


ISSN: 0973-7510
E-ISSN: 2581-690X
Salt-tolerant plant growth-promoting (PGP) actinobacteria play an important role in enhancing plant tolerance to salinity stress and improving crop productivity. Here, 9 morphologically distinct actinobacterial isolates (A-I) were isolated from the rhizosphere soil of Oryza sativa cultivated in the Nalgonda district, Telangana, screened for PGP characteristics (nitrogen fixation, indole-3-acetic acid production (IAA), and phosphate solubilisation) and salt tolerance. Among them, isolate H exhibited superior PGP activity and salt tolerance, growing in media containing up to 10% NaCl. Hence, it was selected for further work. Isolate H produced cellulase, lipase, amylase and showed a positive catalase reaction. It utilised glucose, fructose, galactose, and sucrose, but not maltose. Antibiotic susceptibility analysis indicated that isolate H was partially resistant to penicillin but sensitive to tetracycline, ciprofloxacin, and vancomycin. Molecular identification based on 16S rDNA sequencing (1,356 bp) revealed that isolate H shared 99.78% sequence similarity and 99% query coverage with Streptomyces levis NBRC 15423(T), suggesting that the isolate belongs to the genus Streptomyces. Seed germination efficiency (%) revealed that the 24 hrs culture (T1) of isolate H had a significantly greater effect on rice seed germination than the 48 hrs culture (T2). The germination rate of rice seeds in T1 was 86.67%, followed by T2 (72.00%), the control (62.33%), and distilled water (55.67%), indicating that the 24 hrs grown culture of isolate H may be used to enhance rice seed germination under salt stress. This study provides preliminary evidence of the potential of a Streptomyces levis strain as a PGP bacterium under saline conditions.
Actinobacteria, Plant Growth-Promoting Characteristics, Salt-Tolerant Strain, Antibiotic Susceptibility Analysis, 16S rDNA Sequencing
Numerous microorganisms affected by biotic and abiotic stresses can be found in the rhizosphere or within the zone of influence surrounding plant roots. These include the predominant rhizobacteria known as plant growth-promoting rhizobacteria (PGPR), which are essential for soil health and plant growth because they inhabit the rhizosphere and root surfaces.1 These bacterial groups are important because they participate in a number of biochemical processes in the soil, such as the fixation of atmospheric nitrogen, solubilisation of minerals, such as phosphorus and potassium, production of siderophores that sequester and mobilise iron, and synthesis of plant growth regulators.2 Nitrogenase is a key enzyme involved in biological nitrogen fixation that converts atmospheric nitrogen into forms available to plants, thereby enhancing soil fertility. This is a cost-effective and environmentally friendly alternative to chemical fertilisers.3
Rice (Oryza sativa L.) is a staple food crop in Asia that is affected by various biotic (including rice blast)4 and abiotic stresses.5 Modern agriculture often depends on PGPR with biocontrol traits to improve agricultural production while reducing reliance on chemical fungicides.6
Most of the currently available information on PGPR is related to the phyla Bacteroidetes, Firmicutes, and Proteobacteria. However, very few studies have focused on the PGP characteristics of Actinobacteria.7,8
Actinobacteria (actinomycetes), a phylum of Gram-positive, filamentous bacteria that resemble fungi and possess a high G+C content,9 have the capacity to produce lytic enzymes and secondary metabolites, including PGP substances and antibiotics, making them valuable in agriculture, industry, and medicine.10-12
Salinity stress is one of the major abiotic factors limiting rice productivity worldwide, adversely affecting seed germination, nutrient uptake, plant metabolism, and crop yield.13-15 Under saline conditions, rhizosphere microbial communities play an important role in enhancing plant tolerance and maintaining soil fertility.7 Salt stress mitigation mechanisms employed by PGPR include osmolyte production,16 ACC deaminase activity,17 ion homeostasis and regulation,18 and other general mechanisms associated with salinity stress tolerance.19 Among rhizospheric microorganisms, actinomycetes, particularly Streptomyces species, are recognised for their ability to survive under stressful environmental conditions and produce various PGP metabolites, such as indole-3-acetic acid, siderophores, ammonia, phosphate-solubilising enzymes, and ACC deaminase. These traits can improve nutrient availability, root development, and plant stress tolerance.20-23
In addition, the functional advantages of actinobacteria over other PGPR groups include their ability to produce diverse secondary metabolites,24 extracellular enzymes, stress-resistant spores, and disease-suppressive compounds, as well as their enhanced adaptability to harsh environmental conditions.13,20 Despite several reports on PGPR under salinity stress, studies focusing on salt-tolerant actinomycetes associated with the rice rhizosphere remain limited.22,24,25 Therefore, the isolation and characterisation of salt-tolerant, PGP actinomycetes from the rice rhizosphere are important for identifying potential candidates for sustainable enhancement of rice growth under saline conditions.
Rice cultivation is widely practised in the Nalgonda district of Telangana, and certain agricultural soils are frequently exposed to salinity stress owing to irrigation practices, groundwater quality, and climatic conditions. Hence, the present study emphasises the isolation and identification of salt-tolerant PGP actinobacteria from the rhizosphere of Oryza sativa in the Nalgonda district, evaluation of their PGP traits, and assessment of their potential to promote rice seed germination.
Soil sample collection
Rhizosphere soil samples of Oryza sativa were collected during the crop-growing season from a depth of 10-12.5 cm below the soil surface in Nalgonda district, Telangana, India, at the following GPS coordinates: 17.116634° N, 79.235241° E; 17.115542° N, 79.234527° E; and 17.117239° N, 79.233776° E, using a random sampling method. Three soil samples were collected at each location. Each experiment was performed in triplicate to ensure the reproducibility of the results. After collection, samples were properly sealed in sterile polythene bags and transported to the laboratory.
Isolation of actinomycetes
One gram of soil sample was serially diluted up to 10-6, and 0.1 mL of different dilutions was plated on starch casein agar (g/L): soluble starch, 10; K2HPO4, 2; KNO3, 2; casein, 0.3; MgSO4·7H2O, 0.05; CaCO3, 0.02; FeSO4·7H2O, 0.01; and agar, 15 (pH 7.0 ± 0.2). Starch casein agar contained 80 µg/mL of cycloheximide and 75 µg/mL of nalidixic acid (Himedia, Mumbai, India) to limit fungal and bacterial growth, respectively. A sterilised glass spreader was used to evenly distribute the diluted soil samples on starch casein agar plates. These inoculated plates were incubated at 28 ± 2 °C for 21 days.
Different actinomycete isolates were selected based on specific colony characteristics, including colony colour, texture, size, margin, elevation, aerial mycelium formation, and pigmentation. Colonies were sub-cultured, purified, and stored as slants at 4 °C for further analysis.
PGP tests
The purified actinobacterial isolates were subjected to various PGP tests, including atmospheric nitrogen fixation, IAA production, and phosphate solubilisation. Among all the isolates, a high-potential PGP isolate was selected for further studies.
The nitrogenase activity of the purified actinobacterial isolates was determined by stabbing the isolates separately into Burk’s agar, a nitrogen-free medium (HiMedia, India),26,27 followed by incubation for 3-5 days at 28-30 °C and assessment of growth. The appearance of growth by the isolates indicated a positive result.
IAA production by purified actinobacterial isolates was determined by inoculating them separately into Jensen’s broth (g/L): sucrose, 20; K2HPO4, 1; MgSO4·7H2O, 0.5; NaCl, 0.5; FeSO4, 0.1; NaMoO4, 0.005; and CaCO3,2, supplemented with 2 mg/mL L-tryptophan.28 The test (with the isolate) and control (without the isolate) tubes were kept at 37 °C for 48 hrs with rotation at 125 rpm.29 According to Gordon and Weber, approximately 2 mL of test and control solutions were centrifuged for 1 min at 18,800 x g after the incubation period. Then, 1 mL aliquots of both test and control supernatants were separately mixed with 2 mL of Salkowski’s reagent (150 mL of concentrated H2SO4, 250 mL of distilled water, and 7.5 mL of 0.5 M FeCl3·6H2O) and incubated for 20 min at 28-30 °C. IAA production was confirmed by a change in the broth colour to pink-red compared with the control.30
To determine phosphate solubilisation activity of purified actinobacterial isolates, they were grown on Pikovskaya’s agar.31 Pikovskaya’s agar contained (g/L): dextrose, 10; yeast extract, 0.5; (NH4)2SO4, 0.5; Ca3(PO4)2, 0.5; KCl, 0.2; MgSO4, 0.1; FeSO4, 0.0001; MnSO4, 0.0001; and agar, 15. After inoculation, Petri plates were incubated at 28-30 °C for 7 days. A clear halo zone surrounding the isolates indicated a positive phosphate solubilisation test (conversion of insoluble phosphate into its soluble form).
Effect of % NaCl on growth of actinobacterial isolates
Purified actinobacterial isolates were subjected to NaCl (0%-12%, at 1% intervals) to determine their tolerance levels and the amount of NaCl required for optimal growth. The growth of the isolates was evaluated at 24 hrs and 48 hrs based on measurements of optical density (at 600 nm) using a colorimeter.
Enzyme activities
Based on the PGP traits and salinity tolerance described above, a high-potential PGP isolate (showing high salinity tolerance) was selected from among the purified actinobacterial isolates. Subsequently, it was tested for the production of different hydrolytic enzymes, including cellulase, lipase, and amylase, on carboxymethyl cellulose (CMC), Tween 80 lipase, and starch agar plates.
The cellulase activity of the selected PGP isolate was investigated by inoculating the organism on CMC agar, where 1% CMC was used as the soluble cellulose derivative and substrate. CMC hydrolysis was detected by flooding the agar plate with 1% Congo red dye solution. After 15 min, the dye solution was poured off, and the plate was further flooded with 0.1% NaCl for 15 min and observed for the formation of a clear zone. The undegraded cellulose portions were stained red, whereas clear zones appeared around the colonies where the CMC was degraded.32
The amylase activity of the selected PGP isolate was determined using the clear-zone method on starch agar plates. The plate was flooded with iodine solution for 1 min after incubation for 72 hrs. Formation of a clear zone surrounding the culture indicated a positive starch hydrolysis test, whereas the absence of a zone indicated a negative test.33
The selected PGP isolate was tested for lipase production on 0.1% Tween 80 lipase agar plates.34,35 The formation of a clear zone around the isolate indicated a positive lipase test, whereas the absence of a zone indicated a negative test.
Effect of temperature and pH on growth of the PGP isolate
The selected high-potential PGP isolate was subjected to different temperatures (room temperature (28-30 °C), 28 °C, 37 °C, and 45 °C) to optimise its growth and evaluate pigment production. Different pH conditions (pH 6-9, at intervals of 1 pH unit) were used to identify the optimal pH for growth. Isolate growth was evaluated at 24 hrs and 48 hrs based on measurements of the optical density at 600 nm using a colorimeter.
Biochemical characterisation of the PGP isolate
Different carbohydrate utilisation tests (glucose, fructose, galactose, maltose, and sucrose) were performed for the selected PGP isolate to determine its ability to utilise specific carbohydrates according to the carbohydrate fermentation protocol.36 Phenol red carbohydrate broth was inoculated with the selected PGP isolate, and a colour change from orange-red to yellow after 24 hrs of incubation indicated a positive test (acid production). This test was performed along with a control (without the isolate) for comparison.
To perform the catalase test, a small amount of the selected PGP organism was placed onto a microscope slide without any agar after 24 hrs of growth in nutrient broth. Using a dropper or Pasteur pipette, one drop of 3% H2O2 was placed on the organism on the microscope slide. The formation of bubbles against a dark background, which enhances visibility, indicated a positive result.37
Antimicrobial susceptibility test of the PGP isolate
The antimicrobial susceptibility test of the selected PGP isolate was performed according to CLSI guidelines38 to determine the sensitivity of the organism to penicillin (10 µg), tetracycline (30 µg), ciprofloxacin (5 µg), and vancomycin (30 µg). This test was performed by placing the respective antibiotic discs on the culture and measuring zones of inhibition using the Kirby–Bauer disc diffusion method.39
Molecular characterisation of the PGP isolate
The selected PGP isolate was characterised at the molecular level by isolating its DNA through incubation of a bacterial colony in 50 µL of molecular biology-grade distilled water at 96 °C for 10 min in a thermal cycler. Polymerase chain reaction (PCR) was then performed using the extracted DNA under the following conditions: initial denaturation at 94 °C for 5 min, followed by 30 cycles of denaturation at 94 °C for 30 sec, annealing at 54 °C for 30 sec, and extension at 72 °C for 2 min, followed by a final extension at 72 °C for 5 min to amplify the 16S rDNA gene of the isolate using universal bacterial forward and reverse primers (27F and 1525R).40 The PCR product was then subjected to 16S rDNA sequencing, which was outsourced to the National Collection of Industrial Microorganisms, Pune. The 16S rDNA sequence of the isolate was uploaded to the EzBioCloud database and compared with the 16S rDNA sequences available in this database to identify the isolate based on sequence similarity values. Phylogenetic analysis of the isolate was performed using the ClustalW alignment program in MEGA 11.41
A phylogenetic tree was constructed using the neighbour-joining method with the Kimura 2-parameter model, and bootstrap analysis was performed with 1,000 replications to assess branch support values.
Effect of strain H culture on the growth of rice seeds under salt stress
Seed germination experiments were conducted to study the effects of isolate H on rice seed growth under salt stress. Initially, rice seeds were surface-sterilised in 70% ethanol, followed by treatment with sodium hypochlorite solution, and rinsed thoroughly with sterile distilled water before treatment. The bacterial inoculum was prepared at an approximate density of 108 colony-forming units/mL. Sterilised rice seeds were immersed in distilled water, a nutrient medium consisting of 5% NaCl without isolate H (control), a nutrient medium containing 5% NaCl with isolate H grown for 24 hrs (T1), and a nutrient medium containing 5% NaCl with isolate H grown for 48 h (T2) for 2 hrs each. The rice seeds were then placed separately in four petri dishes containing moist tissue paper and sprinkled daily with sterile distilled water. Seed germination analysis was performed under controlled incubation conditions at 28 ± 2 °C. The percentage germination of rice seeds was observed in triplicate after 7 days. All experiments were conducted in triplicate.
The percentage germination of rice seeds was calculated using the following formula:
% Seed germination = [Number of seeds germinated / Total number of seeds used for germination] × 100
Rice seed germination data were statistically analysed using one-way analysis of variance to compare the distilled water, control, T1, and T2 treatments. Statistical analysis was performed using SPSS version 25.0, and differences were considered statistically significant at P < 0.05. Post-hoc analysis was conducted using Tukey’s honest significant difference test to determine significant differences among treatment means.
Isolation of actinomycetes
Morphologically, nine different actinomycete colonies (serially labelled A-I) were isolated and purified on starch casein agar from rhizosphere soil samples of Oryza sativa. All nine isolates were subjected to PGP tests to determine their PGP potential.
Members of Streptomyces and related actinomycetes play important ecological roles in organic matter decomposition, nutrient recycling, production of bioactive metabolites, and enhancement of plant growth under stress conditions. Their adaptation to saline environments may improve plant tolerance to salt stress through mechanisms, such as phytohormone and siderophore production, phosphate solubilisation, and osmotic stress regulation. A similar ecological significance and functional diversity of actinomycetes have been previously reported.20,42-44 Three isolates, HB6J2, HB8P1, and HB4N3, were reported to have significant salt tolerance and were isolated from 18 soil samples collected from saline soils in Haryana State.45
PGP tests
Among the 9 isolates (A-I) tested, isolate H grew effectively on Burk’s agar, indicating its nitrogen-fixing capacity (Figure 1). The conclusion regarding nitrogen fixation was based on growth in a nitrogen-free medium and associated PGP traits. However, additional direct assays, such as the acetylene reduction assay and nif-gene detection, would further confirm the nitrogen-fixing ability. Compared with the other isolates purified in this study, isolate H also showed IAA production when the broth changed to a reddish colour after addition of salkowski reagent (Figure 2). Many PGP actinomycete isolates produce IAA, which is responsible for enhancing the number of adventitious roots, thereby helping plants absorb more water and nutrients and secrete increased amounts of root exudates that are beneficial to bacteria.46

Figure 1. Growth of isolate H on Burk’s agar

Figure 2. IAA production by isolate H
A clear halo zone formed around isolate H on Pikovskaya’s agar, indicating positive phosphate solubilisation after 7 days of incubation (Figure 3) compared with that observed for the other eight isolates purified in this investigation. In a study of 98 rhizospheric actinomycetes isolated from different wheat and tomato fields in Punjab, Pakistan, only 6 Streptomyces isolates were positive for phosphate solubilisation.47
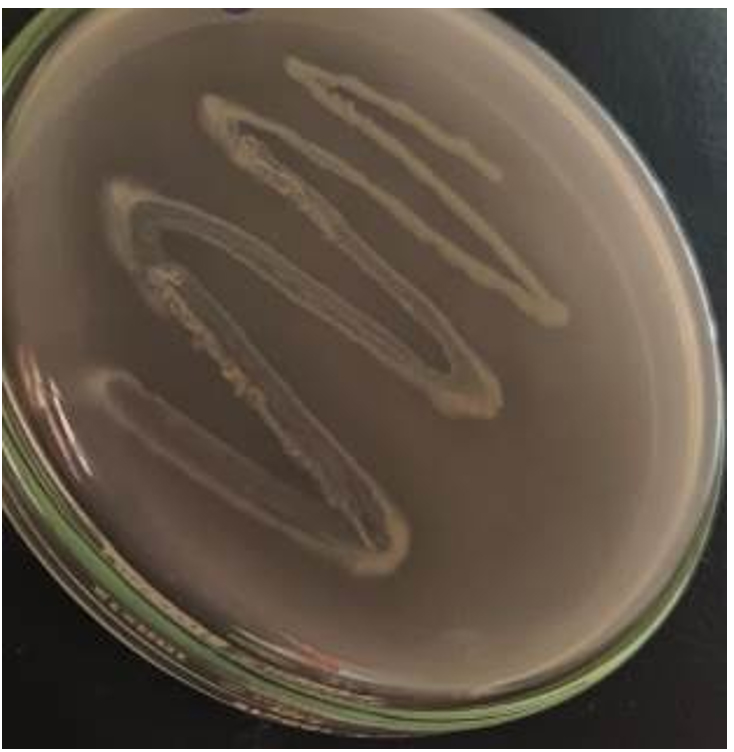
Figure 3. Phosphate solubilisation on Pikovskaya’s agar by isolate H
Effect of % NaCl on growth of actinobacterial isolates
Eight actinobacterial isolates exhibited lower salt tolerance (below 2% NaCl) than that of isolate H, which grew in a medium containing up to 10% NaCl. However, the optimum growth of isolate H was recorded at 2% NaCl. PGPR can reduce the harmful effects of increased salt concentrations on plants and enhance their development in two ways: first, by regulating plant response systems during salt exposure and, second, by producing anti-stress molecules.48
Because isolate H exhibited promising PGP activities and comparatively better salinity tolerance than the other isolates purified in this study, it was chosen for further studies.
Enzyme activities
Clear-zone formation around the culture was observed for isolate H on CMC, starch, and Tween 80 lipase agars, indicating positive cellulase, amylase, and lipase activities, respectively (Figure 4).

Figure 4. Different enzyme activities of isolate H: (a) cellulase activity on carboxymethyl cellulose (CMC) agar; (b) amylase activity on starch agar; and (c) lipase activity on Tween 80 lipase agar
Cellulase production by Streptomyces spp. helps degrade complex cellulose materials present in the rhizosphere, thereby releasing nutrients for plant uptake and supporting microbial colonisation around plant roots.43 Production of extracellular enzymes by actinomycetes also improves rhizosphere colonisation efficiency by enabling microorganisms to utilise diverse organic substrates released from plant roots and soil debris.43
Amylase activity contributes to the breakdown of starch and other polysaccharides into simpler sugars, which enhances nutrient cycling, microbial survival in the rhizosphere, and plant growth.42 Protease production may assist in the degradation of proteinaceous organic matter and suppress certain phytopathogens through the hydrolysis of pathogenic proteins, thereby indirectly promoting plant health.20
Effect of temperature and pH on growth of isolate H
Isolate H showed good growth at room temperature but was not able to produce pigment under these conditions. It produced a darker brown pigment at 28 °C and 37 °C. However, pigment production was lower at 45 °C (Figure 5). The growth of isolate H was observed up to pH 8, but the optimum pH was 7.

Figure 5. Effect of temperature on growth and pigment production by isolate H
Biochemical characterisation
Different carbohydrate utilisation tests performed with isolate H showed a positive reaction (broth colour changed to yellow) for all carbohydrates tested (glucose, fructose, galactose, and sucrose), except maltose (Figure 6). This test was compared with the control (without isolate H). Isolate H tested positive for catalase activity (Figure 7).

Figure 6. Different carbohydrate utilisation tests were performed with isolate H

Figure 7. Catalase test of isolate H
Antimicrobial susceptibility test
Isolate H was partially resistant to penicillin (10 µg), whereas it was sensitive to tetracycline (30 µg), ciprofloxacin (5 µg), and vancomycin (30 µg; Figure 8). Actinobacteria enhance plant growth not only through the production of extracellular enzymes, siderophores, and phytohormones but also through the production of antagonistic metabolites in soil ecosystems.20
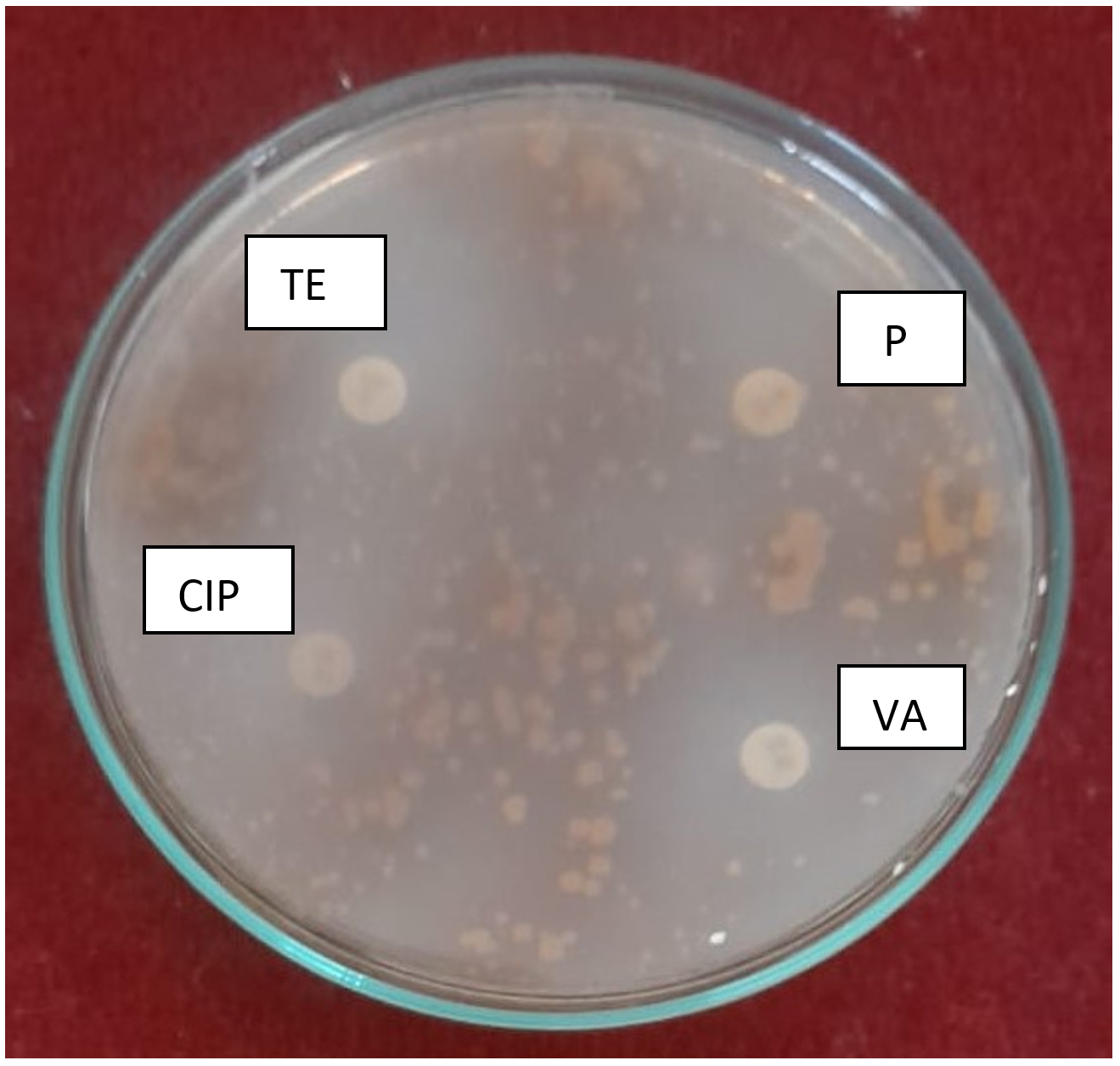
Figure 8. Antibiotic susceptibility test of isolate H
Note: P = penicillin (10 µg); TE = tetracycline (30 µg); CIP = ciprofloxacin (5 µg); and VA = vancomycin (30 µg)
Molecular identification
The 16S rDNA sequence (1,356 bp) of isolate H obtained after outsourcing was uploaded to the EzBioCloud database and compared with the 16S rDNA sequences of other strains present in the database. It showed 99.78% similarity to Streptomyces levis NBRC 15423(T) with 99% query coverage (Figure 9). Streptomyces is the largest genus of actinobacteria, members of which are commonly found in soil and contribute to the decomposition of organic matter, thereby promoting crop production. They are involved in the production of extracellular hydrolytic enzymes and antifungal compounds; therefore, they play a role in biocontrol, particularly against phytopathogens such as those causing rice blast disease.49,50 Some reports have also highlighted their role in improving crop tolerance to salt stress.51

Figure 9. Phylogenetic tree of isolate H constructed using the neighbour-joining method. Bar = 0.01 nucleotide substitutions per site
Effect of isolate H on rice seed germination under salt stress
Rice seed germination results indicated (Table) that the 24 hrs grown isolate H culture (T1) in nutrient medium in the presence of 5% NaCl showed a higher rice seed germination percentage (86.67%) than that of the 48 hrs grown isolate H culture (T2), which showed a germination percentage of 72.00%. The control condition (nutrient medium without isolate H containing 5% NaCl) showed a lower seed germination percentage (62.33%) than that of T2, and treatment with distilled water resulted in the lowest seed germination rate (55.67%). One-way analysis of variance revealed significant differences in rice seed germination among the treatments (P < 0.05). Tukey’s honest significant difference post-hoc test showed that the T1 treatment resulted in significantly higher seed germination than that of the T2, control, and distilled water treatments. The difference between T1 and T2 in seed germination may be associated with variations in the bacterial growth phase, cell viability, or nutrient availability. This report indicated that the more metabolically active 24 hrs grown culture may have produced higher levels of PGP substances than those of the 48 hrs grown culture. Streptomyces spp. are known to improve plant growth under saline conditions through multiple mechanisms, including phytohormone production, nutrient mobilisation, siderophore production, extracellular enzyme secretion, and improved rhizosphere colonisation. These activities may help plants tolerate osmotic and ionic stresses caused by salinity.20 Similar observations were reported by Gopalakrishnan et al.22 and Sreevidya et al.,44 who demonstrated the role of Streptomyces spp. in improving seed germination, enhancing root development and plant growth, and increasing salinity stress tolerance, which may be attributed to their PGP activities, including phytohormone production and stress tolerance mechanisms.
Table. Effect of isolate H on rice seed germination percentage
| No. | Rice seed treatment | Seed germination (%) in replicates | Mean ± Standard deviation (SD) seed germination (%) | Group | ||
|---|---|---|---|---|---|---|
| 1 | T1 (24-hour-grown strain H in nutrient medium with 5% NaCl) | 88 | 87 | 85 | 86.67 ± 1.53 | a |
| 2 | T2 (48-hour-grown strain H in nutrient medium with 5% NaCl) | 74 | 72 | 70 | 72.00 ± 2.00 | b |
| 3 | Control (nutrient medium containing 5% NaCl without isolate H) | 64 | 62 | 61 | 62.33 ± 1.53 | c |
| 4 | Distilled water | 58 | 54 | 55 | 55.67 ± 2.08 | d |
This report provides information on the purification of a salt-tolerant Streptomyces levis strain (isolate H) from Nalgonda district, Telangana, India. The isolate exhibits promising PGP traits that enhance rice seed germination under saline conditions. This report highlights that the 24 hrs grown culture of isolate H (T1) had a significant impact on the germination of rice seeds compared with the 48 hrs grown culture (T2), the control treatment, and distilled water. Further research on this isolate is required to enhance rice crop production.
ACKNOWLEDGMENTS
The authors are thankful to Mahatma Gandhi University, Nalgonda, Telangana, India, for providing the facilities for this research.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Johri BN, Sharma A, Virdi JS. Rhizobacterial diversity in India and its influence on soil and plant health. Adv Biochem Eng Biotechnol. 2003;84:49-89.
Crosseref - Kloepper JW. Current status and future trends in Biocontrol Research and Development in The US. Int symp on clean agri. Sapporo: DECD; 1997:49-52.
- Pareek RP, Chandra R, Pareek N. Role of BNF technology in sustainable agriculture—Proceedings of National symposium. In: Masood A, Chaturvedi SK, Gurha SN, eds. Ind Inst of Pulses Res. 2002:33-42.
- Asibi AE, Chai Q, Coulter JA. Rice blast: A disease with implications for global food security. Agronomy. 2019;9(8):451.
Crossref - Chaiharn M, Chunhaleuchanon S, Lumyong S. Screening siderophore producing bacteria as potential biological control agent for fungal rice pathogens in Thailand. World J Microbiol Biotechnol. 2009;25(11):1919-1928.
Crossref - Jones SE, Elliot MA. Streptomyces exploration: Competition, volatile communication and new bacterial behaviours. Trends Microbiol. 2017;25(7):522-531.
Crossref - Bhattacharyya PN, Jha DK. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J Microbiol Biotechnol. 2012;28(4):1327-1350.
Crossref - Jog R, Nareshkumar G, Rajkumar S. Plant growth promoting potential and soil enzyme production of the most abundant Streptomyces spp. from wheat rhizosphere. J Appl Microbiol. 2012;113(5):1154-1164.
Crossref - Omar AF, Abdelmageed AHA, Al-Turki A, Abdelhameid NM, Sayyed RZ, Rehan M. Exploring the plant growth-promotion of four Streptomyces strains from rhizosphere soil to enhance cucumber growth and yield. Plants. 2022;11(23):3316.
Crossref - Anwesha G, Chowlani M, Saikia R, Mandal DS. Actinobacteria: Diversity and Biotechnological Applications. In Book: Recent Advancements in Microbial Diversity; 2020:217-231.
Crossref - Harir M, Bendif H, Bellahcene M, Fortas Z, Pogni R. Streptomyces secondary metabolites. Basic Biol Appl Actinob. 2018;6:99-122.
Crossref - Ferraiuolo SB, Cammarota M, Schiraldi C, Restaino OF. Streptomycetes as platform for biotechnological production processes of drugs. Appl Microbiol Biotechnol. 2021;105(2):551-568.
Crossref - Kanchan Y, Pooja Y, Kunal S. Microbial-Mediated Molecular Mechanisms to Cope Up Salinity and Drought Stress in Plants. Microbial Based Land Restoration Handbook. 1st ed (2). CRC Press. 2022.
Crossref - Djebaili R, Pellegrini M, Smati M, Del Gallo M, Kitouni M. Actinomycete strains isolated from saline soils: Plant-growth-promoting traits and inoculation effects on Solanum lycopersicum. Sustainability. 2020;12(11):4617.
Crossref - Vurukonda SSKP, Vardharajula S, Shrivastava M, Anil S. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol Res. 2016;184:13-24.
Crossref - Etesami H, Beattie GA. Mining halophytes for plant growth-promoting halotolerant bacteria to enhance the salinity tolerance of non-halophytic crops. Front Microbiol. 2018;9:148.
Crossref - Orozco-Mosqueda MC, Glick BR, Santoyo G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol Res. 2020;235:126439.
Crossref - Ilangumaran G, Smith DL. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front Plant Sci. 2017;8:1768.
Crossref - Nadeem SM, Ahmad M, Zahir ZA, Javaid A, Ashraf M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol Adv. 2014;32(2):429-448.
Crossref - Palaniyandi SA, Yang SH, Zhang L, Suh JW. Effects of Actinobacteria on plant disease suppression and growth promotion. Appl Microbiol Biotechnol. 2013;97(22):9621-9636.
Crossref - Jog R, Pandya M, Nareshkumar G, Rajkumar S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiol (Reading). 2014;160(4):778-788.
Crossref - Gopalakrishnan S, Vadlamudi S, Apparla S, et al. Evaluation of Streptomyces spp for their plant-growth-promotion traits in rice. Can J Microbiol. 2013;59(8):534-539.
Crossref - Silva GC, Kitano IT, Ribeiro IAF, Lacava PT. The potential use of actinomycetes as microbial inoculants and biopesticides in agriculture. Front Soil Sci. 2022;2:833181.
Crossref - Qin S, Xing K, Jiang JH, Xu LH, Li WJ. Biodiversity, bioactive natural products and biotechnological potential of plant-associated endophytic Actinobacteria. Appl Microbiol Biotechnol. 2011;89(3):457-473.
Crossref - Egamberdieva D, Wirth S, Jabborova D, Rasanen LA, Liao H. Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J Plant Interact. 2017;12(1):100-107.
Crossref - Burk D, Lineweaver H. The influence of fixed nitrogen on Azotobacter. J Bacteriol. 1930;19(6):389-414.
Crossref - Park M, Kim C, Yang J, et al. Isolation and characterization of diazotrophic growth promoting bacteria from rhizosphere of agricultural crops of Korea. Microbiol Res. 2005;160(2):127-133.
Crossref - Bric JM, Bostock RM, Silverstone SE. Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl Environ Microbiol. 1991;57(2):535-538.
Crossref - Rahman A, Sitepu IR, Tang SY, Hashidoko Y. Salkowski’s reagent test as a primary screening index for functionalities of rhizobacteria isolated from wild dipterocarp saplings growing naturally on medium-strongly acidic tropical peat soil. Biosci Biotechnol Biochem. 2010;74(11):2202-2208.
Crossref - Gordon SA, Weber RP. Colorimetric estimation of indole acetic acid. Plant Physiol. 1951;26(1):192-195.
Crossref - Goenadi DH, Siswanto, Sugiarto Y. Bioactivation of poorly soluble phosphate rocks with a phosphorus-solubilizing fungus. Soil Sci Soc Am J. 2000;64(3):927-932.
Crossref - Sazci A, Erenler KK, Radford A. Detection of cellulolytic fungi by using Congo red as an indicator: A comparative study with the dinitrosalicyclic acid reagent method. J Appl Bacteriol. 2008;61(6):559-562.
Crossref - Geetha K, Venkatesham E, Hindumathi A, Bhadraiah B. Isolation, screening and characterization of plant growth promoting bacteria and their effect on Vigna Radita (L.) R. Wilczek. Int J Curr Microbiol Appl Sci. 2014;3(6):799-809.
- Sipiczki G, Micevic SS, Kohari-Farkas C, et al. Effects of olive oil and Tween 80 on production of lipase by Yarrowiayeast strains. Processes. 2024;12(6):1206.
Crossref - Ramnath L, Sithole B, Govinden R. Identification of lipolytic enzymes isolated from bacteria indigenous to Eucalyptus wood species for application in the pulping industry. Biotechnol Rep (Amst). 2017;15:114-124.
Crossref - American Society for Microbiology (ASM). Protocols: Carbohydrate Fermentation by Bacteria. 2012.
- Duke PB, Jarvis JD. The catalase test—A cautionary tale. Med Lab Technol. 1972;29(2):203-204.
- Clinical and Laboratory Standards Institute. 33rd ed. CLSI supplement M100. CSLI. Performance Standards for Antimicrobial Susceptibility Testing. Wayne, PA; 2023.
- Bauer AW, Kirby WMM, Sherris JC, Turck M. Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol. 1966;45(4):493-496.
- Yang YW, Chen MK, Yang BY, et al. Use of 16S rRNA gene-targeted group-specific primers for real-time PCR analysis of predominant bacteria in mouse feces. Appl Environ Microbiol. 2015;81(19):6749-6756.
Crossref - Tamura K, Stecher G, Kumar S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38(7):3022-3027.
Crossref - Goodfellow M, Williams ST. Ecology of actinomycetes. Annu Rev Microbiol. 1983;37:189-216.
Crossref - Barka EA, Vatsa P, Sanchez L, et al. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol Mol Biol Rev. 2015;80(1):1-43. Erratum in: Microbiol Mol Biol Rev. 2016;80(4):iii.
Crossref - Sreevidya M, Gopalakrishnan S, Kudapa H, Varshney RK. Exploring plant growth-promotion actinomycetes from vermicompost and rhizosphere soil for yield enhancement in chickpea. Braz J Microbiol. 2016;47(1):85-95.
Crossref - Sharma A, Dev K, Sourirajan A, Choudhary M. Isolation and characterization of salt-tolerant bacteria with plant growth-promoting activities from saline agricultural fields of Haryana, India. J Genet Eng Biotechnol. 2021;19(1):99.
Crossref - El-Tarabily KA. Promotion of tomato (Lycopersicon esculentum Mill.) plant growth by rhizosphere competent 1-aminocyclopropane-1-carboxylic acid deaminase-producing streptomycete actinomycetes. Plant Soil. 2008;308(1):161-174.
Crossref - Anwar S, Ali B, Sajid I. Screening of rhizospheric actinomycetes for various in vitro and in-vivo plant growth promoting (PGP) traits and for Agroactive compounds. Front Microbiol. 2016;7.
Crossref - Fouda A, Hassan SED, Eid AM, El-Din Ewais E. The interaction between plants and bacterial endophytes under salinity stress. In: Mיrillon JM, Ramawat KG, eds. Endophytes and Secondary Metabolites. Cham: Springer International Publishing; 2019:1-17
Crossref - Gonzalez-Franco AC, Robles-Hernandez L. Actinomycetes as biological control agents of phytopathogenic fungi. Technosci Chihuahua. 2009;3(2):64-73.
- Dave A, Ingle S, Perveen K, Bukhari NA, Sayyed R, Mastinu A. Harnessing plant growth-promoting and wilt-controlling biopotential of a consortium of actinomycetes and mycorrhizae in pigeon pea. J Phytopathol. 2024;172(5):e13399.
Crossref - Ravi Kumar K, Awasthi G, Sankhla MS, et al. Current Advances in Microbial Consortia of Actinobacteria and PGPR to improve crop resilience and soil fertility in salinity degraded soils. Ind Biotechnol. 2025;22(3).
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.