


ISSN: 0973-7510
E-ISSN: 2581-690X
The lipid-producing gram-positive bacteria were isolated from soil and it was identified as Streptomyces fradiae JJ1 MK733985.1. Bacterial lipid synthesis was explored using rice washed water through the fermentation research. The exponential growth phase was reached on the fifth day of fermentation, according to the results. The ability of Streptomyces fradiae to produce microbial oil was compared to that of International Streptomyces Project-2 Medium (ISP2) medium. The results showed that RWW culture medium yielded 85.4% fatty acids and 60.3 g/L biomass, while ISP2 medium yielded 83.2% wt fatty acids and 59.5 g/L biomass. The microbial oil was transesterified into Fatty acid methyl esters (FAME), and its characteristics were studied using American Society for Testing Materials (ASTM) standards. The specific gravity was 0.864, the density was 0.863, the gross heating value was 42498 kj/kg, the cetane number was 37, and the acid value was 0.46. Streptomyces fradiae grew with an ISP2 density of 0.835, specific gravity of 0.845, gross heating value of 42326 kj/kg, and acid value of 0.43. According to the findings, Streptomyces fradiae isolated from soil can grow in RWW medium for the first time and provide the same yield as commercial media while being significantly less expensive.
Biodiesel, Streptomyces fradiae, Rice Washed Water, Fermentation, Soil Bacteria
Biodiesel, which is made from biomass, is being considered as a viable alternative to currently used fossil fuels.1 These biofuels include ethanol, butanol, and other alcohol compounds. The biodiesel was made from either vegetable oil or animal fat. Three methods were widely used: pyrolysis, micro emulsification, and transesterification.2 Among these, transesterification with vegetable oil is a popular method. Along with methanol or ethanol, sodium hydroxide (NaOH) or potassium hydroxide (KOH) is used as a hetero genus catalyst in the transesterification process. After reacting with alcohol to form alkoxy groups, NaOH or KOH reacts with triglyceride to form methyl- or ethyl-esters and glycerol. Finally, the product goes through several downstream processes, including repeated washing, to produce high-quality biodiesel.3 This method has some drawbacks, such as the difficulty of removing the catalyst, excess methanol and the formation of soap. Bio diesel production using bio catalyst avoids all of these difficulties, but it is a very expensive process that also requires highly purified enzymes.4 As a result, microorganism-based biodiesel produced through fermentation is the most cost-effective way to produce high-quality biodiesels.5 Previous research has shown that lipid-accumulating microorganisms are ideal for biodiesel production because they accumulate lipids in the form of triacylglycerol, which is transesterified into fatty acid methyl esters (FAME) to produce biodiesel.6 Microorganisms use lignocellulosic waste to produce biofuel that is comparable to existing fuels and suitable for diesel engines.7 Fatty acid ethyl esters (FAEE) were used to create recombinant E. coli-based microbial biodiesel in early 2006. Later, researchers reported that E. coli was used to produce FAEE from Xylan.8 Under favorable environmental conditions, the researchers discovered that some bacteria can store more than 20% of their lipids in internal organelles.9 Currently, wastewaters are employed as a source for microbial lipid production, which not only lowers the cost of biodiesel production but also minimizes pollution produced by wastewater. For example, wastewater effluent from butanol fermentation was utilized to cultivate the oleaginous yeast Trichosporon coremiiforme, which resulted in a 19% lipid content and a 68% chemical oxygen demand (COD) reduction.10 Previously, oleaginous yeast was grown in corn starch water as a substrate, which increased lipid productivity,11 similarly, the filamentous fungi Aspergillus oryzae were grown in potato processing wastewater as a substrate, which increased lipid production capacity.12 In this study, we attempt to investigate rice washed wastewater as a substrate for producing biodiesel. The medium’s key advantage is that it is three times cheaper than any other fermentation media. It reduces manufacturing costs and improves waste management. As a result, novel approaches to the exploitation of waste RWW are urgently needed. We isolated Streptomyces fradiae, a potential biodiesel feedstock generator, from coastal soil sediment samples. Streptomyces fradiae isolated from harsh conditions have extremophilic characteristics and can accumulate substantial amounts of lipids. As a result, the Streptomyces fradiae were used as a catalyst to produce biodiesel from RWW for the first time. The key advantage is that it does not require any complicated separation process to isolate biomass, and it forms natural flocculation with RWW and the biomass is simply separated, and the microbiological did not require any further purifications and contains rich concentration of free fatty acid compared to the usual commercial media, ISP2 and it grow well at normal room temperature, simpler than cultivating other microbes.
Streptomyces fradiae isolation
Fifty-three Actinobacterial isolates were isolated from diverse soil samples obtained from the seashores of Indian east coast Mamallapuram beach, Tamil Nadu, India, and were promptly processed utilizing serial dilution technique for Actinobacteria isolation. Each dilution of soil sample was inoculated using the spread plate method in Actinobacteria isolation medium, which was made up of 20 g Starch, 1 g Potassium nitrate (KNO3), 0.5 g Sodium chloride (NaCl), 0.5 g Dipotassium phosphate (K2HPO4), 0.5 g Magnesium sulphate (MgSO4), 20 µL Ferrous sulfate (FeSO4), agar 18 g, and 1 liter of distilled water. Finally, 15 g of Nalidixic acid was added to prevent gram-negative bacteria growth. Infected samples were cultured on separate plates in the dark for 7 to 14 days at room temperature (30°C). The quadrant streak technique was used to isolate pure cultures. All stock cultures were kept on slants of starch casein agar (SCA) and stored at 4°C for subsequent research based on morphological identification. On new slants, monthly subcultures were created.
Screening of oleaginous Actinobacteria
The potential of pure cultures to produce oil was determined through qualitative examination of the Sudan Black B staining technique. Fixed smears of strains were rinsed in running tap water after being soaked with 0.3% Sudan black for 15 minutes. The slides were then soaked in xylene for 10 minutes before being washed with tap water. Finally, the slides were cleaned and allowed to dry after being saturated in methylene blue (counterstain) for 30 seconds. The light microscope was used to examine the slides.
Biomass cultivation
Streptomyces fradiae, an oleaginous strain, was chosen for bench-scale microbial oil production employing the shake flask fermentation method. Initially, five separate tubes containing 50 mL of Streptomyces fradiae culture were inoculated into five separates 2-liter Erlenmeyer flasks, and each flask was filled with 950 mL of sterilized (autoclaved at 121°C) RWW substrate dilutions of 0%, 25%, 50%, and 75%. To determine the optimal dilution factor, all cultures were incubated at the same time. This experiment was carried out in triplicate to determine the best dilution factor for producing higher biomass with the highest oil content in order to optimize the media. Each dilution received nutrients such as KNO3 1 g, NaCl 0.5 g, K2HPO4 0.5 g, and MgSO4 0.5 g, and the original pH was adjusted to 7.0. The remaining 50 mL inoculum was transferred to the final 2-liter Erlenmeyer flask containing 950 mL of ISP2 medium and incubated for 5 days in a rotary shaker at 180 rpm at 30°Celsius. Every day, 1 mL of culture sample was obtained from each flask to measure the growth pattern using a spectrophotometer at O.D. 660 nm.
Microbial oil extraction
After reaching the exponential growth phase on the fifth day of fermentation, the bacterial biomass was harvested by centrifuging at 6000rpm for 15 minutes. The Bligh and Dyer method13 was used to recover microbial oil from biomass pellets. The cells were homogenized in a 2:1 methanol/chloroform ratio to a final volume 20 times the sample volume. In a separator funnel, the agitated sample was separated into two separate layers, the upper aqueous phase was discarded, and the lower organic phase containing microbial oil was transferred to a pre-weighed glass vial and dried at 80°C for 30 minutes, cooled in a desiccator, and weighed.
Transesterification of microbial oil
A previously published process, with minor modifications, was employed to convert isolated microbial oil to fatty acid methyl esters (FAME).14 The microbial oil was mixed with 334 µL of methanol:toluene (1:1, V/V) solution and 166 µL of 0.5 M sodium methoxide and incubated for 20 minutes at room temperature. Next, 50 µL of 37% HCL and 500 µL of 1M NaCl were added. Fatty acid methyl esters (FAMEs) were extracted by adding 2 mL of hexane and vortexing for 10 seconds. The reaction tube was centrifuged for 5 minutes at 4°C and 3000 rpm, and the upper (organic) phase was collected in a glass tube. The lower (water phase) was extracted twice again, but this time with the addition of 2 mL of hexane-chloroform mixture (4:1 v/v). The organic phase in the glass tube was dried and prepared for the GC measurement according to Hydrochloric acid method.
Gas chromatography
The required FAME sample was fed into gas-liquid chromatography and quantified using the Agilent 6850 GC (Agilent technologies), which was built with a BP21 Capillary column (50 m) and a FAME ionization detector (0.22 mm). A 2 µL sample was accurately injected with hydrogen as the carrier gas at a constant flow rate of 0.6 mL/min. The injector temperature was kept at 250°C and the detector temperature was kept at 275°C, followed by a 5-minute hold at 120°C with an increase of 3°C/min up to 180°C and a second-round increase of 10°C/min up to 220°C and a 31-minute hold at 220°C. Agilent Certify QA-QC Software was used to analyze the data.
The FAME characterization
The density, calorific value, specific gravity, viscosity, flash point, octene number, carbon residue, acid value, and ash content of FAME were determined, and the procedure was followed.
To determine density, the FAME sample was placed on a pycnometer and held at 4°C until it reached 15°C. The density of biodiesel was determined using the following formula after measuring the mass and volume of microbial oil FAME.
D(Kg/m2) = Pycnometer mass containing microbial oil FAME / Volume of microbial oil FAME
The calorific value was measured using a bomb calorimeter.15 FAME 0.5 g microbial oil was placed in a jar and placed inside an oxygenated bomb at 400 psi. The bomb was then placed in an insulated container filled with distilled water, and the fuse wires were attached to it. Two sticks connected to the fuse wires struck a Nichrome wire. The original temperature was recorded until it dropped to 0°C. To ignite the microbial oil FAME, a short circuit was created on the nichrome wire. The temperature was increased until it reached a steady temperature at a given moment in time, which was recorded. The following formula was used to compute the calorific value.
CV (Kj/Kg) = Weight of water + (water equivalent) x Temperature / Weight of the sample
Specific gravity, viscosity, flash point and octene number determinations
The specific gravity, viscosity, flash point and octene number was determined based on previously published methods.16 Briefly, for the specific gravity determinations, three independent 2 mL density vials were filled with 1 ml of microbial oil FAME obtained from RWW, another with ISP2 medium, and the third with 1 ml of distilled water. All density vials were sealed, refrigerated, and weighed without the presence of moisture. The specific gravity was calculated as the mass of microbial oil FAME samples divided by the mass of an equal volume of cold distilled water.
The viscosity was tested at 40°C using a Redwood viscometer. 50 ml of microbial oil FAME sample was collected for analysis in a measuring cylinder placed beneath the oil flow direction. The time of collecting was documented. The viscosity of microbial oil FAME samples was calculated using the formula below.
Viscosity of Microbial oil FAME (mm²/s) = A x Time ‒ B/ Time
Where A and B are two constants for the specific redwood viscometer.
The microbial oil FAME sample was placed in an equipment with a flame spark passed over it, and the flashpoint was determined as the point at which the sample flashed and became semi-solid.
Cetane number is the percentage of normal cetane by volume in a mixture of a-methyl naphthalene and normal cetane that exhibits the same delay in ignition characteristics as combustion advances in a standard engine under certain operational parameters.
Determination of carbon residue, acid value and ash content
For the determination of carbon residues, 5 g of each medium were weighed individually and placed in a separate pre-weighed heatproof glass bulb and then it was heated for 30 minutes at 450°C. then it was allowed to cool at room temperature, the weight was calculated based on the formula.
Carbon residue of microbial oil FAME = Initial Wt. of the bulb ‒ Final Wt. of the bulb x100 / Wt. of the Microbial oil FAME taken
The acid value was determined by titrating 0.5gm of sample against 30 ml of a 2:1 mixture of petroleum ether and ethanol. Phenolphthalein was used as an indicator. To calibrate the entire reaction, 0.1 N potassium hydroxide solution was utilized. The outcome was determined at the time during titration when the pink color developed. AV was calculated using the formula below.
Acid value (mg KOH/g) = Vol. of titrant x Normality of KOH x 56.1/ Wt. of sample (g)
For the determination of ash content, the microbial oil FAME from 25% RWW medium and ISP2 medium were weighed separately in two pre-weighed quartz crucibles and deposited in a 450°C prepared muffle furnace. After 30 minutes, the entirely burnt microbial oil to ash was removed and weighed again. The ash content of a microbial oil FAME sample was calculated using the formula below.
Ash content of microbial oil FAME = Initial weight of Crucible ‒ Final weight of Crucible x 100 / Weight of the Microbial oil FAME
Scientists are working to provide a viable alternative due to rising fuel consumption and the continuous depletion of natural resources. Microbial biomass is required for biofuel synthesis and is being investigated as a potential alternative fuel source for commercial fuel production. Biodiesel generated from microbial resources helps to reduce greenhouse gas emissions and reaches the maximum level of sustainability in the fuel economy.17
Bacterial screening and characterization
The bacteria were first isolated from the soils, and the consortium was created, as shown in Figure 1. Based on the findings, 53 Actinobacteria isolates were isolated from coastal soil sediments for this investigation. All of the isolates were tested and classified based on their capacity to collect lipids. The isolated bacterial species were thought to be extremophiles (microorganisms grown in harsh environments) since they were isolated from coastal habitats where they can increase lipid accumulation to survive during drought circumstances. Figure 2a shows Actinobacteria colonies obtained using the quadrant streak method. Individual colonies were further validated using TEM, as seen in Figure 2b. Streptomyces fradiae morphology revealed sporulated mycelium producing yellow colors and possessing an earthy odor with filamentous rods measuring 0.25 m in length. Figure 3a depicts the results of morphological identification using Sudan Black B staining. In total, 18 Actinobacterial isolates were discovered and categorized based on the findings. The presence of LL-diaminopimelic acid (LL-DAP) in actinobacterial cell walls allowed for easy identification of the actinobacterial isolates.18 According to morphological classification, the majority of the isolates belonged to the Streptomycetaceae and Mycobacterium families. Among the 53 examined isolates, isolate R6 generated microbial oil faster in 25% RWW than in the other dilutions and ISP2 medium. Streptomyces fradiae JJ1 MK733985.1 was identified molecularly after the strong strain R6 was sequenced alone utilizing 16S rRNA sequencing techniques. The partial gene sequence was examined and confirmed; Figure 3b depicts the partial gene sequence.
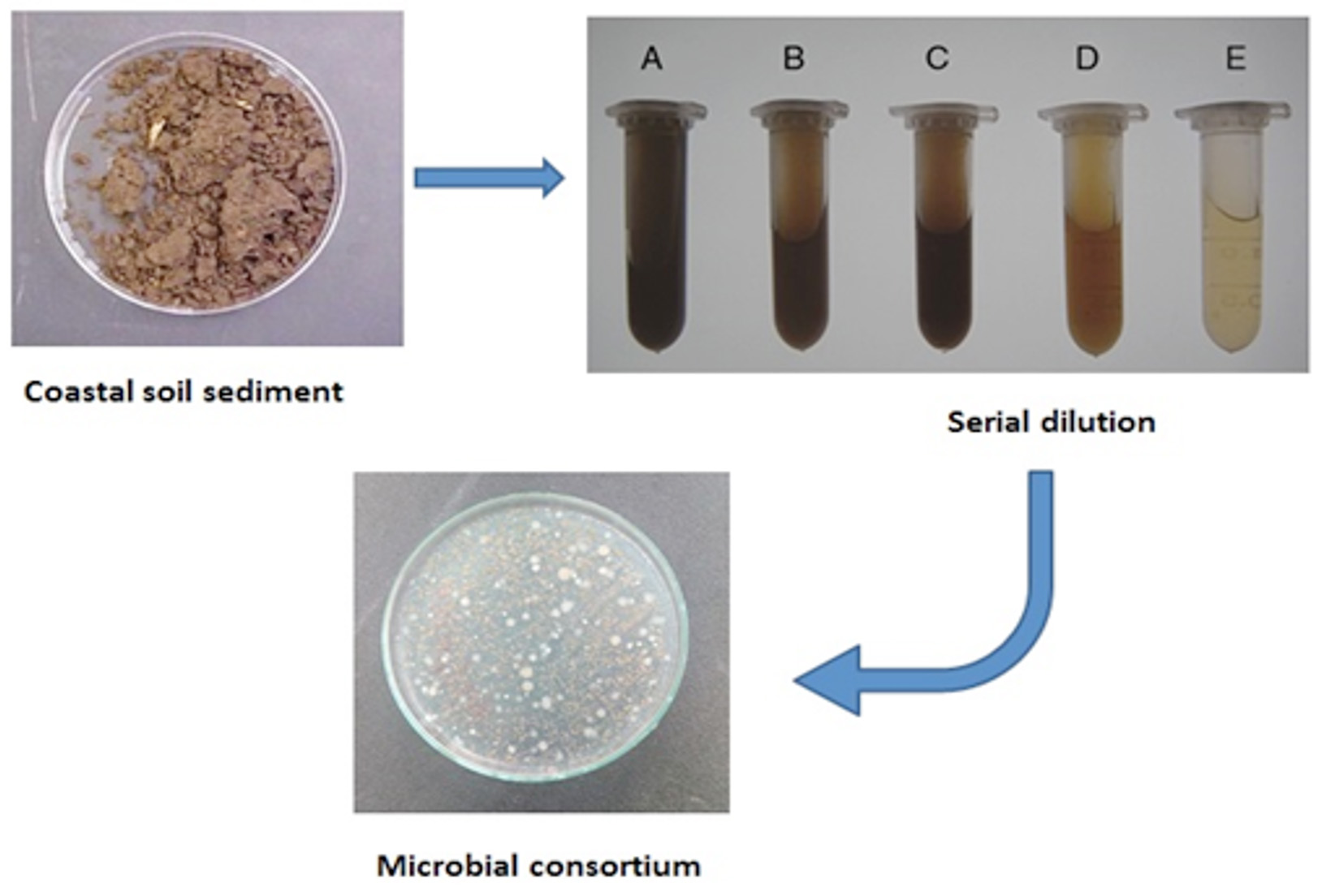
Figure 1. Actinobacteria Isolation Results from Coastal Soil Sediments
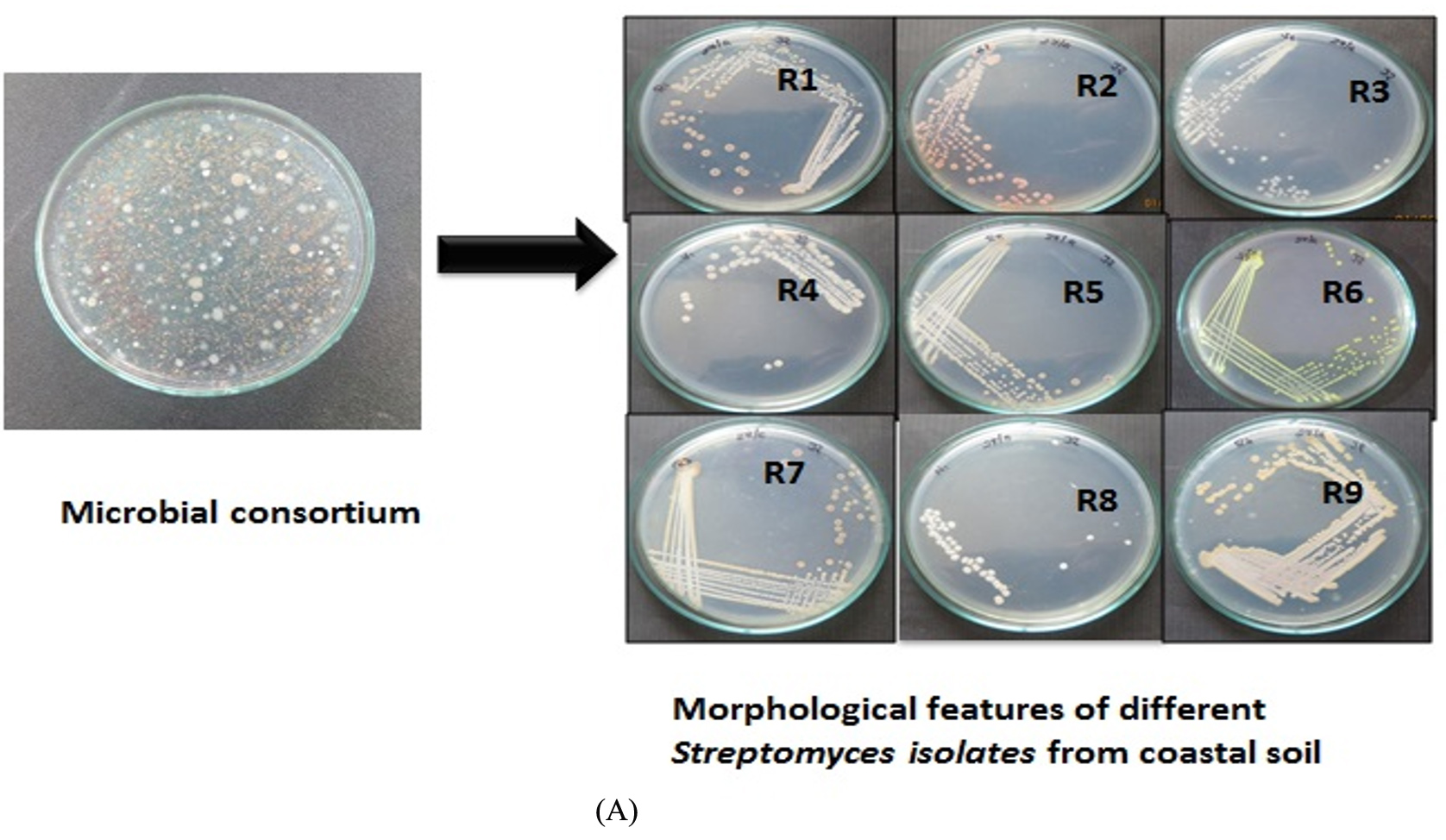

Figure 2. Isolated strains of Oleaginous Actinobacteria from Coastal Soil Sediments and their results of Streptomyces fradiae transmission electron microscopy findings

(A). Phylogenetic tree analysis showing the taxonomic affiliation of 16S rRNA sequences of actino bacteria Streptomyces fradiae

(B). Gene sequences of Streptomyces fradiae
Figure 3. Results of light microscopic observations of Actinobacterial lipids stained with Sudan Black B, as well as a partial gene sequence of Streptomyces fradiae and phylogenic tree analysis
Biodiesel production
Previously, RWW was employed as a source of plant nutrients as well as a very favorable medium for bacterial growth.19 Table 1 shows the composition of the RWW medium. The Streptomyces fradiae JJ1 MK733985.1 strain was grown in RWW and ISP2 media. The lipid content was studied. The results revealed small changes in lipid content, as illustrated in Figure 4. The lipid content of the 25% RWW medium was 7.5 g-1, while the ISP2 medium was 6.9 g-1. The amount of unsaturated fatty acids in the recovered microbial lipids is used to determine fuel quality. If microbial oil contains a high amount of unsaturation in the acyl chains, it tends to oxidize the fuel and cause polymerization in the engine oil, reducing the oxidative stability of the fuel. The microbial oil produced in this process consists of short-chain fatty acids with 12 to 18 carbon atoms and an alkyl chain, indicating a reduced level of unsaturation and acceptable fuel quality, particularly in diesel fuels. Microbial oil extraction is critical in assessing fuel quality. The most extensively used microbial oil extraction technology is the Bligh and Dyer method, which can extract microbial oil from both wet and dry biomass.20 Bacterial lipid has a smaller carbon residue than algal or mixed biodiesel because it has longer fatty acid chains.21 When compared to algal biodiesel, bacterial biodiesel has a higher gross heating value and a lower cetane number.22-23
Table (1):
Rice washed water components
Characteristics |
Values g/L |
|---|---|
Total solids |
40 |
Total suspended solids |
3.5 |
Volatile suspended solids |
3.1 |
Total soluble starch |
38.6 |
Total soluble chemical oxygen demand (COD) |
37 |
Total soluble nitrogen (TSN) |
0.59 |
Total soluble phosphorous (TSP) |
0.54 |
Ammonium nitrogen |
0.187 |
Orthophosphates |
0.102 |
Sulfate |
0.101 |
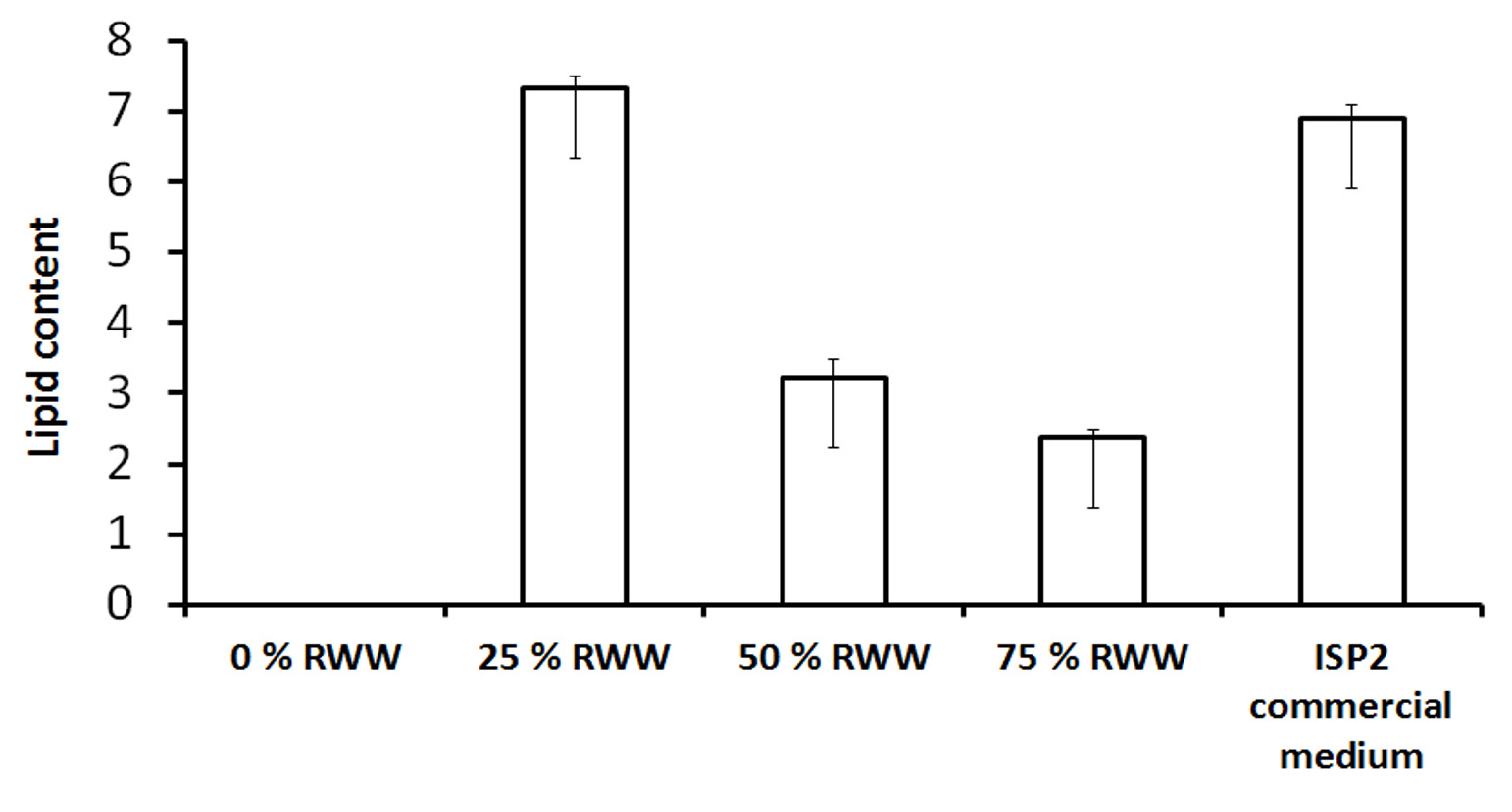
Figure 4. Shows the total lipid content of Streptomyces fradiae grown in 0%, 25%, 50%, and 75% RWW substrates in ISP2 medium, as determined by the sulfo-phospho vanillin assay
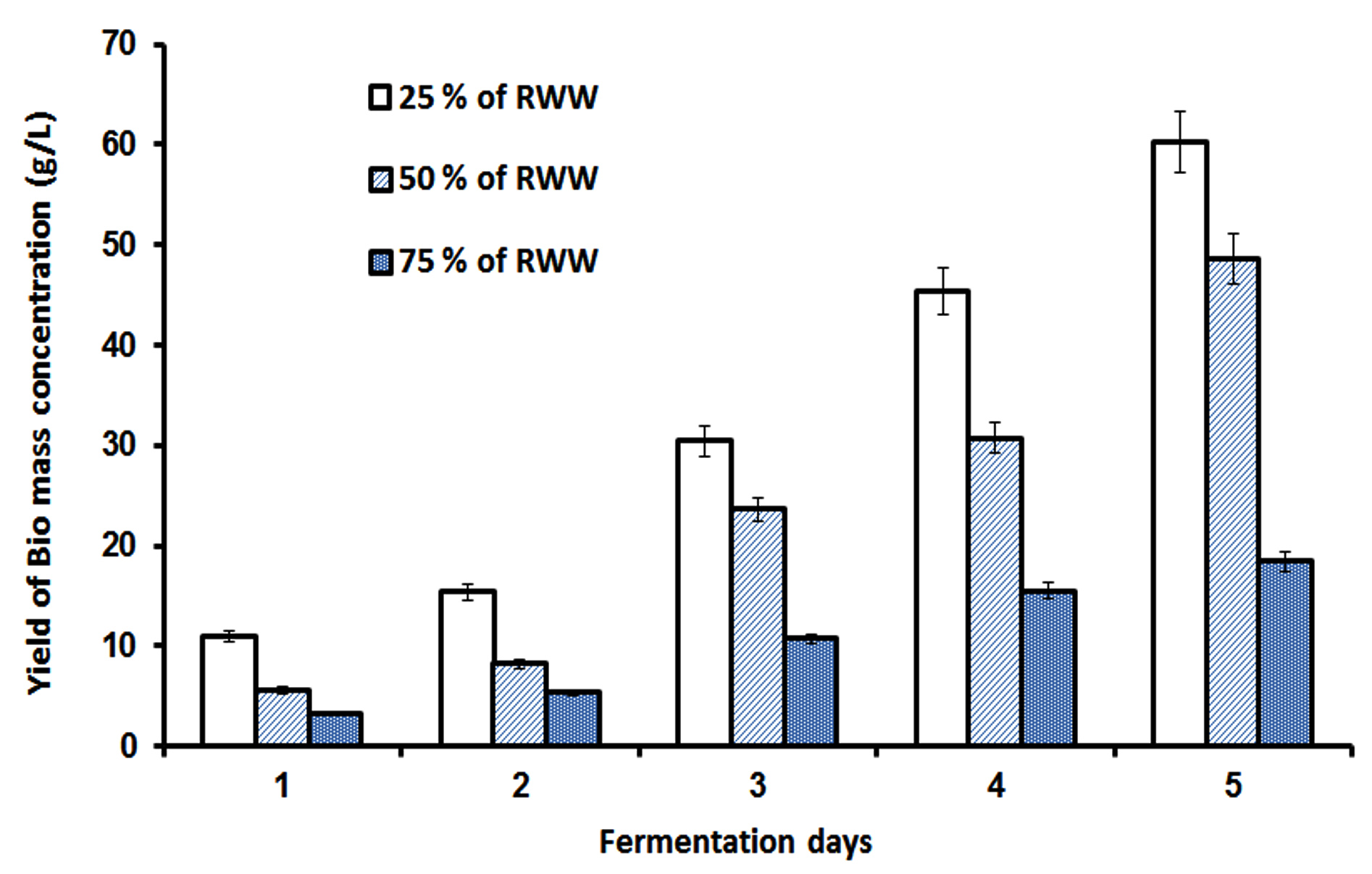
The research we performed also evaluated lipids’ capability as a reliable biodiesel feedstock using the low-cost substrate RWW, and fuel quality was evaluated using ASTM criteria. Based on the physicochemical properties of FAME formed from 25% RWW and ISP2 medium as provided in Table 1, the result suggests that microbial oil produced by Streptomyces fradiae JJ1 MK733985.1 is a potential source of biodiesel feedstock producer in the bioenergy industry.24 This finding was supported by previous study.25 Streptomyces fradiae JJ1 MK733985.1 lipids contain a high concentration of free fatty acids, which can be converted to biodiesel in a single step using acid-catalyzed alcoholysis. This is the most common method for esterifying free fatty acids.26 The biomass production in RWW and ISP2 medium was estimated up to 5 days, as shown in Figures 5a and 5b. Among the combinations, 25% RWW produced more biomass at 62 g/L, while ISP2 medium produced almost as much at 60 g/L. Figure 5c depicts the fermentation process designed in this work. In this process, seed and medium were prepared, after which the fermentation process was performed for up to 5 days, the cell mass was washed and recovered, and finally lipid extraction was performed using the Soxhlet extractor method.
Biofuel chemical analysis
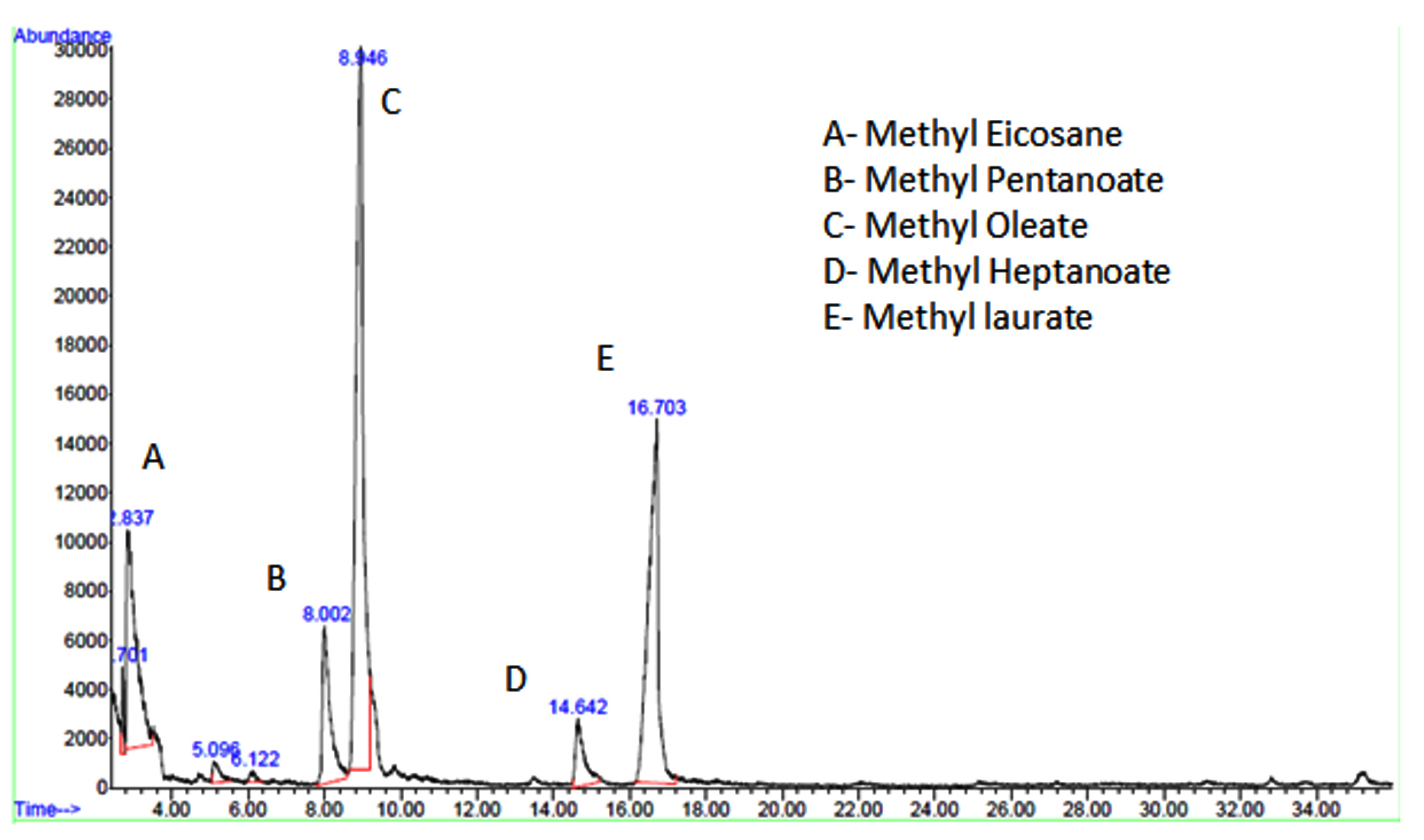
Figure 5d and Table 2 show the results of GC-MS analysis of the produced biodiesel. The results showed that the microbial oil FAME produced with 25% RWW contained both saturated and unsaturated fatty acids. The concentrations of long-chain saturated stearic acid with C18:0 was 145.38 mg/100 g, saturated hexadecanoic acid with C16:0 was 102.48 mg/100 g, and saturated pentadecanoic acid with C15:1 was 83.65 mg/100 g. The total fatty acid components made up 85.4 wt% of the total oil content.
Table (2):
Streptomyces fradiae generated fatty acids in 25%, 50%, and 75% RWW and ISP2 medium and it was estimated by using GC-MS method
Fatty acid chain |
25% RWW |
50% RWW |
75% RWW |
ISP2 |
|---|---|---|---|---|
C6:0 |
5.36 |
ND |
ND |
ND |
C8:0 |
11.74 |
ND |
ND |
ND |
C10:0 |
1.23 |
ND |
ND |
ND |
C11:0 |
1.83 |
ND |
ND |
ND |
C12:0 |
14.25 |
ND |
ND |
ND |
C13:0 |
58.43 |
5.22 |
ND |
2.97 |
C14:1 |
56.16 |
17.63 |
16.34 |
5.38 |
C14:0 |
72.14 |
31.53 |
46.21 |
55.4 |
C15:1 |
83.65 |
66.45 |
53.18 |
80.16 |
C15:0 |
1.68 |
ND |
13.28 |
ND |
C16:1 |
28.13 |
ND |
23.11 |
ND |
C16:0 |
102.48 |
98.16 |
67.12 |
98.35 |
C17:1 |
23.56 |
ND |
54.31 |
ND |
C17:0 |
78.14 |
23.11 |
ND |
ND |
C18:2 |
2.04 |
58.04 |
ND |
138.25 |
C18:1 |
53.91 |
58.23 |
ND |
85.38 |
C18:0 |
145.38 |
112.18 |
98.14 |
48.16 |
C20:1 |
ND |
96.13 |
67.42 |
16.32 |
C20:0 |
ND |
ND |
ND |
ND |
(A)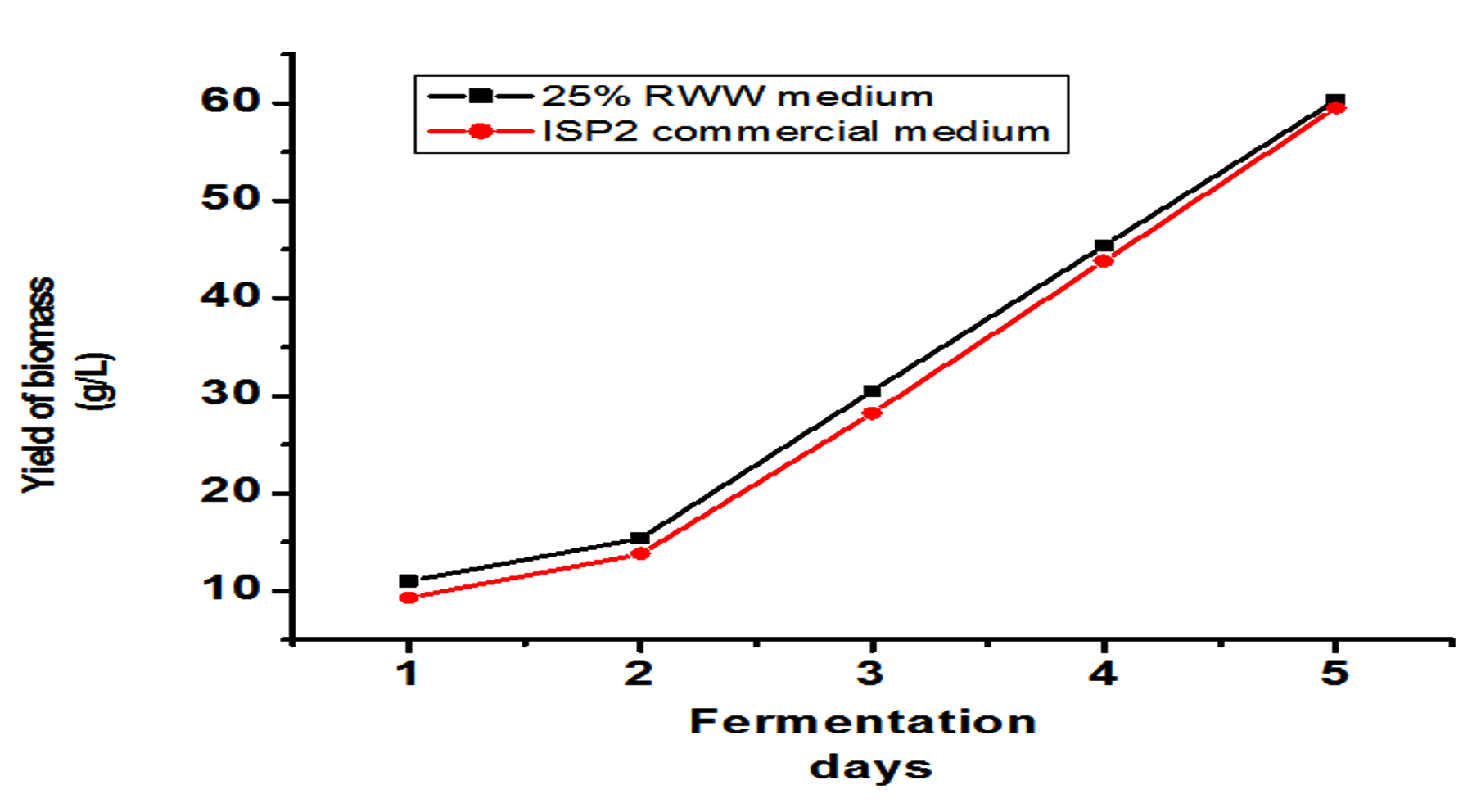
(B) (C)
(C)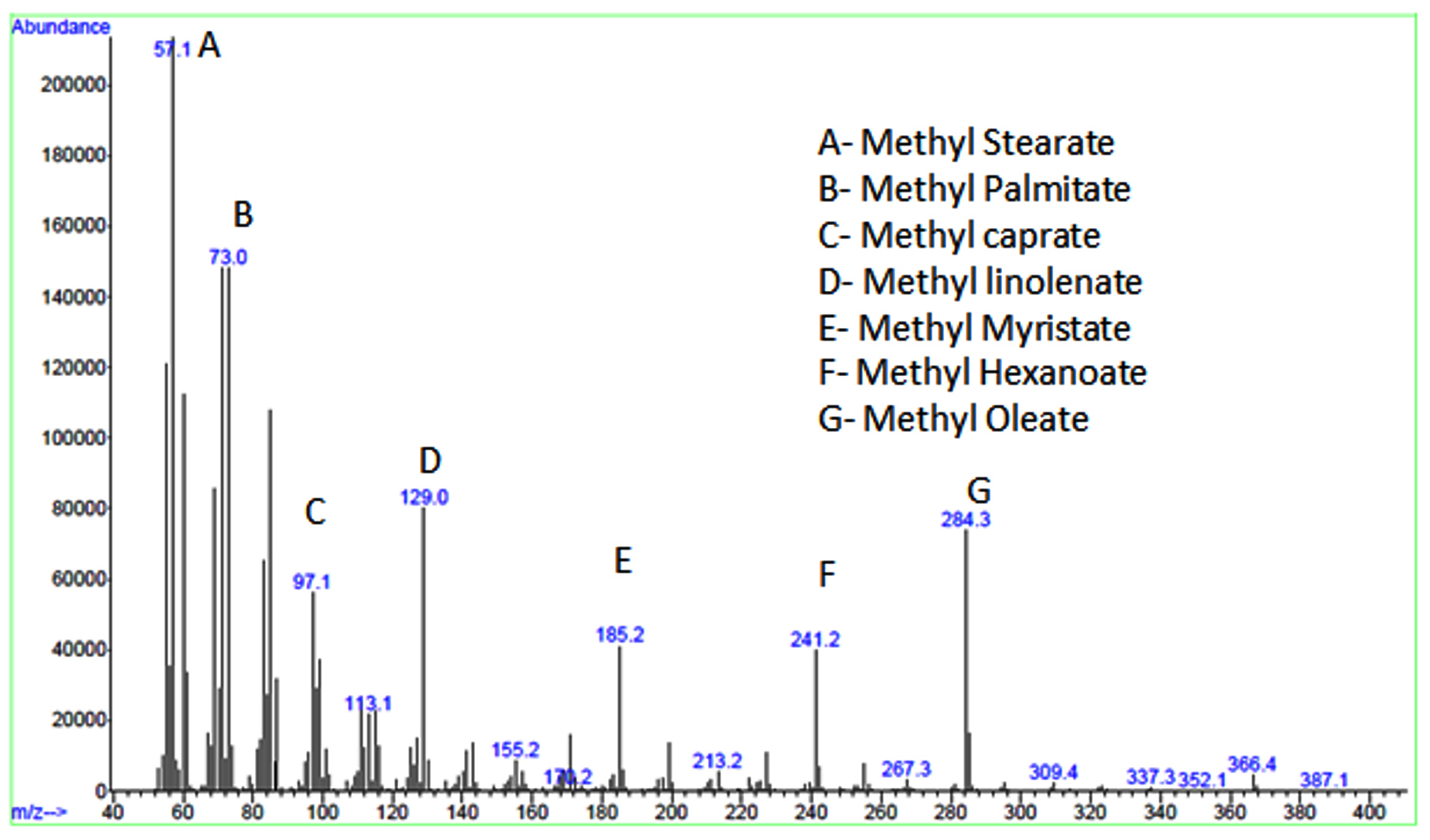
(D)
Figure 5. Biomass production results: (a) From day 1 to 5, compare biomass output from varied concentrations of RWW and commercial ISP2 medium. (b) From day 1 to 5, the biomass yield was compared between 25% RWW and commercial ISP2 medium. (c) Illustrations of FAME extraction from Streptomyces fradiae microbial oil. (d) After-transesterification GC-MS chromatogram results
Saturated stearic acid with C18:0 (112.18 mg/100 g) was found in high concentrations in microbial oil made from 50% RWW, followed by saturated hexadecanoic acid with C16:0 (98.16 mg/100 g) and saturated pentadecanoic acid with C15:1 (66.45 mg/100 g). Total fatty acid compounds made up 68.2 wt% of total fatty acid content.
In the 75% RWW fatty acid profile, saturated stearic acid C18:0 (98.14 mg/100 g) was the most abundant, followed by saturated hexadecanoic acid C16:0 (67.12 mg/100g) and saturated pentadecanoic acid C15:1 (53.18 mg/100g). Total fatty acid compounds made up 48.7 wt% of total fatty acid content.
The fatty acid chain length in ISP2 medium microbial oil FAME ranged from C6 to C18, with 9 fatty acids. Polyunsaturated long-chain linolenic acid C18:2 (138.25 mg/100g), long-chain saturated palmitic acid C16:0 (98.35 mg/100g), and monounsaturated long-chain oleic acid C18:1 (85.38 mg/100g) were found to have the greatest fatty acid contents. Total fatty acid compounds made up 83.2 wt% of total fatty acid content.
The microbial oil extracted from 25% RWW and ISP2 medium was transesterified to fatty acid methyl esters (FAME), and the physical, chemical, and mechanical properties of biodiesel were evaluated using ASTM standards, as shown in Table 3. According to the results of those measurements, the density of microbial oil FAME was 0.863 for 25% RWW and 0.835 for ISP2 medium. Microbial oil FAME from 25% RWW has a specific gravity of 0.864 and an ISP2 of 0.835. The gross heating value of microbial oil FAME produced from 25% RWW is 42498 kj/kg, which is somewhat higher than the gross heating value of ISP2 medium (42326 kj/kg). Furthermore, the cetane number of 25% RWW microbial oil FAME (37) was slightly higher than that of ISP2 produced microbial oil FAME (36). Furthermore, the ash level of microbial oil FAME formed from 25% RWW differed significantly from 0.005% to 0.003% for ISP2 medium derived microbial oil FAME. With fatty acid chain lengths more or less equivalent, the carbon residue computed for 25% RWW produced microbial oil FAME (7.3%) was lower than that determined for ISP2 derived microbial oil FAME (7.6%).
Table (3):
The physiochemical properties of biodiesel generated by Streptomyces fradiae in 25% RWW and ISP2 media were compared
Fatty acid chain |
ASTM standard |
25%RWW |
ISP2 |
|---|---|---|---|
Acid number (mg KoH/g of acid) |
0.46 |
0.43 |
|
Density (g/cm3) |
0.8800 |
0.863 |
0.835 |
Cetane number |
37 |
36 |
|
Ash content (% wt) |
0.005 |
0.003 |
|
Carbon residue (% wt) |
5-15 |
7.3 |
7.6 |
Flash point (°C) |
90-130 |
95 |
94 |
Viscosity |
6.81 |
6.84 |
|
Specific gravity |
0.864 |
0.845 |
|
Calorific value (Kj/Kg) |
42498 |
42326 |
Many research groups have previously reported pure cultures of numerous Streptomyces species.27 However, in this work, we isolated Streptomyces fradiae JJ1 MK 733985 from soil, which can store excess carbon as TriAcylGlycerols (TAGs) and also supplied a ready-to-use source for biodiesel synthesis, and their productivity and oil accumulation is comparable to previous findings. Previously reported papers only used free sugars; also, there is little information on the characteristics of the oil produced by these systems. However, in this study, the bacteria seen produced biodiesel from the cheapest source in a fermentation method that is simple to scale up in high volumes. Biodiesel production on a lab scale using waste rice washing water and commercial media. The findings demonstrated that the microbe can produce biofuel from both sources. The lipid structures seen in the cells and mycelia of the bacterium isolated in this study gather a significant amount of lipids. The GC-MS analysis indicated that the bacterium synthesized C14-C16 long-chain fatty acids as well as fatty acid methyl esters and triglycerides. The presence of fatty acid methyl esters suggested that the strain could produce fatty acid methyl esters directly from starch; however, methanol was not detected in GC-MS filtrates in this investigation. A comparison study was conducted between the 25%RWW medium and the ISP2 medium, and the results revealed the presence of common fatty acids with varying chain lengths. Stearic and oleic acids were also discovered in the lipids of Streptomyces fradiae JJ1 MK733985, which are primarily required for the synthesis of high-quality, low-emission biodiesel. Rice-washed water (RWW) was found to be an excellent substitute for commercial growth media, with promising lipid output and productivity. Additional approaches that combine genetic engineering and metabolic investigations will increase our understanding of microbial lipid buildup and its consequences.
This work proved the ability of the isolated oleaginous Actinobacteria Streptomyces fradiae JJ1 MK733985.1 to create microbial oil. The stain was grown in rice washed water (RWW) with the same efficiency as commercially available ISP2 medium. The study’s findings revealed a lipid production of 7.5 g/L, a biomass yield of 62 g/L, and a bioreactor productivity of 1-48 g/L/h within 48 h. The RWW source was five times less expensive and more widely available than commercially supplied ISP2 media. Future research to improve the efficiency of biofuel production using Streptomyces fradiae JJ1 MK733985.1 as a microbe with a low-cost medium on a wide scale will lead to a new breakthrough in inexpensive alternative fuel generation with minimal effort. It will additionally contribute to reducing waste and increase recyclable material recycling, thereby saving the environment.
ACKNOWLEDGMENTS
The authors would like to acknowledge the Director, Sathyabama Institute of Science and Technology, Chennai, for providing facilities. The author Janice R acknowledges the financial assistance provided by the Ministry of Human Resource Development, Department of Higher Education (F.NO.5-6/2013-TS-V11).
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Williams PJ, Laurens LM. Microalgae as biodiesel and biomass feedstocks: Review and analysis of the biochemistry, energetics and economics. Energy Environ Sci. 2010;3(5):554-590.
Crossref - Nayab R, Imran M, Ramzan M, et al. Sustainable biodiesel production via catalytic and non-catalytic transesterification of feedstock materials-A review. Fuel. 2022;328:125254.
Crossref - Salvi BL. Transesterification methods. Production of Biodiesel from Non-Edible Sources. 2022:117-151.
Crossref - Ferrero GO, Faba EM, Rickert AA, Eimer GA. Alternatives to rethink tomorrow: Biodiesel production from residual and non-edible oils using biocatalyst technology. Renewable Energy. 2020;150:128-135.
Crossref - Jacob A, Ashok B, Alagumalai A, Chyuan OH, Le PT. Critical review on third generation micro algae biodiesel production and its feasibility as future bioenergy for IC engine applications. Energy Convers Manag. 2021;228:113655.
Crossref - Cripwell RA, My R, Treu L, et al. Additional glucoamylase genes increase ethanol productivity on rice and potato waste streams by a recombinant amylolytic yeast. Bioresour Technol. 2023;388:129787.
Crossref - Patel A, Karageorgou D, Rova E, et al. An Overview of Potential Oleaginous Microorganisms and Their Role in Biodiesel and Omega-3 Fatty Acid-Based Industries. Microorganisms. 2020;8(3):434.
Crossref - Tomas-Pejo E, Morales-Palomo S, Gonzalez-Fernandez C. Microbial lipids from organic wastes: Outlook and challenges. Bioresour Technol. 2021;323:124612.
Crossref - Mhlongo SI, Ezeokoli OT, Roopnarain A, et al. The Potential of Single-Cell Oils Derived From Filamentous Fungi as Alternative Feedstock Sources for Biodiesel Production. Front Microbiol. 2021;12:637381.
Crossref - Al-Shorgani NKN, Al-Tabib AI, Kadier A, Zanil MF, Lee KM, Kalil MS. Continuous Butanol Fermentation of Dilute Acid-Pretreated De-oiled Rice Bran by Clostridium acetobutylicum YM1. Sci Rep. 2019;9(1):4622.
Crossref - Shaigani P, Awad D, Redai V, et al. Oleaginous yeasts- substrate preference and lipid productivity: a view on the performance of microbial lipid producers. Microb Cell Fact. 2021;20(1):220.
Crossref - Srinivasan N, Thangavelu K, Sekar A. Sanjeev B, Sivakumar U. Aspergillus caespitosus ASEF14, an oleaginous fungus as a potential candidate for biodiesel production using sago processing wastewater (SWW). Microb Cell Fact. 2021;20:179.
Crossref - Yaakob MA, Mohamed RMSR, Al-Gheethi A, Gokare RA, Ambati RR. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells. 2021;10(2):393.
Crossref - Langseter AM, Dzurendova S, Shapaval V, Kohler A, Ekeberg D, Zimmermann B. Evaluation and optimisation of direct transesterification methods for the assessment of lipid accumulation in oleaginous filamentous fungi. Microb Cell Fact. 2021;20(1):59.
Crossref - Kaisan MU, Anafi FO, Nuszkowski J, Kulla DM, Umaru S. Calorific value, flash point and cetane number of biodiesels from cotton, jatropha and neem binary and multi-blends with diesel. Biofuels. 2020;11(3):321-327.
Crossref - Raja VK, Muthu GMI, Goswami L, et al. Analytical methods in biodiesel production. Biomass valorization to bioenergy, 2020:197-219.
Crossref - Nabayi A, Sung CTB, Zuan ATK, Paing TN, Akhir NIM. Chemical and Microbial Characterization of Washed Rice Water Waste to Assess Its Potential as Plant Fertilizer and for Increasing Soil Health. Agronomy. 2021;11(12):2391.
Crossref - Nabayi A, Sung CTB, Zuan ATK, Paing TN. Fermentation of Washed Rice Water Increases Beneficial Plant Bacterial Population and Nutrient Concentrations. Sustainability. 2021;13(23):13437.
Crossref - Lee EH, Jeong HM, Kim EA, Lee YR, Shim JH. Development of Fermented Rice Water to Improve the Quality of Garaetteok, a Traditional Korean Rice Cake. Foods. 2023;12(3):642.
Crossref - Ulmer CZ, Jones CM, Yost RA, Garrett TJ, Bowden JA. Optimization of Folch, Bligh-Dyer, and Matyash sample-to-extraction solvent ratios for human plasma-based lipidomics studies. Anal Chim Acta. 2018;1037:351-357.
Crossref - Koh HG, Kang NK, Kim EK, et al. Isolation and Characterization of Novel Chlorella Species with Cold Resistance and High Lipid Accumulation for Biodiesel Production. J Microbiol Biotechnol. 2019;29(6):952-961.
Crossref - Kuranska M, Malewska E. Waste cooking oil as starting resource to produce bio-polyol – analysis of transesteryfication process using gel permeation chromatography. Industrial Crops and Products. 2021;162:113294.
Crossref - El-Sheekh M, El-Dalatony MM, Thakur N, Zheng Y, Salama ES. Role of microalgae and cyanobacteria in wastewater treatment: Genetic engineering and omics approaches. Int J Environ Sci Technol. 2022;19:2173-2194.
- Folayan AJ, Anawe PAL, Aladejare AE, Ayeni AO. Experimental investigation of the effect of fatty acids configuration, chain length, branching and degree of unsaturation on biodiesel fuel properties obtained from lauric oils, high-oleic and high-linoleic vegetable oil biomass. Energy Reports. 2019;5:793-806.
Crossref - Zhang Y, Zhong Y, Lu S, Zhang Z, Tan D. A Comprehensive Review of the Properties, Performance, Combustion, and Emissions of the Diesel Engine Fueled with Different Generations of Biodiesel. Processes. 2022;10(6):1178.
Crossref - Gopi R, Thangarasu V, Ramanathan A. A critical review of recent advancements in continuous flow reactors and prominent integrated microreactors for biodiesel production. Renew Sustain Energy Rev. 2022;154:111869.
Crossref - Alam K, Mazumder A, Sikdar S, et al. Streptomyces: The biofactory of secondary metabolites. Front Microbiol. 2022;13:968053.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.