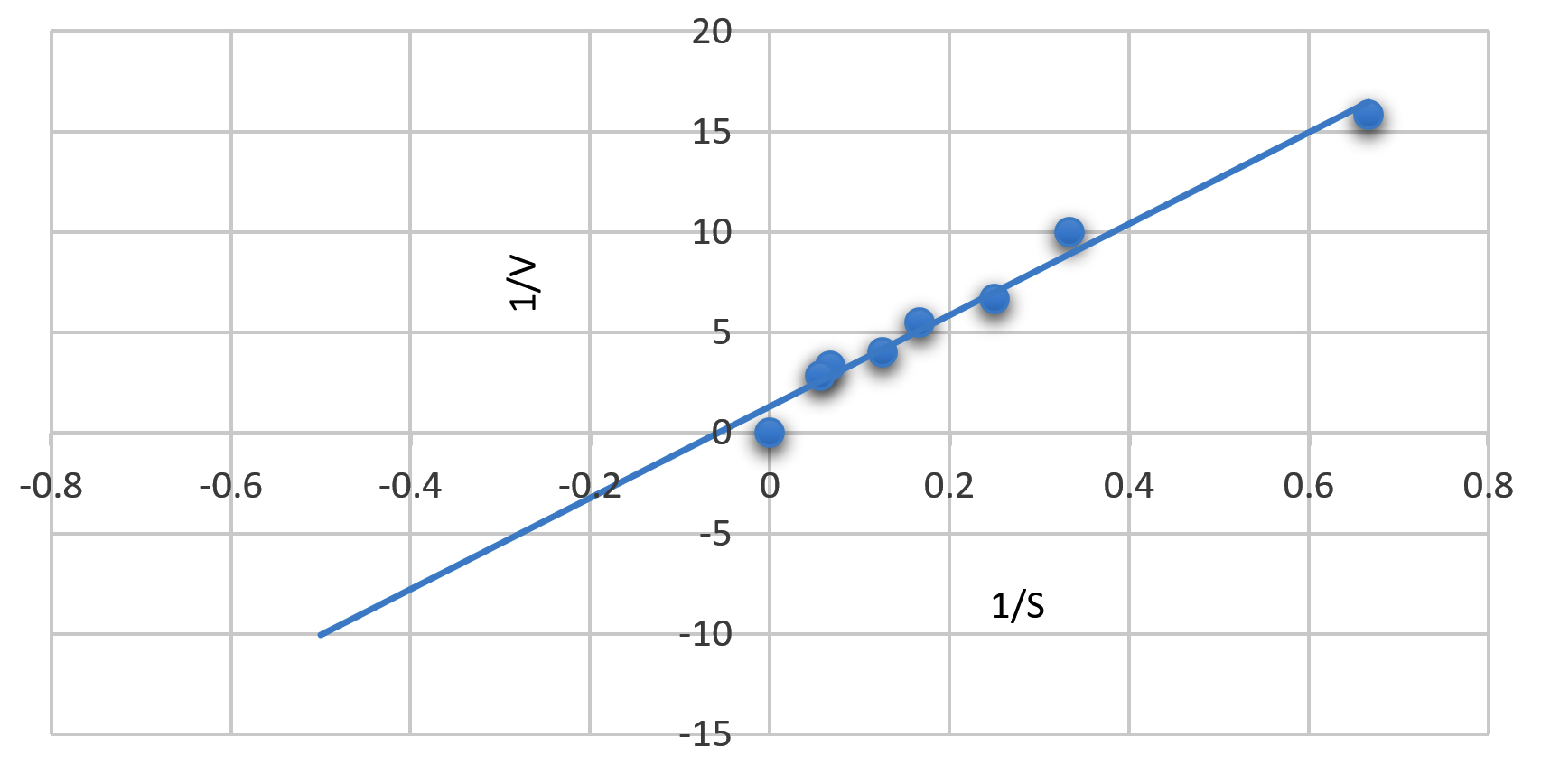
Fig. 8. LB Plot showing Km and Vmax of immobilized Kurthia gibsonii Mb126 chitinase.
Effect of metal ions on the immobilized Kurthia gibsonii Mb126 chitinase
Metal ions act as cofactors in enzymatic activity and they help to stabilize the enzyme–substrate binary complexes. Metal ions like Ca2+, and Mg2+ improved the activity of immobilized enzyme, whereas Ag+, Fe2+, Cu2+ and Zn2+ inhibited enzyme activity. The immobilized chitinase activation by Ca2+ was also observed,49 and the inhibition by Ag+, was also reported.46
Effect of metal ions on the immobilized Kurthia gibsoni chitinase was shown in Table 1.
Table (1):
Effect of metal ions on the immobilized Kurthia gibsonii chitinase.
Metal ion (10 mM) |
Relative activity of immobilised chitinase% |
|---|---|
Control |
100±0.18 |
Ca2+ |
123±0.25 |
Ag1+ |
67±1.25 |
Fe2+ |
45±2.54 |
Mg2+ |
186±3.2 |
Zn2+ |
40±1.03 |
Cu2+ |
16±0.64 |
Effects of ethylenediaminetetraacetic acid and detergents
The effects of detergents and EDTA on the immobilized Kurthia gibsonii Mb126 were studied (Table 2). Triton X 100 and EDTA at a concentration of 10 mM decreased the relative activity of the immobilized enzyme by about 29±0.65% and 23±0.21%, respectively. SDS decreased the activity of immobilized chitinase to 47±0.32%. The immobilized chitinase could retain 75±1.34% of its activity in the presence of CTAB. These results were in accordance with those reported in the literature that showed the negative effects of SDS and EDTA on the enzymatic activity of immobilized enzymes.50,51
Table (2):
Effects of ethylenediaminetetraacetic acid and detergents.
Chemicals (10 mM) |
Relative activity of immobilised chitinase(%) |
|---|---|
Control |
100±0.18 |
EDTA |
23±0.21 |
SDS |
47±0.32 |
Triton X 100 |
29±0.65 |
CTAB |
75±1.34 |
Effects of DMSO, n-hexane, acetone, methanol, ethanol and n-propanol
The effects of organic solvents on the immobilized Kurthia gibsonii Mb126 were studied (Table 3). A minute reduction (88%) in relative activity of immobilized chitinase was caused by n-propanol and acetone. Free chitinase was completely inhibited by DMSO, n-hexane, acetone, methanol, ethanol and n-propanol, however, these organic solvents have no remarkable effect on the activity of immobilized chitinase. The ability to resist organic solvents provides enormous applications in industrial field.
Table (3):
Effects of DMSO, n-hexane, acetone, methanol, ethanol and n-propanol.
Organic solvents (10%V/V) |
Relative activity of immobilised chitinase% |
|---|---|
DMSO |
100±0.18 |
n-Hexane |
98±0.24 |
Acetone |
87±0.45 |
Methanol |
100±0.78 |
Ethanol |
98±0.28 |
n-Propanol |
88±0.65 |
The reusability of immobilized chitinase
The reusability of immobilized Kurthia gibsonii Mb126 chitinase has been studied because of the importance of repeated use. Commercial chitinase are costly (more than Rs. 40,000 for 1g), hence reutilization is critical for many bioprocesses. In contrast to soluble free chitinase, immobilized chitinase can be recovered for repeated use. The chitinase immobilized on glutaraldehyde-treated chitosan by covalent had the excellent capacity to retain its residual activity even after 16 use and the results indicated that the autolysis of chitinase was limited by immobilization (Fig. 9) immobilized Kurthia gibsonii Mb126 chitinase retained 100% of its residual activity till 11 cycle. The 5% reduction in chitinase activity after 11th cycle may be due to the accumulation of product on the immobilized enzyme, which will cause diffusional limitation of the substrate. The immobilized chitinase of Pseudomonas aeruginosa k-187 could retain 70% of its activity even after 10 batches,39 the immobilized chitinase of Aspergillus awamori EM66 maintained enzyme activity for several cycles,40 and the immobilized chitinase of Alternaria infectoria preserved activity after 8 cycles.49

Fig. 9. Shows the reusability of immobilized Kurthia gibsonii Mb126 chitinase.
Storage stability of immobilized Kurthia gibsonii Mb126 chitinase
The relative activity of immobilized chitinase–chitosan beads was 78% after 20 months of storage at 4°C and autolysis of chitinase was limited by immobilization (Fig. 10). The immobilized chitinase of Thermomyces lanuginosus was remain stable after 1 month at 4°C.46 According to Danial et al.,17 for the production of the stable and reproducible biocatalysts, storage stability has important role.
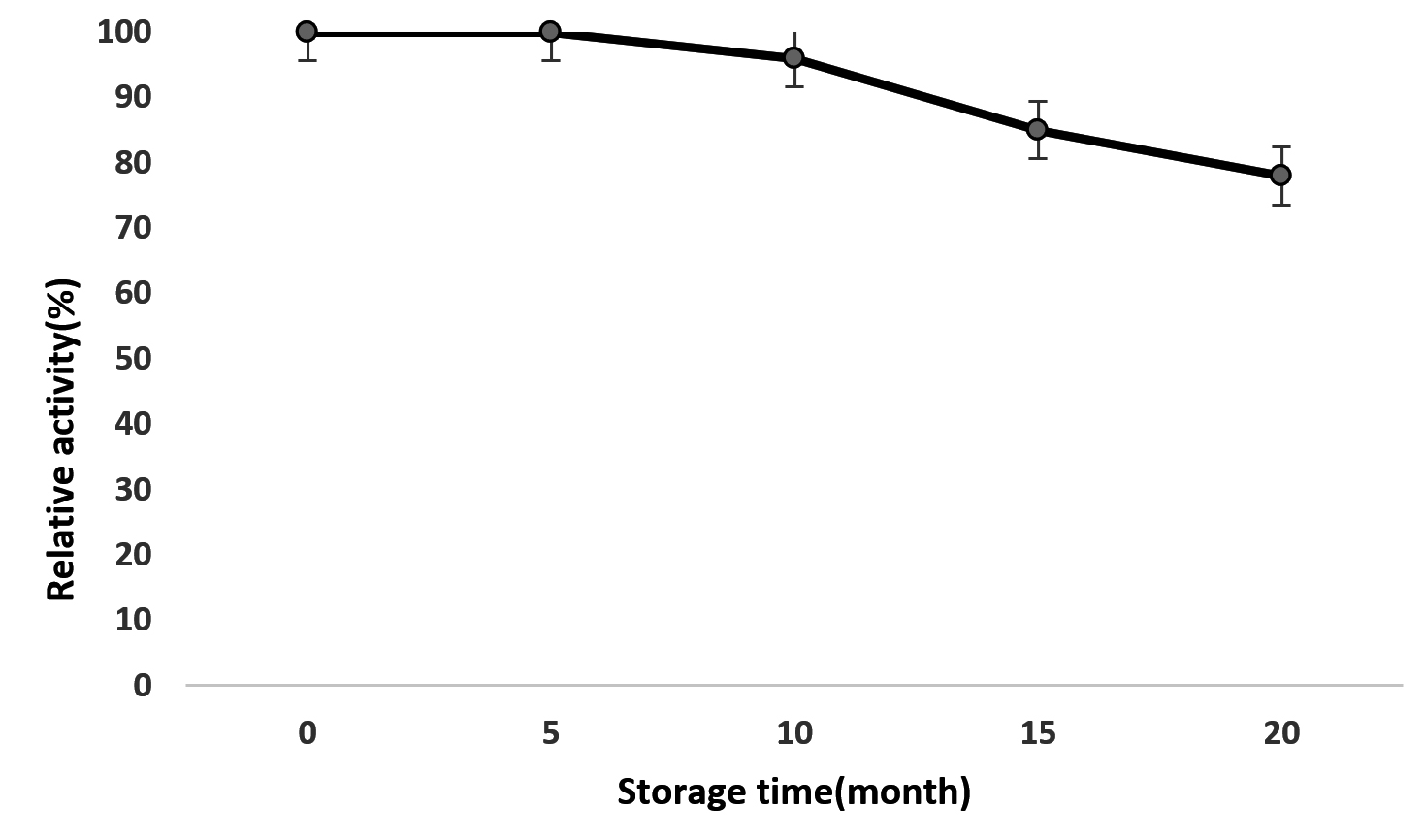
Fig. 10. Storage stability of immobilized Kurthia gibsonii Mb126 chitinase.
On comparing the characteristics can conclude that the immobilized Kurthia gibsonii Mb126 chitinase had more superior performance in thermal (92% of its initial activity at 60°C for 1.5h), operational (90% residual activity after 16 cycles), and storage stability (78% of its initial activity at 4°C for 20 months), than free chitinase.
Deproteinization of prawn shell waste by immobilized Kurthia gibsonii Mb126 chitinase
The seafood processing industry produces huge amounts of shellfish waste, and it creates environmental problems because degrade slowly. The amount of chitin in prawn shell is approximately 14-27%.52 Commercial manufacture of chitin from prawn shell waste involves deproteinization by strong bases and demineralization by strong acids leads to the accumulation of modified harmful end products like deacetylated polymers.53 Bioconversion of prawn shell waste is one of the most environment-friendly cost-effective process. Our data showed a high level of deproteinization (38.4 to 0.8%) with immobilized chitinase than the free enzyme (38.4 to 14.3%) of shrimp waste without any additional supplements in a time period of 20 days (Fig. 11). This indicates good potential for application of this immobilized chitinase in the utilization of prawn shell waste.

Fig. 11. Deproteinization of prawn shell waste by immobilized Kurthia gibsonii Mb126 chitinase.
Enzyme immobilization is one of the most important techniques for getting operational stability. In this study, chitinase immobilization of Kurthia gibsonii Mb126 chitinase on chitosan was carried out with immobilization yield of 106%. The specific activity of immobilized chitinase was 4.3-fold greater than that of the free chitinase. The factors affecting the immobilization technique such as concentration of glutaraldehyde and chitinase and immobilization time were studied. The temperature and pH optima of the immobilized chitinase and free enzyme were exactly same, i.e., 7.5 and 40°C respectively. Immobilized Kurthia gibsonii Mb126 chitinase had more superior performance in thermal (92% of its initial activity within 1.5h at 60°C), operational (90% residual activity after 16 cycles), and storage stability (78% of its initial activities at 4°C for 20 months), than free chitinase. Fermentation of prawn shell waste with immobilized chitinase indicated a high level of deproteinization. Protein content was reduced from 38.4 to 0.8% with immobilized chitinase. The high activity of immobilized enzyme in the degradation of prawn shell waste indicates an excellent potential of Kurthia gibsonii Mb126 chitinase on chitosan for managing prawn shell waste. Our study reports suggest that the fermentation of prawn shell waste with immobilized chitinase was simple, eco-friendly, cost-effective, and very useful model for enhancing enzymatic activity, reusability and stability, thus suitable for large scale bioprocess of chitinous waste.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
ETHICS STATEMENT
This article does not contain any studies with human participants or animals performed by any of the authors.
AVAILABILITY OF DATA
All datasets generated or analyzed during this study are included in the manuscript
- Kumar ABV, Tharanathan RN. A comparative study on depolymerization of chitosan by proteolytic enzymes. Carbohydr Polym. 2004;58(3):275-283.
Crossref - Rattanakit N, Plikomol A, Yano S, Wakayama M, Tachiki T. Utilization of shrimp shellfish waste as a substrate for solid-state cultivation of Aspergillus sp. S1-13: Evaluation of a culture based on chitinase formation which is necessary for chitin-assimilation. J Biosci Bioeng. 2002;93(6):550-556.
Crossref - No HK, Hur EY. Control of Foam Formation by Antifoam during Demineralization of Crustacean Shell in Preparation of Chitin. J Agric Food Chem. 1998;46(9):3844-3846.
Crossref - Rueda N, Albuquerque TL, Bartolome-Cabrero R, et al. Reversible Immobilization of Lipases on Heterofunctional Octyl-Amino Agarose Beads Prevents Enzyme Desorption. Molecules. 2016;21(5):646.
Crossref - Tipton K, Boyce S, History of the enzyme nomenclature system. Bioinformatics. 2000;16(1):34-40.
Crossref - Salum TFC, Villeneuve P, Barea B, et al. Synthesis of biodiesel in column fixed-bed bioreactor using the fermented solid produced by Burkholderia cepacia LTEB11. Process Biochem. 2010;45(8):1348-54.
Crossref - Ali B, Khan KY, Majeed H, et al. Production of ingredient type flavoured white enzyme modified cheese. J Food Sci Technol. 2019;56(4):1683-1695.
Crossref - Ahmed AAQ, Babalola OO, McKa T. Cellulase- and xylanase producing bacterial isolates with the ability to saccharify wheat straw and their potential use in the production of pharmaceuticals and chemicals from lignocellulosic materials. Waste Biomass Valor. 2018;9(5):765-775.
Crossref - Shrestha A, Sultana R, Chae JC, Kim K, Lee KJ. Bacillus thuringiensis C25 which is rich in cell wall degrading enzymes efficiently controls lettuce drop caused by Sclerotinia minor. Eur J Plant Pathol. 2015:142(3):577-589.
Crossref - Mathivanan N, Kabilan V, Murugesan K. Purification, characterization, and antifungal activity of chitinase from Fusarium chlamydosporum, a mycoparasite to groundnut rust, Puccinia arachidis. Can J Microbiol. 1998;44(7):646-651.
Crossref - Balasubramanian N, Juliet GA, Srikalaivani P, Lalithakumari D. Release and regeneration of protoplasts from the fungus Trichothecium roseum. Can J Microbiol. 2003;49(4):263-268.
Crossref - Mendonsa ES, Vartak PH, Rao JU, Deshpande MV. An enzyme from Myrothecium verrucaria that degrades insect cuticles for biocontrol of Aedes aegypti mosquito. Biotech Letters. 1996;18(4):373-376.
Crossref - Vyas P, Deshpande M. Enzymatic hydrolysis of chitin by Myrothecium verrucaria chitinase complex, and its utilization to produce SCP. J Appl Gen Microbiol. 1991;37(3):267-275.
Crossref - Aam BB, Heggset EB, Norberg AL, Sorlie M, Varum KM, Eijsink VG. Production of chitooligosaccharides and their potential applications in medicine. Mar Drugs. 2010;8(5):1482-1517.
Crossref - Elnashar MM, Awad GE, Hassan ME, Mohy Eldin MS, Haroun BM, El-Diwany AI. Optimal immobilization of β-galactosidase onto κ-carrageenan gel beads using response surface methodology and its applications. Scientific World Journal. 2014;2014:571682.
Crossref - Elnashar MM, Mostafa H, Morsy NA, Awad GEA. Biocatalysts: isolation, identification and immobilization of thermally stable lipase onto novel supports. Ind Eng Chem Res. 2013;52(42):14760-14767.
Crossref - Danial EN, Elnashar MMM, Awad GEA. Immobilized inulinase on grafted alginate beads prepared by the one-step and the two-steps methods. Ind Eng Chem Res. 2010;49(7):3120-3125.
Crossref - Muzzarelli RAA. Immobilization of enzymes on chitin and chitosan. Enzyme Microb Technol. 1980;2(3):177-184.
Crossref - Juang RS, Wu FC, Tseng, RL. Use of chemically modified chitosan beads for sorption and enzyme immobilization. Adv Environ Res. 2002;6(2):171-177.
Crossref - Kumar MNVR. A review of chitin and chitosan applications. React Func Polym. 2000;46(1):1-27.
Crossref - Yazdani-Pedram M, Retuert J, Quijada R. Hydrogels based on modified chitosan, synthesis and swelling behavior of poly (acrylicacid) graphed chitosan. Macromol. Chem. Phys. 2000;201(9):923-930.
Crossref - Valentina N, Michela S, Elena G. β Galactosidase entrapment in silica gel matrices for a more effective treatment of lactose intolerance. J Molecul Catal B Enzym. 2011;71(1-2):10-15.
Crossref - Rodrigues RC, Virgen-Ortiz JJ, dos Santos JCS, et al. Immobilization of lipases on hydrophobic supports: immobilization mechanism, advantages, problems, and solutions. Biotechnol Adv. 2019;37(5):746-770.
Crossref - Liu D-M, Dong C. Recent advances in nano-carrier immobilized enzymes and their applications. Process Biochem. 2020;92:464-475.
Crossref - Zahirinejad S, Hemmati R, Homaei A, et al. Nano-organic supports for enzyme immobilization: Scopes and perspectives. Colloids Surf. B. 2021;204:111774.
Crossref - Zhang D H, Yuwen LX, Peng LJ. Parametere affecting the performance of immobilized enzyme. J Chem. 2013;2013:946248.
Crossref - Sandhya C, Adapa LK, Nampoothiri KM, Binod P, Szakacs G, Pandey A. Extracellular chitinase production by Trichoderma harzianum in submerged fermentation. J Basic Microbiol. 2004;44(1):49-58.
Crossref - Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem. 1959;31(3):426-428.
Crossref - Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein Measurement with the Folin Phenol Reagent. J Biol Chem. 1951;193(1):265-275.
Crossref - Carrara CR, Rubiolo AC. Immobilization of Galactosidase on Chitosan. J Biotech. 1994;10(2):220-224.
Crossref - Rao MS, Stevens WF. Fermentation of shrimp biowaste under different salt concentrations with amylolytic and nonamylolytic Lactobacillus strains for chitin production. Food Technol Biotechnol. 2006;44:83-87.
- Dumitriu S, Chornet E. Immobilization of xylanase in chitosan-xanthan hydrogels. Biotech Prog. 1997;13(5):539-545.
- Seo DJ, Jang YH, Park RD, Jung WJ. Immobilization of chitinases from Streptomyces griseus and Paenibacillus illinoisensis on chitosan beads. Carbohydr Polym. 2012;88(1):391-394.
Crossref - Bhushan B. Production and characterization of a thermostable chitinase from a new alkalophilic Bacillus sp. BG-11. J Appl Microbiol. 2000;88(5):800-808.
Crossref - Bilal M, Jing Z, Zhao Y, Iqbal HMN. Immobilization of fungal laccase on glutaraldehyde cross-linked chitosan beads and its bio-catalytic potential to degrade bisphenol A. Biocatal Agric Biotechnol. 2019;19:101174.
Crossref - Spagna G, Andreani F, Salatelli S, Ranagnoli D, Pifferi PG. Immobilization of a-Larabinofuranosidase on chitin and chotsan. Process Biochem. 1998;33(1):57-62.
Crossref - Cristovao RO, Silverio SC, Tavares APM, et al. Green coconut fiber: a novel carrier for the immobilization of commercial laccase by covalent attachment for textile dyes decolourization. World J Microbiol Biotechnol. 2012;28(9):2827-2838.
Crossref - Garcia-Galan C, Berenguer-Murcia A, Fernandez-Lafuente R, Rodrigues RC. Potential of different enzyme immobilization strategies to improve enzyme performance. Advanced Synthesis & Catalysis. 2011;353(16):2885-2904.
Crossref - Wang SL, Chio SH. Reversible immobilization of chitinase via coupling to reversibly soluble polymer. Enzyme Microb Technol. 1998;22(7):634-640.
Crossref - Esawy MA, Awad GEA, Wahab WAA, et al. Immobilization of halophilic Aspergillus awamori EM66 exochitinase on grafted k-carrageenan-alginate beads. 3 Biotech. 2016;6(1):29.
Crossref - Sakai K, Yokota A, Kurokawa H, Wakayama M, Moriguchi M. Purification and characterization of three thermostable endochitinases of a noble Bacillus strain, MH-1 isolated from chitin-containing compost. App Environ Microbiol. 1998;64(9):3397-3402.
Crossref - Rehman HU, Aman A, Silipo A, Qader SA, Molinaro A, Ansari A. Degradation of complex carbohydrate: immobilization of pectinase from Bacillus licheniformis KIBGE-IB21 using calcium alginate as a support. Food Chem. 2013;139(1-4):1081-1086.
Crossref - Dincer A, Becerik S, Aydemir T. Immobilization of tyrosinase on chitosan-clay composite beads. Int J Biol Macromol. 2012;50(3):815-820.
Crossref - Yewale T, Singhal RS, Vaidya AA. Immobilization of inulinase from Aspergillus niger NCIM 945 on chitosan and its application in continuous inulin hydrolysis. Biocatal Agric Biotechnol. 2013;2(2):96-101.
Crossref - Sanjay G, Sugunan S. Enhanced pH and thermal stabilities of invertase immobilized on montmorillonite K-10. Food Chem. 2006;94(4):573-579.
Crossref - Prasad M, Palanivelu P. A novel method for immobilization of a thermostable fungal chitinase and the properties of the immobilized enzyme. Biotechnol Appl Biochem. 2014;61(4):441-445.
Crossref - Kidibule PE, Costa J, Atrei A, Plou FJ, Fernandez-Lobato M, Pogni R. Production and characterization of chitooligosaccharides by the fungal chitinase Chit42 immobilized on magnetic nanoparticles and chitosan beads: selectivity, specificity and improved operational utility. RSC Adv. 2021;11(10):5529-5536.
Crossref - Mini KP, Mini KD, Mathew J. Purification and characterization of chitinase enzyme from Kurthia gibsonii Mb126. J Env Biotech Res. 2016;2(1):37-44.
- El-Shora HM, Khalaf SA, El-Sheshtawi SAH. Biochemical Characteristics of Immobilized Chitinase from Alternaria Infectoria Microbiol. 2017;22(1):1-10.
Crossref - Sanchez-Otero MG, Valerio-Alfaro G, Garcia-Galindo HS, Oliart-Ros RM. Immobilization in the presence of Triton X-100: modifications in activity and thermostability of Geobacillus thermoleovorans CCR11 lipase. J Ind Microbiol Biotechnol. 2008;35(12):1687-1693.
Crossref - Keskin S, Ertunga NS, Bektas KI. Covalent Immobilization of α-Amylase from Thermophilic Geobacillus sp. TF14 on Chitosan Beads. Sak J Biol Sci. 2017:21(6):1342-1348.
Crossref - Suresh PV, Chandrasekaran M. Utilization of prawn waste for chitinase production by the marine fungus Beauveria bassiana by solid state fermentation. World J Microbiol Biotechnol. 1998;14:655-660.
Crossref - Yang JK, Shih IL, Tzeng YM, Wang SL. Production and purification of protease from a Bacillus subtilis that can deproteinize crustacean wastes. Enzyme Microb Technol. 2000;26(5-6):406-413.
Crossref
© The Author(s) 2022. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.