


ISSN: 0973-7510
E-ISSN: 2581-690X
The larvae of the black soldier fly (BSFL), Hermetia illucens (Linnaeus, 1758) (Diptera: Stratiomyidae), can survive in environments contaminated with various bacteria by producing antimicrobial compounds. This study, for the very first time, investigated the potential antibacterial activity of hemolymph extracted from BSFL in Malaysia using diffusion and dilution methods. Prior to extraction, the larvae were infected with either Methicillin-resistant Staphylococcus aureus (MRSA) or Escherichia coli. Then, the hemolymph was collected. Serial dilutions from 200 to 12.5 mg/ml of the hemolymph extracts were screened against ten different bacteria. The results showed inhibition of eight out of ten tested bacteria (i.e., MRSA, Staphylococcus aureus, Streptococcus pyogenes, Bacillus subtilis, Micrococcus luteus, Escherichia coli, Klebsiella pneumoniae and Acinetobacter sp.). We found that immunological-challenge larvae have stronger antimicrobial activity than the control groups. The Minimum Inhibitory Concentrations (MIC) for bacteria against for infected larvae were 12.5 mg/ml for MRSA, S. pyogenes, B. subtilis, M. luteus, E. coli, and K. pneumoniae. As for bactericidal activity, the MBC of E. coli infected larvae was 25mg/ml against S. pyogenes and B. subtilis. In conclusion, BSFL hemolymph has antibacterial activity against a range of bacteria and could be a candidate for novel antimicrobial development.
Antibacterial Agents, BSFL, E. coli, Hemolymph Extract, Hermetia illucens, MRSA
Antimicrobial resistance (AMR) is defined as a loss of microorganism response to the antimicrobial agents including antibiotics. It is associated with several health care problems such as increase in mortality rate, prolonged hospitalization, and increase in treatment costs.1 Also, it was predicted that bacterial AMR for 23 pathogens with 88 pathogen-drug combinations had led to 4.95 billion fatalities reported from 204 countries.2 Antimicrobial resistance can develop naturally through genetic transmission over time; however, misuse and overuse of antibiotics in healthcare and agriculture sectors have accelerated this process.3 Bacteria have several mechanisms of developing resistance to antimicrobial agents, including limiting uptake of a drug, altering a drug target, inactivating a drug, active efflux of a drug, as well as bypassing of metabolic pathways inactivated by antibiotics.4 To date, three strategies to overcome antimicrobial resistance have been highlighted: (1) discovering new antibiotics, (2) improving the efficacy of existing antibiotics, and (3) developing alternatives to traditional antibiotics.5
Innate immunity is the first line of defense in both mammals and insects. It is generally activated by a series of signaling cascades responsible for triggering specific immune responses.6 Specifically in insects, the defense mechanisms triggered several immune responses, including production of cytotoxic chemicals, phagocytosis, and release of antimicrobial compounds from fat bodies into the haemolymph.7,8 Several studies have isolated and characterized antimicrobial peptides from different insects such as cockroaches,9 flies,10 and moths.11 Hence, the antimicrobial compounds isolated from insects have the potential as new antimicrobial agents.
Black soldier flies are known as effective waste recyclers, and their larvae could be utilized as animal feed, fertilizer, and natural biodiesel.12-14 The ability of black soldier fly larvae to survive and thrive in bacteria-rich environments has indicated their superior immune system that protects them from pathogenic microorganisms. To date, there have been many studies reporting the discovery of antimicrobial compounds such as cecropin-like peptides from black soldier fly larvae,15 that have the potential as sources of new antibacterial agents. It is well known that the production of different compounds by plants and animals depends on environmental factors such as rainfall, temperature, water, and seasons which can affect their biological activity.16 Hence, we hypothesized that insect production of antimicrobial compounds may also depend on environmental factors and may vary geographically. Higher temperatures and humidity in tropical countries such as Malaysia encourage the proliferation of a variety of microbes. However, no previous studies have attempted to target the antimicrobial activity of BSFL under tropical climate against main pathogenic bacteria. Therefore, the aim of this research was to investigate the antimicrobial activity of BSFL in Malaysia as the potential antimicrobial agents.
Rearing of the black soldier fly larvae
The 5-day-old larvae of H. illucens were purchased from Life Origin Sdn. Bhd, Malaysia. They were reared at 30°C, 60% relative humidity, and a photoperiod of 14:10 light and dark in an incubator (Memmert, Germany). The larvae were fed with a mixture of organic wastes such as rotten fruits and vegetables added with 200g chicken feed until they reached the sixth in star stage (~13 days).17 Then, they were divided into three groups, each group consisting of about 200-250 larvae (~300 g). Each experiment performed was conducted in triplicate.
Preparation of larvae hemolymph extract
The first and second groups of black soldier fly larvae were pricked deeply at the posterior segment of the larvae with 1 cc syringe containing 0.5-McFarland bacterial suspension of Methicillin-resistant Staphylococcus aureus (MRSA) and E. coli, respectively, to stimulate the immunity and production of antibacterial peptides, while the third group was used as the control. All three groups were then incubated overnight at the same rearing temperature, humidity, and feeding material. After eight hours incubation, the larvae were cut with small scissors, and the hemolymph was collected in sterile 1.5-microcentrifuge tubes, with a 0.001g of N-phenylthiourea powder added to prevent melanization. The collection was performed on ice. The hemolymph crude extract was then centrifuged and filtered with a cellulose acetate membrane before being dried in a vacuum concentrator, Concentrator Plus (Eppendorf, Germany) until a solid residue remained. For the diffusion tests, the original weight of the hemolymph dry extract used was determined at 200 mg using a beam balance FX-300i (A&D Company Limited, Japan). The samples were diluted by adding 1 ml of autoclaved distilled water. The final concentration of the extract was 200 mg/ml. The product was stored at -20°C until further use. To determine the minimum inhibitory concentration (MIC), the hemolymph extract was further diluted to a serial concentration of 100 mg/ml, 50 mg/ml, 25 mg/ml, and 12.5 mg/ml.
Diffusion assays
In this study, ten different gram positive and gram negative bacteria, the common causative agent of skin, soft tissue and intestinal infections, were selected for the screening: MRSA (clinical isolate), S. aureus (ATCC 6538), Streptococcus pyogenes (ATCC 21547), Staphylococcus epidermidis (ATCC 12228), Micrococcus luteus (ATCC 10240), Bacillus subtilis (ATCC 6633), E. coli (ATCC 25922), Acinetobacter sp. (clinical isolate), Klebsiella pneumoniae (ATCC 13883), and Pseudomonas aeruginosa (ATCC 15442). The clinical isolates were provided by the Microbiology and Parasitology Unit, Clinical Diagnostic Laboratories, Hospital Al-Sultan Abdullah, Universiti Teknologi MARA (UiTM, Malaysia). ATCC bacteria (Remel, United Kingdom) were purchased from BIO3 Scientific SDN BHD. Bacterial suspensions were prepared at a concentration of 0.25 MacFarland in tryptic soy broth (TSB) and grown in Muller-Hinton agar medium (MH). Two sets of experiments were performed using the disk and well methods. The hemolymph extract with 200mg/ml were pipetted into each well and disk prepared. Common antibiotics such as vancomycin (VA, 30µg/disk), penicillin G (P, 5 µg/disk), gentamicin (CN, 10 µg/disk), polymyxin B (PB, 300µg/disk) and ciprofloxacin (CIP, 5 µg/disk) (Oxoid, UK) were used as positive controls. After 12-hours incubation, the inhibition zones of bacteria growth were recorded in all experimental and control groups.
Minimum inhibitory concentration (MIC)
For the MIC assay, the same bacteria used in the previous section were prepared at 0.25 MacFarland and added to a 96-well plate. The hemolymph extract at serial concentrations mentioned above was added to the bacteria at a 1:1 ratio. One group of wells (bacteria only) served as a negative control and another group of wells (broth only) served as a sterile reference. MIC concentration was measured before and after overnight incubation at 37 °C at wavelength of 595 nm using a microplate reader. The absorbance values were then color-corrected using the modified formula.18
Adjusted OD = OD post-incubation – [ OD pre-incubation + OD Blank / 2 ]
Adjusted OD refers to the optical density value after being color-corrected, OD post-incubation refers to the values read from spectrophotometer after 8-hours incubation while pre-incubation refers to the readings of the same well prior to incubation. OD Blank refers to hemolymphs without the addition of bacterial suspension.
Minimum bactericidal concentration (MBC)
From the microplate used for MIC, 10µl of each concentration was added to MH agar for growth monitoring. They were incubated overnight at 37°C. The concentration that showed no bacterial growth was recorded as MBC.
Statistical analysis
Statistical analysis was performed using Prism 8 (GraphPad, USA). One-way analysis of variance (ANOVA) was used to compare between different prior bacterial infections of the larvae and the inhibition zones of produced against the bacteria tested. Then, Tukey’s multiple comparison tests were used to compare between groups, p value < 0.05 was considered statistically significant.
Determination of bacterial susceptibility based on diffusion assays
The antimicrobial activity of hemolymph extracts from non-infected larvae and infected larvae with MRSA and E. coli were evaluated using two diffusion tests-well and disk methods (Table 1, Figure). In general, crude extract of hemolymph from infected BSF larvae showed inhibitory effect against eight bacteria using the disk diffusion method (MRSA, S. aureus, S. pyogenes, M. luteus, B. subtilis, E. coli, Acinetobacter sp. and K. pneumoniae) and when using the well diffusion technique, showed inhibitory effects against seven bacteria such as MRSA, S. aureus, M. luteus, B. subtilis, E. coli, Acinetobacter sp. and K. pneumoniae. In addition, the hemolymph of the MRSA-infected larvae showed significantly higher antimicrobial activity against bacteria such as B. subtilis (p = 0.0001), M. luteus (p = 0.0049) and E. coli (p = 0.0377) than the non-infected larvae (Table 2). Meanwhile for E. coli-infected larvae, they showed significant increase compared to the non-infected larvae against bacteria such as MRSA (p = 0.0149), M. luteus (p = 0.0039) and E. coli (p = 0.0377) (Table 2). There were no significant differences of antimicrobial activity of hemolymph extracts obtained from the larvae infected with MRSA and E. coli against bacteria such as MRSA (p = 0.8790), S. aureus (p = 0.4435), S. pyogenes (p = 0.5015), B. subtilis (p = 0.1024), M. luteus (p = 0.3131), E. coli (p = 0.6452), K. pneumoniae (p =0.3779), and Acinetobacter sp. (p = 0.3131).
Table (1):
Susceptibility of 10 species of pathogenic bacteria to hemolymph extracts from black soldier fly larvae (BSFL) using both well and disk diffusion techniques. Antibiotics used as positive control were indicated as the following: Vancomycin 30 µg/disk =VA-30, penicillin G 5 µg/disk =P-5, Gentamicin 10 µg/disk =CN-10, polymyxin B 300 µg/disk =PB-300, and ciprofloxacin 5 µg/disk= CIP-5
| Bacteria tested | Mean inhibition zones (mm) of the extract from BSFL± Standard error (Mean±SE) | p value | F value | Positive Control | ||
|---|---|---|---|---|---|---|
| Non-infected | Infected with MRSA | Infected with E. coli | ||||
| MRSA Well diffusion Disk diffusion |
6.0±0 6.7±0.26 |
9.3±0.29 9.7±0.47 |
8.0±0.35 10.3±0.27 |
0.18 (NS)) | 2.74 | VA30 26.3±0.17 |
| S. aureus Well diffusion Disk diffusion |
6.0±0 6.3±0.26 |
10.3±0.42. 9.0±0.19 |
9.7±0.45 7.6±0.48 |
0.44 (NS) | 1.00 | VA-30 20.3±0.20 |
| S. epidermidis Well diffusion Disk diffusion |
6.0±0 6.0± |
6.006.0±0 | 6.0±06.0±0 | – | – | VA30 16.7±0.33 |
| S. pyogenes Well diffusion Disk diffusion |
6.0±07.3±0.12 | 6.0±0 7.3±0.12 |
6.0±07.0±0.12 | 0.05* | 7.00 | P-515.7±0.1 |
| M. luteus Well diffusion Disk diffusion |
8.7±0.11 7.3±0.12 |
24.3±0.07 20.3±0.07 |
23.0±0.24 19.7+0.08 |
0.79 (NS) | 0.25 | VA-3029.0±0.11 |
| B. subtilis Well diffusion Disk diffusion |
6.0±06.3±0.13 | 7.0±0.128.3±0.12 | 6.7±0.227.0±0.22 | 0.02* | 13.00 | VA-3017.7±0.45 |
| E. coli Well diffusion Disk diffusion |
6.0±06.0±0 | 6.3±0.12 7.7±0.12 |
7.0±0.127.7±0.12 | 0.11 (NS) | 4.00 | CN-1021.7±0.38 |
| Acinetobacter Well diffusion Disk diffusion |
6.0±0 6.0±0 |
6.3±0.13 6.0±0 |
6.0±0 6.7±0.26 |
0.17 (NS) | 2.80 | PB-30019.0±0.26 |
| K. pneumoniae Well diffusion Disk diffusion |
6.0±0 6.0±0 |
7.7±0.32 8.0±0.12 |
7.7±0.20 7.0±0.31 |
0.16 (NS) | 3.00 | CN-1021.7±0.26 |
| P. aeruginosa Well diffusion Disk diffusion |
6.0±06.0±0 | 6.0±0 6.0±0 |
6.0±06.0±0 | – | – | CIP-528.0±0.13 |
BSFL = black soldier fly larvae, MRSA = Methicillin-resistant Staphylococcus aureus, p value: (*<0.05); (**<0.005); (***<0.0005), (****<0.0001). NS refers Not Significant.
Table (2):
Tukey’s Comparison Test between experimental and control groups of black soldiers fly larval hemolymph extracts against seven species of pathogenic bacteria
| Bacteria | Non-infected BSFL versus MRSA-infected BSFL | Non-infected BSFL versus E. coli infected BSFL | E. coli infected BSFL versus MRSA-infected BSFL |
|---|---|---|---|
| p value | |||
| MRSA | 0.3209 | 0.0149* | 0.8790 |
| S. aureus | 0.0634 | 0.2259 | 0.4435 |
| S. pyogenes | 0.5015 | >0.9999 | 0.5015 |
| B. subtilis | <0.0001**** | 0.3131 | 0.1024 |
| M. luteus | 0.0049** | 0.0039** | 0.3131 |
| E. coli | 0.0377* | 0.0377* | 0.6452 |
| K. pneumoniae | 0.3779 | 0.3779 | 0.3779 |
| Acinetobacter sp. | 0.3779 | 0.6452 | 0.3131 |
p value: (*<0.05); (**<0.005); (***<0.0005), (****<0.0001). NS refers Not Significant.
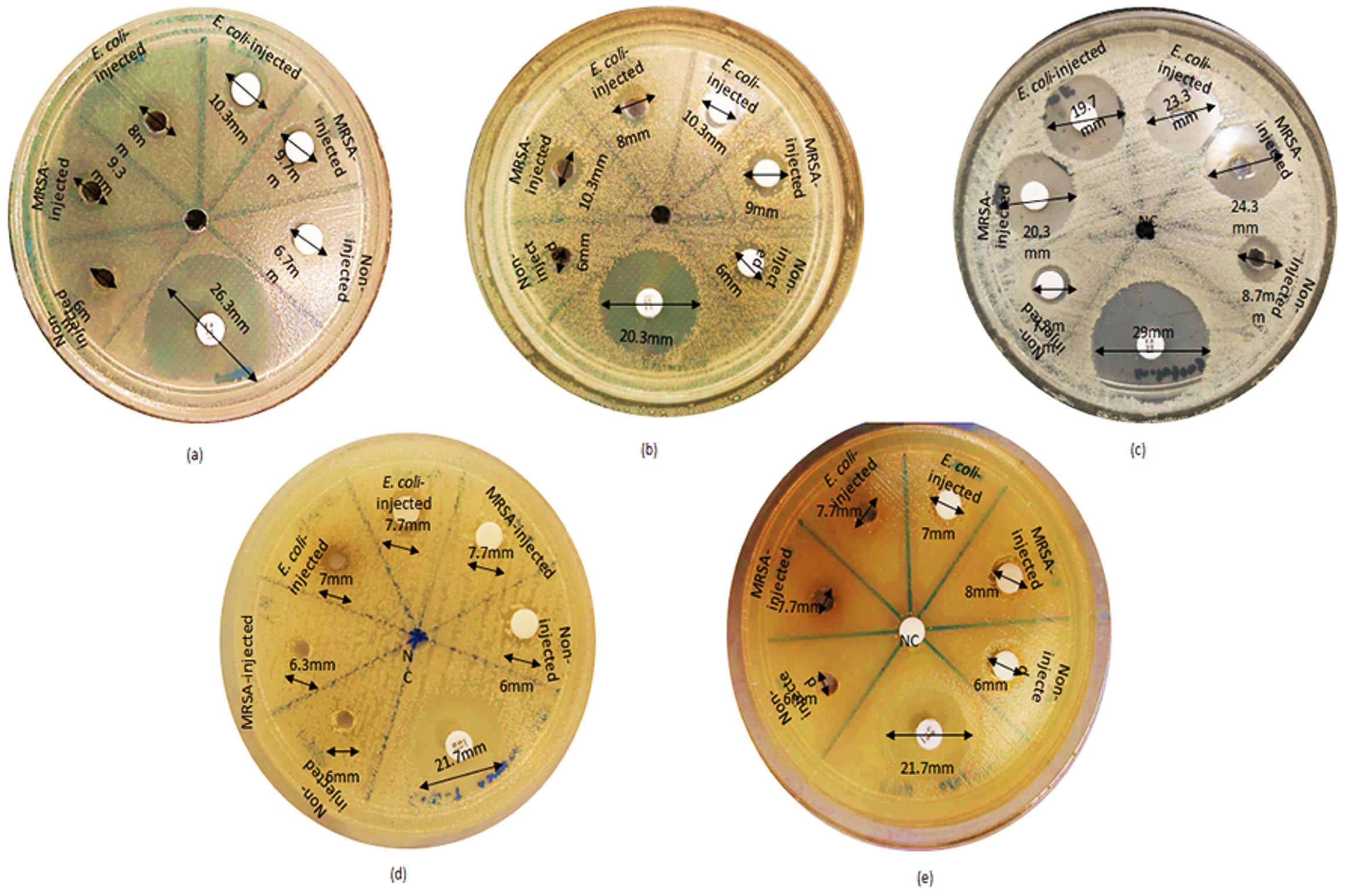
Figure. Inhibitory effects of black soldier fly larval hemolymph extracts against five species of pathogenic bacteria: (a) MRSA; (b) S. aureus; (c) M. luteus; (d) E. coli; (e) K. pneumoniae
Minimum inhibitory concentration (MIC)
The MIC assay was performed with serial dilutions of 200 mg/ml, 100 mg/ml, 50 mg/ml, 25 mg/ml, and 12.5 mg/ml of hemolymph crude extracts from the non-infected and infected larvae against eight pathogenic bacteria selected based of bacteria susceptibility from well and disk diffusion tests(Table 3). The MIC for MRSA was 12.5, 50 and 100 mg/ml for hemolymph extracts from E. coli-infected, MRSA-infected, and non-infected larvae, respectively. For S. aureus, the MIC was 50 mg/ml for the extract from larvae infected with MRSA and E. coli, and 25 mg/ml for non-infected larvae. Meanwhile for S. pyogenes, the MICs were 50 mg/ml for the extracts from non-infected and MRSA-infected larvae, while the MIC for the extract from E. coli-infected larvae was 12.5 mg/ml. As for B. subtilis, the MIC was 100 mg/ml for non-infected, and 12.5 mg/ml for MRSA and E. coli infected larvae. The non-infected larvae were unable to inhibit E. coli, Acinetobacter sp., K. pneumoniae. Meanwhile, for infected larvae, the MICs for E. coli and K. pneumoniae were 12.5 mg/ml for both. As for Acinetobacter sp., the MIC was 100 mg/ml for E. coli-infected larvae.
Table (3):
The minimum inhibitory concentration (MIC) of hemolymph crude extract from non-infected, E. coli- infected and MRSA-infected black soldier fly larvae (BSFL) against eight species of pathogenic bacteria
| Minimum Inhibitory Concentration (MIC) (mg/ml) | |||
|---|---|---|---|
| Bacteria | Non- infected | Infected with MRSA | Infected with E. coli |
| MRSA | 100 | 50 | 12.5 |
| S. aureus | 25 | 50 | 50 |
| S. pyogenes | 50 | 50 | 12.5 |
| B. subtilis | 12.5 | 25 | 12.5 |
| M. luteus | 100 | 12.5 | 12.5 |
| E. coli | NI | 12.5 | 12.5 |
| K. pneumoniae | NI | 12.5 | 12.5 |
| Acinetobacter sp. | NI | NI | 100 |
NI refers to No Inhibition
Minimum bactericidal concentration (MBC)
The MBC test was performed using the same range of concentrations of hemolymph extract as those of the MIC plate (Table 4). Most of the tested bacteria demonstrated growth, except for B. subtilis and S. pyogenes, which showed no growth at the presence of the hemolymph extract from E. coli-infected larvae. The MBC for both B. subtilis and S. pyogenes was 25 mg/ml.
Table (4):
The minimum bactericidal concentration (MBC) of hemolymph extract of black soldier fly larvae (BSFL) against eight species of pathogenic bacteria
| Bacteria | Minimum bactericidal concentration (mg/ml) | |||||
|---|---|---|---|---|---|---|
| 200 | 100 | 50 | 25 | 12.5 | ||
| MRSA | + | + | + | + | + | |
| BSF Non infected larvae | S. aureus | + | + | + | + | + |
| S. pyogenes | + | + | + | + | + | |
| B. subtilis | + | + | + | + | + | |
| M. luteus | + | + | + | + | + | |
| E. coli | + | + | + | + | + | |
| K. pneumoniae | + | + | + | + | + | |
| Acinetobacter sp. | + | + | + | + | + | |
| MRSA | + | + | + | + | + | |
| S. aureus | + | + | + | + | + | |
| BSF MRSA infected larvae | S. pyogenes | + | + | + | + | + |
| B. subtilis | + | + | + | + | + | |
| M. luteus | + | + | + | + | + | |
| E. coli | + | + | + | + | + | |
| K. pneumoniae | + | + | + | + | + | |
| Acinetobacter sp. | + | + | + | + | + | |
| MRSA | + | + | + | + | + | |
| S. aureus | + | + | + | + | + | |
| S. pyogenes | – | – | – | – | + | |
| BSF E. coli infected larvae | B. subtilis | – | – | – | – | + |
| M. luteus | + | + | + | + | + | |
| E. coli | + | + | + | + | + | |
| K. pneumoniae | + | + | + | + | + | |
| Acinetobacter sp. | + | + | + | + | + | |
(+) indicates growth of bacteria; (-) indicates no growth of bacteria.
Hemolymph is a circulating fluid that flows through the body of insects and transports various nutrients. It is composed of various complex materials such as nutrients, water, inorganic and organic solutes such as amines, carbohydrates, peptides, and proteins. Previous studies showed that the hemolymph of BSF contains antimicrobial peptides (AMPs) responsible for antimicrobial activity.15 In insects, these AMPs are generally stored in fat bodies and activated to be released upon infection.19 In this study, we provided screening of antibacterial effects of crude extracts from hemolymph of Malaysian strain BSF using disk and well diffusion methods. We found that the antimicrobial activities of hemolymph extract were more prominent when the hemolymph extract was used in the disk diffusion test than in the well diffusion method. This could be due to the hemolymph extract being liquid and therefore dries rapidly in the agar well than on the plates. Other studies also showed that the disk diffusion method was more useful than the well diffusion assay for detecting the antimicrobial activity of various animal and plant extracts.20,21
It was found that the BSFL infected with E. coli and MRSA before extraction showed higher antimicrobial activities compared to the non-infected larvae. Generally, AMPs are expressed in insect hemolymph at minimal amount due to several factors such as metabolic changes, aging, and stressors in non-infected larvae.22 However, when invaded with bacteria, a series of signaling cascades were activated by immune pathways such as Toll and IMD (immunodeficiency).23 As a result, there were increased expression of inflammatory factors and AMPs following the immune responses.24 These resulted in stronger antimicrobial responses of the infected larvae compared to the non-infected ones as portrayed in our study.
Hemolymph extracted from BSF larvae immunized by E. coli showed promising bactericidal properties at low concentration compared to MRSA-infected larvae. Previous studies also showed an increase in bacteriolytic activities in hemolymph upon infection with E. coli -LPS (lipopolysaccharide), which was attributed to increased lysozyme and defense protein production in insects.25,26 When infected with various bacteria, gram negative bacteria triggered the IMD pathways while gram positive bacteria activated the Toll pathways.27 Extracellular recognition factors started protease cascades that activated the Toll receptor ligand Spatzle in order to activate the Toll pathway in Drosophila while IMD pathways control the activity of a third NF-kB protein in Drosophila called Relish and regulated the production of the majority of AMPs.28,29 Different transcripts and AMP levels were produced because of variations in activation methods. In contrast to gradual and prolonged induction of the genes by Toll pathways, it was found that IMD pathways were more engaged in the robust, accurate, and higher expression of AMP genes of flour beetle, Tribolium castaneum.30 Therefore, we deduced that E. coli-infected BSF larvae might also share similar immune responses, hence resulting in stronger antimicrobial activities compared to MRSA-infected larvae. However, to fully understand the immune mechanisms involved in post-bacteria infected larvae, further experiments on immune pathways of BSF larvae are necessary.
In short, the hemolymph of Malaysia strain BSFL-infected with bacteria showed greater antimicrobial activity compared to the non-infected larvae. Between the bacteria, E. coli-infected larvae showed better antimicrobial activity against S. pyogenes and B. subtilis than MRSA. More future works are needed especially on the purification and elucidation of the antimicrobial compounds responsible for the antibacterial activity from Malaysian strain BSFL.
ACKNOWLEDGMENTS
The authors are thankful to the Institute for Medical Molecular Biotechnology (IMMB), Faculty of Medicine, Universiti Teknologi MARA (UiTM) for providing the facilities to conduct this research.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
NA collected the data and performed the experiments. HA contributed to methodology and data validation. NA and HA performed formal analysis. CCH and NS secured funding. NA wrote the manuscript. NS, AK, SS and HA reviewed the manuscript. AK, SS, CCH and HA edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This work was funded by Universiti Teknologi MARA (UiTM) under internal grants, namely FMRG: 600-TNCPI 5/3/DDF (MEDIC) (006/2021), GIP: 600-RMC/GIP 5/3 (043/2021), and 600-RMC/GIP 5/3 (038/2023).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- World Health Organization. 2021. Antimicrobial resistance. https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance. Accessed 17 November 2021.
- Murray CJ, Ikuta KS, Sharara F, et al. Global burden of bacterial antimicrobial resistance in 2019:a systematic analysis. Lancet. 2022;399(10325):629-655.
Crossref - Dadgostar P. Antimicrobial resistance:implications and costs. Infect Drug Resist. 2019;12:3903-3910.
Crossref - Reygaert WC. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018;4(3):482-501.
Crossref - Ye J, Chen X. Current promising strategies against antibiotic-resistant bacterial infections. Antibiotics. 2023;12(1):67.
Crossref - Silverman N, Maniatis T. NF-kB signaling pathways in mammalian and insect innate immunity. Genes Dev. 2001;15:2321-2342.
Crossref - Hoffmann JA. Innate immunity of insects. Curr Opin Immunol. 1995;7(1):4-10.
Crossref - Manniello MD, Moretta A, Salvia R, et al. Insect antimicrobial peptides:Potential weapons to counteract antibiotic resistance. Cell Mol Life Sci. 2021;78(9):4259-4282.
Crossref - Lee H, Hwang JS, Lee DG. Periplanetasin-4, a novel antimicrobial peptide from the cockroach, inhibits communication between mitochondria and vacuoles. Biochem J. 2019;476(8):1267-1284.
Crossref - Hanson MA, Lemaitre B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr Opin Immunol. 2020;62:22-30.
Crossref - Wojda I, Staniec B, Sulek M, Kordaczuk J. The greater wax moth Galleria mellonella:biology and use in immune studies. Pathog Dis. 2020;78(9):1-15.
Crossref - Raksasat R, Lim JW, Kiatkittipong W, et al. A review of organic waste enrichment for inducing palatability of black soldier fly larvae:Wastes to valuable resources. Enviro Pollut. 2020;267:115488.
Crossref - Tomberlin JK, Van Huis A. Black soldier fly from pest to ‘crown jewel’of the insects as feed industry:an historical perspective. J Insect Food Feed. 2020;6(1):1-4.
Crossref - Beesigamukama D, Mochoge B, Korir NK, et al. Low-cost technology for recycling agro-industrial waste into nutrient-rich organic fertilizer using black soldier fly. Waste Manage. 2021;119:183-194.
Crossref - Park SI, Yoe SM. A novel cecropin like peptide from black soldier fly, Hermetia illucens:Isolation, structural and functional characterization. Entomol Res. 2017;47(2):115-124.
Crossref - Netshiluvhi TR, Eloff JN. Influence of annual rainfall on antibacterial activity of acetone leaf extracts of selected medicinal trees. S Afr J Bot. 2016;102:197-201.
Crossref - Kim WT, Bae SW, Park HC, et al. The larval age and mouth morphology of the black soldier fly, Hermetia illucens(Diptera:Stratiomyidae). Int J Ind Entomol. 2010;21:185-187.
- Sahalan, AZ, Omar B, Mohamed AY, Jeffery J. Antibacterial activity of extracted haemolymph from larvae and pupae of local fly species, Musca domestica and Chrysomya megacephala. Jurnal Sains Kesihatan Malaysia. 2007;4(2):1-11.
- Li S, Yu X, Feng Q. Fat body biology in the last decade. Ann Rev Entomol. 2019;64:315-333.
Crossref - Boyanova L, Gergova G, Nikolov R, et al. Activity of Bulgarian propolis against 94 Helicobacter pylori strains in vitro by agar-well diffusion, agar dilution and disc diffusion methods. J Med Microbiol. 2005;54(Pt 5):481-483.
Crossref - King T, Dykes G, Kristianti R. Comparative evaluation of methods commonly used to determine antimicrobial susceptibility to plant extracts and phenolic compounds. J AOAC Int. 2008;91(6):1423-1429.
Crossref - Staczek S, Malgorzata C, Agnieszka ZB. Unraveling the role of antimicrobial peptides in insects. Int J Mol Sci. 2023;24(6):5753.
Crossref - Engstrom Y. Induction and regulation of antimicrobial peptides in Drosophila. Dev Comp Immunol. 1999;23(4-5):345-358.
Crossref - Buchon N, Silverman N, Cherry S. Immunity in Drosophila melanogaster – from microbial recognition to whole-organism physiology. Nat Rev Immunol. 2014;14(12):796-810.
Crossref - Anderson RS, Cook ML. Induction of lysozyme-like activity in the haemolymph and hemocytes of an insect, Spodoptera eridania. J Invertebr Path. 1979;33(2):197-203.
Crossref - Han YS, Chun J, Schwartz A, Nelson S, Paskewitz SM. Induction of mosquito haemolymph proteins in response to immune challenge and wounding. Dev Comp Immunol. 1999;23(7-8):553-562.
Crossref - Tanji T, Hu X, Weber AN, Ip YT. Toll and IMD pathways synergistically activate an innate immune response in Drosophila melanogaster. Mol Cell Biol. 2007;27(12):4578-4588.
Crossref - Valanne S, Wang JH, Ramet M. The Drosophila toll signaling pathway. J Immunol. 2011;186(2):649-656.
Crossref - Myllymaki H, Valanne S, Ramet M. The Drosophila IMD signaling pathway. J Immunol. 2014;192(8):3455-3462.
Crossref - Yokoi K, Koyama H, Minakuchi C, Tanaka T, Miura K. Antimicrobial peptide gene induction, involvement of Toll and IMD pathways and defense against bacteria in the red flour beetle, Tribolium castaneum. Results Immunol. 2012;2:72-82.
Crossref
© The Author(s) 2023. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.