


ISSN: 0973-7510
E-ISSN: 2581-690X
Pectobacterium wasabiae is a dreadful causal agent of potato soft rot. It relied mainly on the production a wide range of plant cell wall-degrading enzymes to cause disease. Of the different enzymes, the endo-b-1,4-glucanase may be important in pathogenesis, being responsible for hydrolyzing the cellulose. In the process of pathogen infection, cellulase secreted by the pathogen is major determinant of pathogenicity, which played an important role in softening and decomposition of cell wall. But so far, the related research very rarely. However, most characteristic of the cellulase in the soft rot bacteria was not clear so far. Here, a putative endo-b-1,4-glucanase gene from P. wasabiae SCC3193 was previously synthesized according the bias codon of yeast, heteroexpressed in Pichia pastoris, and further investigated for its characteristic. Pectobacterium wasabiae SCC3193 is a genetically well characterized model in soft rot research. SDS-PAGE analysis showed that the recombinant endo-b-1,4-glucanase (PcegAI) produced in P. pastoris has a molecular mass of approximately 30kDa. When CMC was used as substrate, it exhibited maximal activity at pH 4.0 with an optimum temperature of 40°C, and stable at pH 5-8. The Km and specific activity values for CMC are 64.5 mM and 7 ×103 U mg-1 respectively. Most metallic ions had no influence on the activity of endo-b-1,4-glucanase at a concentration of 1 mM except for Cu2+ and Mn2+. These characteristics suggested that PcegAI may play a major role in degrading of cellulose polymers. The high activity of this enzyme also has potential in biomass opening, such as the bioconversion of lignocellulosic materials and bioethanol production.
endo-b-1,4-glucanase; Pectobacterium wasabiae; heterologous expression; Pichia pastorias; enzymatic characteristics.
Soft rot disease is economically one of the most devastating bacterial diseases affecting plants worldwide. There are large quantities of a wider range of cell wall-degrading enzymes causing soft-rot1. Due to the capacity to synthesize and secrete several extracellular enzymes including pectate lyase, pectin lyase, polygalacturonase, cellulase, protease, and elicitor protein harpin, the soft rot bacteria Pectobacterium is responsible for significant, global economic losses of crops and ornamental plants, both in the field and in storage. Pectate lyase, pectin lyase, and endopolygalaturonase cause cell separation and changes in cell membrane permeability, and cellulase and protease can augment the action of the pectinases, thus contributing to virulence2. Emphasis is placed on the role of cellulase in attacking plant cell wall and membrane components leading to plant tissue maceration.
It is generally known that cellulose is the most abundant component of plant biomass, accounting for about half of the organic material in the biosphere and cellulose degrading enzymes are widely spread in nature. As the major polysaccharide constituent of plant cell walls, cellulose consists of long chains of b-1,4-linked glucose units and water insoluble form that cannot be utilized directly by most organisms3. It attracted considerable research attention, and its degradation has become a problem of fundamental interest due to the high degree of crystallinity of the substrate4. As we all know, extensive hydrolysis of crystalline cellulose requires the synergistic action of different types of cellulases. It is currently accepted that through synergetic action with three different types of cellulolytic enzymes, it could convert the cellulosic materials to glucose, which is further turned into bioethanol5. Among them, endo-b-1,4-glucanase catalyzes the first step of plant polymer hydrolysis by randomly breaking down the internal b-1,4-glycoside linkages6-8.
Many prokaryotic and eukaryotic organisms, including bacteria, fungi, termites, and plants, express genes encoding endo-b-1,4-glucanase. The development and utilization of glucanase possess great significance to solve agricultural and industrial raw materials, alcohol fermentation9, energy crisis, waste management, animal feed production10 and other issues. Consequently, it has very broad application prospects, and there has been a rapid growth in demand for endo-b-1,4-glucanase, especially those that possess particular biochemical properties, e.g., neutral characteristics11,12 or high specific activities13-15.
The complete and annotated genome sequence of the plant-pathogenic enterobacterium has been reported previously. Pectobacterium wasabiae SCC3193 is a kind of strong pathogenic gram-negative bacteria which causes soft-rot and blackleg in a variety of important crops16. In this research, we report the high expression of the putative endoglucanases gene from P. wasabiae SCC3193 in Pichia pastorias. P. pastoris is an ideal exogenous gene expression system, which can use methanol as a sole carbon source and induce efficient secretion and expression of exogenous proteins.
The study emphasis of this paper is endo-b-1,4-glucanase. Firstly, through the homology comparison, we find a gene similar to endo-b-1,4-glucanase.In order to research on the function of the gene, we synthesized the modified gene, the original codon sequence in which was instead with yeast bias. After protein properties research, we convinced that it is indeed a endo-b-1,4-glucanase gene, and infer the genes involved in cell wall degradation. In addition, the enzyme is high specific activity , acid resistance and suitable for industrial fiber processing. Some properties of the enzyme were investigated and discussed.
Chemicals, Plasmids, Strains, and Culture Medium
CMC was purchased from Sigma Chemical Co., Ltd (St Louis, USA). Enzymes (Taq DNA polymerase and T4 DNA ligase) were purchased from Sangon (Shanghai, China). Protein markers and restriction endonucleases were purchased from Takara (Dalian, China). All other chemicals were of analytical grade and commercially available. Pichia pastoris strain GS115 and plasmid pPIC9 K were purchased from Invitrogen Corporation (San Diego, CA, USA). Pichia pastoris GS115 was used for heterologous expression of the target gene. Expression of inserts was controlled by the methanol-inducible alcohol oxidase gene (AOX1) promoter. P. pastorisGS115 was cultivated at 30°C in YPD medium for propagation. BMGY and BMMY liquid medium were used for P. pastoris recombinants growth.
Chemical synthesis of PceglAI gene, vector construction and transformation
Based on the amino acid sequences of the annotated putative xyloglucan-specific endo-b-1,4-glucanase A (GenBank accession no. YP_006283904) from Pectobacterium wasabiae SCC3193, we designed nineteen oligonucleotide primers to synthesize the endo-b-1,4-glucanase gene . In the renamed synthesized gene PceglAI, the codons for the signal peptides have been deleted and replaced with the condons for sequences KREAEA recognized by Kex2 protease. In addition, condons for six polyhistidine tag were added at the end of gene. Appropriate mutations of all restriction sites in the gene were created but Xho I and Not I were added upward and downward the gene (Fig.1a). The putative endo-b-1,4-glucanase gene PceglAI was chemically synthesized via the PCR-based two-step DNA synthesis (PTDS) method17. The condition of PCR that synthesizes the PceglAI gene was as follows: first 94°C 30 s, 65°C 40 s, 72°C 1 min 30 s; 5 cycles and then 94°C 30 s, 70°C 40 s, 72°C 1 min 30 s; 25 cycles. After Xho I and Not I digestion, the gene was inserted into a modified yeast vector pYPX88 Xiong to form PceglAI expression vector pYR1725. Compared to the original vector pPIC9K, the new vector pYPX88 containing a synthesized á-mating factor prepro-secretion signal MF4I (GenBank accession no. AY145833). Moreover, the Bam HI site between the AOX1 promoter and signal was deleted so that the promoter is linked with the signal peptide directly. The resulting plasmid was linearized with Sal I, and transformed into P. pastoris by electroporation (Bio-Rad Genepulser, Hercules, USA). The transformants were screened on selective plates (SD, 0.8 M sorbitol, 2% glucose, and 2% agar).
Expression of recombinant in P. Pastoris
The transformat strains were grown at 28°C and 250 rpm for 24 h in shaking flasks containing 25 ml liquid medium of BMGY (SD, 1% yeast extract, 2% peptone, 1.34% YNB, 0.000004% biotin, 1% glycerol). The cells were harvested by centrifugation at 10,000 rpm for 7 min. After rinsing with sterilized water, the cells were adjusted to a volume of liquid BMMY (SD, 0.000004% biotin) with OD600=1.0. The cells were incubated at 28°C for 3 days with constant shaking 250 rpm, and 100% methanol was added to a final concentration of 1% every 24 h to induce enzyme expression. The recombinant enzymes were secreted into the culture medium, collected by centrifugation, and subjected to endo-b-1,4-glucanase activity assay. Aliquots of culture supernatant were sampled every day followed by measuring the enzyme activity.
Purification of enzyme and protein analysis
The culture was centrifuged and the supernatant was used as the endo-b-1,4-glucanase source. The crude endo-b-1,4-glucanase was precipitated with (NH4)2SO4 (80%). Desalting was performed on Sephadex G-15 column (Sigma-Aldrich), followed by loading onto Ni2+ -NTA agarose affinity column (Sigma-Aldrich) according to manufacturer’s instructions. The endo-b-1,4-glucanase obtained after purification were analyzed on 12 % (w/v) SDS-PAGE using the Mini-2 protein gel electrophoresis system (Bio-Rad Lab., Hercules, USA). The separated protein bands were stained with 0.2 % Coomassie brilliant blue R-250. Protein concentrations were determined by the Bradford method, using bovine serum albumin as standard. Deglycosylation of the natural endo-b-1,4-glucanase was performed using an N-glycosidase F deglycosylation kit (Roche, Penzberg, Germany) according to the manufacturer’s instructions. For deglycosylation of the recombinant endo-b-1,4-glucanase, ten micrograms of recombinant enzyme were incubated with 100U of endoglycosidase H (Endo-H) at 37 °C for 1 h. Unless otherwise stated, all procedures were performed at 4°C. Related sequences were obtained by searching the database using the software BLAST. The amino acid sequences were aligned using the ClustalX, Phylogenetic analyses were performed in MEGA 6, using the maximum evolution approach. The automated protein structure homology-modelling server, SWISS-MODEL was used to generate the 3D structure of the enzyme. And the Discovery Studio ViewerPro 6.0 was used to visualize and analyze the structure of the model.
Conditions of the endo-b-1,4-glucanase activity assay
The endo-b-1,4-glucanase activity was determined using the 3,5-dinitrosalicylic acid (DNS) method18. The reaction system consisted of 0.07 ml of appropriately diluted enzyme ,0.07 ml citric acid–Na2HPO4 (pH 5.0) and 0.14 ml of 1% (w/v) CMC-Na dissolved in McIIvaine buffer. Assay mixtures were incubated at 37°C for 2h after initiating the reaction by the addition of enzyme, and terminated by the addition of 0.28 ml DNS reagent and boiled for exactly 10 min. After cooling down to room temperature in ice water bath, the reaction mixture was subjected to spectrophotography assay at 540 nm. One unit of enzyme activity was defined as the amount of enzyme that catalyzed the formation of 1ìmol glucose per minute under the assay conditions. All enzymological assays were carried out at least in triplicate and enzymological data in the graphs represent the averages of three replicates.
Characterization of the purified recombinant enzyme
Kinetic studies were done with freshly prepared enzyme mixed with various concentrations of CMC-Na added to McIlvaine buffer at 37°C for 1h. The velocities of enzyme-catalyzed reactions were measured at 540 nm for CMC-Na. The kinetic parameters, Km, were calculated form Lineweaver-Burk plots of values derived from three independent reactions.
Optimal pH and temperature values for enzyme activity were studied. PH, temperature stability, substrate specificity, the substrate of protection, the effect of metal ions, pepsin and trypsin on endo-b-1,4-glucanase activity, were determined as described previously19, with minor modifications.
To estimate the effect of pH on endo-b-1,4-glucanase activity was assayed using McIlvaine buffer at different pH ranges. The pH stability was examined by monitoring residual enzyme activity on carboxymethylcellulose sodium (CMC-Na) in McIlvaine buffer after preincubate the recombinant endo-b-1,4-glucanase in different pH(2-8) buffers at 37°C for 24h. The optimal temperature was investigated between 20 and 70°C at 10°C increments. Thermostability of the enzyme was determined by incubating the enzyme in citric acid-disodium hydrogen phosphate (McIlvaine) buffer (pH 5.0) at 30, 40, 50, and 60°C. Aliquots were removed at various times (from 10 to 60 min at 10 min intervals) and assessed with carboxymethylcellulose sodium (CMC-Na) as the substrate, as described above.
To evaluate the effect of metal ions, the enzyme was assayed in McIlvaine buffer (pH 5.0) containing 1% carboxymethylcellulose sodium (CMC-Na) with or without 1 mM or 10 mM metal ions [e.g.,CuSO4, CaCl2, Ca(NO3)2, Fe(NO3)2, MnSO4, ZnSO4, MnCl2, CoCl2, MgSO4, FeCl3, LiCl, CrCl3, NaCl]. One unit was defined as the amount of endo-b-1,4-glucanase oxidizing 1lmol CMC-Na per minute. The protein concentration was determined by Bradford assay kit (Sigma-Aldrich). All experiments were done in triplicate and expressed as mean values with standard errors.
CTCGAGAAAAGAGAGGCTGAAGCTGCCTCTTCCAGCAACGATGCTGACAAACTCTACTTT
K R E A E A A S S S N D A D K L Y F
GGCAACAACAAGTACTACCTCTTCAACAACGTTTGGGGTAAGGATGAGATCAAAGGCTGG
G N N K Y Y L F N N V W G K D E I K G W
CAGCAGACCATCTTCTACAACTCTCCAATCTCTATGGGGTGGAACTGGCACTGGCCATCC
Q Q T I F Y N S P I S M G W N W H W P S
TCTACTCACTCTGTCAAAGCCTATCCATCTCTTGTTTCTGGTTGGCATTGGACTGCTGGC
S T H S V K A Y P S L V S G W H W T A G
TACACTGAGAACTCTGGCTTGCCAATCCAACTCTCTTCCAACAAGTCTATCACCTCCAAC
Y T E N S G L P I Q L S S N K S I T S N
GTCACCTACTCCATCAAGGCTACTGGTACTTACAATGCTGCCTATGACATCTGGTTTCAC
V T Y S I K A T G T Y N A A Y D I W F H
ACTACTGACAAAGCCAACTGGGATTCTTCTCCAACTGACGAGTTGATGATCTGGCTCAAT
T T D K A N W D S S P T D E L M I W L N
GACACCAACGCTGGTCCAGCTGGTGACTACATTGAGACTGTCTTTCTTGGTGACTCCTCT
D T N A G P A G D Y I E T V F L G D S S
TGGAACGTCTTCAAAGGCTGGATCAACGCTGACAATGGTGGTGGTTGGAATGTCTTCTCC
W N V F K G W I N A D N G G G W N V F S
TTCGTCCACACTTCTGGCACCAACTCTGCCTCTCTCAACATCAGACACTTCACTGACTAC
F V H T S G T N S A S L N I R H F T D Y
CTGGTTCAGACCAAGCAGTGGATGTCTGACGAGAAGTACATCTCCTCTGTTGAGTTCGGT
L V Q T K Q W M S D E K Y I S S V E F G
ACTGAAATCTTCGGTGGTGATGGTCAGATTGACATCACTGAGTGGAGAGTTGACGTCAAG
T E I F G G D G Q I D I T E W R V D V K
CACCATCACCATCACCATTAAGAGCTC
H H H H H H *
Protease resistance was tested by incubating 0.05ml of endo-b-1,4-glucanase with pepsin (pH2.5) or trypsin (pH7.0) . The digested protein mixtures were tested for the remaining activity as described above, using McIIvaine buffer, pH 5.0, at 37°C for 1h, 3h, 5h and 7h.
Thermal stability of the enzyme that protected by the substrate
To evaluate the thermostability of the enzyme that protected by the substrate, it was assayed in solution containing 1% carboxymethylcellulose sodium at 50°C.Aliquots were removed at various times (from 10 to 60 min at 10 min intervals) and assessed with 1% carboxymethylcellulose sodium (CMC-Na) dissolved in McIlvaine buffer (pH 5.0). Furthermore, ck was the enzyme assayed in water at 70°C, and then dealed with 1.5% carboxymethylcellulose sodium (CMC-Na) dissolved in McIlvaine buffer (pH 5.0).
Synthesis of endoglucanase gene and construction of expression vector
Endoglucanase not correct: Codon optimized for Pichia pastoris (it is a yeast, but it is enough with the name)
To obtain a high level of expression, we designed and resynthesized the recombinant gene endoglueanases gene PceglAI using optimized genetic codons bias of yeast P. pastoris. The synthesized 747-bp recombinant gene was 74.96% identical to the wild type endo-b-1,4-glucanase A gene (GenBank accession no. YP_006283904). To improve the efficiency of gene transcription and RNA stability, the G+C content of the synthesized endoglueanases gene was keep to about 50%, and all hairpin structures and motifs containing consecutive ATs were eliminated by using degenerate codons. nineteen of 60 nt oligonucleotides were used for its synthesis with the PTDS method and the errors presented in the synthesized gene were corrected using multi-site mutagenesis method. The termination codon of recombinant endoglucanase is UAA but wide-type is UAG. Compared with wild-type gene, the recombinant gene with six histidine tag at the end is beneficial to purify (Fig.1a). The gene encoding the mature endo-b-1,4-glucanase was fused in-frame to the yeast mating pheromone b-factor signal sequence after inserted into expression vector. In the construct pYR1725, the endo-b-1,4-glucanase gene was controlled by the alcohol oxidase1 (AOX1) promoter.
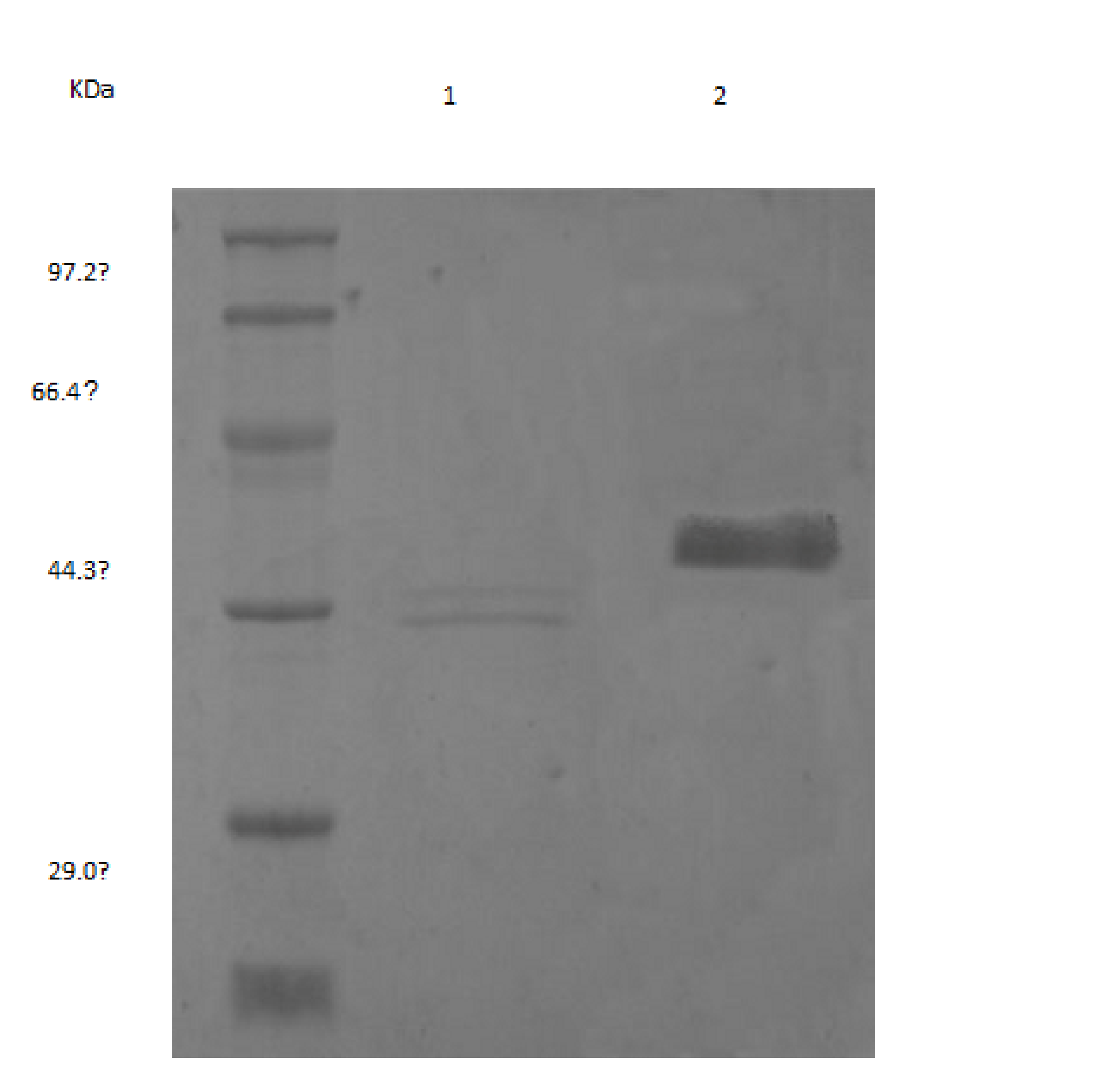
Fig. 1. a. The nucleotide sequences deduced amino acid sequences of the synthesized PcgelAI gene. The sequences KREAEA reconized by Kex2 protease and six histone were highlighted with double line. Three putaitive N-glycosylation sites (N-X-S/T) were showed in square frame. The sequences used for primer design are shown in bold with arrow to indicate their directions. The stop codon is indicated by an asterisk.
b. SDS-PAGE of recombinant endo-b-1,4-glucanase secreted by P.pastorsis. Lane M: molecular mass standard, Lane 1: purified recombinant endo-b-1,4-glucanase by Ni2+–NTA agarose affinity column, Lane 2: deglycosylated recombinant endo-b-1,4-glucanase with Endo H, the lane of 29KDa was Endo H, and the lane of 28KDa was endo-b-1,4-glucanase.
High expression of PceglAI in P. Pastoria
In the recombinant Pichia strains, the heterologous genes were situated downstream of the AOX1 promoter and the coding sequence of the signal peptide of the yeast amating factor. With induction by methanol, the recombinant strains would express and secrete recombinant gene into the culture broth.After 3 day incubation approximately 180 clones were generated of which several clones were selected for assay of endo-b-1,4-glucanase activity. One clone showing the highest endo-b-1,4-glucanase activity from the culture supernatant was chosen for further studies. The recombinant strain could be induced by 1% methanol in liquid BMMY to produce endo-b-1,4-glucanase efficiently. The PceglAI was purified and detected by SDS-PAGE. The molecular mass of purified enzyme from yeast cell was approximately 28 KDa, and had a single protein band (Fig.1b). The enzyme kinetics of recombinant endo-b-1,4-glucanase against CMC-Na was investigated.It revealed that the apparent Km value of the enzyme determined from the Lineweaver-Burk plot was estimated to be 64.5 mM. And the specific activity of PceglAI was 7×103 U mg-1.
Amino-acid sequences analysis and tertiary structure of the endoglucanase
In this paper, the phylogeny of PceglAI was studied by construct phylogenetic tree with amino acid sequence of endoglucanase from the bacteria, fungi, plants and animals. The complete amino acid sequence of Pectobacterium wasabiae SCC3193 has been deposited in the NCBI nucleotide sequence database under accession number YP_006283904 (Fig.2). Proteins similar to Pectobacterium wasabiae SCC3193 were collected by web BLAST search showed the high amino acid sequence identity (87%) with Pectobacterium carotovorum subsp. Carotovorum PCC21, 66% and 52% identity, respectively, with Bacillus licheniformis DSM13=ATCC14580 and Paenibacillus mucilaginosus3016.
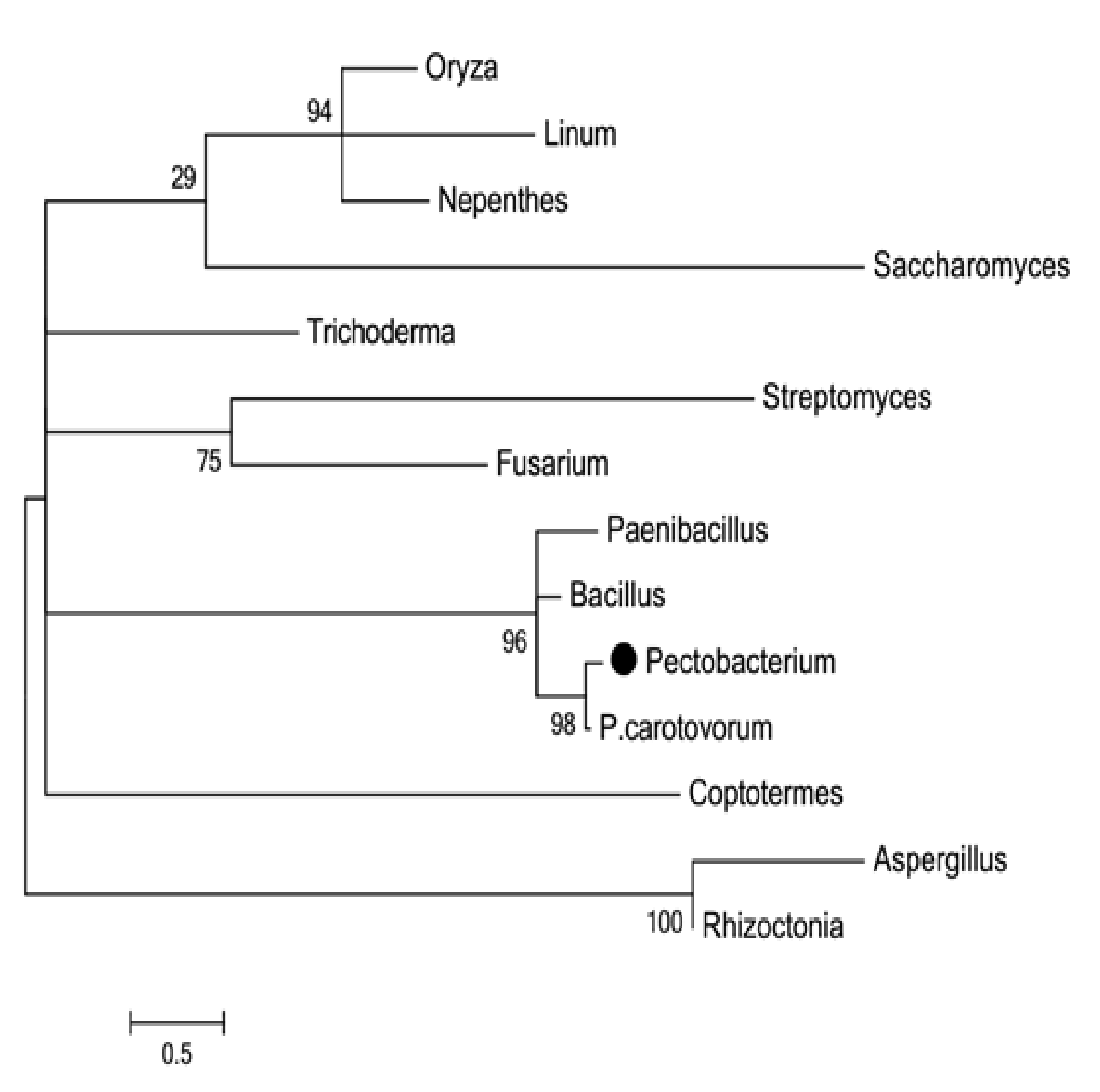
Fig. 2. Phylogenetic tree for the endo-b-1,4-glucanase gene PceglAI. The tree was constructed as described under Materials and Methods. Branches are drawn to scale as indicated by the scale bar (=0.1 substitutions/1000 residues). Bootstrap values are indicated above the nodes. Pectobacterium wasabiae SCC3193 is highlighted and other species are indicated by common names
Based on the putative tertiary structures, the putative endo-b-1,4-glucanase gene PceglAI from P. wasabiae SCC3193 belongs to the GH12 family of glycoside hydrolases. A “jelly-roll” fold of a six stranded antiparallel b-sheet packing on the outside of a nine-stranded mostly antiparallel b-sheet curved around an active site cleft. The two curved b-sheets A and B are linked by interconnecting loops, loop A6-B4 encompasses a two-turn á-helix, while the convex side of the sheet forms hydrophobic interactions with sheet A and with the helix. There are more aromatic amino-acid residues in the cleft of the bacterial enzymes. We found several amino acid residues, include Gly31, Pro61, Pro80, Trp110, Met128, Trp130, Trp168, Gly212, and two catalytic residues, Glu126 and Glu214, were conserved as some hyperthermophilic endoglucanase. Among these conserved amino acids, two glutamic acid residues might be the catalytic nucleophile and proton donor that have been reported in many enzymes20, other eight conserved amino acids might be necessary to the thermostability of protein and binding of the substrate22. The reducing end of the active site cleft is closed by a loop designated the cord, which is of fixed conformation and contains a conserved Pro-X-Gly motif, it may causes the substrate to distort from the relaxed conformation seen in cellulose and aids hydrolysis by interact with the cord. This is like to the family 12 cellulases and the family 11 xylanases, while the same crystal structure has been seen in Rhodothermus marinus endoglucanase Cel12A.
High expression of PceglAI in P. Pastoria
In the recombinant Pichia strains, the heterologous genes were situated downstream of the AOX1 promoter and the coding sequence of the signal peptide of the yeast amating factor. With induction by methanol, the recombinant strains would express and secrete recombinant gene into the culture broth.After 3 day incubation approximately 180 clones were generated of which several clones were selected for assay of endo-b-1,4-glucanase activity. One clone showing the highest endo-b-1,4-glucanase activity from the culture supernatant was chosen for further studies. The recombinant strain could be induced by 1% methanol in liquid BMMY to produce endo-b-1,4-glucanase efficiently. The PceglAI was purified and detected by SDS-PAGE. The molecular mass of purified enzyme from yeast cell was approximately 28 KDa, and had a single protein band (Fig.2). The enzyme kinetics of recombinant endo-b-1,4-glucanase against CMC-Na was investigated.It revealed that the apparent Km value of the enzyme determined from the Lineweaver-Burk plot was estimated to be 64.5 mM. And the specific activity of PceglAI was 7×103 U mg-1.
Effect of pH and temperature on endo-b-1,4-glucanase activity and stability
The pH profile for endo-b-1,4-glucanase activity against carboxymethylcellulose sodium (CMC-Na) showed its optimum pH was 4.0 (Fig.3a). The activity of endo-b-1,4-glucanase was stable after incubation for 24 h at 37 °C in 1% citrate–phosphate buffer at pH 5-8. Moreover,it showed that the enzyme activity increase slightly at pH 6.0 (Fig.3b).
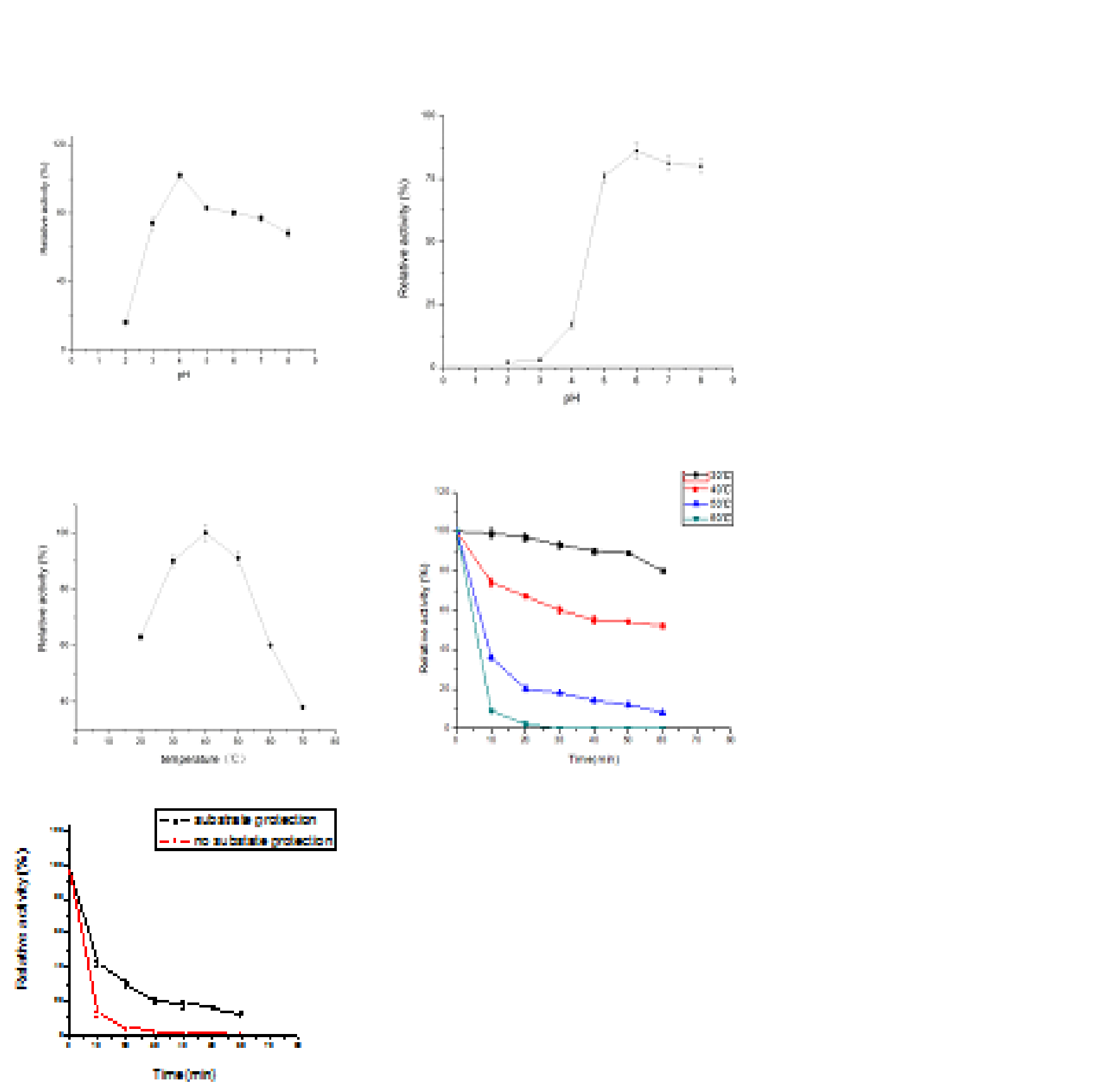
Fig. 3. Effect of pH on endo-b-1,4-glucanase: a.Optimum pH of PceglAI was determined at various pH from 2.0 to 8.0. b. pH stability of PceglAI was assayed after incubating the endo-b-1,4-glucanase for 24 h at 37°C. Values are the mean±SD of three separate measurements.In adition,effect of temperature on endo-b-1,4-glucanase: c. Optimal temperature of PceglAI was measured with temperatures from 20 to 70 °C in increments of 10°C. d. The thermostability of PceglAI was assayed via incubation of the purified endo-b-1,4-glucanase at 30, 40, 50 or 65°C with different durations(10 min increment). e.Thermal stability of the enzyme that protected by the substrate.Ten minutes later, under the aegis of the substrate, the thermostability of endo-b-1,4-glucanase increased four times. Values are the mean±SD of three separate measurements
The effect of temperature on enzyme activity showed the velocity of endo-b-1,4-glucanase catalysis was maximal at 40°C, its activity was high from 20 to 60°C (Fig.3c). The endo-b-1,4-glucanase is stability below 40°C, after incubated at 30°C for 60 min, the remaining activity was 80%. Inactivation of enzyme is powerful when the temperature was over 40°C, 65 % of activity persisted at 40°C for 10 min, declined quickly when temperature was above 50°C, 36 % at 50°C for 10 min and completely depleted at 60°C for 10 min (Fig.3d).
Substrate protection on thermal stability of the enzyme
The endo-b-1,4-glucanase was treated in 1% carboxymethylcellulose sodium at 50°C. Compared with the enzyme in water, the thermostability was enhanced significantly. Ten minutes later, under the aegis of the substrate, the thermostability of endo-b-1,4-glucanase increased four times.Treatment for 20, 30, 40 minutes, the thermostability of enzyme improved 10-fold, 18- fold,20-fold (Fig.3e).
Effect of metallic ions on endo-b-1,4-glucanase activity
The effects of metal ions on enzyme activity were determined (Fig.4). Most metallic ions had no influence on the activity of endo-b-1,4-glucanase at a low concentration of 1 mM Whereas addition of 1 mM Cu2+ and Mn2+ resulted in a inhibition of activity to 70% and 60% respectively. When the concentration reached to 10 mM, Na+ and Cr+ could slight promoted its activity, Zn2+ significantly promote the activity, and Cu2+, Mn2+, Fe3+ and Co2+ have the effect of inhibition.
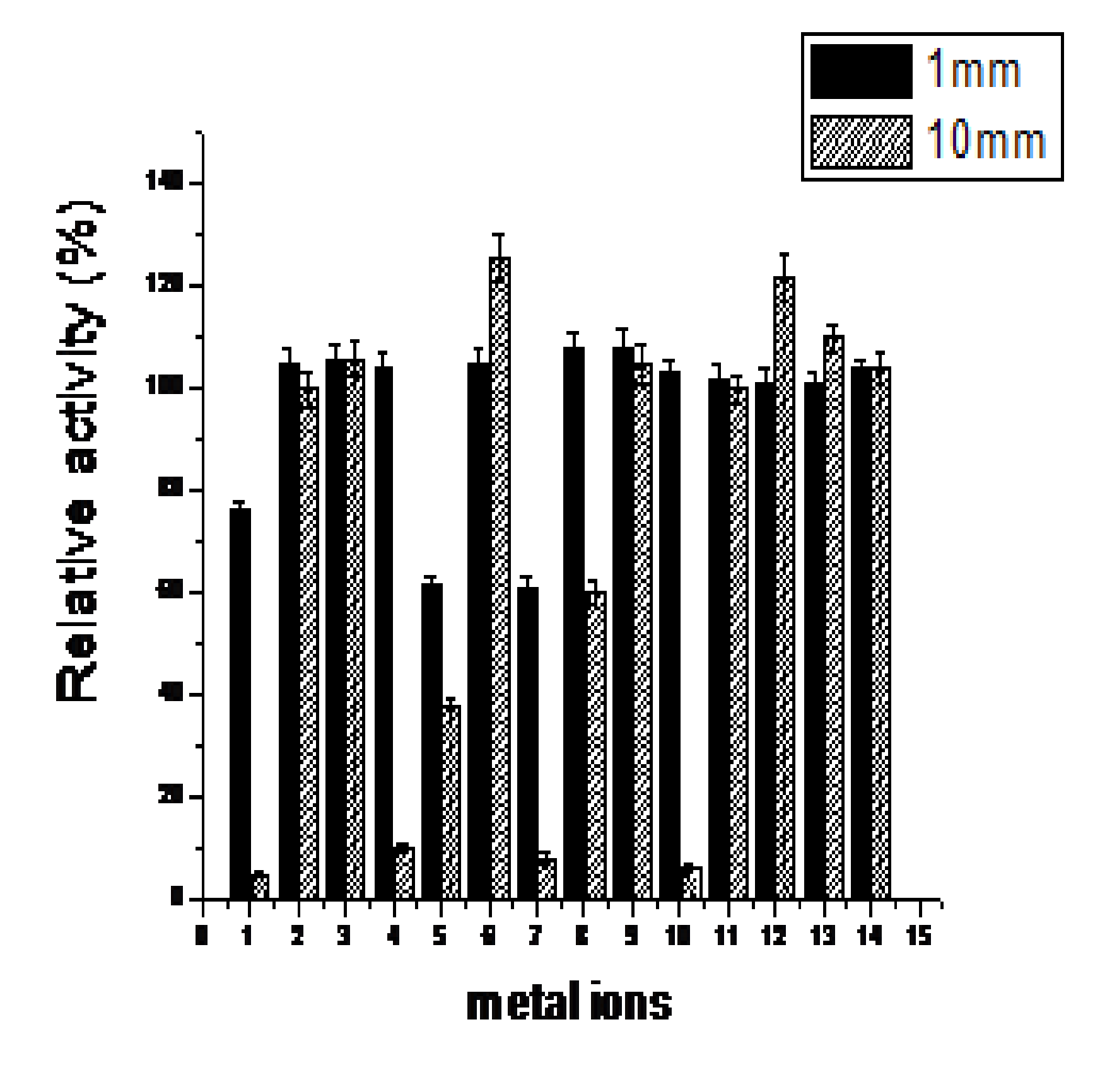
Fig. 4. Effect of metal ions on endo-b-1,4-glucanase. 1-14 is CuSO4, CaCl2, Ca(NO3)2, Fe(NO3)2, MnSO4, ZnSO4, MnCl2, CoCl2, MgSO4, FeCl3, LiCl, CrCl3, NaCl, H2O, respectively.Values are the mean±SD of three separate measurements
Effect of pepsin and trypsin on glucanase activity
Residual activity of the recombinant endo-b-1,4-glucanase, after treating with pepsin or trypsin after 1h, 3h, 5h, and 7h (Fig.5). The identification results of coloration showed that both pepsin and trypsin decrease the activity of enzyme. As show in this study, after 3h, the activity of enzyme treated with trypsin ramained 60 %, but the enzyme in pepsin almost inactive.In addition,the activity of the enzyme is 50% after 5h in trypsin.
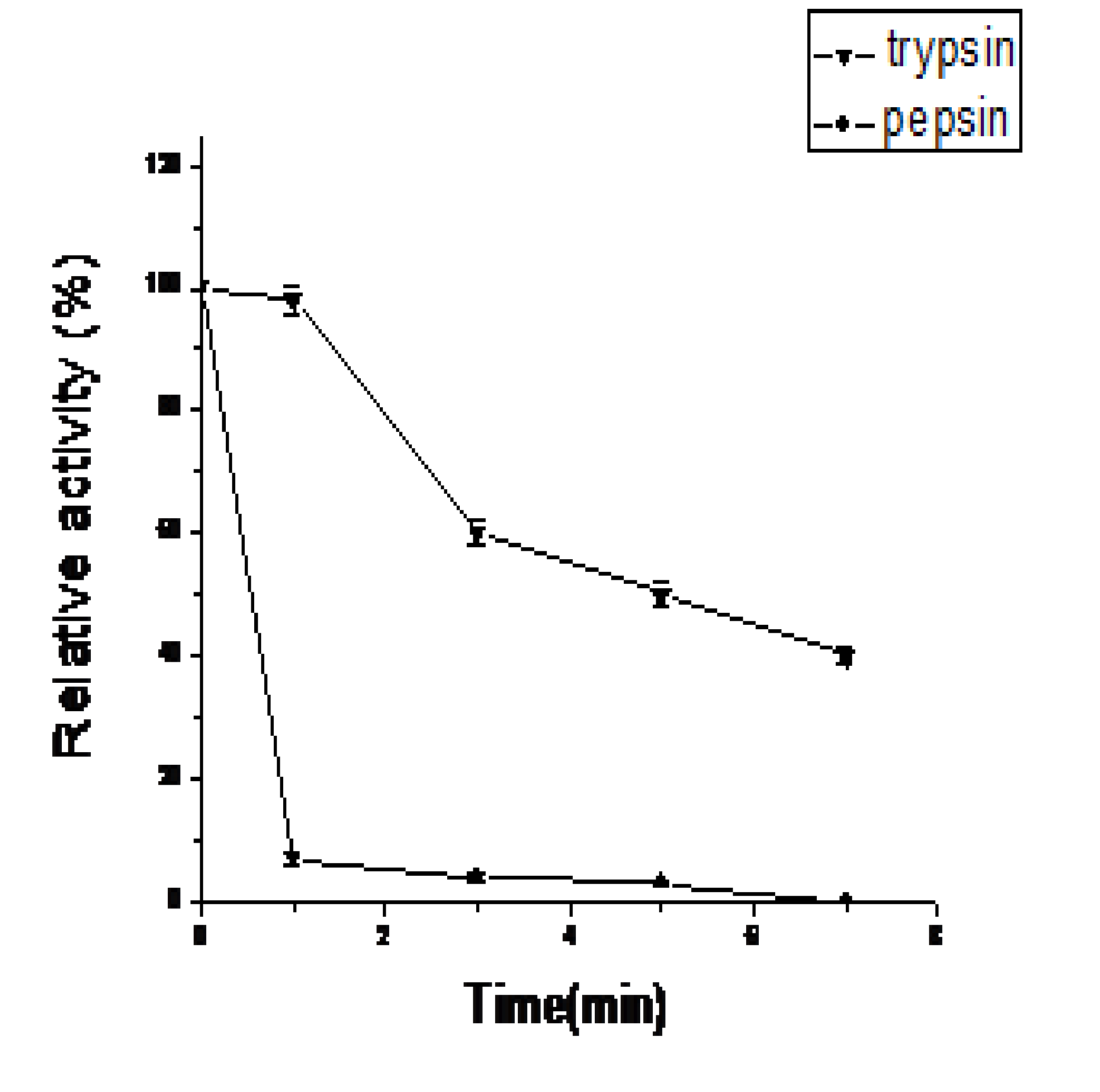
Fig. 5. Effect of protease on endo-b-1,4-glucanase: Residual activity of PceglAI , was determined after treating with pepsin and pepsin in McIlvaine buffer (pH 5.0) containing 1% carboxymethyl cellulose sodium (CMC-Na). Values are the mean±SD of three separate measurements
Pectobacterium wasabiae SCC3193 has been a model strain in the research of soft rot molecular biology. The pathogenicity is due to their ability to degrade the vascular bundle cells of plants through secreting enzyme system including pectinase, cellulase, and protease and so on. In the present study, we have synthesized and highly expressed a putative endo-b-1,4-glucanase from Pectobacterium wasabiae SCC3193 in P. pastoris. Then we study its characterization with hydrolytic activity against CMC-Na carefully.
The optimal pH of cellulases from other sources varied from acidic to alkaline. PceglAI is an enzyme that appears to be optimally adapted to work under acidic conditions as the optimal pH is 4.0 and with a comparatively slow decrease in activity on both acid and alkaline sides. More than 70% of the activity is found in the range pH 3-8, similar to the pH activity curve of the low molecular mass endo-b-1,4-glucanases of Trichoderma reesei. Moreover, PceglAI has the same pH optima as that of some fungal endo-b-1,4-glucanases but is more active at lower pH (more than 50% activity at pH 2.5) . The acidophilic property would play an important role in some biotechnological applications, for example, the acidic cellulases could be used as animal feed supplements as a means of improving animal performance.
Generally, most of endo-b-1,4-glucanases are stable up to 40-55°C. This enzyme degraded CMC showed a maximal activity at 40°C. Some factors, reported in previous article, that fraction of buried atoms, accessible surface area, and lengths of loops may also influence the thermostability of enzymes20 . It has been reported that glycosylation has significant effects on enzyme thermostability21, the optimum pH value22, and resistance to proteases23, 24. Thermostability is an important factor for commercial enzymes, especially cellulases owing to their enormous potential for industrial applications .
The effects of Cu2+ and Mn2+ were same at 1mM and 10mM. Addition of Cu2+ inhibit the actiyity to apparently 70% is somewhat alike to, endoglucanase from Bacillus subtilis DLG25. And 1mM Mn2+ resulted in an inhibition of activity to 15%., In the presence of 10 mM Cu2+ and Mn2+, the enzyme activity was serious repressed, an observation quite attract the gaze of people. In previous studies, the endo-b-1,4-glucanase activity was inhibited by 1 mM Mn2+ 26, 27 but stimulated by 10 mM Mn2+. Mn2+ acted as an activator for endoglucanase had not been reported to date. The activity of enzyme treated with trypsin ramain 95%, but the enzyme in pepsin almost inactive after 1h. This shows that the enzyme cloned from the model soft rot phytopathogen Pectobacterium wasabiae SCC3193 has resistance to trypsin to a certain extent, and no resistance to pepsin.
The overall architecture of PceglAI is the classic b-jelly roll, based on the template Bacillus Licheniformis. It is similar to the cellulases from Rhodothermus marinus Cel12A, Streptomyces lividans CelB2 and Hypocrea jecorina Cel12A(Trichoderma reesei Cel12A), and closely resembling the topology of the glycoside hydrolase family 11 xylanases. The relationship between the conserved amino acid residues of GH12 and their molecular structures have been illustrated in some researches, such as endoglucanase Cel12B from Thermotoga maritima. The residues Gly31, Gly212, Pro61, Pro80 and increase in the aromatic content of active sites might assumed for the thermostability of the enzyme. The reason for proline contribute to thermostability, part of the explanation may be it could provide closer packing of each region. The Trp110, Trp130, Trp168 and Met128 predicted to be related to the binding of substrate. The tryptophan residue may be essential in mediating the total cooperativity of the response of the enzyme to substrate, so it exhibited a pronounced effect on the binding of substrate and the enhancement of the total enzyme activity. It is clear that the bacterial enzymes are more diverse, the major differences are between the connecting loops, whereas the b-strands constituting the jelly-roll scaffold are largely conserved. The loops connecting the b-strands show variations in their lengths and dispositions, some apparently giving rise to different substrate affinity and specificity.
During the bioconversion process, lignocellulosic biomass is generally pre-treated with acid at high temperatures, followed by enzymatic hydrolysis, thus adjustment of pH and temperature is necessary. Some researchers have explore the mechanism underlying enzyme thermostability. The comparison of two structures of the mesophilic and hyperthermophilic endoglucanase, Crennell’s study mentioned that large increase in the number of ion pairs and the stabilisation of a highly mobile loop on the periphery of the active site, together with sequence changes to counter deamidation appeared to be major features contributing to the increased thermostability. Despite the mesophilic is more similar to hyperthermophilic across the structure as a whole, the reducing end of the cleft is an area where there are substantial sequence differences between the mesophile and thermophile. It has been reported that the reducing end is both the area of greatest main chain movement on substrate binding and the area of greatest difference in substrate or product binding strength between the hyperthermophilic and mesophilic enzymes . Besides those enumerated above, it was demonstrated that loop and turn played a key role in thermostability of protein, especially for the proteins that proline is located in loop or turn region , it is likely to be because proline in the polypeptide chain possesses less conformational freedom than other amino acids.
PceglAI might have the potential to be a key enzyme in bioethanol production from cellulose due to its temperature and pH stability. In the procession of producing ethanol, cellulose is hydrolyzed using acids at high temperatures. In the subsequent step of hydrolysis, a thermophilic and acidophilic cellulase may simplify the reaction process and reduce production costs28. In addtion, acidophilic stability and high resistance to proteolysis very likely allow PceglAI to function under the acidic conditions and high degree of proteolytic degradation in the digestive tract of animals. This indicate that PceglAI can be a candidate for improving nutrient utilization in the animal feed industry. (Gene cloning and expression of a new acidic family 7 endo-b-1,3-1,4-glucanase from the acidophilic fungus Bisporasp. MEY-1)
In conclusion, we have identified a novel enzyme PceglAI, with acidic pH optimum, broad range of pH stability, and relatively high thermostability. These properties make this endo-b–4-glucanase a good candidate for use in industrial applications. Through a series of enzymology properties research, we confirm that the endo-b-1,4-glucanase expressed by YP_006283904 gene is a powerful enzyme in Pectobacterium wasabiae. It has high specific activity, we infer that the endo-b-1,4-glucanase may be involved in the pathogenic process. This is the first time to purify and characterize an endo-b-1,4-glucanase from Pectobacterium wasabiae, and the result may provide more insight into the cellulase system of this organism.
ACKNOWLEDGMENTS
The research was supported by the National Natural Science Foundation of China (31672439), the Key Project Fund of the Shanghai Municipal Committee of Agriculture (gongzi 2014-7-1, zhongzi2016-1-2), International Scientific and Technological Cooperation (13440701700).
- Park, Y. W., Lim, S. T., Cho, S. J., and Yun, H. D. Characterization of Erwinia carotovora subsp.carotovora LY34 endo-1,4-b-Glucanase Genes and Rapid Identification of Their Gene Products. Biochem. Bioph. Res. Co., 1997; 241, 636-641.
- Pérombelon, M. C. M. Potato diseases caused by soft rot erwinias: an overview of pathogenesis. Plant Pathol., 2002; 51, 1-12.
- Chahed, H., Ezzine, A., Mlouka, A. B., Hardouin, J., Jouenne, T. and Marzouki, M. N. Biochemical characterization, molecular cloning, and structural modeling of an interesting b-1,4-glucanase from Sclerotinia Sclerotiorum. Mol Biotechnol. 2014; 56, 340-350.
- Bhat, M. K. and Bhat, S. Cellulose degrading enzymes and their potential industrial applications. Biotechno. Adv., 1997; 15, 583-620.
- Demain, A. L., Newcomb, M. and David Wu, J. H. Cellulase, clostridia, and ethanol. Microbiol. Mol. Biol. R. 2005; 69, 124-154.
- Henrissat, B. Cellulases and their interaction with cellulose. Cellulose. 1994; 1,169–196
- McCarthy, T., Hanniffy, O., Savage, A. V., Tuohy, M. G. Catalytic properties and mode of action of three endo-b-glucanases from Talaromyces emersonii on solubleb-1,4- and b-1,3;1,4-linked glucans. Int. J. Bio. Macromol. 2003; 33, 141-148.
- Zhang, Y. P., and Lynd, L. R. Toward an aggregated understanding of enzymatic hydrolysis of cellulose: noncomplexed cellulase systems. Biotechnol. Bioeng., 2004; 88, 797-824
- Celestino, K. R., Cunha, R.B., and Felix, C.R. Characterization of a b-glucanase produced by Rhizopus microsporus var. microsporus, and its potential for application in the brewing industry. BMC Biochem. 2006; 7, 23.
- Walsh, G.A., Murphy, R.A., Killeen, G.F., Headon, D.R., Power, R.F. Technical note: detection and quantification of supplemental fungal b-glucanase activity in animal feed. J. Anim. Sci., 1995; 73, 1074-1076.
- Koga, J., Baba, Y., Shimonaka, A., Nishimura, T., Hanamura, S., and Kono, T. Purification and characterization of a new family 45 endoglucanase, STCE1, from Staphylotrichum coccosporum and its overproduction in Humicola insolens. Appl. Environ. Microb. 2008; 74, 4210-4217.
- Schulein, M. Enzymatic properties of cellulases from Humicola insolens. J. Biotechn. 1997; 57, 71–81.
- Gu, F.Y., Chen, C.Y., Shi, J.J., and Qian, S.J. Advances in cellulase and its development tendency. J. Microbiol. 2008; 28, 83-87.
- Ljungdahl, L.G. The cellulase/hemicellulase system of the anaerobic fungus Orpinomyces PC-2 and aspects of its applied use. Ann. NY. Acad. Sci., 2008; 1125, 308-321.
- Zhang, Y. H. P., Himmel, M. E., and Mielenz, J. R. Outlook for cellulase improvement: Screening and selection strategies. Biotechnol. Adv. 2006; 24, 452-481.
- Czajkowski, R., Pérombelon, M. C. M., van Veen, J. A., and van der Wolf J.M. Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeyaspecies: a review. Plant Pathol. 2011; 60, 999-1013.
- Xiong, A.S., Yao, Q.H., Peng, R.H., Li, X., Fan, H.Q., Cheng, Z.M., and Li, Y. A simple, rapid, high-fidelity and cost-effective PCR-based two-step DNA synthesis method for long gene sequences. Nucleic Acids Res. 2004; 32: 1-10.
- Zhao, J. Q., Shi,P. J., Huang, H. Q., Li, Z. Y., Yuan, T. Z., Yang, P. L., Luo, H. Y., Bai, Y. G., and Yao, B. A novel thermoacidophilic and thermostable endo-b-1,4-glucanase from Phialophorasp. G5: its thermostability influenced by a distinct b-sheet and the carbohydrate-binding module. Appl. Microbiol. Biot., 2012; 95, 947-955.
- Thitinard, N., Akira, W., and Yasuhiko, A. Extracellular laccase produced by an edible basidiomycetous mushroom, Gridola frondosa: purification and characterization. Biosci. Biotech. Bioch, 2011; 75, 538-543.
- Pereira, J. H., Chen, Z. W., McAndrew, R. P., Sapra, R., Chhabra, S. R., Sale, K. L., Simmons, B. A., and Adams, P. D. Biochemical characterization and crystal structure of endoglucanase Cel5A from the hyperthermophilic Thermotoga maritime. J. struct. Bio. 2010; 172, 372-379.
- Clark, S.E., Muslin, E.H., and Henson, C.A. Effect of adding and removing N-glycosylation recognition sites on the thermostability of barley á-glucosidase. Protein Eng. Des. Sel. 2004; 17: 245-249.
- Somera, A.F., Pereira, M.G., Souza Guimaraes, L.H., Polizeli, M de L., Terenzi, H.F., Melo Furriel, R.P., and Jorge, J.A. Effect of glycosylation on the biochemical properties of b-xylosidases from Aspergillus versicolor. J. Microbiol. 2009; 47, 270-276.
- Wyss, M., Pasamontes, L., Friedlein, A., Remy, R., Tessier, M., Kronenberger, A., Middendorf, A., Lehmann, M., Schnoebelen, L., Rothlisberger, U, et al. Biophysical characterization of fungal phytases (myo-inositol hexakisphosphate phosphohydrolases): molecular size, glycosylation pattern, and engineering of proteolytic resistance. Appl. Environ. Microb., 1999; 65, 359-366.
- Takegawa, K., Yoshikawa, M., Mishima, T., Nakoshi, M., and Iwahara, S. Determination of glycosylation sites using a protein sequencer and deglycosylation of native yeast invertase by endo-b-N-acetylglucosaminidase. Biochem. Int. 1991; 25, 585-592.
- Robson, L. M., and Chambliss, G. H. Characterization of the cellulolytic activity of a Bacillus isolate. Appl. environ. microb. 1984; 47, 1039-1046.
- Mawadza, C., Hatti-Kaul, R., Zvauya, R., and Mattiasson, B. Purification and characterisation of cellulases produced by two Bacillus strain. J. Biotechnol. 2000; 83, 177-187.
- Kim J. Y., Hur, S. H., and Hong, J.-H. Purification and characterization of an alkaline cellulase from a newly isolated alkalophilic Bacillus sp. HSH-810. Biotechnol. Lett. 2005; 27, 313-316.
- Huang, Y. W., Krauss, G., Cottaz, S., Driguez, H. and Lipps, G. A highly acidstable and thermostable endo-beta-glucanase from the thermoacidophilic archaeon Sulfolobus solfataricus. Biochem. Soc. 2005; 385, 581-588.
© The Author(s) 2018. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.