


ISSN: 0973-7510
E-ISSN: 2581-690X
The seed kernels of Caesalpinia bonducella, a traditional medicinal plant in India, are widely used to treat various disorders, including polycystic ovary syndrome. The seed kernel possesses anti-bacterial properties against many pathogenic bacteria. However, their impact on Lactobacillus spp., a prominent gram-positive gut bacterium, has not been studied till date. The present study employed both in vitro and in silico methods to illustrate the effect of seed extract of C. bonducella against Lactobacillus rhamnosus GG. For this, disc diffusion assay was performed with 100, 500, and 1000 µg/ml of aqueous and methanolic seed extract against L. rhamnosus and E. coli, and the zone of inhibition was measured. While both the extracts inhibited the growth of E. coli, it did not show any zone of inhibition against L. rhamnosus. The latter possess surface layer proteins, SlpX and SlpA, which prevented the influx of the phytocompounds of C. bonducella, as demonstrated by molecular docking using Autodock Vina. Docking results showed that the binding of the phytocompounds to the SlpX and SlpA proteins was not in the active pockets. These findings conclude that C. bonducella seed kernel extracts are safe against the gut bacteria L. rhamnosus.
Antibacterial Activity, Molecular Docking, Polycystic Ovary Syndrome, Phytocompounds, Surface Layer Protein
Caesalpinia bonduc (L.) Roxb., a traditional medicinal plant belonging to the family Caesalpiniaceae, is found in Africa and South Asia, particularly in the tropical regions of India. This plant possesses various secondary metabolites like alkaloids, flavonoids, glycosides, saponins, tannins, and triterpenoids. Major phytocompounds found in the seed kernels are bonducellin, citrulline, a-caesalpin, b-caesalpin, g-caesalpin, oleic acid, palmitic acid, stearic acid, aspartic acid, and arginine.1-3 C. bonducella seed kernel (Figure 1) has numerous therapeutic properties, including anti-bacterial, anti-pyretic, anti-diuretic, anthelmintic, antioxidant, analgesic, anti-diarrhoeal, anti-anaphylactic, anti-asthmatic, anti-viral, anti-amoebic, and anti-cancer. It also possesses anti-diabetic, anti-inflammatory, anti-estrogenic, and anti-androgenic properties; hence, it is widely used in the treatment of polycystic ovary syndrome (PCOS) in Indian women.4-9
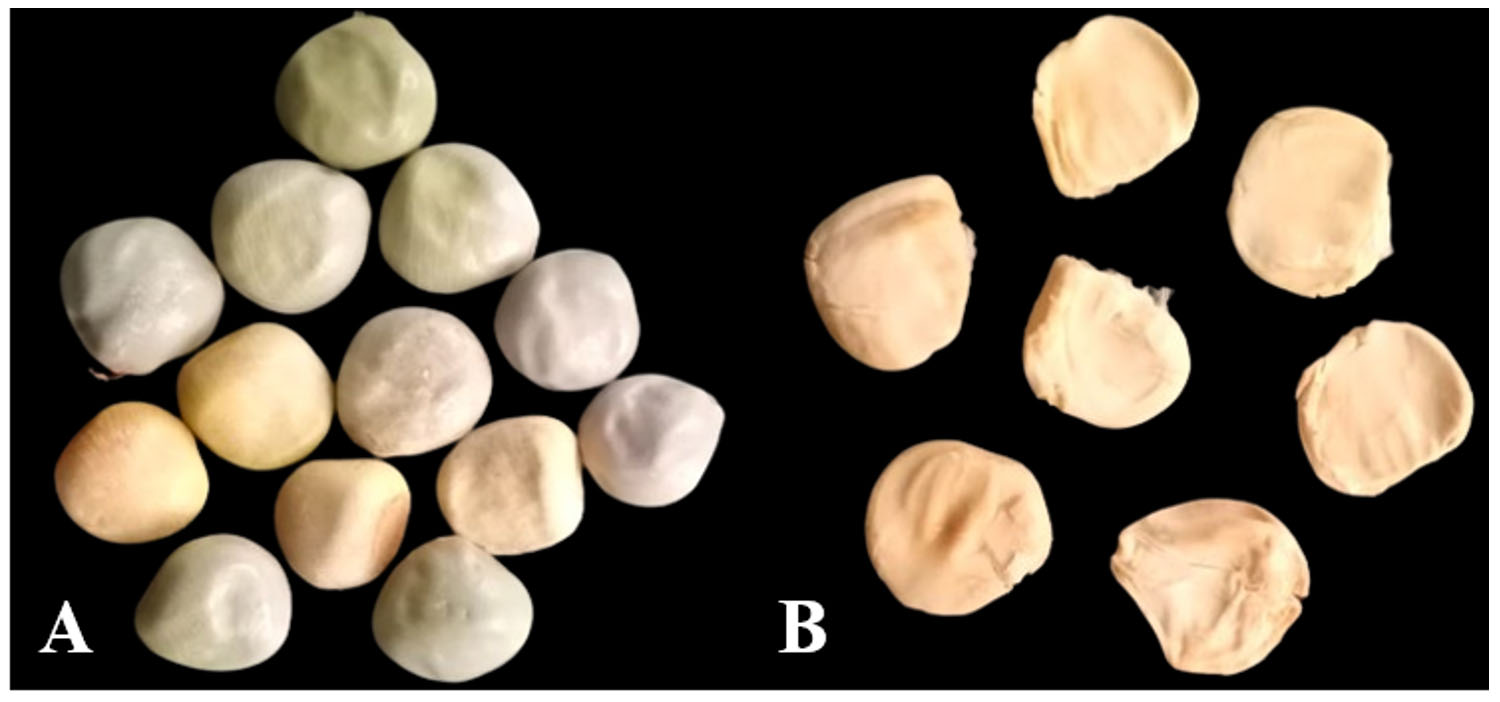
Figure 1. A) Caesalpinia bonducella seed; B) Caesalpinia bonducella seed kernels
PCOS is an endocrine disorder that affects women of childbearing age.10 Research on human trials has revealed that there is a significant change in the taxonomic diversity of gut bacteria in PCOS patients.11 It has been shown that dysbiosis in the gut is directly related to PCOS and obesity.12 Alternative medicine for PCOS is popular in India, and C. bonducella seed kernel with equal parts of pepper mixed with ghee or honey for 48 days is prescribed by Siddha practitioners. Although there is a plethora of evidence that C. bonducella seeds are toxic to human pathogens like Staphylococcus aureus, Pseudomonas aeruginosa, Mycobacterium smegmatis, Salmonella typhi, Escherichia coli, Proteus mirabilis, Klebsiella pneumoniae, and Vibrio cholerae,13-15 its effect on probiotic gut bacteria has not been investigated.
The gut microbiota is a complex biological community, comprising more than thousand microbial species.16 The microbiota in our gut typically sustains our health, which ensures resistance to pathogen colonization, produces short-chain fatty acids (SCFAs) that fuel epithelial cells, regulates gene expression, and produces vitamins and toxins, in addition to regulating cholesterol metabolism and bile deconjugation, all of which are vital functions for human existence.17 The gut microbiota is composed primarily of the phyla Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia. Among these phyla, 90% are composed of Bacteroidetes and Firmicutes. More than two hundred distinct genera comprise the Firmicutes phylum, including Ruminicoccus, Bacillus, Lactobacillus, and Clostridium.18 The most significant probiotic bacteria in the gut microbiome belong to the genus Lactobacillus, which is a member of the phylum Firmicutes. Lactobacillus, a frequently consumed probiotic, makes up 6% of the total bacteria in the human duodenum and 0.3% in the human colon.19 It kills the pathogens by producing lactic acid, acetic acid, propionic acid, bacteriocins, and reactive functional niches in the gut.20
Lactobacillus species also influence the functions and behaviors associated with the central nervous system, where species like Lactobacillus helveticus NS8 reduce depression, anxiety, and cognitive dysfunction and increase serotonin and neuropeptides in the hippocampus.21 Lactobacillus rhamnosus regulates the GABA receptors that reduce anxiety and depression.22 Dysbiosis in the gut microbiota results in the alteration of the beneficial and harmful bacterial ratio and is associated with many diseases and disorders, especially obesity, insulin resistance, and diabetes, that are directly associated with PCOS.23 To balance gut barrier integrity and mucosal barrier defense and improve host immunological responses, these gut-dwelling Lactobacillus species converse not only with one another but also with the gut epithelial lining.24 Hence, it is essential to know if the phytocompounds consumed for treating various disorders alter the gut bacterial community.
In this study, we investigated the effects of methanolic and aqueous extracts of
C. bonducella seed kernels on L. rhamnosus, the most prevalent gut bacteria. Further, we performed molecular docking of the bioactive compounds in the seed kernel against the surface layer proteins of Lactobacillus spp., (SlpX and SlpA), as these proteins play a crucial role in protecting the bacteria from the outer environment.
Bacterial culture
Two strains of bacteria were used in this research. E. coli TOP10 M15 was procured from the Microbial Type Culture Collection (MTCC). L. rhamnosus GG was isolated from kimchi (fortified food) by spread-plating the serially diluted food powder. Both cultures were validated using 16S rDNA sequencing. The forward primer -5’AGGCTACGCTAACCGATGTC 3’ and reverse primer -5’AACGGCATACATTAACGCGC 3’ were used in the 20 µL reaction containing 100 ng of isolated genomic DNA as a template, two primers (10 pmol each) described by Scarpellini et al.25, 0.2 mM of dNTPs, 6.0 units of Taq polymerase, and 10 x buffer with 3 mM MgCl2. The reaction protocol was: 94°C for 5 mins, 30 cycles of 94°C for 1 min, 58°C for 2 mins, 72°C for 2 min, and then 72°C for 10 mins, carried out in a thermal cycler (Aligent Sure cycler 8800). Purified PCR products of about 1500 bp were sequenced, and BLAST was performed to identify the strains. All strains were stored at -20°C in glycerol until use.
Composition of culture medium
E. coli TOP10 M15 was grown in Luria-Bertani broth (LB) agar composed of tryptone (10 g/L), yeast extract (5 g/L), sodium chloride (10 g/L), and agar (15 g/L) with a pH adjusted to 7.5 ± 0.2 and incubated at 37°C for 24 hours. L. rhamnosus GG was grown in De Man-Rogosa-Sharpe (MRS) agar, composed of proteose peptone (10 g/L), HM peptone B (10 g/L), yeast extract (5 g/L), dextrose (glucose) (20 g/L), polysorbate 80 (Tween 80) (1 g/L), ammonium citrate (2 g/L), sodium acetate (5 g/L), magnesium sulfate (0.1 g/L), manganese (II) sulfate (0.05 g/L), di-potassium hydrogen orthophosphate (2 g/L), agar (12 g/L), adjusted to pH 6.5 ± 0.2, and incubated at 37°C for 48 hours.
Seed extraction
Fresh seeds of C. bonducella Roxb. were collected from the National Institute of Siddha, Chennai, India (11° 15′ 42.9984 N; 78° 23′ 8.5668 E) and stored at 4°C. Prof. P. Jayaraman, a taxonomist from Presidency College in Chennai, India, validated the seeds. The collected seeds were shade-dried for 2 weeks, and the seed coats were broken to collect the seed kernels, which were shade-dried for 10 more days and pulverized into a fine powder. The powder was then air-dried for 30 minutes. About 5 g of the seed powder was measured and dissolved in 50 ml of methanol and water, sealed, and incubated for 12 hours at 28 ± 2°C in a shaking incubator at 125 rpm. After 12 hours, the entire mixture was filtered using Whatman filter paper grade 1 (90 mm), and the same process was repeated for 3 subsequent days to extract most of the active ingredients. The methanol and aqueous extracts were dried and stored at 4°C until further analysis.
Disc diffusion assay
The effect of methanol and aqueous crude extracts of the seed kernel (100, 500, and 1000 µg/ml) were screened against L. rhamnosus GG and E. coli using the disc diffusion method. Sterile discs (6 mm diameter, Himedia) were saturated with the above-mentioned concentrations of the extracts and placed on the MRS agar and LB agar medium, uniformly spread with L. rhamnosus GG, and E. coli, respectively. The inoculum was approximately 106 cfu/ml. A standard antibiotic (Ampicillin, 30 µg/disc) was imbued in the sterile disc and was used as a positive control. The plates were then incubated at 4°C for one hour, which is efficient for the diffusion of the test material. Then the plates were incubated at 37°C and checked for zones of inhibition after 24 and 48 hours.
Selection and preparation of target proteins
Surface layer proteins play a crucial role in the communication between the cell and the environment. Surface layer proteins SlpX and SlpA, which are essential for protecting Lactobacillus spp. against the external environment, were selected as target proteins for docking against the phytocompounds of C. bonducella seeds.
The three-dimensional structures of the surface layer proteins, SlpX (PDB ID: 7QFJ) with a resolution of 2.50Å determined with X-ray diffraction and SlpA (PDB ID: 7QFG) with a resolution of 1.65Å determined with X-ray diffraction were obtained from the protein data bank (PDB) (https://www.rcsb.org/). Any associated groups, including PO42- and water molecules, were eliminated from the proteins in PyMol software, and the protein structures were saved in pdbqt format for docking.26 The binding pockets of the proteins were predicted by an online tool CASTp (http://sts.bioe.uic.edu/castp/) by uploading the protein pdb files and setting the radius to 1.4Åֵ.27
Selection and preparation of ligands
A comprehensive literature search was conducted to gain insights into the bioactive compounds that are found in C. bonducella seed kernels. All relevant information was gathered from well-known databases, such as PubMed, Scopus, Google Scholar, and EMBASE; associated articles were located by looking up their references. Bioactive compounds of C. bonducella, secondary metabolites, bonduc nut, LC-MS, GC-MS, and additional pertinent terms were the keywords used in the literature search. The search did not exclude any specific region, period, or language. The structures of the compounds retrieved from the literature were obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov). The structures were uploaded to PyRx and converted from the spatial data file (SDF) to PDB format using Open Babel (https://www.cheminfo.org/), an open-source software. The 2D structure of the derived bioactive compounds was drawn using ChemSketch version 2022.1.2 (https://www.acdlabs.com/). The Universal Force Field was applied to all the selected ligands before the molecular docking procedure.
Molecular docking
For molecular docking, the National Biomedical Computation Resources PyRx open-source software for virtual screening, which incorporates Autodock Vina, was utilized. The Vina Wizard software was utilized to bind two different surface layer proteins (SlpA and SlpX) against 54 compounds from the seeds. The docking site of SlpA was defined at the active site with a grid box size of 80x80x80, a grid center of -26.055Å, -6.121Å, 33.527Å, grid space of 0.375Å. Similarly, for SlpX, a grid box size of 80x80x80, a grid center of 10.524Å, 0.441Å, 7.755Å, grid space of 0.375Å. The binding energy scores were used to predict the efficiency with which the compounds would interact with the target proteins.26 Visualization of the docked protein-ligand complex was processed by PyMOL 2.3 software.
C. bonducella has been widely used to manage disorders like PCOS, Diabetes mellitus, insulin resistance, bacterial infections, diarrhea, and urinary tract infections. Prior research has examined the seed kernel extract’s ability to combat pathogenic bacteria, including gram-positive bacteria like Bacillus subtilis, as well as gram-negative bacteria such as P. aeruginosa, E. coli, and P. mirabilis.28,29 However, the impact of the seed extract on the human gut bacteria has not been studied yet. Our study focused on the effect of C. bonducella seed against L. rhamnosus GG, the most prominent bacterium in the human gut.
The antibacterial assay was performed for L. rhamnosus GG and E. coli with 3 different concentrations of the methanolic and aqueous extracts of C. bonducella seed kernel (100, 500, and 1000 µg), with ampicillin (30 µg) as the positive control (Table 1). No zone of inhibition was observed for L. rhamnosus GG in both methanolic and aqueous extracts, whereas E. coli showed a distinct zone of inhibition for 1000 µg of methanolic extract and all 3 concentrations of aqueous extract, with an increase in the zone size in proportion to the concentration of the extracts (Figure 2). This states that growth of L. rhamnosus GG is not affected by both the seed extracts, whereas that of E. coli is affected.
Table (1):
Effect of the seed extracts against E. coli and L. rhamnosus
| Sample | Concentration (µg/disc) | E. coli (mm) | L. rhamnosus (mm) |
|---|---|---|---|
| Ampicillin | 30 | 16 ± 1 | 25 ± 2 |
| Methanolic extract | 100 | – | – |
| 500 | – | – | |
| 1000 | 24 ± 1 | – | |
| Aqueous extract | 100 | 10 ± 1 | – |
| 500 | 22 ± 1 | – | |
| 1000 | 23 ± 2 | – |
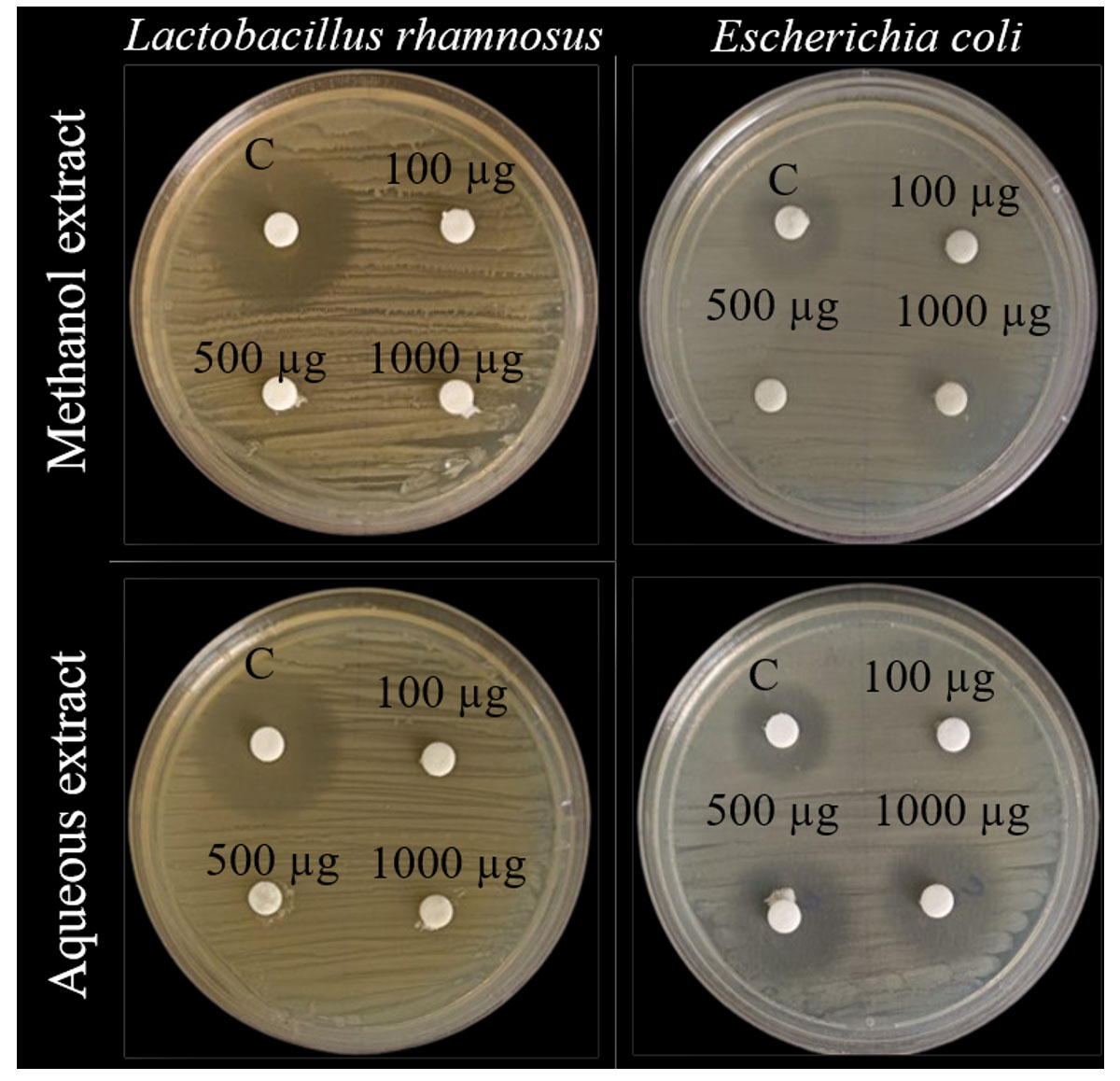
Figure 2. In-vitro disc diffusion assay for aqueous and methanol extracts of Caesalpinia bonducella seed kernel. The zones of inhibition are observed on the cultured plates
The result shows that the seed kernel does not affect the growth of L. rhamnosus while inhibiting that of E. coli. We speculated that the exclusive surface layer proteins present in Lactobacillus species may be responsible for L. rhamnosus GG’s protection against the phytocompounds. These surface layer proteins have been found in many Lactobacillus spp., including L. brevis, L. buchneri, L. helveticus, L. hilgardii, L. acidophilus, L. amylovorus, L. crispatus, L. gallinarum, and L. rhamnosus.30,31 Numerous strains of Lactobacillus are capable of inhibiting pathogen adhesion to the intestinal epithelial cells and mucus with the help of their surface layer proteins, SlpX and SlpA, with molecular weights ranging from 30-130 kDa.30,32 These proteins also assist these gut microbes to survive in a hostile gut environment that is rich in digestive enzymes and bile salts.33
In silico analyses were performed to investigate if the phytocompounds bind to the active binding pockets of the Slps. For this, the 3D structure of the two Slps were derived from the Protein Data Bank and was used to predict the binding pocket with the CASTp tool. The tool predicted 10 amino acid sites for SlpA protein and 14 amino acid sites for SlpX as binding pockets (Table 2). These two proteins were docked with the 54 phytocompounds in the seed kernel of C. bonducella that were retrieved from the previously reported literature and their binding affinity is listed in Table 3.
Table (2):
The active pockets of the proteins SlpA and SlpX predicted with CASTp tool
Protein |
Area (Å2) |
Volume (Å3) |
Binding pockets |
|---|---|---|---|
SlpA (PDB:7QFG) |
64.462 |
50.394 |
His323, Asn324, Ala325, Tyr326, Tyr328, Asn375, Ala377, Asn378, Gly415, Ala416 |
SlpX (PDB:7QFJ) |
134.916 |
69.305 |
Gly294, Thr295, Leu296, Tyr297, Gly298, Asn299, Ile304, Val329, Ser342, Val344, Val359, Lys360, Thy361, Ser362 |
Table (3):
Binding affinity of 54 phytocompounds of Caesalpinia bonducella seeds against SlpX and SlpA
| No. | Compound Name | Structure | Binding affinity | |
|---|---|---|---|---|
| SlpX | SlpA | |||
| 1. | 1-undecanol (8184) | -4.1 | -4.3 | |
| 2. | 1-Phenanthrenecarboxylic acid tetradecahydro 7-(2-methoxy-2-oxoethylidene)-1,4a,8-trimethyl-9-oxo-methyl ester (5379250) |  |
-6.5 | -4.5 |
| 3. | 15,17,19,21-Hexatriacontatetrayne (537054) | -3.3 | -4.1 | |
| 4. | 16-dehydroprogesterone (101964) |  |
-6.3 | -7.1 |
| 5. | 2,4 bis (1 methyl 1 phenyl ethyl phenol (102877) | 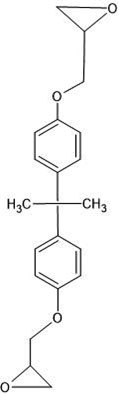 |
-6.4 | -6.9 |
| 6. | 2-Hydroxy-4-methoxy-7-methyl-7,8,9,10,11,12,13,14-octahydro-6-oxabenzocyclododecen-5-one (602765) | 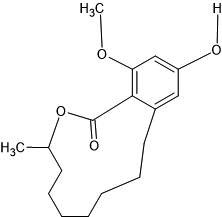 |
-6.7 | -6.2 |
| 7. | 4-o-methylmannose (345716) | 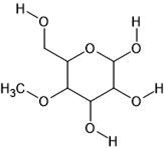 |
-4.9 | -4.9 |
| 8. | 5-hydroxymethyl-furfural (237332) | 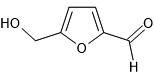 |
-4.3 | -4.5 |
| 9. | 6b-Hydroxymethandienone (13241205) | 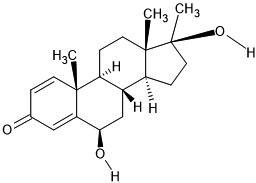 |
-6.6 | -7 |
| 10. | 7,11-Dihydroxyprogesterone (539326) | 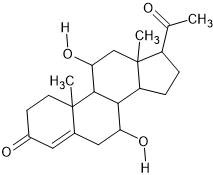 |
-6.9 | -6.6 |
| 11. | 9,12-octadecadien-1-ol (5462912) | 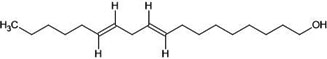 |
-3.9 | -5 |
| 12. | 9,12-Octadecadienoic acid (Z, Z) 2-hydroxy-1-(hydroxymethyl) ethyl ester (5365676) | 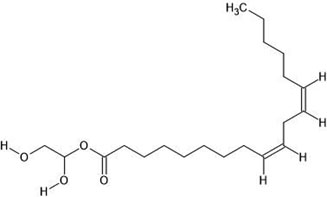 |
-4.1 | -4.4 |
| 13. | 9,12-Octadecadienoic acid methyl ester (8203) | -4.5 | -5.2 | |
| 14. | Androst-4-en-11-ol-3,17-dione-9-thiocyanato- (91694000) | 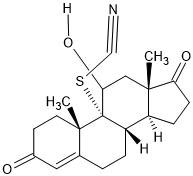 |
-6.7 | -4.6 |
| 15. | Alpha-tocopherol (57393415) | 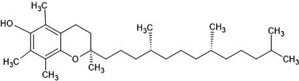 |
-5 | -6.4 |
| 16. | Alpha-caesalpin (21679154) | 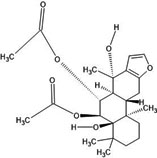 |
-6.4 | -6.2 |
| 17. | Anthraegostatrine (313024) | 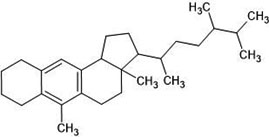 |
-6.2 | -7.3 |
| 18. | Benzimidazole 2-[1-(4-isopropylbenzyl) ethyl] (589515) |  |
-6.6 | -7.1 |
| 19. | Beta-sitosterol (222284) |  |
-6.2 | -6.2 |
| 20. | Beta-caesalpin (71440416) |  |
-6.6 | -6.5 |
| 21. | Bonducellin (14079439) |  |
-6.1 | -6.5 |
| 22. | Caesalpin F (101937720) |  |
-5.7 | -6.4 |
| 23. | Caesalpinin (15329770) | 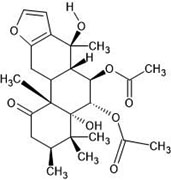 |
-6.3 | -6.5 |
| 24. | Campesterol (173183) | 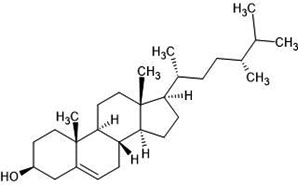 |
-7 | -6.7 |
| 25. | Carpesterol (21155918) | 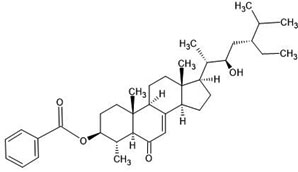 |
-6.7 | -8.1 |
| 26. | Cassane furano-diterpenes (102286656) |  |
-7.3 | -7.2 |
| 27. | Cholesterol (5997) | 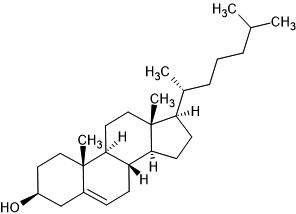 |
-6 | -6.6 |
| 28. | Corticosterone 21 acetate (255846) | 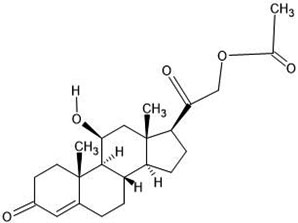 |
-6.4 | -6.3 |
| 29. | D5-avenasterol (91753899) | 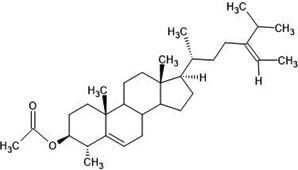 |
-6.5 | -6.5 |
| 30. | Eicosyl acetate (110347) | -3.5 | -3.7 | |
| 31. | Epsilon-caesalpin (21679153) | 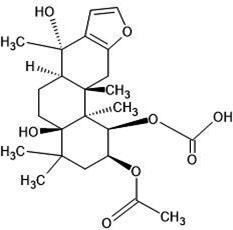 |
-6.2 | -7.2 |
| 32. | Ergosterol (6436903) | 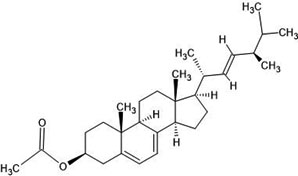 |
-6.9 | -7 |
| 33. | Ethyl oleate (5363269) |  |
-3.8 | -4 |
| 34. | Fucosterol (5281328) |  |
-6.7 | -6.6 |
| 35. | Hexadecanoic_acid_2-hydroxy-1-(hydroxymethyl) _ethyl_ester (129853056) | -4.5 | -4.1 | |
| 36. | Linolei acid (5280450) |  |
-4.7 | -5.3 |
| 37. | Lycopene (446925) | -7.2 | -6.7 | |
| 38. | N-nonadecanol-1 (890281) |  |
-4.4 | -3.6 |
| 39. | Nandrolone (9904) |  |
-6.5 | -6.8 |
| 40. | Octacosane (12408) | -3.8 | -3.3 | |
| 41. | Oleic acid (445639) |  |
-4.2 | -4.2 |
| 42. | Palmitic acid (985) | -4.2 | -4.5 | |
| 43. | Pentadecanoic acid (13849) | -3.9 | -3.8 | |
| 44. | Phenol,2_(1_phenyl_ethyl) (95322) |  |
-6.8 | -7 |
| 45. | Phorbol 12,13,20 triacetate (499954) |  |
-6 | -6.4 |
| 46. | Pregnenolone (8955) | 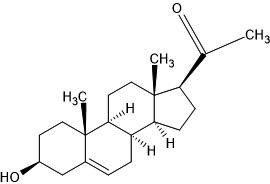 |
-6.2 | -6.8 |
| 47. | Retinol (445354) | 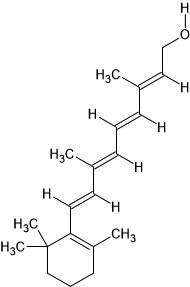 |
-6.2 | -6.4 |
| 48. | Retinyl acetate (638034) | 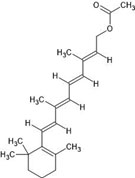 |
-6.3 | -6.1 |
| 49. | Retrofractamide A (11012859) | 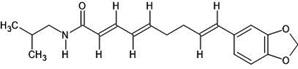 |
-5.7 | -6 |
| 50. | Rhodopin (5365880) | -5.1 | -4.9 | |
| 51. | Simvastatin (54454) | 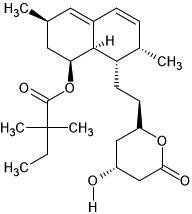 |
-6.6 | -7.3 |
| 52. | Stearic acid (5281) | -4.1 | -4.9 | |
| 53. | Stigmasterol (5280794) |  |
-6.4 | -6.7 |
| 54. | Tanshinone IIA (164676) | 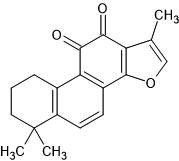 |
-6.7 | -7.8 |
SlpA showed a higher binding affinity with compounds, including Carpesterol, tanshinone IIA, and anthraegostatrine, with a binding score of -8.1, -7.8, and -7.3 kcal/mol, respectively. The hydrogen bond interaction between the complex of both Carpesterol and tanshinone IIA with SlpA is at Lys358 with a length of 2.2Å, whereas anthraegostatrine did not show any hydrogen bond interaction with the protein (Figure 3). SlpX showed a higher binding affinity with compounds cassane furano-diterpenes, lycopene, and campesterol, with binding scores of -7.3, -7.2, and -7 kcal/mol, respectively. The hydrogen bond interaction between the complexes of cassane furano-diterpenes is with Thr189 and Thr280, with a length of 3.2Å and 2.7Å, respectively. Campesterol showed two hydrogen bond interactions at Thr303 with a length of 2.4Å and Ser305 with a length of 3.0Å, whereas lycopene had no hydrogen bond interaction with SlpX (Figure 4).
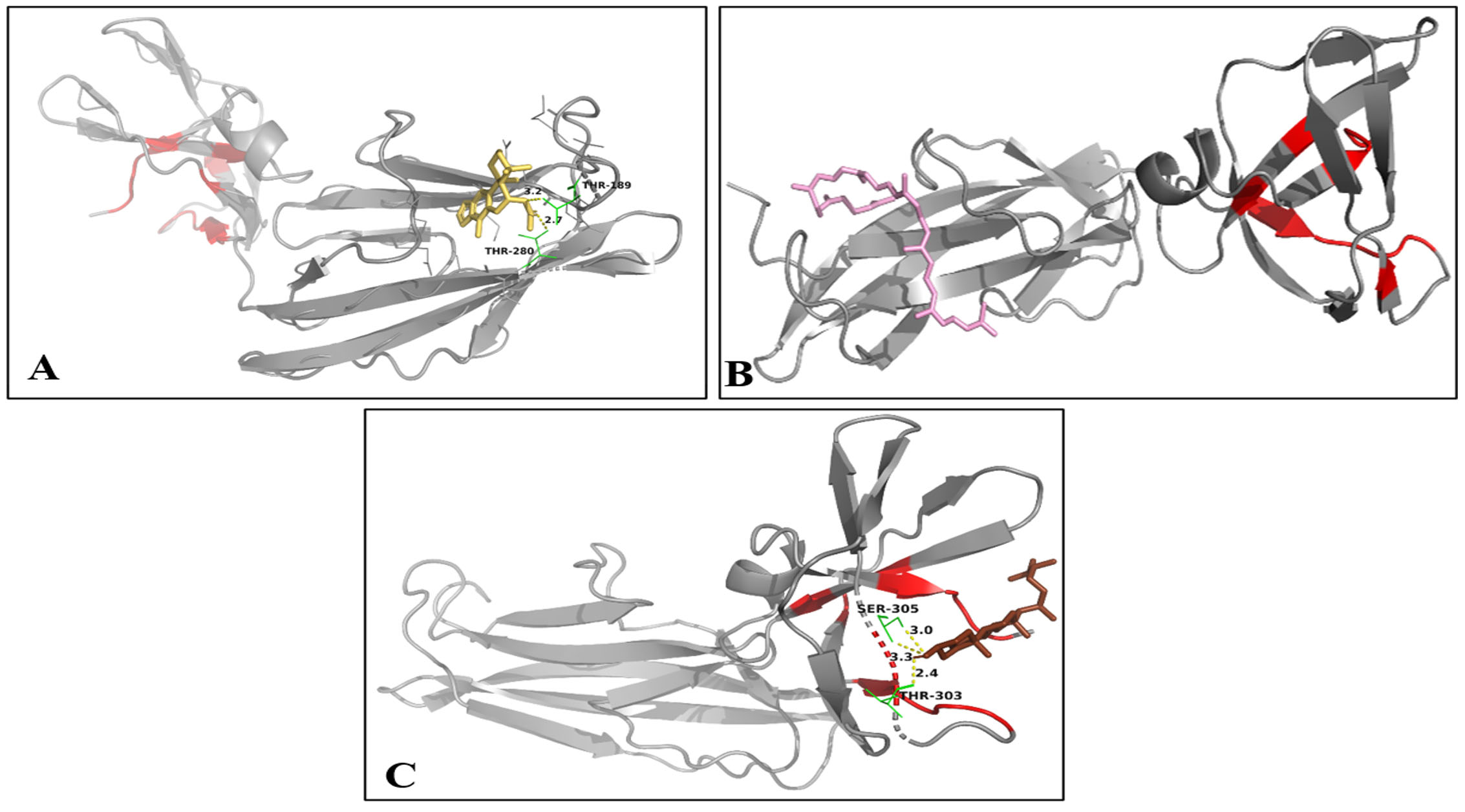
Figure 3. Binding of protein SlpX with compound A) Cassane furano-diterpene B) Lycopene C) Campesterol. (Red indicates active sites of protein; yellow indicates hydrogen bond; grey indicates the protein; yellow indicates Cassane furano-diterpene; pink indicates Lycopene; brown indicates Campesterol)
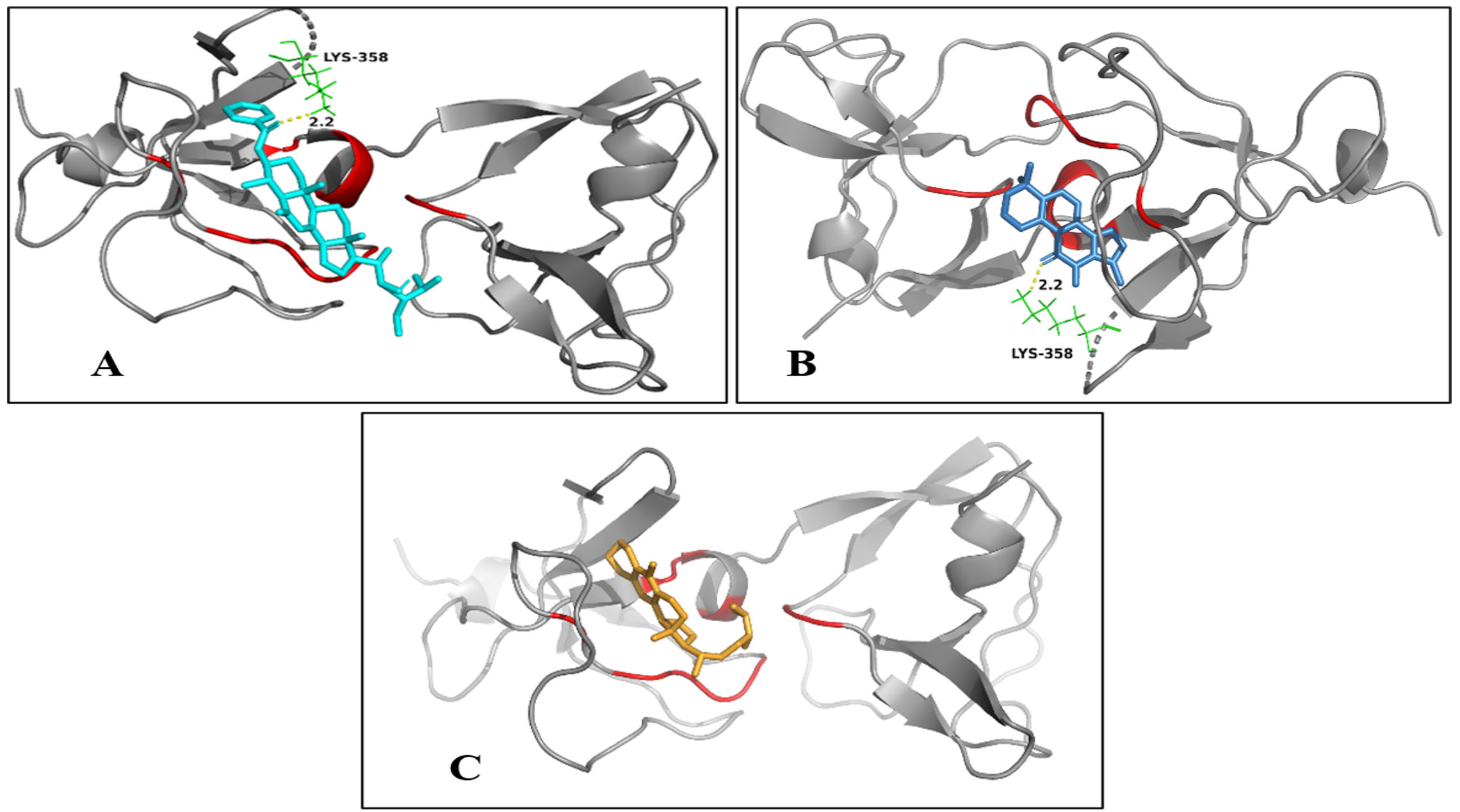
Figure 4. Binding of Protein SlpA with Compounds A) Carpesterol, B) Tanshinone IIA, and C) Anthraegostatrine. (Red indicates active sites of protein; yellow indicates hydrogen bond; gray indicates the protein; turquoise indicates carpesterol; blue indicates tanshinone IIA; orange indicates anthraegostatrine)
Although these compounds showed higher binding scores with the Slp proteins, their interaction with the Slp amino acid residues did not match the amino acid residues at the active binding pocket (Table 2). Thus, it shows that the phytocompounds do not bind tightly to the Slps and are hence prevented from entering the bacterial cell.
E. coli lacks these proteins and may be susceptible to the phytocompounds, thereby inhibiting its growth. Likewise, Clostridium spp., having huge potential as a probiotic by producing metabolites including secondary bile acids, butyrate, acetic acid, and propionic acid, helps in strengthening and energizing intestinal epithelial cells and intestinal barrier,34 and Enterococcus spp., produces small peptides that are bacteriocin, having antimicrobial properties inhibiting the growth of E. coli, P. aeruginosa, L. monocytogenes, and V. cholerae in the human gut.35 These Clostridium spp. and Enterococcus spp. families in the gut also possess Slp35,36 that the seed’s phytocompounds may not kill, and thus the gut microbiota will not be disturbed by consuming the seed kernel as a medicine. However, pathogenic bacteria like S. aureus, P. aeruginosa, M. smegmatis, S. typhi, P. mirabilis, K. pneumoniae, and V. cholerae lack Slps, making them vulnerable to and inhibited by the phytocompounds. Our finding put forth that C. bonducella seed extract affects those bacteria without Slp proteins and is safe against probiotic gut bacteria.
Limitations of this study
The study’s scope is confined to only L. rhamnosus, hence, requires further validation with other gut bacteria to ascertain the safe use of C. bonducella seed for various disorders.
Caesalpinia bonducella seeds are used to treat various disorders, notably fever, edema, malaria, diabetes, and PCOS, and are shown to possess antibacterial properties against several pathogens. Lactobacillus is a prominent probiotic bacterium colonizing the human intestinal tract and plays a crucial role in preventing intestinal damage caused by certain bacterial infections. In our research, the impact of the seed extract of C. bonducella against Lactobacillus rhamnosus was tested where seed extract did not hinder the growth of the bacteria. Molecular docking of the surface layer proteins, SlpX and SlpA, to the phytocompounds of C. bonducella seed kernel, revealed no binding to active binding pockets. This suggests that C. bonducella seed extracts are not toxic to Lactobacillus spp., which is prevalent in the gut microbiome.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
BU contributed to the study’s conception and design. AP and MK contributed equally to material preparation, data collection, and analysis. SMH performed molecular docking studies. AP and MK wrote the manuscript. BU edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This research work was funded by the SRM Institute of Science and Technology, Kattankulathur, Chennai, India.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Gupta M, Mazumder UK, Kumar RS, Sivakumar T, Gomathi P, Rajeshwar Y. Antioxidant defense system induced by a methanol extract of Caesalpinia bonducella in rat liver. J Pharmacol Sci. 2005;43(5):411-419.
Crossref - Gaur RL, Sahoo MK, Dixit S, et al. Antifilarial activity of Caesalpinia bonducella against experimental filarial infections. Ind J Med Res. 2008;128:65-70
- Lilaram, Ahmed RN. Effect of ethanolic seed extract of Caesalpinia bonducella on female reproductive system of albino rat: a focus on antifertility efficacy. Asian Pacific J Trop Dis. 2012;2(2):S957-S962.
Crossref - Neogi NC, Nayak KP. Biological investigation of Caesalpinia bonducella Flem. Indian J Pharmacol. 1958;20:95-100. Accessed April 10, 2024.
- Gayaraja S, Shinde S, Agarwal SL. Antiasthmatic properties of Caesalpinia bonduc leaves. Indian J Pharmacol. 1978;10:86-89. Accessed April 10, 2024.
- Dhar ML, Dhar MM, Dhawan BN, Mehrotra BN, Ray C. Screening of Indian plants for biological activity: I. Indian J. Exp. Biol. 1968;6(4):232-247. https://pubmed.ncbi.nlm.nih.gov/5720682/. Accessed April 14, 2024.
- Shukla S, Mehta A, Mehta P, Vyas SP, Shukla S, Bajpai VK. Studies on anti-inflammatory, antipyretic and analgesic properties of Caesalpinia bonducella F. seed oil in experimental animal models. Food Chem Toxicol. 2010;48(1):61-64.
Crossref - Salunke KR, Ahmed RN, Marigoudar SR, Lilaram. Effect of graded doses of Caesalpinia bonducella seed extract on ovary and uterus in albino rats. J Basic Clin Physiol Pharmacol. 2011;22(1-2):49-53
Crossref - Murugesan BM, Muralidharan P, Hari R. Effect of ethanolic seed extract of Caesalpinia bonducella on hormones in mifepristone induced PCOS rats. J Appl Pharm Sci. 2020;10(2),072-076.
Crossref - Sivasankari R, Usha B. Reshaping the gut microbiota through lifestyle interventions in women with PCOS: a review. Indian J. Microbiol. 2022;62(3),351-363.
Crossref - Rizk MG, Thackray VG. Intersection of polycystic ovary syndrome and the gut microbiome. J Endocr Soc. 2021;5(2):bvaa177.
Crossref - Liu R, Zhang C, Shi Y, et al. Dysbiosis of gut microbiota associated with clinical parameters in polycystic ovary syndrome. Front Microbiol. 2017;8:324.
Crossref - Billah MM, Islam R, Khatun H, et al. Antibacterial, antidiarrhoeal, and cytotoxic activities of methanol extract and its fractions of Caesalpinia bonducella (L.) Roxb leaves. BMC Complement Altern Med. 2013;13:1-7.
Crossref - Kumar SRS, Shastri SL, Venkatesh R, Pradeepa K, Krishna V. Antibacterial potential of Caesalpinia bonducella extracts and their isolated phytoconstituents: in vitro and in silico analysis. J Appl Biol Biotechnol. 2019;7(3):41-46.
Crossref - Arif T, Mandal TK, Kumar N, et al. In vitro and in vivo antimicrobial activities of seeds of Caesalpinia bonduc (Lin.) Roxb. J Ethnopharmacol. 2009;123(1):177-180.
Crossref - Lagier JC, Khelaifia S, Alou MT, et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat Microbiol. 20161(12):16203.
Crossref - Harmsen HJ, de Goffau MC. The human gut microbiota. Microbiota of the human body: Implications in health and disease. 2016;902:95-108.
Crossref - Rinninella E, Raoul P, Cintoni M, et al. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms. 2019;7(1),14.
Crossref - Huang R, Wu F, Zhou Q, et al. Lactobacillus and intestinal diseases: Mechanisms of action and clinical applications. Microbiol Res. 2022;260:127019.
Crossref - Dempsey E, Corr SC. Lactobacillus spp. for gastrointestinal health: current and future perspectives. Front Immunol. 2022;13:840245.
Crossref - Desbonnet L, Clarke G, Traplin A, et al. Gut microbiota depletion from early adolescence in mice: Implications for brain and behavior. Brain Behav Immun. 2015;48:165-173.
Crossref - Liang S, Wang T, Hu X, et al. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neurosci J. 2015;310:561-577.
Crossref - Yurtda G, Akdevelioglu Y. A new approach to polycystic ovary syndrome: the gut microbiota. J Am Coll Nutr. 2020;39(4):371-382.
Crossref - Martin R, Chamignon C, Mhedbi-Hajri N, et al. The potential probiotic Lactobacillus rhamnosus CNCM I-3690 strain protects the intestinal barrier by stimulating both mucus production and cytoprotective response. Sci Rep. 2019;9(1):5398.
Crossref - Liu W, Bao Q, Jirimupu, et al. Isolation and identification of lactic acid bacteria from Tarag in Eastern Inner Mongolia of China by 16S rRNA sequences and DGGE analysis. Microbiol Res. 2012 Jan 20;167(2):110-115.
Crossref - Kandasamy V, Sathish S, Madhavan T, Balasundaram U. In-Silico Screening of Phytochemical Compounds in Caesalpinia Bonducella L. Seeds Against the Gene Targets of Ovarian Steroidogenesis Pathway. J Microbiol Biotechnol Food Sci. 2023;13(2):e6124-e6124.
Crossref - Tian W, Chen C, Lei X, Zhao J, Liang J. CASTp 3.0: computed atlas of surface topography of proteins. Nucleic Acids Res. 2018;46(W1),W363-W367.
Crossref - Tambekar DH, Khante BS. Antibacterial properties of traditionally used medicinal plants for enteric infections by adivasi’s (Bhumka) in melghat forest (Amravati District). Int J Pharm Sci Res. 2011;1(9):120-128.
Crossref - Savaji P, Navanit S, Nayeem A. Phytochemical extraction and antibacterial studies of Caesalpinia bonducella seed extracts. Mapana Journal of Sciences. 2014;13(4):47-54.
Crossref - Zhang W, Wang H, Liu J, Zhao Y, Gao K, Zhang J. Adhesive ability means inhibition activities for Lactobacillus against pathogens and S-layer protein plays an important role in adhesion. Anaerobe. 2013;22:97-103.
Crossref - Hynonen U, Palva A. Lactobacillus surface layer proteins: structure, function and applications. Appl Microbiol Biotechnol. 2013;97(12):5225-5243.
Crossref - Wakai T, Kano C, Karsens H, Kok J, Yamamoto N. Functional role of surface layer proteins of Lactobacillus acidophilus L-92 in stress tolerance and binding to host cell proteins. Biosci Microbiota Food Health. 2021;40(1),33-42.
Crossref - Shi Z, Li X, Fan X, et al. The SlpX protein plays a crucial role in the intestinal juice tolerance of Lactobacillus acidophilus CICC6074. Food Biosci. 2024;59(4):103865.
Crossref - Guo P, Zhang K, Ma X, He P. Clostridium species as probiotics: potentials and challenges. J Animal Sci Biotechnol. 2020;11:24.
Crossref - He Y, Xu X, Zhang F, et al. Anti-adhesion of probiotic Enterococcus faecium WEFA23 against five pathogens and the beneficial effect of its S-layer proteins against Listeria monocytogenes. Can J Microbiol. 2019;65(3):175-184.
Crossref - Fagan RP, Fairweather NF. Biogenesis and functions of bacterial S-layers. Nat Rev Microbiol. 2014;12(3):211-222.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.