


ISSN: 0973-7510
E-ISSN: 2581-690X
Probiotics are microorganisms that are safe and stable under product development conditions and are used as adjuncts to food and drugs to promote health, including the acceleration of nutrient digestion. Here, we present evidence on the ability of two isolates, Lacticaseibacillus paracasei BCRC-16100 and L. paracasei ZFM54, to (1) enhance carbohydrate digestion, (2) tolerate processing conditions, and (3) demonstrate safety in terms of antimicrobial resistance (AMR). These approaches include whole-genome sequence (WGS) analysis, gene expression, and bioactivity assays. WGS revealed genes encoding enzymes involved in carbohydrate digestion, tolerance to processing conditions, and AMR. The ability of the two strains to digest carbohydrates was confirmed by glucose release when cultured alongside starch. The isolates also showed versatility across a range of temperatures and alcohol concentrations, indicating their suitability for product development. Genes cause AMR, particularly against vancomycin, through three mechanisms: transporter control, transcriptional regulation, and efflux pumps. Furthermore, promoter, gene expression, and transposable element analyses showed that some upregulated AMR genes in the presence of antimicrobials were transposable. Altogether, we show the potential of the two isolates for incorporation into products as probiotics to improve carbohydrate digestion, while considering precautions regarding mobile AMR genes that may compromise safety.
Probiotics, Nutrient Digestion, Antibiotic Resistance, Antibacterial Activity, Whole Genome Sequence
In the last ten years, there has been significant progress in probiotic research, with many studies indicating the essential role of probiotics in maintaining human health. Probiotics are increasingly gaining popularity and market share owing to their potential health benefits, such as improved bioavailability of macronutrients, altered gut microbiota diversity, and treatment of several diseases. Microbes in the genera Pediococcus, Lactococcus, Enterococcus, Streptococcus, Propionibacterium, and Bacillus are generally considered probiotics, provided they pass screenings for function and safety.1
Several studies have shown that probiotics can improve the digestion of carbohydrates such as lactose,2 and lactose-intolerant patients supplemented with lactobacilli or bifidobacteria in their diet showed improved bioavailability and digestion of lactose.3-6 Moreover, probiotics facilitate the digestion of resistant starch. When the fiber passes through the stomach and small intestine, undigested Bifidobacteria in the large intestine catalyze its hydrolysis.7,8 There is also evidence of enhanced protein digestion and absorption with probiotics through multiple mechanisms, such as the production of proteases, regulation of microbiota to favor microbes with peptidase activities, and improved absorption of peptides and amino acids through accelerated transport in the intestine.9 Lactobacillus-derived proteases have been shown to catalyze the hydrolysis of complex polypeptide chains.10 Some of the most common hydrolytic enzymes in microbes are aspartic proteases, cysteine proteases, metalloproteases, and serine proteases.11 These findings indicate the potential of probiotics to facilitate the utilization of nutrients from food sources with low digestibility, which consequently improves digestion and the overall nutritional value.
In addition to digestive benefits, probiotics can alter the gut microbiota, favoring microbial groups that can improve health in three ways, (1) modifying the internal host environment, (2) changing the composition of the gut microbiome, and (3) inhibiting pathogen proliferation. First, a shift in the diversity of the gut microflora changes the internal environment of the host gut by increasing the amount of specific metabolic compounds.12 Metabolites from probiotics, including organic acids, short-chain fatty acids, teichoic acids, peptidoglycans, plasmalogens, neurotransmitters, biosurfactants, amino acids, and flavonoid-derived compounds such as desaminotyrosine, equol daidzein, noratirriol, terpenoids, and phenolic compounds, have been found to have positive effects on consumer health.13-15 Second, observations in animal models suggest that the composition of the gut microbiome changes with probiotic supplementation.16 A wide range of studies have shown that a perturbed microbial community in the gut can be restored using probiotics, which promote the competitive exclusion of pathogen binding.17-19 Probiotics can modulate the gut microbiota by producing antimicrobial peptides (AMPs) such as bacteriocins, which function as a natural bacterial immune defense system by binding to the corresponding receptor of both narrow- and broad-spectrum microorganisms. Additionally, probiotics produce other metabolites such as lactic acid, which are harmful to several pathogenic strains of microorganisms.20-23 Thus, bacteria that produce lactic acid as the end product of carbohydrate fermentation are generally recognized as safe (GRAS) by the Food and Drug Administration (FDA) in the Philippines. Scant nutrients in the environment trigger the production of bacteriocins that compete for space and resources, thereby excluding potential pathogens from the gut microbiome and altering the composition of the intestinal microbiota. Finally, alteration of the gut microbiome and modification of the host internal environment through consumption of probiotics consequently treated gastrointestinal diseases in controlled clinical trials24-25 and improved disease susceptibility.26
Probiotics may provide multiple health benefits, but it is equally essential that microbes used as probiotics are non-pathogenic, non-toxigenic, versatile, and remain stable and viable for long periods of storage and harsh conditions for incorporation in both functional food and dietary supplements and drugs.20 Probiotics should be generally safe for the intended use and meet standards for purity, identity, and potency. A thorough evaluation of the pathogenicity, physiological and metabolic activities, and intrinsic properties is required. Despite the variation in regulatory requirements in each country, a comprehensive assessment of probiotic strains intended for use in foods and human supplements must be conducted,27 through methods that include (1) assessment of physiological activities, (2) strain identity, and (3) screening for antimicrobial resistance. First, the industrial production of probiotic strains should be based on the properties of the strains involved and their ability to withstand stress during processing and storage.28 Second, the unambiguous identification of the probiotic strains of interest should be considered, and whole-genome sequencing (WGS) is used for this purpose. Finally, only probiotic strains which do not have antimicrobial resistance (AMR) genes should be selected for use in food and supplements.29 Phenotypic assessment of antimicrobial resistance genes consists of testing the probiotic strain against a set of clinically important antimicrobials and determining colony forming units for each antimicrobial exposure with respect to the original log concentration of the strain culture. The annotated genome sequences of probiotics are utilized to inspect the genetic elements responsible for metabolic activities, physiological activities, hemolytic activity, toxins, and antimicrobial resistance. The horizontal transfer capability of antimicrobial resistance is further determined by gene identification either on a plasmid or in close proximity to mobile elements, such as transposases. The vancomycin-resistant phenotype present in several strains of Lactobacillus is a well-known example of intrinsic AMR, which is attributed to the properties of the cell wall that prevent the binding of the antimicrobial agent.30 Based on the European Food Safety Authority EFSA in 2012, any functional AMR genes found within the genome of probiotic strains should be characterized as intrinsic or transmissible. This classification will help in determining the risk of potential spread, which is a major public health concern.31 Countries with modern regulatory structures utilize advanced technology to establish probiotic safety profiles, with a greater emphasis on WGS.
Candidate probiotic strains are ubiquitous. They can be isolated from human and animal origins, such as the gastrointestinal tract and breast milk, and various food biotopes, such as fermented food products and dairy products, as well as from different parts of plants.32 Several potential probiotic strains have been isolated from fermented food products of plant origin.33 This broad range of inexhaustible sources poses a challenge for probiotic strains to adapt to various environments, and these abilities vary significantly among species and even at the strain level34; hence, there is significant variation in their genomic and metabolomic profiles. These variations arise from genetic mutations and horizontal gene transfer, leading to phenotypic variations, metabolic changes, and antimicrobial resistance. Additionally, environmental factors, such as nutrient availability, temperature, pH, and competition with other microorganisms, exert selective pressures that shape bacterial populations, further contributing to the diversity of bacterial characteristics across various sources.35
Probiotics are becoming increasingly popular among consumers because they offer multiple health benefits. This trend necessitates the isolation of probiotics from different sources, ensuring their safety, and evaluating their functional use as supplements. Here, we elucidated the potential benefits of L. paracasei BCRC-16100 and L. paracasei ZFM54 from Nipa sap in improving the digestion of carbohydrates and proteins. Moreover, we explain the bottlenecks in safety with an emphasis on AMR and the underlying biomolecular mechanisms. Finally, we provide insights into the known safety limitations of these two isolates.
Isolation and screening of Lactic Acid Bacteria (LAB)
The de Man Rogosa Sharpe agar (Sigma-Aldrich, Massachusetts, USA) supplemented with 1% CaCO3 was used as the medium for cultivation and isolation of the two probiotic isolates from Nipa sap. One hundred microliters of the serially diluted sample were aseptically inoculated onto MRSA plates using the spread-plate method. The plates were incubated for 24 h at 37°C. Individual colonies with a colony morphology of putative LAB grown overnight were further subjected to standard purification of bacterial cultures. Purified putative LAB were subjected to Gram staining test (Sigma-Aldrich Massachusets, USA) and a catalase test using 3% H2O2. Moreover, a hemolysin test using Sheep Blood Agar (Remel Tryptic Soy Agar with 5% Sheep Blood) was conducted using gram-positive and catalase-negative putative LAB. Isolates that exhibited γ-hemolysis were stored and sent for molecular identification.
Identification of Isolates by Capillary Sequencing
The genomic DNA (gDNA) of L. paracasei BCRC-16100 and L. paracasei ZFM54 was extracted using the Quick-DNA Fungal/Bacterial Miniprep Kit (Zymo Research, USA) according to the manufacturer’s protocol. The Polymerase Chain Reaction (PCR) amplicons were purified using AMPure XP beads (Cat. No. 163881, Beckman Coulter/Agencourt, California, USA). One microliter of the purified PCR amplicons was loaded onto a 1% agarose gel run at 120 V for 45 min with an Invitrogen 1 kb Plus DNA Ladder. Capillary sequencing involved the incorporation of fluorescently labeled chain-terminator ddNTPs. The reaction components included amplicons, the corresponding primers, and an ABI BigDye® Terminator v3.1 Cycle Sequencing Kit (Cat No. 4337455). The cycling parameters on the thermal cycler were as follows: pre-hold at 4°C, 96°C for 1 min, 25 cycles at 96°C for 10 s, 50°C for 5 s, 62°C for 4 min, and hold at 4°C. Ethanol precipitation was performed to remove the unincorporated ddNTPs, excess primers, and primer dimers. Capillary electrophoresis was carried out on an ABI 3730xl DNA Analyzer using a 50 cm 96-capillary array, POP7 Polymer (Cat No. 4393714), and 3730xl Data Collection Software v3.1. Base calling was performed using Sequencing Analysis Software v5.4.
Whole genome sequencing
Library was prepared using a TruSeq DNA Nano Kit (Illumina, USA), and sequencing was conducted using an Illumina MiSeq instrument and a paired-end read format of 2 ׳ 150 bp for 300 cycles at the Philippine Genome Center, Quezon City, Philippines.
Genome assembly and annotation
Genome assembly was performed using BaseSpace Sequence Hub. Read quality was assessed using FastQC v1.0.0. After which, the quality of the reads was filtered using FastQ Toolkit where the base trimming, quality trimming, and read filtering settings were expanded to trim reads at the 52 -end by 15 bp and 32 -end by 15 bp, trim bases at the 52 -end and 32 -end with a quality score less than 28, and minimum quality score to 28, respectively. Genome assembly was conducted using SPAdes Genome Assembler, showing the generated contigs, scaffolds, and length of the assembly, whereas annotation of the genome was performed using Prokka Genome Annotation v.1.1.1, where tRNA, rRNA, CDS, and CRISPRs were identified from the assembly.
Stress tolerance test
The thermotolerance, pH tolerance, and alcohol tolerance of L. paracasei BCRC-16100 and L. paracasei ZFM54 were evaluated under various conditions. For thermotolerance, bacterial suspensions (3.02 × 106 CFU/mL) were prepared and inoculated on de Man Rogosa Sharpe agar (Sigma-Aldrich) and incubated at various temperatures ranging from 25°C to 50°C for 24 h. The sizes of the colonies grown on the plate were measured using the grid method. The same process was applied for testing pH tolerance using different pH levels (3, 4, 5, and 7), where the acidity of MRS agar was adjusted with 1 mol/L NaOH and HCl solutions. Alcohol tolerance was tested using MRS agar mixed with varying ethanol concentrations of 5%, 10%, 15%, and 20%.
Antimicrobial susceptibility test
The isolates L. paracasei BCRC-16100 and L. paracasei ZFM54 were subjected to antimicrobial susceptibility using commonly used antimicrobial (BD BBL™ Sensi-Disc™, BD Canada) from various classes with various pharmacological actions, namely, natural penicillins, glycopeptides, aminoglycosides, macrolides, lincosamides, and fluoroquinolones. Kirby-Bauer technique (disc-diffusion) was employed where 100 µL of bacterial culture was inoculated and spread in the plate containing sterile Mueller Hinton Agar (TM Media, India) and discs of Vancomycin (30 µg), Clindamycin (2 µg), Gentamycin (10 µg), Ofloxacin (5 µg), Erythromycin (15 µg), and Streptomycin (300 µg) were used as antimicrobial wafers. The Zone of Inhibition (ZOI) was measured after 48 h using digital calipers. The results were interpreted as follows: resistant/R (<15 mm), intermediate/I (16-20 mm), and sensitive/S (>21 mm). This test was done in triplicates, and antimicrobial-free disc plates were used as negative controls. Moreover, viable cells of the probiotic isolates suspended in Mueller Hinton Broth (TM Media, India) containing antimicrobials at known concentrations were quantified to correlate with the semi-quantitative data obtained from the disc diffusion method using a standard formula.
Carbohydrate hydrolysis
The hydrolysis of L. paracasei BCRC-16100 and L. paracasei ZFM54 was qualitatively identified through an agar well diffusion assay using a solution of 23 g of nutrient agar (TM Media, India) and 10 g of potato starch in 1 L of distilled H2O. The solidified agar was bored using sterile cork borers with diameter of +10 mm. 300 µL of de Man Rogosa Sharpe broth (Sigma-Aldrich) were placed in 3 agar wells as control. 100 µL aliquot of 48 h bacterial cultures (1-2 × 107 cells approximately) were inoculated onto wells of the starch agar, with pH 6.5. The plates were incubated separately for 24 h and 48 h at 35°C. The clearance zone was measured in mm at 24 h and 48 h using a digital caliper (Linear Tools).
Prediction of promoter elements
Bioinformatic tools such as BPROM and PRODORIC were used to predict the promoter elements involved in the expression of genes associated with antimicrobial resistance, stress tolerance, and hydrolytic activity of L. paracasei BCRC-16100 and L. paracasei ZFM54. The upstream regions of the genes were extracted from their WGS and subjected to analysis on the BPROM website, where the results show possible –10 and –35 boxes of predicted promoters, their positions in the submitted sequence, and possible transcription factors. These transcription factors were further analyzed in the virtual footprint of the PRODORIC website, in which their potential binding sites had the highest relative scores.
Gene expression and polymerase chain reaction
For gene expression, total RNA was isolated using TRIzol (Invitrogen Inc., USA) and quantified using a NanoDrop 2000 (Thermo-Fisher Inc.). Two micrograms of total RNA was treated with RQ1-RNAse free DNase (Thermo Fisher Inc.), and 1 µg of the DNase-treated RNA was used for cDNA synthesis using PrimeScript RT reagent kit (Takara Bio, CA, USA). The expression analysis was performed using TB green Premix Ex Taq II (Takara Bio, CA, USA) on Bio-Rad CFX 96 C1000 with the conditions: 95°C for 30 s and 40 cycles of 95°C for 5s + 60°C for 30 s. Product specificity was verified using melting curve analysis. The Ct values of all genes were normalized against 16S rRNA expression as a reference. The primer sequences used for gene expression are listed in Table 1. Polymerase chain reaction (PCR) was performed on genomic DNA to amplify 16S rRNA using Emerald Amp MAX PCR Master Mix (Takara Bio). The primers are presented in Table 1.
Table (1):
Gene ID from NCBI of reference sequences and primers used for PCR and gene expression analysis using qPCR of L. paracasei BCRC-16100 and L. paracasei ZFM54
| Gene ID | Gene Name_Strain | Sequence (5’-3’) |
|---|---|---|
| qPCR |
||
| P96712 | bmr3_ BCRC-16100 | Forward: TATCGGGAACCTAGTCGTGAA
Reverse: CGCCTGAATGCCAAGTAGAA |
| P71879 | stp_ BCRC-16100 | Forward: CGCTGTTGGAGCTAGATCTTT
Reverse: GAATCAGACTCGGTGCTACTAAC |
| Q9CHL8 | lmrA_ BCRC-16100 | Forward: CGTTGTGCCGCTTGTTATTC
Reverse: TAAACGCTGCCAGATTCTCTT |
| P52600 | emrY_ BCRC-16100 | Forward: CTGTTTACTGGCGGGATGAT
Reverse: CCACCAGCAAGGATGAGAAA |
| O07550 | yheI_ BCRC-16100 | Forward: GCCAACTGGTTTCCTTTGTG
Reverse: GTTCATCGACCCGATCATAACT |
| P71879 | stp_ ZFM54 | Forward: GGTCTGATCGGTTCCTTTGT
Reverse: GATCAAAGCGACAATCCCAATC |
| O07550 | yheI_ ZFM54 | Forward: GCCAACTGGTTTCCTTTGTG
Reverse: GTTCATCGACCCGATCATAACT |
| yheH_ ZFM54 | Forward: CGAAGCGACGAATGAGGAATA
Reverse: AACCGCAATCAACGCAAAG |
|
| P96712 | bmr3_ ZFM54 | Forward: CCGGGTTTCACGATTGGTAT
Reverse: GCACTCAAGCCTAATGTTTCTTC |
| tetA_1_ ZFM54 | Forward: GCATAACGTCGGTCCCATTA
Reverse: GATCGCCGTAACGACCTAATC |
|
| Q9CHL8 | lmrA_ ZFM54 | Forward: CAACCTCACCAAAGTCGTTAGA
Reverse: GCTCTTCCGCATGGTGATAA |
| tetA_2_ ZFM54 | Forward: GCATAACGTCGGTCCCATTA
Reverse: GATCGCCGTAACGACCTAATC |
|
| tetO_ ZFM54 | Forward: CACTGACGAAAGCCGTAGAA Reverse: TCGGCGGCAAAGGTTAAT |
|
| marR_ ZFM54 | Forward: GCTCAGTGAGTGCCACAATTA Reverse: CTCGCTTGTCAGTTGGATTAGG |
|
| PCR | ||
| 16S rRNA | Forward: AGAGTTTGATCMTGGCTCAG Reverse: GGTTACCTTGTTACGACTT |
|
Data analysis
Experiments subjected to Statistical analyses were performed in triplicates. To compare the bioactivity of macronutrient hydrolysis and other phenotypic characteristics between the two isolates, means were compared using Student’s t-test. Antimicrobial resistance assays were performed in a completely randomized design. Data were arcsine-transformed and subjected to one-way ANOVA. Differences in mean antimicrobial susceptibility to different antimicrobial drugs were determined using Tukey’s multiple comparison test. Statistical analyses were performed using SAS statistical software (version 9.4, SAS Institute Inc.).
Phenotypic and genomic characteristics and identification of isolates
The two LAB isolates obtained from Nipa sap were subjected to phenotypic characterization and identification (Figure 1a-c). Both bacterial isolates were gram-positive and had the shape of bacilli. Both isolates tested negative for catalase test and exhibited γ hemolysis, as no clear, greenish, or opaque zones formed around the colonies (Figure 1a). These phenotypes are common in LAB.36
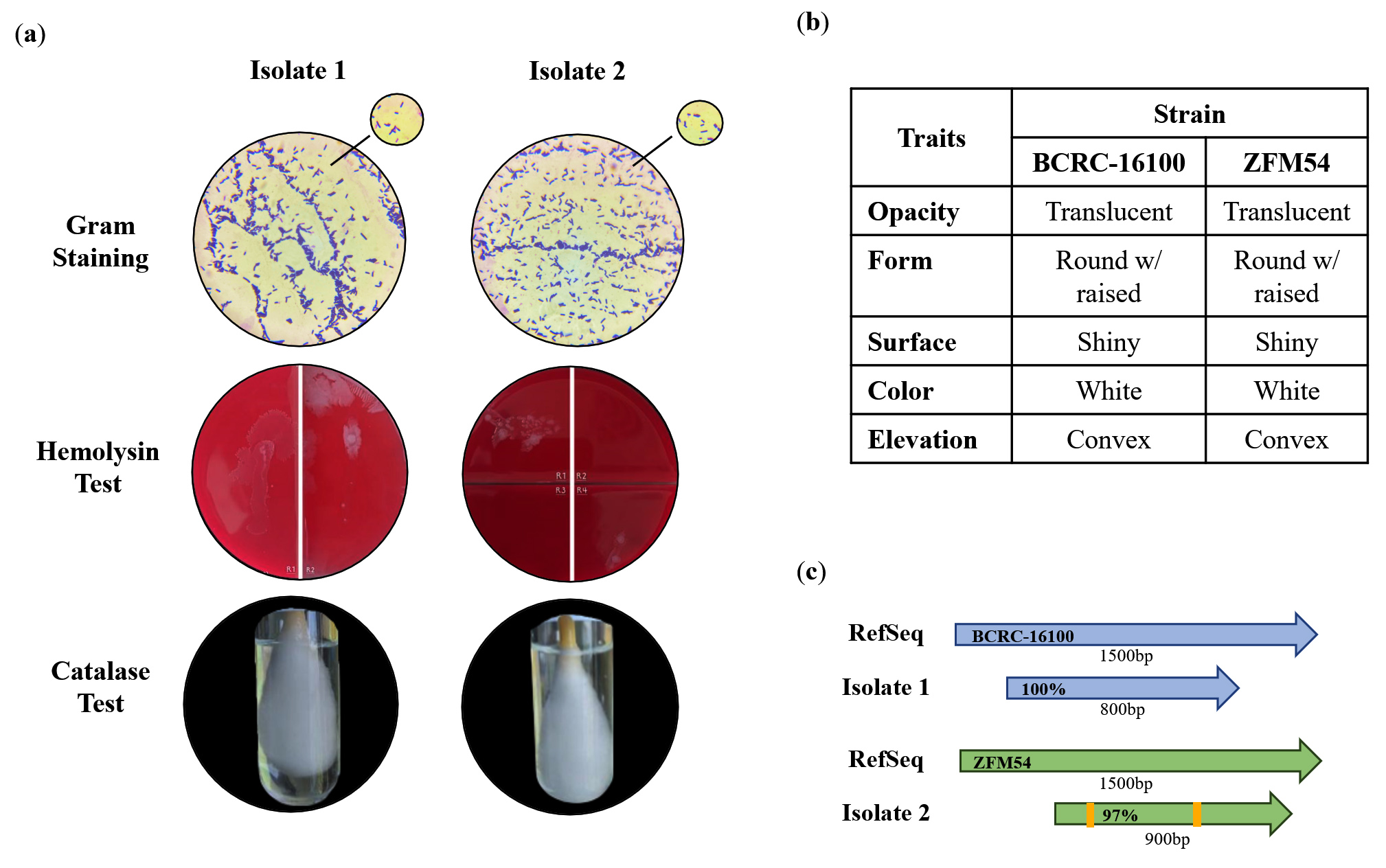
Figure 1. Phenotypic characteristics and identification of lactic acid bacterial (LAB) isolates from nipa sap, Lacticaseibacillus paracasei BCRC-16100 and Lacticaseibacillus paracasei ZFM54. (a) Tests for safety in the two isolates indicating gram-positive, hemolysin negative, catalase negative for both, (b) observable physical appearance and (c) identity using 16S rRNA. Gram stain test (Purple cell morphology = gram-positive, pink cell morphology = gram-negative); hemolysin test (yellow to translucent inhibition = β-hemolysis, translucent with greenish inhibition = α-hemolysis, no clearing zones = γ-hemolysis; catalase test (presence of bubbles = positive, absence of bubbles -negative)
Thus, it was inferred that these putative LAB are safe for further utilization as probiotics that can be incorporated into drugs or functional foods. The colonies of the two isolates were translucent, round, shiny, and convex (Figure 1b), consistent with the characteristics of a LAB isolate described in other studies.37-42 Using 16S rRNA, the isolates were identified as L. paracasei BCRC-16100 and L. paracasei ZFM54 with 100% homology to the database sequences with accession numbers NZ_CP086132 and NZ_CP032637, respectively (Figure 1c). Visualization of the whole genome of Lacticaseibacillus paracasei BCRC-16100 showed 2,885 contigs (3,029,123 bp), and Lacticaseibacillus paracasei ZFM54 showed 2,945 contigs (3,015,887 bp). In addition to differences in genome length, the two isolates also showed variability in the coding sequences on the forward strand, genomic scaffolds, coding sequences on the reverse strand, GC content, and GC skew (Figure 2a-b). The potential binding sites with the highest relative scores in the upstream regions of the identified functional genes are shown in Table 2.
Table (2):
AMR Genes identified in the genome sequences of L. paracasei BCRC-16100 and L. paracasei ZFM54, associated with their corresponding transcription factors (TF) and respective binding site scores. Core score signifies the primary score indicating the strength of the binding interaction, while relative score represents the score normalized relative to other binding interactions
Antimicrobial Resistance (AMR) Genes |
Transcription Factors |
Binding Site |
Score (Core score) |
Relative score |
|---|---|---|---|---|
stp |
phoB |
CTTTCATCAATTATATTCAGTC |
4.91
(3.35) |
0.79 |
yhel |
araC |
TATGAATTGATCTGC |
3.21
(2.92) |
0.79 |
fnr |
TTGATATAAATCAA |
3.21
(2.92) |
0.79 |
|
fis |
GTAGACGATGTAGAC |
3.21
(2.92) |
0.93 |
|
yheH |
arcA |
TGTTAAACAA |
3.62
(2.59) |
0.88 |
purR |
AAATTCGAACATTA |
3.21
(2.92) |
0.79 |
|
bmr3 |
fnr |
TTGAGGACAAACAA |
10.54
(7.72) |
0.87 |
ihf |
CAATAGTT |
4.69
(4.07) |
0.97 |
|
lrp |
CCCATTTTAA |
7.19
(6.58) |
0.98 |
|
nagC |
GTTAATTGATTTTGCGAAATAGG |
3.21
(2.92) |
0.79 |
|
tetA_1 |
fis |
GCGGAAAAAATGACC |
3.41
(2.92) |
0.99 |
arcA |
TGTTATTTAA |
3.99
(3.04) |
0.97 |
|
purR |
AAATTCGAACATTA |
3.21
(2.92) |
0.79 |
|
cpxR |
GTAAAGGTGTCTAA |
11.14
(8.91) |
0.93 |
|
crp |
TGAGCGGTATAACCGC |
6.86
(5.57) |
0.90 |
|
metR |
TGAAAATTTTTCA |
3.21
(2.92) |
0.79 |
|
lmrA |
ihf |
CAATTGTT |
4.69
(4.07) |
0.97 |
lrp |
CAGCCTTTTATC |
2.10
(1.63) |
0.91 |
|
cpxR |
GTAATTTAGGTAA |
3.21
(2.92) |
0.79 |
|
fhlA |
GTAATGGTCCAAAGTC |
3.21
(2.92) |
0.79 |
|
ompR |
TTTAACACTAAA |
8.58
(7.07) |
0.99 |
|
tetA-2 |
purR |
ATTTAAACGGTTGCGT |
3.21
(2.92) |
0.79 |
argR |
CGAATTACCATACT |
7.92
(5.32) |
0.80 |
|
tetO |
purR |
ATTTAAACGGTTGCGT |
3.21
(2.92) |
0.79 |
lrp |
CGTTTTTTAG |
7.27
(6.58) |
0.99 |
|
argR |
CTAATTTCTATCCA |
7.99
(6.21) |
0.80 |
|
dnaA |
TTATCCACA |
3.21
(2.92) |
0.79 |
|
marR |
fnr |
TTGTTGTCGAACAC |
10.46
(7.72) |
0.86 |
emrY |
Ihf |
CAAGGGTT |
4.69
(4.07) |
0.97 |
glpR |
GTTCGGTTAAAAACAGA |
8.72
(6.52) |
0.85 |
|
phoB |
ATAACATACTTAAGTTACTAAT |
5.07
(3.28) |
0.81 |
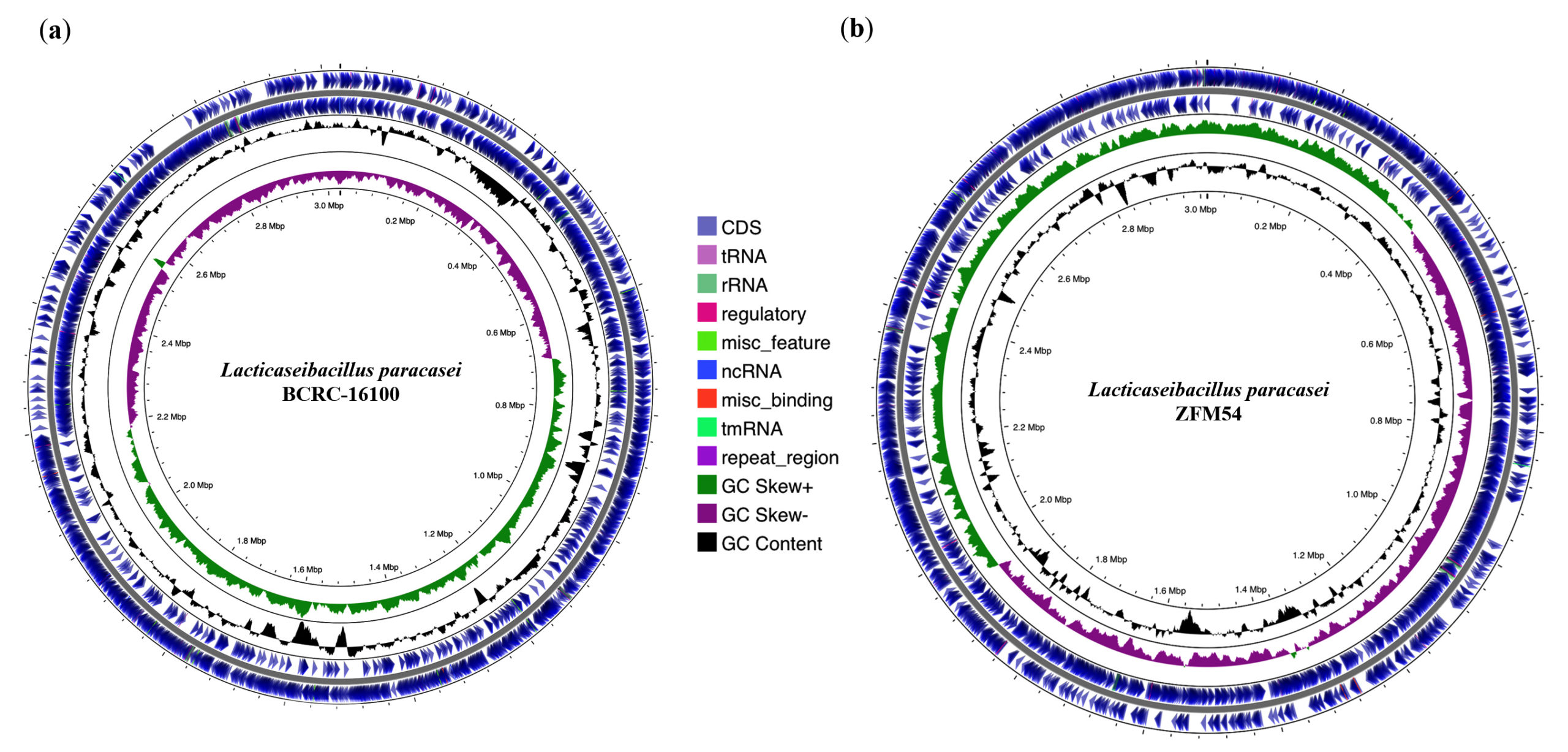
Figure 2. Whole genome sequence (WGS) assembly and analysis of Lacticaseibacillus paracasei BCRC-16100 and Lacticaseibacillus paracasei ZFM54. (a) Visualization of regions of interest in the whole genome of L. paracasei BCRC-16100 showing 2,885 contigs (3,029,123 bp) and (b) Visualization of regions of interest in the whole genome of L. paracasei ZFM54 showing 2,945 features (3,015,887 bp). Circles from the outside to the center illustrate the following characteristics: (1) coding sequences on forward strand, (2) coding sequences on reverse strand, (3) GC content, (4) GC skew, and (5) genomic scaffolds
Tolerance in different conditions
Figure 3a shows that L. paracasei ZFM54 grew within a temperature range of 35-45°C, with 45°C as the optimal temperature. The optimal temperature for the growth L. paracasei BCRC-16100 was the same, but there was a significant decline in growth at any deviation in temperature (±1°C). However, it has been previously reported that the optimal growth of different Lactobacillus spp. occurs at 37°C.43 The growth of the two isolates was similar when tested at different concentrations of alcohol, suggesting the ability of the isolates to survive when incorporated into functional food products or drugs with alcohol content ranging from 5% to 20% (Figure 3b). For both probiotic isolates, growth at pH 5.0, and 7.0, was significantly higher than that at pH 3 and 4 (Figure 3c). This is consistent with the result of a previous study indicating that the optimal pH range for the growth of Lactobacillus spp. is 5.5-6.5 which may vary depending on the strain.43
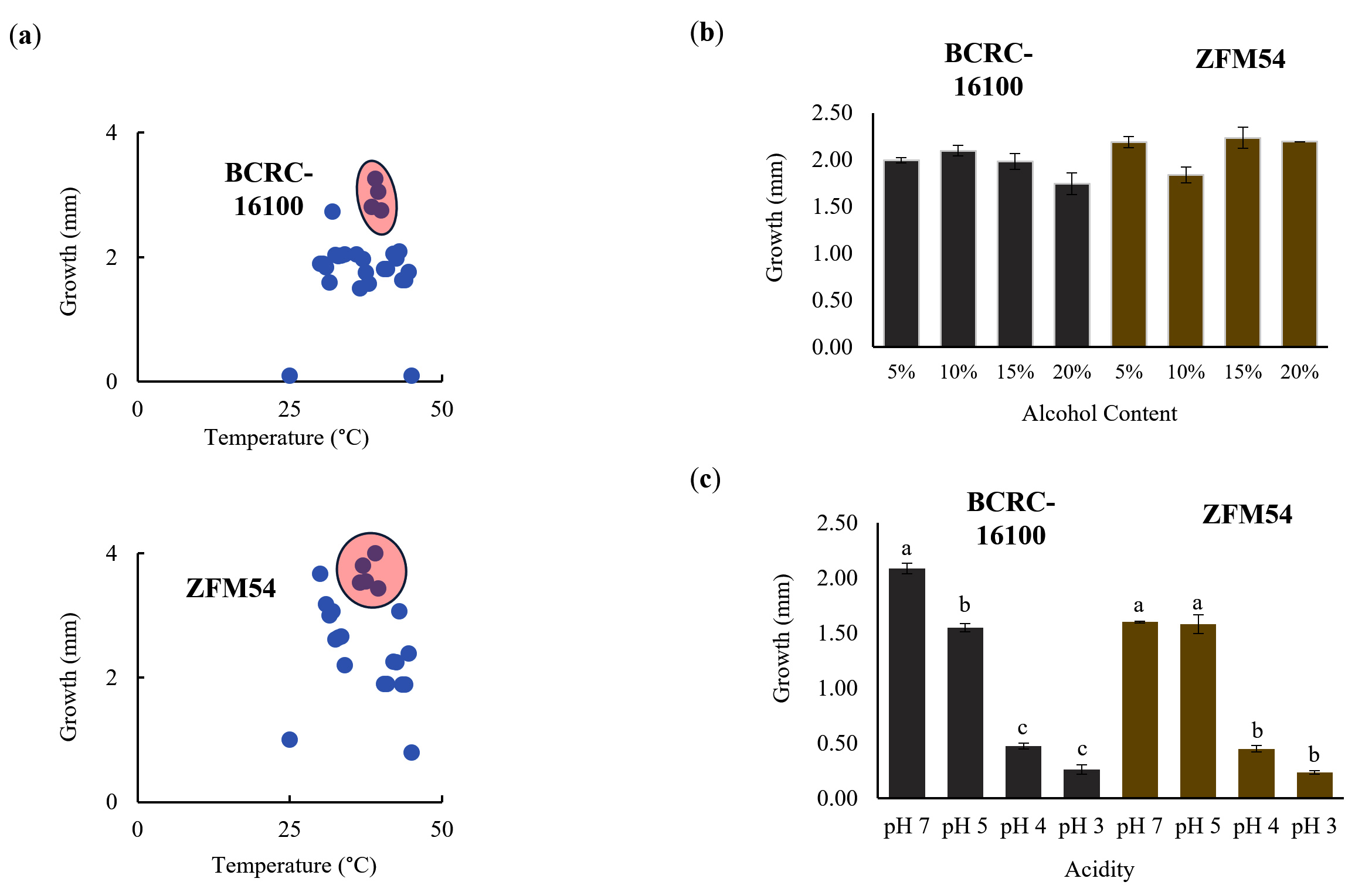
Figure 3. Tolerance of lactic acid bacterial isolates from nipa sap, Lacticaseibacillus paracasei BCRC-16100 and Lacticaseibacillus paracasei ZFM54. L. paracasei ZFM54 has wider range of tolerance across (a) temperature and (b) presence of alcohol while (c) L. paracasei BCRC-16100 performs better in a range of pH. Means within each bar (comparison within the same strain) having the same letter are not significantly different in Tukey’s multiple comparison at α = 0.05. and significant for condition as source of variation at p < 0.01. Error bars represent standard error within 3 biological replicates of each type of condition
The differences in the frequency of stress tolerance genes and variability in the type and number of transcription factors (TFs) binding to cis-regulatory elements offer an explanation for the differential or similar growth responses of the two isolates in a particular stressful condition (Figure 4a-c). The growth of L. paracasei ZFM54 is less influenced by temperature, and a higher frequency of TFs that potentially bind to the promoter region of the thermoregulatory gene dnaK may play a significant role in this tolerance. A previous study showed that in known thermotolerant Bacillus pumilus, dnaK is highly expressed.44 Although it cannot be ruled out that there may be differences in dnaK of different species and strains, other genes present in the genomes of the two isolates play synergistic or antagonistic roles in conferring tolerance to certain temperatures.
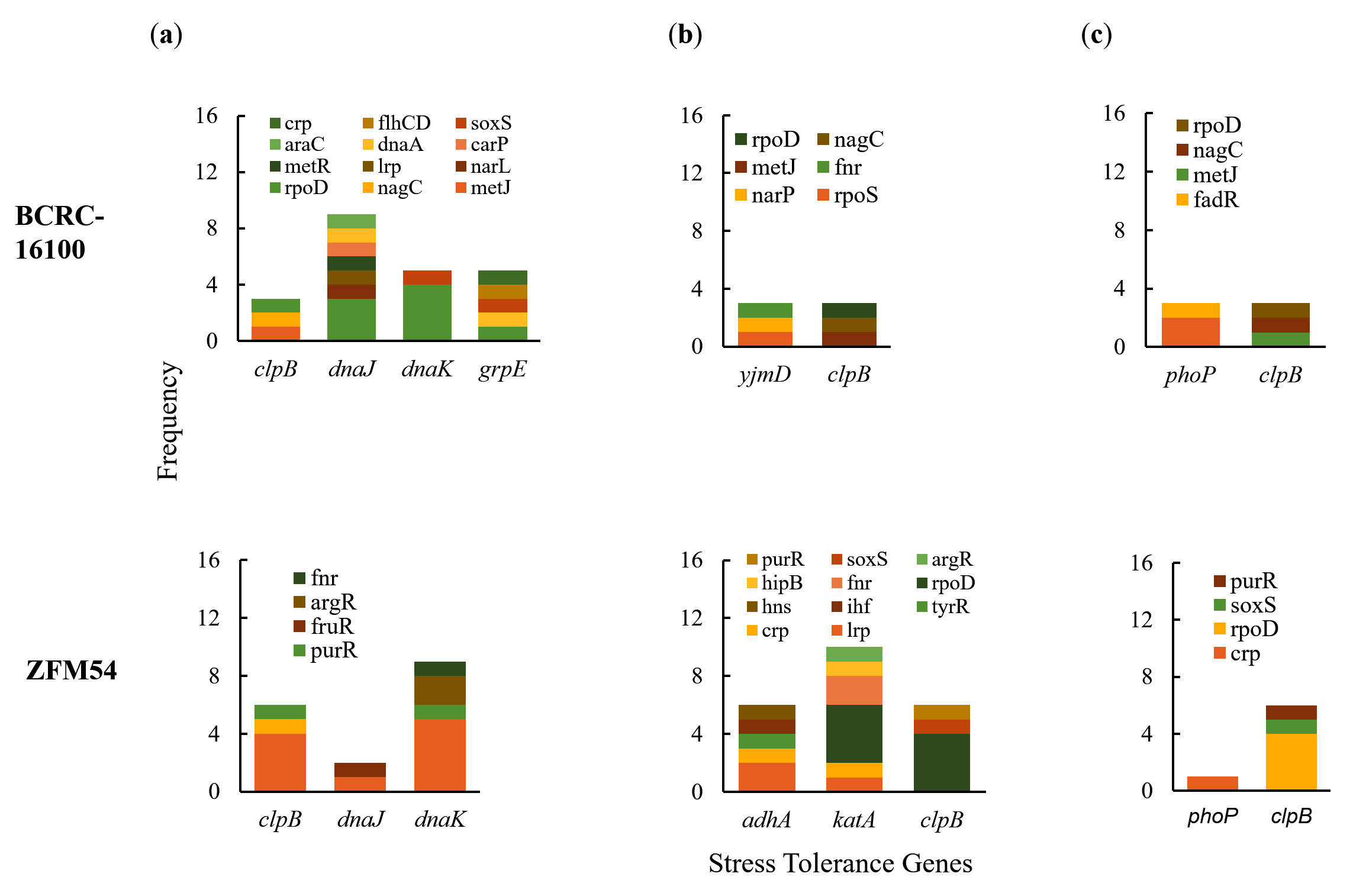
Figure 4. Promoter analysis of genes associated to (a) thermoregulation, (b) alcohol tolerance, and (c) pH response in Lacticaseibacillus paracasei BCRC-16100 and Lacticaseibacillus paracasei ZFM54. There is differential frequency in the number of tolerance genes and transcription factors binding to each gene responsible for stress response. Colors in the bars indicate the number of each type of transcription factors (TF) binding to a stress tolerance gene
Antimicrobial resistance
The antimicrobial resistance of the two probiotic isolates showed a similar pattern for the six types of antibiotics both at 24 h and 48 h post-treatment (Figure 5a-b). At 24 h, L. paracasei BCRC-16100 and L. paracasei ZFM54, showed complete resistance to clindamycin, vancomycin, gentamicin, ofloxacin, erythromycin, and streptomycin. However, at 48 h post-treatment, five antimicrobials (clindamycin, gentamycin, ofloxacin, erythromycin, and streptomycin) inhibited the growth of the two isolates, except for vancomycin (Figure 5a). Differences in the zones of inhibition (mm) were significantly different (p = 0.0005, =0.05) for the various antimicrobials for each of the two isolates (Figure 5b). A larger zone of inhibition was observed for erythromycin in both isolates. The results of disk diffusion were corroborated by the colony-forming unit quantity (CFU/mL) when the two isolates were grown independently in liquid broth supplemented with antimicrobials for 48 h. The CFU of both isolates was significantly higher in the presence of vancomycin (BCRC-16100 = 6.330413773 and ZFM54 = 5.252853031), suggesting resistance to this antimicrobial and supporting the results of the disk diffusion analysis. These analyses indicated the differential resistance of L. paracasei BCRC-16100 and L. paracasei ZFM54 depending on the type of antimicrobial and the time-of-exposure. Furthermore, this suggests the prevalence of AMR genes with active cis-regulatory elements in these isolates, enhancing their survival against antibacterial agents.
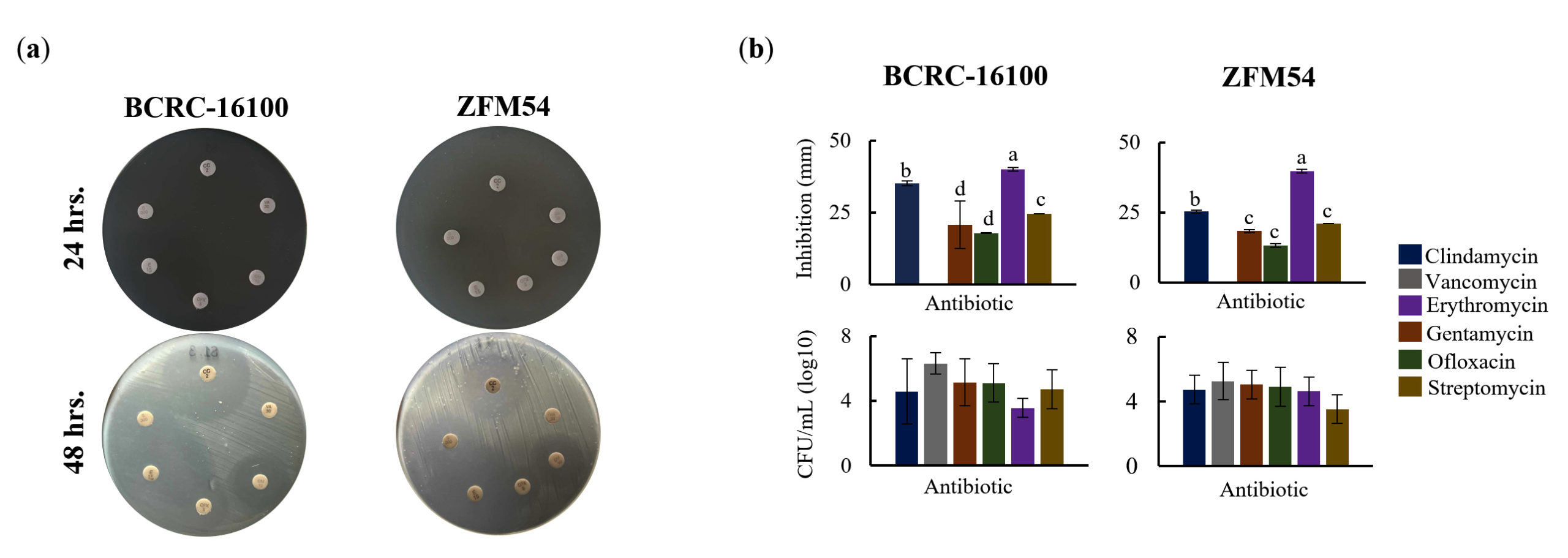
Figure 5. Higher antimicrobial resistance in Lacticaseibacillus paracasei ZFM54 compared to Lacticaseibacillus paracasei BCRC-16100 suggested by higher growth in six (6) types of atimicrobials and expression of antimicrobial resistance (AMR) genes. (a) disk diffusion assay, zone of inhibition in antimicrobial disks and (b) and colony forming units (CFU) of the two isolates in different antimicrobial at 12 h. From top clockwise in (a) the antimicrobials are clindamycin, vancomycin, gentamycin, ofloxacin, erythromycin, and streptomycin. Means within each bar (comparison within the same strain) having the same letter are not significantly different in Tukey’s multiple comparison at =0.05. and significant for type of antimicrobial as source of variation at p ≤ 0.01. Error bars represent standard error within 3 biological replicates of each type of condition
Genomic Analysis of Antimicrobial Resistance (AMR) genes
AMR genes were present in both probiotic isolates and differed in terms of frequency, activity, and genomic and amino acid structures (Figure 6a-d). There were five (bmr3, stp, lmrA, emrY, yheI) and nine (stp, yheI, yheH, bmr3, tetA_1, lmrA, tetA_2, tetO, marR) AMR genes in L. paracasei BCRC-16100 and L. paracasei ZFM54, respectively (Figure 6a). Briefly, there are two mechanisms of AMR (transcriptional regulation and transporter activity) in L. paracasei BCRC-16100, whereas there are three mechanisms in L. paracasei ZFM54, suggesting that the latter has a wider range of resistance to antimicrobials (transcriptional regulation, transporter activity, and efflux system). In L. paracasei BCRC-16100, the AMR genes are mostly responsible for encoding transporter protein subunits (bmr3, stp, emrY, yheI), and one is a transcriptional repressor (lmrA). The two isolates shared four AMR genes, namely bmr3, stp, lmrA, yheI and unique AMR genes in L.paracasei ZFM54 include yheH, tetA_1, tetA_2, tetO, and marR where three are efflux pumps (tetA_1, tetA_2, and tetO), one encodes a transcriptional regulator (marR), and the other encodes a transporter protein subunit (yheH).
Carbohydrate hydrolytic activity
Although four AMR genes were shared between L. paracasei BCRC-16100 and L. paracasei ZFM54, analysis of the homology of these four genes relative to each other and to other AMR genes, indicated differences. Among the four shared AMR genes in the two isolates, yheI and stp were the most divergent, as suggested by the distances between their clusters or clades in the phylogenetic analysis (Figure 6b). Given the overlap and diversity of similar AMR genes in the two isolates, genes responsible for AMR were tested for differential gene expression analysis in the presence of vancomycin (Figure 6c), an antimicrobial agent to which both isolates showed resistance (Figure 5a). In both isolates, AMR genes encoding transcriptional regulators (lmrA and marR) were upregulated after vancomycin treatment. This suggests a similar mechanism for vancomycin resistance in Enterococci where vancomycin activates VanR which then activates other AMR genes.45 Other AMR genes that were significantly upregulated by vancomycin were emrY and yheI which encode transporter protein subunits and tetA_1, which encodes an efflux pump. Among the AMR genes, only emrY has the highest probability to be transposed laterally due to the presence of transposable elements in the -54 bp (52 ) and +637 bp (32 ) of the gene (Figure 6d).
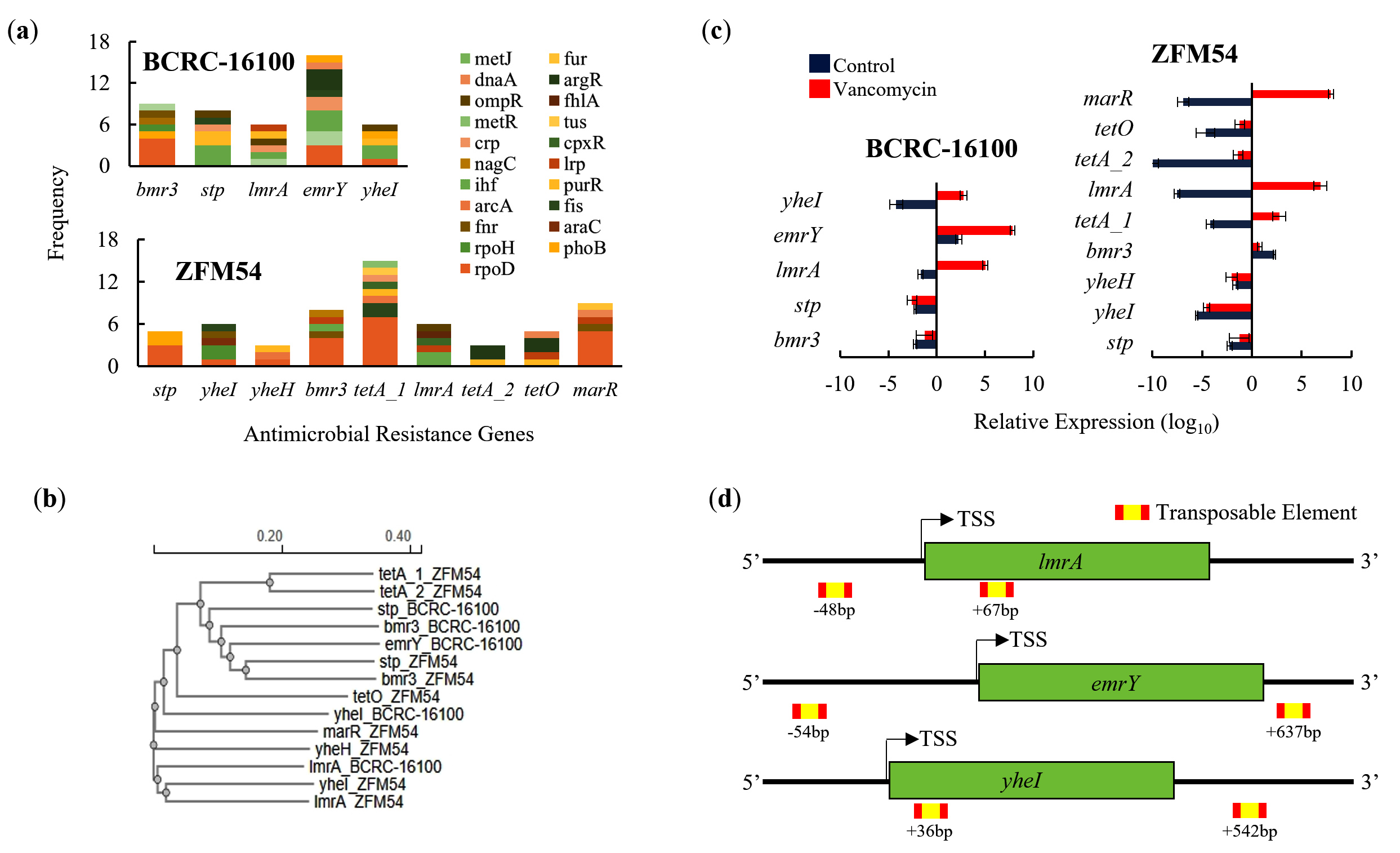
Figure 6. Promoter, phylogeny, gene expression, and transposable elements of antimicrobial resistance (AMR) genes in Lacticaseibacillus paracasei BCRC-16100 and Lacticaseibacillus paracasei ZFM54. (a) AMR genes and frequency of each type of transcription factor (TF) binding in the promoter region (1000 bp) in every gene. More resistance genes are found in the genome of L. paracasei ZFM54 and the frequency of TFs that can bind to each of the AMR genes promoter region vary. (b) Phylogeny of AMR genes showing differential clustering of some similar genes (stp and bmr3) shared by the two isolates. (c) Expression of AMR genes in the two isolates. (d) Transposable elements located in the upstream (5’) and downstream (3’) regions (500 bp) of AMR genes. Colors in the bars indicate the number of each type of TF binding to a stress tolerance gene. Complete list of AMR genes with the cis-regulatory elements where TFs bind is shown. Relative expression is against the internal control (16S rRNA) is shown. Transcription Start Site (TSS). Error bars represent standard error within 3 biological replicates
Carbohydrate hydrolytic activity
The two isolates were analyzed for their ability to hydrolyze carbohydrates, TF binding to the promoter, and the expression of genes responsible for hydrolysis (Figure 7). In the starch assay, both isolates showed qualitative and quantitative hydrolytic activities (Figure 7a) and (Figure 7b). Multiple genes were found in the WGS, which may be responsible for carbohydrate hydrolytic activity, with eight and 11 genes in L. paracasei BCRC-16100 and L. paracasei ZFM54, respectively (Figure 7c). Notably, promoter analysis revealed increased TF binding to bga, lacG and bglA suggesting the importance of these three genes in carbohydrate hydrolysis. However, gene expression analysis revealed that during carbohydrate hydrolysis, rafA, malL, and bglC genes were upregulated (Figure 7d). It can be argued that despite the presence of more cis-regulatory elements in some carbohydrate hydrolysis genes, the observed difference in expression can be explained by the positional effect of where the TF binds, which has been previously described to influence gene activity.46
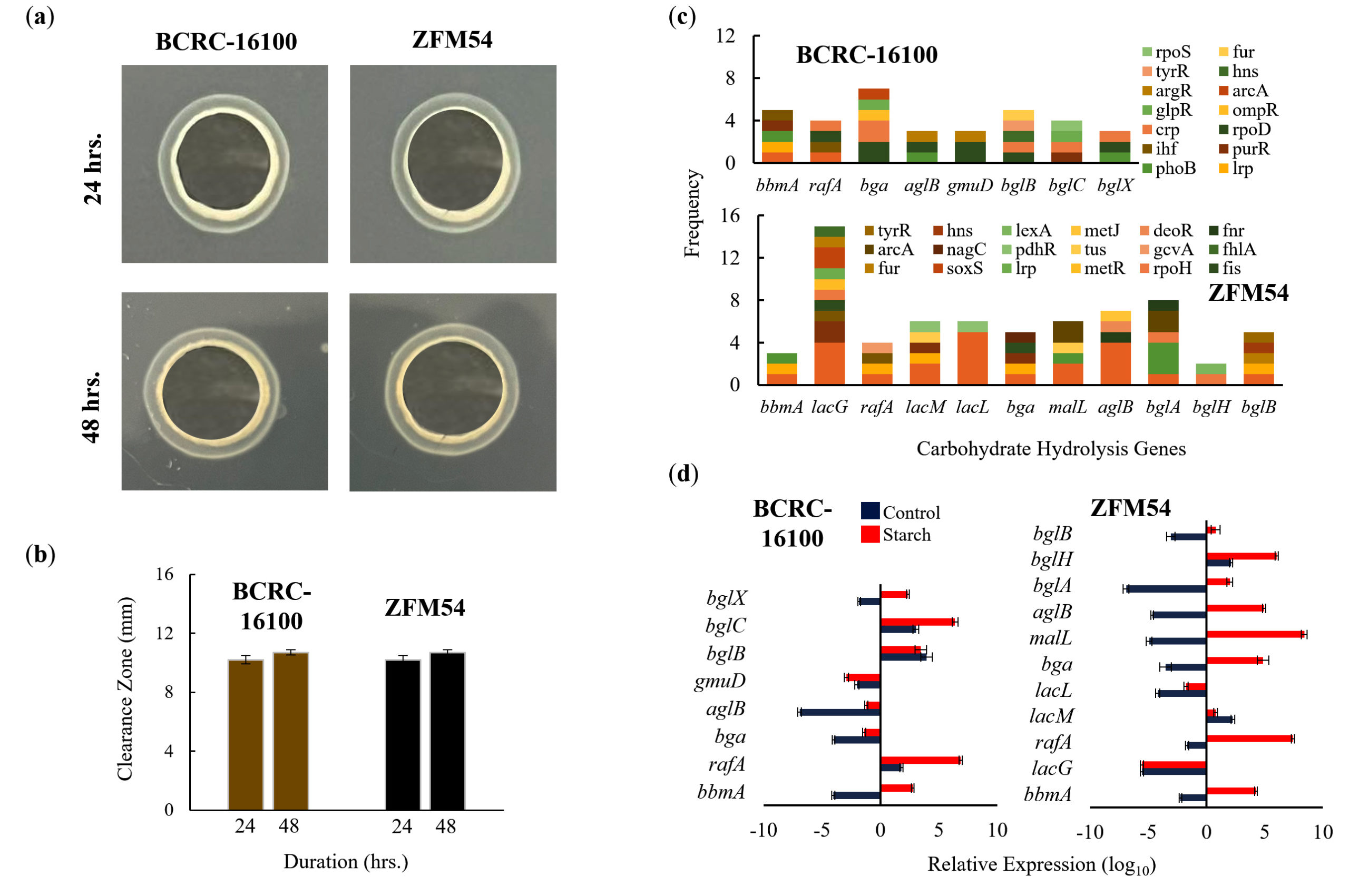
Figure 7. Carbohydrate hydrolytic activity of Lacticaseibacillus paracasei BCRC-16100 and Lacticaseibacillus paracasei ZFM54. (a) Amylase activities of BCRC-16100 and ZFM54 presented by zone of clearance, (b) the quantitative zone of clearance expressed in mm, (c) promoter analysis of carbohydrate hydrolysis genes, and (d) expression of amylases genes. Colors in the bars indicate the number of each type of transcription factors (TF) binding to a stress tolerance gene. Relative expression is against the internal control (16S rRNA) is shown. Error bars represent standard error within 3 biological replicates
Probiotics can be isolated from animal and dairy sources, and novel or similar genotypes obtained from different sources may have different characteristics.47-49 Studies exploring plants as sources of probiotic isolates are limited.50
A diverse range of indigenous food plants (IFPs) in the northern Philippines can be utilized as sources of probiotics, such as Nipa. Two strains of the same probiotic species were isolated from this source namely L. paracasei BCRC-16100 and L. paracasei ZFM54. Some known health benefits of the two strains include amelioration of allergic airway, antimicrobial activity, enhancement of intestinal microbiota, and stress modulation.51-53 The intrinsic heterogeneity across strains of L. paracasei makes the species a strong candidate for the development of probiotic isolates that offer health benefits and lead to the development of drugs or functional foods for the abovementioned purposes.54,55 Although there have been previous efforts to identify the potential health benefits of L. paracasei, reports have been inconclusive and do not offer strong evidence to support product development initiatives.
To assess the potential of L. paracasei BCRC-16100 and L. paracasei ZFM54 as commercial probiotics, WGS, gene expression, and biochemical analyses were performed. Several genes confer thermotolerance in both isolates, which allow growth at a wide range of temperatures, and dnaK and dnaJ are likely responsible for this trait.56 These genes show the highest potential for regulation through the association of promoter regions with multiple transcription factors (TFs). Both dnaK and dnaJ belong to a chaperone system in microbes, that is known to confer a wide tolerance to temperature.57 The two isolates grew optimally at pH 5-7, however, promoter analysis of genes responsible for this trait suggested that clpB is responsible for differential growth depending on pH. The gene clpB is highly conserved in bacteria and provides tolerance to oxidative stress, nutrient starvation, and low pH.58-60
In addition to flexibility under stressful conditions, we explored the AMR genes in each strain along with their corresponding molecular mechanisms and transposability. The two strains were highly resistant to vancomycin, and gene expression analysis suggested the involvement of four genes in conferring this resistance, namely, lmrA, marR, emrY and yheI, and the mechanisms of resistance, included efflux pump, transcriptional control, and transport regulation (Figure 6). Moreover, these AMR genes possess transposable elements in the upstream (52) and downstream (32) region. These strains can be used as probiotics to avoid potential lateral transfer of AMR genes to pathogenic microbes.61
Several genes that encode carbohydrate-active enzymes were determined using WGS, which correlated with enzymatic hydrolytic activities in vitro. Among the identified genes, rafA, malL, and bglC were most likely responsible for the carbohydrate-hydrolytic activities of L. paracasei BCRC-16100 and L. paracasei ZFM54 (Figure 7). Moreover, the abovementioned genes were upregulated at the transcript-level when the two isolates were cultured with starch.
In conclusion, L. paracasei BCRC-16100 and L. paracasei ZFM54 can be used as probiotics that can be incorporated into drugs and functional foods to promote the digestion of carbohydrates in food products with low digestibility, but with careful consideration of the AMR genes present in these strains.
ACKNOWLEDGMENTS
The authors are thankful to the Commission on Higher Education (CHED), Philippines for supporting the project. The authors are also thankful to Dr. Policarpio B. Joves for providing a microbiology laboratory, and the Philippine Genome Center of the University of the Philippines Diliman for their extension program titled, Whole Genome Sequence @ Your Fingertips.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
This project was supported by the Commission on Higher Education (CHED) with grant number LAKAS 2022-005.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- de Brito Alves JL, de Sousa VP, Neto MPC, et al. New insights on the use of dietary polyphenols or probiotics for the management of arterial hypertension. Front Physiol. 2016;7:448.
Crossref - Hamad G, Ombarak RA, Eskander M, et al. Detection and inhibition of Clostridium botulinum in some Egyptian fish products by probiotics cell-free supernatants as bio-preservation agents. LWT. 2022;163:113603.
Crossref - Roobab U, Batool Z, Manzoor MF, Shabbir MA, Khan MR, Aadil RM. Sources, formulations, advanced delivery and health benefits of probiotics. Curr Opin Food Sci. 2020;32:17-28.
Crossref - Lin MY, Savaiano DA, Harlander S. Influence of nonfermented dairy products containing bacterial starter cultures on lactose maldigestion in humans. J Dairy Sci. 1991;74(1):87-95.
Crossref - Kim HS, Gilliland SE. Lactobacillus acidophilus as a dietary adjunct for milk to aid lactose digestion in humans. J Dairy Sci. 1983;66(5):959-966.
Crossref - Lin M, Yen CL, Chen SH. Management of lactose maldigestion by consuming milk containing Lactobacilli. Dig Dis Sci. 1998;43(1):133-137.
Crossref - Vitellio P, Celano G, Bonfrate L, Gobbetti M, Portincasa P, De Angelis M. Effects of Bifidobacterium longum and Lactobacillus rhamnosus on gut microbiota in patients with lactose intolerance and persisting functional gastrointestinal symptoms: A randomized, double-blind, cross-over study. Nutrients. 2019;11(4):886.
Crossref - De Vuyst L, Leroy F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifidobacterial competitiveness, butyrate production, and gas production. Int J Food Microbiol. 2011;149(1):73-80.
Crossref - Turroni F, Milani C, Duranti S, Mahony J, van Sinderen D, Ventura M. Glycan utilization and cross-feeding activities by bifidobacteria. Trends Microbiol. 2018;26(4):339-350.
Crossref - Riviere A, Selak M, Lantin D, Leroy F, De Vuyst L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front Microbiol. 2016;7:979.
Crossref - Veloorvalappil NJ, Robinson BS, Pradeep S, et al. Versatility of microbial proteases. Adv Enzyme Res. 2013;01(03):39-51.
Crossref - Rao MB, Tanksale AM, Ghatge MS, Deshpande VV. Molecular and biotechnological aspects of microbial proteases. Microbiol Mol Biol Rev. 1998;62(3):597-635.
Crossref - Erginkaya Z, Konuray-Altun G. Potential biotherapeutic properties of lactic acid bacteria in foods. Food Bioscience. 2022;46:101544.
Crossref - Nataraj BH, Ali SA, Behare PV, Yadav H. Postbiotics-parabiotic: the new horizons in microbial biotherapy and functional foods. Microbial Cell Factories. 2020;19(1).
Crossref - Stiles ME. Biopreservation by lactic acid bacteria. Antonie Van Leeuwenhoek. 1996;70(2-4):331-345.
Crossref - Zhao N, Su Y, Jian C, et al. Lactobacillus ameliorates SD-Induced stress responses and gut dysbiosis by increasing the absorption of Gut-Derived GABA in rhesus monkeys. Front Immunol. 2022;13:15393.
Crossref - Gareau MG, Sherman PM, Walker WA. Probiotics and the gut microbiota in intestinal health and disease. Nat Rev Gastroenterol Hepatol. 2010;7(9):503-514.
Crossref - Dowarah R, Verma AK, Agarwal N. The use of Lactobacillus as an alternative of antibiotic growth promoters in pigs: A review. Anim Nutr. 2017;3(1):1-6.
Crossref - Hooper LV, Gordon JI. Commensal host-bacterial relationships in the gut. Science. 2001;292(5519):1115-1118.
Crossref - Yang SC, Lin CH, Sung CH, Fang JY. Antibacterial activities of bacteriocins: application in foods and pharmaceuticals. Front Microbiol. 2014;5.
Crossref - Servin AL. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol Rev. 2004;28(4):405-440.
Crossref - Konings WN, Kok J, Kuipers OP, Poolman B. Lactic acid bacteria: the bugs of the new millennium. Curr Opin Microbiol. 2000;3(3):276-282.
Crossref - Cotter PD, Hill C, Ross RP. Bacteriocins: Developing innate immunity for food. Nat Rev Microbiol. 2005;3(10):777-788.
Crossref - Preidis GA, Versalovic J. Targeting the human microbiome with antibiotics, probiotics, and prebiotics: Gastroenterology enters the metagenomics era. Gastroenterology. 2009;136(6):2015-2031.
Crossref - Thomas CM, Versalovic J. Probiotics-host communication. Gut Microbes. 2010;1(3):148-163.
Crossref - Frank DN, Zhu W, Sartor RB, Li E. Investigating the biological and clinical significance of human dysbioses. Trends Microbiol. 2011;19(9):427-434.
Crossref - Roe AL, Boyte ME, Elkins CA, et al. Considerations for determining safety of probiotics: A USP perspective. Regul Toxicol Pharmacol. 2022;136:105266.
Crossref - Fossi BT, Ndjouenkeu R. Probiotic potential of thermotolerant lactic acid bacteria isolated from “Gari “a cassava-based African fermented food. J Appl Biol Biotechnol. 2017;5(4):50401.
Crossref - Ouwehand AC, Forssten S, Hibberd AA, Lyra A, Stahl B. Probiotic approach to prevent antibiotic resistance. Ann Med. 2016;48(4):246-255.
Crossref - Gueimonde M, Sanchez B, de Los Reyes-Gavilan CG, Margolles A. Antibiotic resistance in probiotic bacteria. Front Microbiol. 2013;4:202.
Crossref - Jindal AK, Pandya K, Khan ID. Antimicrobial resistance: A public health challenge. Med J Armed Forces India. 2015;71(2):178-181.
Crossref - Zommiti M, Feuilloley MGJ, Connil N. Update of Probiotics in Human World: A nonstop source of benefactions till the end of time. Microorganisms. 2020;8(12):1907.
Crossref - Zielinska D, Rzepkowska A, Radawska A, Zielinski K. In vitro screening of selected probiotic properties of Lactobacillus strains isolated from traditional fermented cabbage and cucumber. Curr Microbiol. 2014;70(2):183-194.
Crossref - Linares-Morales JR, Cuellar-Nevarez GE, Rivera-Chavira BE, Gutierrez Mendez N, Perez-Vega SB, Nevarez-Moorillon GV. Selection of lactic acid bacteria isolated from fresh fruits and vegetables based on their antimicrobial and enzymatic activities. Foods. 2020;9(10):1399.
Crossref - Nguyen J, Lara-Gutierrez J, Stocker R. Environmental fluctuations and their effects on microbial communities, populations and individuals. FEMS Microbiol Rev.2020;45:fuaa068.
Crossref - Yasmin I, Saeed M, Khan WA, et al. In vitro probiotic potential and safety evaluation (hemolytic, cytotoxic activity) of Bifidobacterium strains isolated from raw camel milk. Microorganisms. 2020;8(3):354.
Crossref - Siregar MTP, Kusdiyantini E, Rukmi MI. Isolasi dan karakterisasi bakteri asam laktat pada pangan fermentasi mandai. Diponegoro University. 2014;3(2).
- Manzoor A, Ul-Haq I, Baig S, Qazi JI, Seratlic S. Efficacy of locally isolated lactic acid bacteria against antibiotic-resistant uropathogens. Jundishapur J Microbiol. 2016;9(1):18952.
Crossref - Mahulette F, Mubarik NR, Suwanto A, Widanarni W. Microbiological and physicochemical characteristics of inasua traditional fish fermented from maluku islands. Biosaintifika. 2018;10(2):298-305.
Crossref - Rahayu HM, Qurbaniah M. Selection of Tempoyak lactic acid bacteria as candidate strain for yoghurt starter culture. Biosaintifika. 2019;11(1):39-46.
Crossref - Nasution MHB, Ramadhani S, Fachrial E. Isolation, characterization and antibacterial activities of lactic acid bacteria isolated from Batak’s special food “Dali ni Horbo”. Jurnal Natur Indonesia. 2020;18(1):1.
Crossref - Nasri, Harahap U, Silalahi J, Satria D. Antibacterial activity of lactic acid bacteria isolated from Dengke Naniura of Carp (Cyprinus carpio) against diarrhea-causing pathogenic bacteria. Biodiversitas. 2021;22(8).
Crossref - Slizewska K, Chlebicz-Wojcik A. Growth kinetics of probiotic lactobacillus strains in the alternative, cost-efficient semi-solid fermentation medium. Biology. 2020;9(12):423.
Crossref - Kumar M, Prasanna R, Lone SA, Padaria JC, Saxena AK. Cloning and expression of dnaK gene from Bacillus pumilus of hot water spring origin. Appl Transl Genom. 2014;3(1):14-20.
Crossref - Guffey AA, Loll PJ. Regulation of resistance in vancomycin-resistant enterococci: the VANRS two-component system. Microorganisms. 2021;9(10):2026.
Crossref - Investigators P, Groups N, Leads DP, Analysts L. An integrated encyclopedia of DNA elements in the human genome. Nature. 2012;489(7414):57-74.
Crossref - Lin DC. Probiotics as functional foods. Nutr Clin Pract. 2003;18(6):497-506.
Crossref - Salminen S, Gueimonde M, Isolauri E. Probiotics that modify disease risk. J Nutr. 2005;135(5):1294-1298.
Crossref - Abdou AM, Hedia RH, Omara ST, El-Fatah MMA, Kandil MM, Bakry MA. Interspecies comparison of probiotics isolated from different animals. Vet World. 2018;11(2):227-230.
Crossref - Karami S, Roayaei M, Hamzavi H, et al. Isolation and identification of probiotic Lactobacillus from local dairy and evaluating their antagonistic effect on pathogens. Int J Pharm Investig. 2017;7(3):137-141.
Crossref - Haghshenas B, Nami Y, Almasi A, et al. Isolation and characterization of probiotics from dairies. Iran J Microbiol. 2017;9(4):234-243.
- Menendez E, Garcia-Fraile P. Plant probiotic bacteria: solutions to feed the world. AIMS Microbiol. 2017;3(3):502-524.
Crossref - Lin EK, Chang WW, Jhong JH, Tsai WH, Chou C, Wang IJ. Lacticaseibacillus paracasei GM-080 ameliorates allergic airway inflammation in children with allergic rhinitis: from an animal model to a double-blind, randomized, placebo-controlled trial. Cells. 2023;12(5):768.
Crossref - Hill D, Sugrue I, Tobin C, Hill C, Stanton C, Ross RP. The Lactobacillus casei Group: history and health related applications. Front Microbiol. 2018;9:2107.
Crossref - Jones RM. The use of Lactobacillus casei and Lactobacillus paracasei in clinical trials for the improvement of human health. Academic Press. 2017:99-108.
Crossref - Balzaretti S, Taverniti V, Guglielmetti S, et al. A novel rhamnose-rich hetero-exopolysaccharide isolated from Lactobacillus paracasei DG activates THP-1 human monocytic cells. Appl Environ Microbiol. 2017;83(3):e02702.
Crossref - Kim JS, Liu L, Vazquez-Torres A. The DNAK/DNAJ chaperone system enables RNA Polymerase-DKSA complex formation in Salmonella experiencing oxidative stress. MBio. 2021;12(3):03443-20.
Crossref - Meibom KL, Dubail I, Dupuis M, et al. The heat-shock protein ClpB of Francisella tularensis is involved in stress tolerance and is required for multiplication in target organs of infected mice. Mol Microbiol. 2008;67(6):1384-1401.
Crossref - Glaza P, Ranaweera CB, Shiva S, et al. Repurposing p97 inhibitors for chemical modulation of the bacterial ClpB-DnaK bichaperone system. J Biol Chem. 2021;296:100079.
Crossref - Tripathi P, Singh LK, Kumari S, Hakiem OR, Batra JK. ClpB is an essential stress regulator of Mycobacterium tuberculosis and endows survival advantage to dormant Bacilli. Int J Med Microbiol. 2020;310(3):151402.
Crossref - Sieber KB, Bromley RE, Hotopp JCD. Lateral gene transfer between prokaryotes and eukaryotes. Exp Cell Res. 2017;358(2):421-426.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.