


ISSN: 0973-7510
E-ISSN: 2581-690X
Rhizoctonia solani is an important plant pathogen which is responsible for causing a number of plant diseases in a variety of cropping plants. The current studies were carried out to explore antifungal potential of bacteria isolated from rhizospheric region of Daucus carota against Rhizoctonia. Of the total 21 bacterial isolates isolated, only 3 isolates namely CVO-3, CVO-4 and CRY-O-2 were shown antifungal potential against Rhizoctonia. Besides having antifungal potential these isolates also have some plant growth promoting attributes that will be an extra benefit of using these isolates as a biofungicides against R. solani. Thus, these isolates could be used as biofungicides against Rhizoctonia to prevent a variety of plant diseases like sheath blight in Oryza sativa, stem canker and black scurf in Solanum tuberosum and seedling diseases in Brassica napus. The isolates CVO-3, CVO-4 and CRY-O-2 were also characterized biochemical level by using methods based on Determinative Bacteriology given in Bergey’s manual and at molecular level using 16S rRNA sequencing and sequences of 16S rDNA segment were submitted to NCBI in GenBank database with the accession numbers OR775574 (Lysinibacillus boronitolerans ss3cvo3), OR775575 (Lysinibacillus boronitolerans ss4cvo4) and OR708523 (Shouchella rhizosphaerae ss1cryo2). The present investigation is an approach towards the use of these bacterial isolates from carrot rhizosphere to control the activities in the rhizosphere of other cropping plants.
Daucus carota, Rhizoctonia solani, PGPR, Lysinibacillus boronitolerans, Shouchella rhizosphaerae
Rhizoctonia solani is well known ubiquitous fungal pathogen of number of several agriculturally important crop plants. Pathogens survives in soil in form of sclerotia or mycelia on infected plant debris and also survives in water logged condition as it floats on surface of water.1 In suitable conditions the spores germinate and infect crop plants.2 Due to infection on several crop plants pathogen cause qualitative and quantitative damage leading to significant loss in productivity.3-5 Some isolates are highly virulent and responsible for causing pre- or post-emergence of damping-off and root rot with characteristic water-soaked lesions on root and hypocotyl which ultimately results in root necrosis and cortex tissue maceration.6 Sheath blight in rice plant caused through R. solani is a serious disease of rice.7 R. solani is known to related with root rot complex of rapeseed.8 R. solani, the pathogen causes sheath blight on Oryza sativa and black scurf of Solanum tuberosum.3 R. solani has been most frequently identified and studied in having range of host plant species more than 250 and also capable of causing disease in several diverse environmental conditions.9,10 R. solani shows pathogenicity against Poaceae, Linaceae, Moraceae, Brassicaceae, Solanaceae, Fabaceae, Amaranhtaceae, Rubiaceae, Asteraceae, Malvaceae and Araceae.11 R. solani has already been reported as a most common pathogen present in fields where winter wheat, i.e. Triticum aestivum is grown as rotation crop with oil seed rape12. Vegetatively propagated plants are susceptible to Rhizoctonia as it causes rot at the base of the cutting. The rhizospheric regions are hot spots for microorganisms which contains both the beneficial as well as harmful microorganisms. The plant growth promoting rhizobacteria (PGPR) comprises a group of beneficial bacteria residing in rhizosphere of plants and can promote the growth of plants.13 Growth promotion activities of PGPR relies either on direct mechanisms involving enhancement in nutrient supply and production of growth promoters or on indirect mechanisms involving inhibition of the pathogenic microbial forms capable to establish infection in host plants. Application of PGPR is a modern, ecofriendly and sustainable approach in agriculture which acts as an alternative of chemical fertilizer and pesticides. The current study was carried out to search out antifungal potential of bacterial isolates from the rhizospheric region of the D. carota against R. solani. The bacterial cultures were isolated from carrot rhizosphere and isolates having antifungal potential against R. solani were screened for their plant growth promoting attributes. The present investigation is an approach towards the use of rhizospheric bacterial isolates of D. carota to manage the damage of other crop plants and reduction in yield. Microbes are important role players in the soil and it is known that mixing of soil adds heart to the soil. Plants growing in the field also regulates the growth and productivity of next crop sown in the field, this is due to the activity of rhizospheric microbes associated with a particular plant which enhance productivity of other crop plants as it is known to happens naturally during crop rotation14 which is an aspect to increase the soil fertility, maintaining the quality of soil, interference in pest cycles and weed subduing.15 These processes are mediated by activities of soil microorganisms,16-18 which have given a clue to use rhizospheric bacteria of the carrot for prevention of diseases in some other crop plants.
Sampling and isolation of rhizospheric bacteria
Soil samples were collected from various orange carrot grown fields of District Jaunpur. The soil adhered on the roots were collected carefully from carrot plants growing in fields. The soil samples were smashed and sieved through 9 mesh per square inch sieve. One gram of this soil sample was taken from different fields of carrot and added to 9 ml distilled water. Ten-fold dilutions were prepared and spread on different media containing plates. For isolation, different selective media such as N-free Jensen’s Agar, NBRI-P media, Methyl Red Agar, and Crystal Violet Agar were used. Plates were then incubated for 3 days at 30°C in incubator for the growth of rhizospheric microbes. Few bacterial colonies were selected after the appearance of the colonies on plates. Selection was based on different morphological features and growth on different media plates. The pure culture of the selected bacterial isolates was prepared by streaking the bacterial isolates on respective media plates and further repeated sub-culturing on Nutrient Agar (NA) plates. All the culture media and compositions used were from Himedia (Mumbai, India).
Determination of antifungal activities
All the bacterial isolates were tested for their antifungal potential against fungal pathogen, R. solani by using modified cross streak method.19 The experiment fungus R. solani was obtained from culture collection center of ICAR-NBAIM (Indian Council of Agricultural Research-National Bureau of Agriculturally Important Microorganisms), Kushmaur, Mau Nath Bhanjan, UP, 275103, India. Initially, fungal cultures were grown on the PDA (Potato Dextrose Agar) plates and disks of this fungus was prepared by cutting the plugs by using cork borer from the fungal plates. Then plugs of fungus placed in the center of NA PDA (1:1) mixed plates. Finally, bacterial isolates were streaked both sides of fungal plugs at equal distance of 1 cm.20 A control plate also made with only fungal plug on petri plate without bacterial isolates. Then, plates incubated in BOD at 30°C for 5-7 days.
Determination of biochemical activities of bacterial isolates
All three selected bacterial isolates were characterized for their biochemical activities like carbohydrate fermentation (glucose, lactose, fructose and mannitol), IMViC test and catalase test. The results of biochemical tests were analysed using Bergey’s manual of Determinative Bacteriology.21
Determination of hydrolytic enzyme production
Hydrolytic enzyme production like protease and amylase was determined through point inoculating bacterial isolates on Skim Milk Agar (SMA) plates and on Starch Agar plates respectively.22
Determination of plant growth promoting activities
The bacterial isolates having antifungal potential against Rhizoctonia were screened for their plant growth promotion activities like:
Determination of HCN production
HCN producing abilities of isolates were analysed by using method of Bakker and Schipper 1987.23 Isolates were streaked on nutrient agar slants (having 4.4 g/l of glycine) in test tubes. A small strip of filter paper (Whatman No. 1) of about 1cm width and 4 cm length was cut and soaked in picric acid solution and placed by hanging it at the top of the test tubes. Mouth of test tubes were covered properly with parafilm (Himedia) to avoid any contamination. The test tubes were incubated at 30°C for 4-5 days. After incubation the positive results were recorded qualitatively as filter paper turns reddish-brown from yellow.
Determination of Ammonia production
Ammonia production by selected three isolates were observed according to Cappuccino and Sherman 1992.22 Isolates were inoculated in peptone water broth and incubated in BOD incubator at 37°C for 72 hours. Positive results were recorded as change of colour of broth from yellow to brown after addition of freshly prepared Nessler’s reagent.
Determination of Siderophore production
Production of siderophore were determined by inoculating bacterial isolates on CAS media plates (Chrome Azoural S agar).24,25 A yellow clear zone formed around colonies, indicated positive result for siderophore production by bacterial isolates.
Determination of Phosphate solubilization
The phosphate solubilization activity of isolates were determined via point inoculation of the pure bacterial isolates on plates containing Pikovskya’s medium. Plates were then incubated at 30°C for 3 days in BOD incubator. Finally examined for the formation of clear zone around the bacterial colonies for positive result of phosphate solubilization.26
Determination of Potassium solubilization
Potassium solubilization by the bacterial isolates were determined by point inoculation and formation of zone after incubation on plates containing Aleksandrov medium.27
Determination of Cellulase production
Cellulase production activity was determined by point inoculation of bacterial isolates on CMC media plates followed by incubation of 3 days at 30°C. After 3 days of incubation, the plates were flooded by 1% Congo red solution (15 min). Then, the solution was drained off and plates were flooded with 1 M NaCl solution (15 min). Cellulase production by the isolates was confirmed by formation of a clear zone around the colonies.28
Determination of IAA production
The IAA (Indole acetic acid) production by isolates were determined via using colorimetric method in presence as well as in absence of tryptophan by using method of Brick et al.29 Protein produced by bacterial isolates were estimated quantitively by using method of Lowry et al.,30 Values were determined with standard curve made by using concentrations of IAA ranging from 10-100 µg/ml separately for both IAA and Protein. Finally, the IAA production by isolates were estimated in micro gram per gram (µg/g) of protein produced by bacterial isolates.31
Determination of Gibberellic acid production
Production of phytohormone-gibberellin (Gibberellic Acid) by bacterial isolates were determined by using protocol of Ashry et al.32 Bacterial isolates were inoculated in nutrient broth having tryptophan (1 mM), incubated at 37°C for 48 hours keeping it in rotatory shaker at 100 rpm. Bacterial isolates were transferred in centrifuge tube and then centrifuged at 10,000 rpm for 5 min. The supernatant was collected and 1 ml of Folin-Ciocalteu reagent (FCR) and 1 ml of HCl is added in test tubes. Appearance of greenish blue colour was recorded as positive result for production of Gibberellic acid. Tubes were then kept in boiling water bath for 5 minutes and then cooled at room temperature. Quantitative estimation of gibberellic acid production was estimated after taking absorbance at 760 nm. Standard curve was prepared by using gibberellic acid (range used, 10-100 µg/ml) solution.
Molecular characterization of Isolates
Molecular characterization of isolates CVO-3, CVO-4 and CRY-O-2 was done by using 16S rRNA sequencing gene. The genomic DNA and 16S rRNA gene of bacterial isolates from rhizospheric region of D. carota, was amplified by using 16s forward (5’GGATGAGCCCGCGGCCTA3’) and reverse (5’CGGTGTGTACAAGGCCCGG3’) primers. Isolates CVO-3, CVO-4 and CRY-O-2 were characterized at molecular level by sequencing of 16S rRNA gene. 1.5 kbp, high–fidelity PCR polymerase enzyme was used for amplification of 16S rDNA fragment and product obtained was sequenced bi-directionally. The identification of bacteria and their closest neighbours was done by aligning and analysing sequence data using GenBank database through NCBI-BLAST (NCBI BLASTR home page). The nucleotide sequences were submitted to NCBI GenBank database. Inference of evolutionary history, drawn by using Neighbor-Joining method.33 The evolutionary distance was computed through the Maximum Composite Likelihood method34 and in units of number of base substitutions per site. Evolutionary analyses were conducted in MEGA11.35
Isolation of rhizospheric bacteria
Rhizospheric soil of orange carrots were used for isolation of PGPR. Total 21 bacterial strains were isolated and purified from rhizospheric region of the different orange carrot grown fields of District Jaunpur.
Determination of antifungal activities and screening of bacterial isolates
Out of 21 bacterial isolates were tested for their antifungal activities against Rhizoctonia, only 3 bacterial isolates CRY-O-2, CVO-3 and CVO-4 gave positive result for antifungal activities against Rhizoctonia (Figure 1 and 2). Antifungal activities of these bacterial isolates were estimated quantitatively on the basis of reduction in growth of Rhizoctonia as compared to control plate. All these experiments were performed in triplicates.
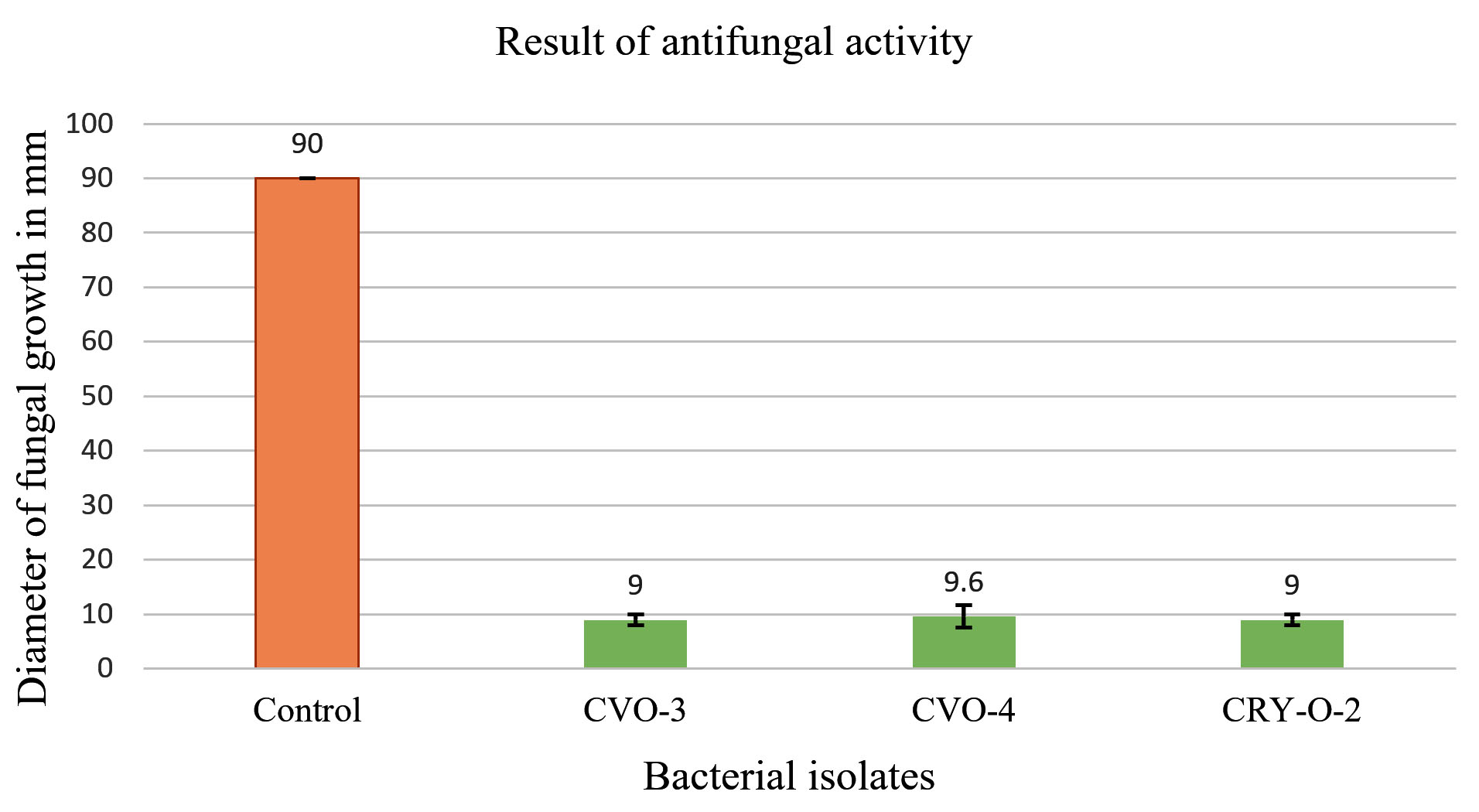
Figure 1. Antifungal Activities of bacterial isolates against Rhizoctonia
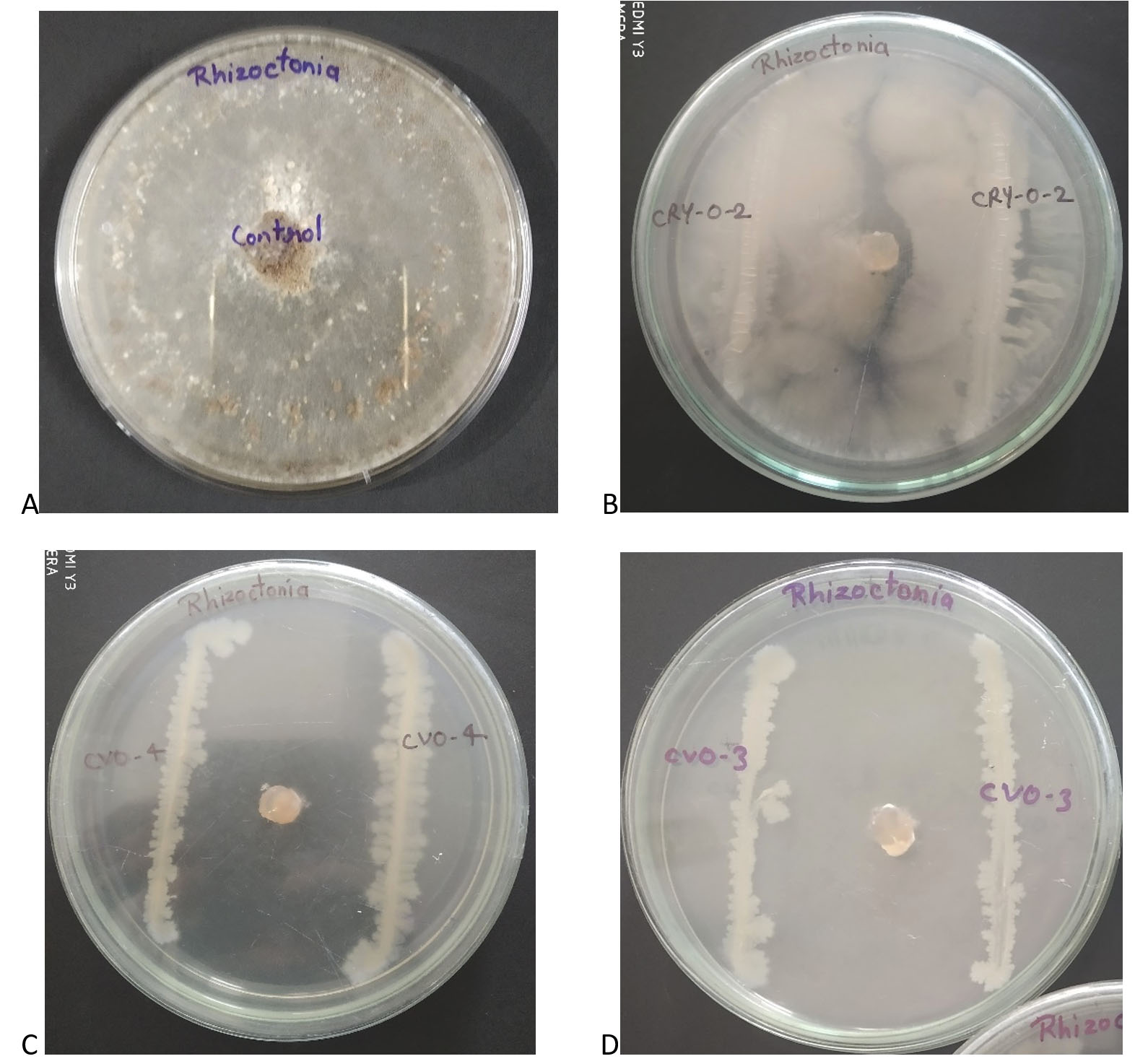
Figure 2. Results of the antifungal activity of bacterial isolates against R. solani done by using cross streak method. A. Control plate showing full fungal growth without any inhibition, B. Antifungal activity of CRY-O-2, C. Antifungal activity of CVO-4 and D. Antifungal activity of CVO-3
Characterization of isolates
Gram Staining procedure was used for initial characterization of the isolates. All the selected three isolates CRY-O-2, CVO-4 and CVO-3 were found as Gram’s positive.
Determination of biochemical activities
All three isolates were screened for their biochemical activities such as carbohydrates fermentation, IMViC test and catalase test. All the three isolates were tested for glucose, fructose, mannitol and lactose fermentation, only CVO-3 and CVO-4 gave negative result for lactose fermentation but gave positive result for glucose, mannose and fructose with acid production. On the other hand, CRY-O-2 fermented all four carbon sources with acid production. Isolates CRY-O-2, CVO-3 and CVO-4 gave positive result for catalase test. CRY-O-2 gave positive result for Indole production and Voges Proskauer while gave negative result for Methyl Red and Citrate Utilization. On the other hand, CVO-3 and CVO-4 gave negative result for Indole production, Voges Proskauer and Methyl Red and gave positive result for only Citrate Utilization. All three isolates gave positive result for catalase test. These experiments were done in triplicates and result are given in Table 1.
Table (1):
Result of IMViC test, Catalase test and Carbohydrate fermentation
| Culture Name | Indole production | Methyl Red | Voges Proskauer | Citrate Utilization | Catalase Test | Carbohydrate fermentation | |||
|---|---|---|---|---|---|---|---|---|---|
| Glc. | Lact. | Mann. | Fruc. | ||||||
| CRY-O-2 | +ve | -ve | +ve | -ve | +ve | AG | AG | AG | AG |
| CVO-3 | -ve | -ve | -ve | +ve | +ve | AG | – | AG | AG |
| CVO-4 | -ve | -ve | -ve | +ve | +ve | AG | – | AG | AG |
Abbreviations Glc. = glucose, Lact. = lactose, Mann. = mannose and Fruc. = fructose
Determination of hydrolytic enzyme production
All three isolates were tested for hydrolytic enzymes production such as protease and amylase. Isolate CVO-3 gave 8.67 mm zone on skim milk agar plate for protease production and 6.33 mm zone on Starch agar plate for amylase production. CVO-4 gave 8.67 mm zone for protease and 9.0 mm zone for amylase and CRY-O-2 gave 13.33 mm zone for protease and 16.0 mm zone for amylase. Results are shown in Figure 3.
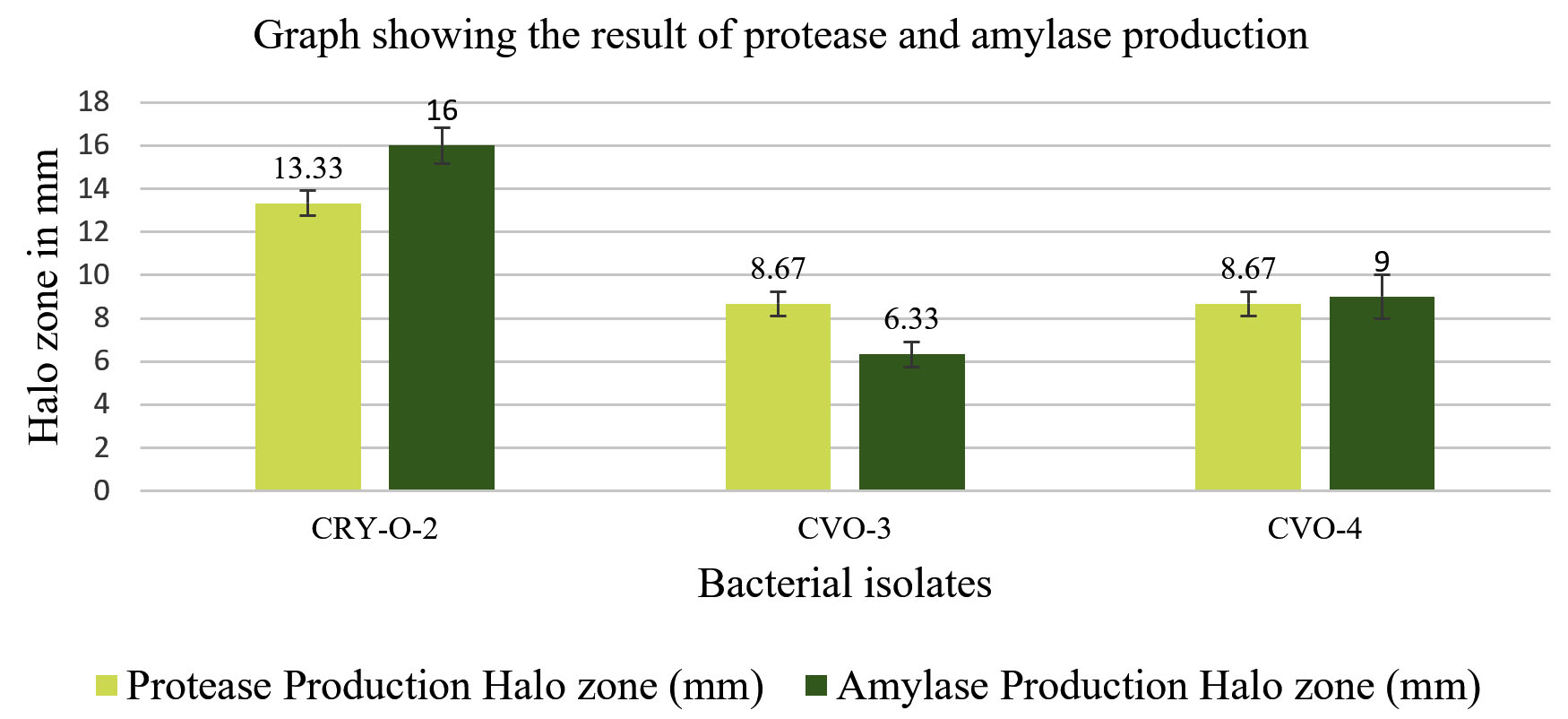
Figure 3. Result of hydrolytic enzyme production by CRY-O-2, CVO-3 and CVO-4 (protease in yellow bars and amylase in green bars)
Determination of the Plant Growth Promoting Activities
Isolates were qualitatively estimated for HCN and Ammonia production. Isolates CVO-3 and CVO-4 gave negative result for HCN production. On the other hand, isolate CRY-O-2 gave positive result for the same. All the three bacterial isolates gave positive result for ammonia production (Table 2). Other plant growth enhancing attributes like siderophore production, solubilization of phosphate, potassium solubilization, production of cellulase and IAA production estimated quantitively. CVO-3 gave positive result for production of siderophore, phosphate solubilization and cellulase production while gave negative result for potassium solubilization. CRY-O-2 and CVO-4 gave positive result for all 4 activities such as siderophore production, phosphate solubilization, potassium solubilization, cellulase production. All the three bacterial isolates also gave positive result for IAA production in presence as well as in absence of Tryptophan (Figure 4 and 5).
Table (2):
Result of HCN and ammonia production
Plant Growth Promoting Activity (Qualitative analysis) |
CVO-3 |
CVO-4 |
CRY-O-2 |
|---|---|---|---|
HCN Production |
Negative |
Negative |
Positive |
Ammonia Production |
Positive |
Positive |
Positive |
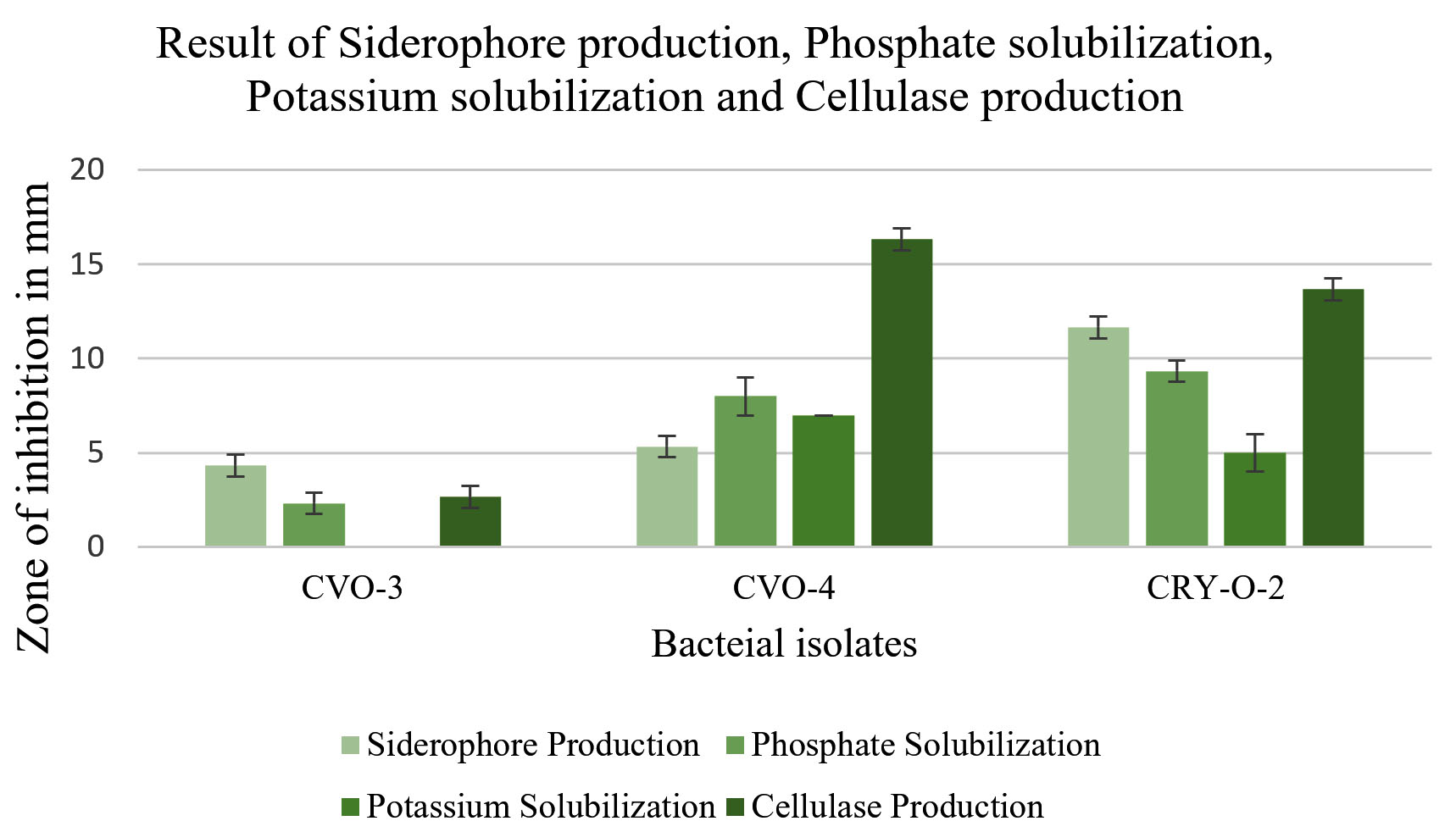
Figure 4. Result of Siderophore production, Phosphate solubilization, Potassium solubilization and Cellulase production
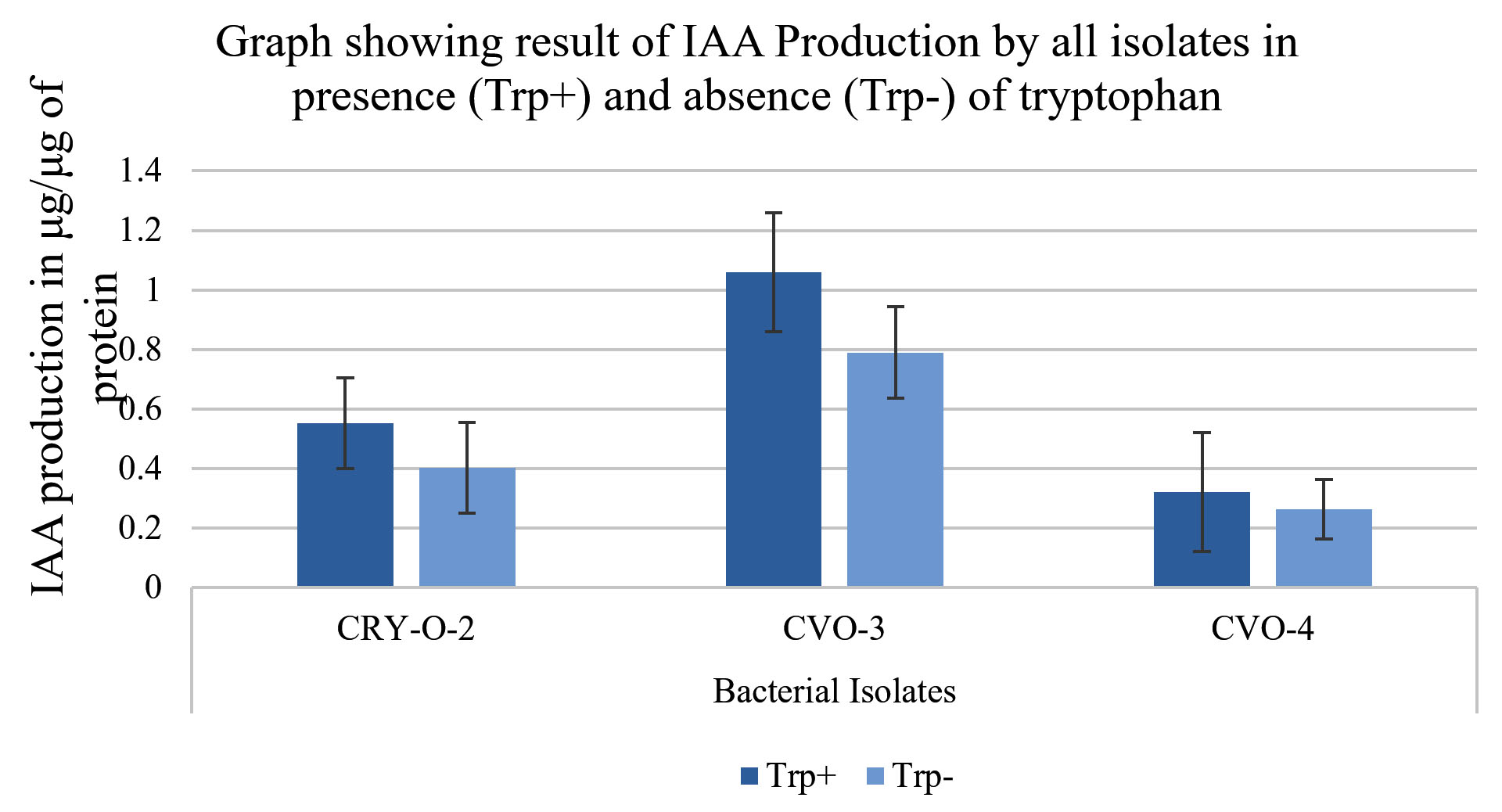
Figure 5. Result of IAA Production in presence (Trp+) and absence (Trp-) of tryptophan
Result of Gibberellic Acid production
All three bacterial isolates gave positive result for the production of gibberellic acid (GA). CRY-O-2 gave minimum result for GA production (23.89 µg/ml), while CVO-3 gave maximum GA production (38.41μg/ml) (Figure 6).
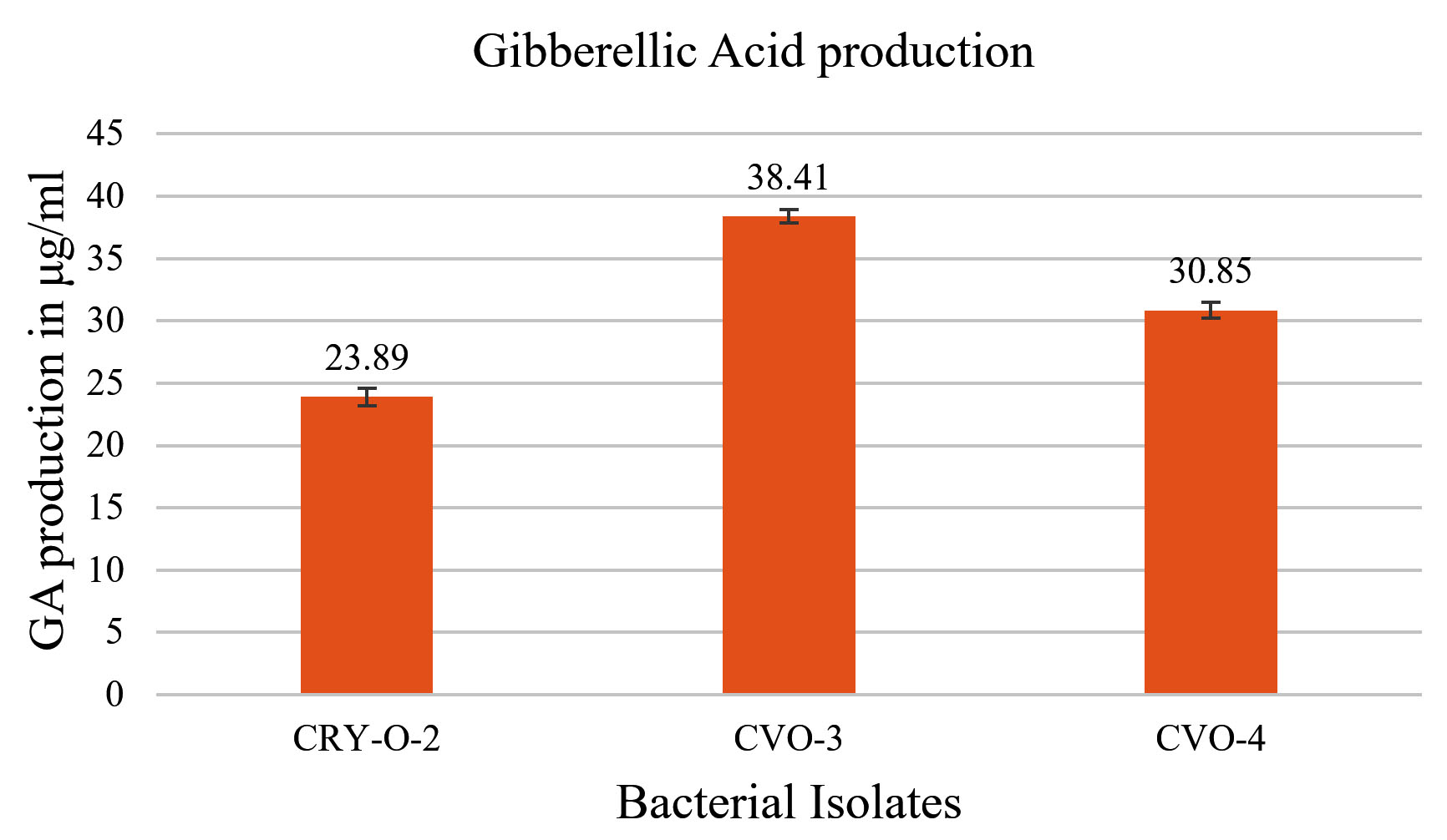
Figure 6. Result of Gibberellic Acid (GA) production by all three isolates
Result of molecular characterization
Molecular characterization of isolates CVO-3, CVO-4 and CRY-O-2 was done by using 16S rRNA sequencing gene. After the analysis and alignment of sequence data of the isolates were identified as strains of Lysinibacillus boronitolerans and Shouchella rhizosphaerae. The partial 16S rDNA segment of nucleotide sequences submitted to NCBI GenBank database with accession numbers OR775574 (Lysinibacillus boronitolerans ss3cvo3), OR775575 (Lysinibacillus boronitolerans ss4cvo4) and OR708523 (Shouchella rhizosphaerae ss1cryo2). Nine nucleotide sequences involved in this analysis. Phylogenetic tree showing the relation among all three studied bacterial isolates and their closely related bacteria from NCBI database has been shown in Figure 7.

Figure 7. Neighbor-joining phylogenetic tree showing the relationship between all three studied bacterial isolates and their closely related genus from the NCBI database
Plant pathogens cause severe diseases in crop plants and result in decreased crop yield and fruit quality. Traditionally, the pathogens were mainly controlled through chemical agents but this approach was not ecofriendly. Potato and rice are major food items used throughout the world. One of the major pathogens of these crops is R. solani, which is a soil-borne pathogenic fungi which is responsible for causing sheath blight in O. sativa,2 black scurf and stem canker in potato,3 seedling disease in B. napus,4-7 rot diseases in stem and roots of dicotyledons belonging to Brassicaceae (10;8) and ‘root rot’ or ‘bare patch’ on monocotyledons of family Poaceae.11 It is also one of the most common pathogens present in agriculture fields where T. aestivum is grown as rotation crop with oil seed rape.12 Thus, R. solani can cause a significant loss in productivity in many of the agriculturally important crops.11 The bacterial isolates CRY-O-2, CVO-4 and CVO-3 exhibited potent antifungal potential against R. solani in laboratory experiments. The plant growth promoting activities such as HCN production, Ammonia production, solubilization of Phosphate and Siderophore production of isolates will make them a promising candidate for sustainable agriculture. These isolates will not only suppress the growth of R. solani by their antifungal activity but they also increase the nutrient availability for the crops. Siderophore production activity of these isolates is also important in reducing the iron toxicity in crop fields.36,37 Besides these benefits, siderophore production also promote the plant growth by means of antibiosis.38,39
Positive result was obtained for Gibberellic acid production ranging from 23 µg/ml to 38 µg/ml by all three selected the isolates and similar result for Gibberellic acid production was obtained by Gunja et al.,40 where Gibberellic acid production was found ranging from 10.2-112.4 µg/ml. In the present investigation Carrot plant was used for the isolation of rhizospheric microbes because of its medicinal properties41 and production of β-carotene in high amount42 so present investigation could be” before “an approach to use the rhizospheric bacteria from one crop to control the activities of phytopathogens in the rhizosphere of another crop, as it naturally happens during crop rotation which is oldest known biofertilizer concept in agriculture. The present investigation is an approach to use the rhizospheric bacteria from one crop to control the activities of phytopathogens in the rhizosphere of another crop, as it naturally happens during crop rotation which is oldest known biofertilizer concept in agriculture. The results obtained in the plate trials for antifungal activities by cross streak method suggests for a better future of bacterial isolates CRY-O-2, CVO-4 and CVO-3 as potent biofungicides against R. solani in cropping fields as a biocontrol agent for variety of crops.
Use of the plant growth promoting microbes for crop plants is an attractive and economic approach for sustainable agriculture. The potential benefit of these growth promoting microbes rather than using classical approaches such as chemical fertilizers, is their ecofriendly nature and cost-effective approach. It has also been established that rhizospheric growth promoting microbes are effective candidates for enriching the crop productivity and soil fertility. The present study reveals that Lysinibacillus boronitolerans ss3cvo3, Lysinibacillus boronitolerans ss4cvo4 and Shouchella rhizosphaerae ss1cryo2 are prominent candidates for applicating as a biocontrol agent against pathogens of plants such as R. solani. They inhibit the growth of fungal pathogen through production of hydrolytic enzymes, production of siderophore, IAA production, solubilization of phosphate, solubilization of potassium and production of cellulase. Thus, producing their effect by direct and indirect mechanisms for growth promotion of agricultural crops. The antifungal activity of these microbes elaborates their biological control spectrum and because of these properties selected microbes would be useful for controlling various plant pathogens as alternative of chemical fungicides under field conditions.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Shree Prakash Tiwari, Associate Professor, Department of Microbiology, VBSPU Jaunpur and Dr. Alok Kumar Srivastava, Director, NBAIM, Mau for providing necessary lab facility and research fungus (R. solani), respectively.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Verma PR. Biology and control of Rhizoctonia solani on rapeseed: a review. Phytoprotection. 1996;77(3):99-111.
Crossref - Ou S. Rice diseases, 2nd Ed. Kew, Surrey, UK: Commonwealth Mycological Institute Pub., 1985.
- Tsror L. Biology, Epidemiology and Management of Rhizoctonia solani on Potato. J Phytopath. 2010;158(10):649-658.
Crossref - Tahvonen RH, Hollo J, Hannukkala A. Rhizoctonia solani damping-off on spring turnip rape and spring rape (Brassica spp.). Finland J Agric Sci. 1984;56(2):143-154.
Crossref - Khangura RK, Barbetti MJ, Sweetingham MW. Characterization and pathogenicity of Rhizoctonia species on canola. Plant Dis. 1999;83(8):714-721.
Crossref - Kataria H, Verma P. Rhizoctonia solani damping-off and root rot in oilseed rape and canola. Crop Prot. 1992;11(1):8-13.
Crossref - Senapati M, Tiwari A, Sharma N, et al. Rhizoctonia solani K hn Pathophysiology: Status and Prospects of Sheath Blight Disease Management in Rice. Front Plant Sci. 2022;13:881116.
Crossref - Gugel RK, Yitbarek SM, Verma PR, Morrall RAA, Sadasivaiah RS. Etiology of the Rhizoctonia root-rot complex of canola in the Peace River region of Alberta. Can J Plant Pathol Rev Can De Phytopathol.1987;9(2):119-128.
Crossref - Baruch Sneh, Lee Burpee, Akira Ogoshi., Indentification of Rhizoctonia Species 2nd Ed. APS Press. 1991.
- Tewoldemedhin YT, Lamprecht SC, Mcleod A, Mazzola M. Characterization of Rhizoctonia spp. recovered from crop plants used in rotational cropping systems in the Western Cape province of South Africa. Plant Dis. 2006;90(11):1399-1406.
Crossref - Akber MA, Fang X. Research Progress on Diseases Caused by the Soil-Borne Fungal Pathogen Rhizoctonia solani in Alfalfa. Agronomy. 2024;14(7).
Crossref - Brown MJ, Woodhall J, Mooney SJ, Ray RV. The occurrence and population dynamics of Rhizoctonia solani in soil of winter wheat. Proc. Crop Prot. Nor. Brit. 2014:107-112.
- Kloepper JS, Schroth MN, Miller TD. Effect of rhizosphere colonization by plant growth promoting rhizobacteria on potato plant development and yield. Phytopathology. 1980;70(11):1078-1082.
Crossref - Bullock D. Crop rotation. Plant Sci. Crit Rev. 1992;11(4):309-326.
Crossref - Smith RG, Gross KL, Robertson GP. Effects of crop diversity on agroecosystem function: crop yield response. Ecosystems. 2008;11(3):355-366.
Crossref - Kennedy A. Bacterial diversity in agroecosystems. Agric Ecosyst Environ. 1999;74:65-76.
Crossref - Kennedy A, Smith K. Soil microbial diversity and the sustainability of agricultural soils. Plant Soil. 1995;170:75-86.
Crossref - Parkinson D, Coleman DC. Microbial communities, activity and biomass. Agric Ecosyst Environ. 1991;34(1-4):3-33.
Crossref - Velho-Pereira S, Kamat NM. Antimicrobial Screening of Actinobacteria using a Modified Cross-Streak Method. Indian J Pharm Sci, 2011;73(2):223-228.
Crossref - Jamali H, Sharma A, Roohi, Srivastava AK. Biocontrol potential of Bacillus subtilis RH5 against sheath blight of rice caused by Rhizoctonia solani. J Basic Microbiol. 2019;60(3):268-280.
Crossref - Breed RS, Morray EGD, Smith NR, [ed.], Bergey’s Manual of Determinative Bacteriology, 7th Ed. Baltimore, United State of Amarica: The Williams & Wilkins Company. 1957.
Crossref - Cappuccino J, Sherman N. Microbiology: A Laboratory Manual, 3rd Ed. New York, USA: Benjamin/Cumming Pub. Co., 1992.
- Bakker A, Schippers B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas sp. Mediated plant growth stimulation. Soil Biol Biochem. 1987;19(4):451-457.
Crossref - Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987;160(1):47-56.
Crossref - Louden BH, Haarmann D, Lynne AM. Use of Blue Agar CAS Assay for Siderophore Detection. J Microbiol Biol Educ. 2011;12(1):51-53.
Crossref - Pikovskaya RI. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Mikrobiology. 1948;17:362-370.
- Sugumaran P, Janarthanam B. Solubilization of potassium containing minerals by bacteria and their effect on plant growth. World J Agric Sci. 2007;3(3):350-355.
- Apun K, Jong BC, Salleh MA. Screening and isolation of a cellulolytic and amylolytic Bacillus from sago pith waste. J Gen Appl Microbiol. 2000;46(5):263-267.
Crossref - Brick JM, Bostock RM, Silverstone SE. Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl Environ Microbiol. 1991;57(2):535-538.
- Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with Foiin phenol reagent. J Biol Chem. 1951;193(1):265-275.
Crossref - Hunter and William J. Indole-3-acetic acid production by bacteroids from soybean root nodules. Physiol Plant. 1989;76(1):31-36.
Crossref - Ashry NM, Alaidaroos BA, Mohamed SA, Badr OA, El-Saadony MT, Esmael A. Utilization of drought-tolerant bacterial strains isolated from harsh soils as a plant growth promoting rhizobacteria (PGPR). Saud J Biol Sci. 2022;29(3):1760-1769.
Crossref - Nei M, Saitou N. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406-425.
Crossref - Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci U S A. 2004;101(30):11030-11035.
Crossref - Tamura K, Stecher G, Kumar S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol. 2021;38(7):3022-3027.
Crossref - O’Brien S, Hodgson DJ, Buckling A. Social evolution of toxic metal bioremediation in Pseudomonas aeruginosa. Proc Biol Sci. 2014;281(1787):20140858.
Crossref - Becker M, Asch F. Iron toxicity in rice-conditions and management concepts. J Plant Nutr Soil Sci. 2005;168(4):558-673.
Crossref - Timofeeva AM, Galyamova MR, Sedykh SE. Bacterial Siderophores: Classification, Biosynthesis,Perspectives of Use in Agriculture. Plants, 2022;11(22):01-22.
Crossref - Parmar HY, Chakraborty H. Effect of Siderophore on Plant Growth Promotion. Int. Jour. Appl. Pur. Sci. Agri. (IJAPSA), 2016;2:60-68.
- Vasant G, Bhatt S, Raghav R. Isolation and Molecular Characterization of Plant Growth Promoting Rhizobacteria from Groundnut (Arachis Hypogaea L.) Rhizosphere. Curr. Agri. Res., 2023; 11(1):337-347.
Crossref - Varshney K, Mishra K, An analysis of health benefits of carrot. Int. J. Innov. Res. Eng. Manage. 2022; 9, 211-214.
Crossref - Karnjanawipagul P, Nittayanuntawech W, Rojsanga P, Suntornsuk L. Analysis of β-carotene in carrot by spectrophotometry. Mahidol Univ. J. Pharm. Sci. 2010, 37;8-16
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.