


ISSN: 0973-7510
E-ISSN: 2581-690X
Endophytic yeasts were isolated from different organs of 18 plant species collected in Uzbekistan. More than 50 yeast isolates were obtained, of which 41 were identified using Matrix-Assisted Laser Desorption Ionization-Time of Flight (MALDI-TOF MS), while three strains were further confirmed by molecular methods based on 18S rRNA and ITS region sequencing. The identified genera included Pichia, Meyerozyma, Metschnikowia, Hanseniaspora, Kluyveromyces, and Naganishia. The antimicrobial activity of ethyl acetate extracts obtained from lyophilized whole cultures of 29 selected strains was evaluated against Gram-positive bacteria (Staphylococcus aureus, Bacillus subtilis), Gram-negative bacteria (Pseudomonas aeruginosa, Escherichia coli), and the pathogenic yeast Candida albicans using the agar well diffusion method. Several isolates exhibited pronounced and broad-spectrum antimicrobial activity, with Metschnikowia pulcherrima 02, Meyerozyma guilliermondii 03, Pichia kudriavzevii 66, and isolate 023 showing the highest inhibition zones. To elucidate the biochemical basis of the observed antimicrobial effects, the most active strains were subjected to reverse-phase Liquid Chromatography-Tandem Mass Spectrometry (Nano-LC-MS/MS) analysis. Bioactive metabolites with known antimicrobial properties, including alternariol, andrastin D, altersolanol A, fumiquinazoline F, 1,4-naphthoquinone and lanosterol were putatively identified. The presence of these metabolites suggests that the antimicrobial activity of the investigated endophytic yeasts may be associated with metabolite production. This study represents one of the first reports on the isolation and metabolite profiling of endophytic yeasts from plants in Uzbekistan and the detection of these metabolites suggests that the observed antimicrobial activity may be associated with bioactive secondary metabolites produced by the isolates.
Endophytic Yeasts, Antimicrobial Activity, Nano-LC-MS/MS, Secondary Metabolites, Uzbekistan
Endophytic microorganisms, including fungi, bacteria, and yeasts, are widely distributed as microorganisms that live in the internal tissues of plants and establish various relationships with them. Endophytic microorganisms provide various beneficial functions to plants, including improved growth, protection against pathogens, and increased resistance to plant stress. Endophytic microorganisms are characterized by their antibacterial and antifungal activities, and their metabolites are widely used in various biotechnological and medical fields.1,2 During the past few decades, endophytes have attracted much attention as an important and novel source of bioactive natural products with potential applications in medicine.3,4 In addition, yeast is also a source of several secondary metabolites with various industrial applications, such as phenols, flavonoids, stilbenoids, isoprenoids, sesquiterpenes, etc.5,6 Some studies have shown that the crude extract of yeast contains important metabolites that can act as antioxidants, anti-inflammatory agents, anti-proliferative agents, etc.7-9 Although antibiotics have been widely used in animal husbandry, it is important to address the problem of antibiotic supply, as a substitute and effective means, especially for high-quality products, particularly for the production of high-quality products.3,10
Endophytic yeasts have been found to be beneficial to various plants, in particular, species such as S. cerevisiae, Metschnikowia pulcherrima, Pichia guilliermondii can act against harmful plant pathogens. Metabolites isolated from the endophytic yeast Pichia guilliermondii Ppf9 showed strong antibacterial activity against a wide range of bacteria, which opens up the possibility of their future use as antibacterial agents.11
Metschnikowia pulcherrima is one of the endophytic yeasts widespread in grapes, and has antibacterial and antifungal activity through its pulcherrimin pigment.12 This pigment has been shown to complex with iron ions and thus play an important role in inhibiting the growth of pathogenic microorganisms.13 Another important yeast species, Wickerhamomyces anomalus, is known to produce Candida toxins, which are active against other pathogens.10,14
Endophytic microorganisms are used in biological preservation,15 to extend the shelf life of food products, sausages, cheese, yogurt, and other fermented foods,10 and the antibacterial and antifungal compounds they produce are important in the food industry and biological preservation processes.16,17
Crude extracts of yeasts, especially those obtained from plants such as Capsicum annuum L., contain many beneficial properties. Their antibacterial, antifungal, and anti-inflammatory activities constitute the full spectrum of metabolites produced by endophytes. These properties ensure their use in medicine and other industrial fields.18
Endophytic yeasts and their antimicrobial activities, especially their antibacterial, antifungal, and biopreservative abilities, open up a wide range of applications in plant microbiology, agriculture, and medicine. The antimicrobial activities of endophytic yeasts, the beneficial properties of their metabolites, and their role in biopreservatives indicate their future potential. It is also clear that further research is needed for their application in healthcare and industry.1
Despite the increasing interest in endophytic yeasts as sources of bioactive compounds, their diversity and antimicrobial potential remain poorly explored in Central Asia, particularly in Uzbekistan. In addition, studies integrating antimicrobial screening with metabolite profiling approaches such as Nano-LC-MS/MS are still limited. Therefore, the present study aimed to provide one of the first systematic surveys of endophytic yeasts from selected plant hosts in Uzbekistan, including their isolation, diversity assessment, antimicrobial screening, and putative metabolite profiling. This work provides regional baseline data rather than introducing a new class of antimicrobial agents.
Sample collection, isolation and identification of endophytic yeasts
Endophytic yeasts were isolated at the Physiologically Active Compounds Biochemistry and Biotechnology Laboratory of the Institute of Microbiology, Academy of Sciences of the Republic of Uzbekistan. A total of 50 endophytic yeast isolates were obtained from the generative and storage organs of 18 plant species, 36 samples collected from Tashkent city and the surrounding Tashkent region in 2024-2025 using an enrichment method.19 For each plant species, 3-5 individual plants were sampled, and 2 organs, the generative and storage organs were collected per plant, with 3 biological replicates per organ. The plant species were chosen because they are native to Uzbekistan and are widely distributed across the country, making them relevant and representative for this study. Generative and storage organs, specifically flowers and fruits, were selected because of their higher sugar content, making them suitable substrates for yeast growth.
Flower samples were collected in March and April from trees located in the Yunusabad district of Tashkent a few hours prior to the experiments. Fruit samples were collected in May from different regions of Uzbekistan, including strawberry (Fragaria Victoria) from Hasanboy (Tashkent region), strawberry (Korolevskiy cultivar) from Qibray district (Tashkent region), apricot from Surkhandarya province, white and black mulberry from Yangiyul district (Tashkent region), and sweet cherry from Namangan province. Stored apples harvested in the previous season were also included in the study. All plant materials were identified based on their morphological characteristics.
The samples were processed under aseptic conditions. Surface sterilization was carried out using 70% ethanol (3-5 min), followed by treatment with 3% sodium hypochlorite, and subsequently rinsed three times with sterile distilled water. The final rinse water was plated onto agar medium as a sterility control, and no microbial growth was observed, confirming effective surface sterilization. Sterilized tissue surface imprinted as control and microbial growth was not observed.
The sterilized materials were cut into fragments of approximately 5 mm. The plant fragments were then transferred into flasks containing liquid Sabouraud medium supplemented with cefotaxime (200 mg/L) to suppress bacterial growth during the initial isolation stage. After the primary enrichment step, cefotaxime was not used in any subsequent cultivation procedures. Because cefotaxime was added during the enrichment step, our method may have favored yeasts that can tolerate this antibiotic while suppressing some bacteria. This means we might have missed yeasts that depend on bacterial partners, so the isolates we recovered may not fully represent the natural diversity found in the original samples. The cultures were incubated at 28 °C, 140 rpm for 3-5 days on a rotary shaker.
After visual changes such as turbidity and gas formation were observed, the enriched cultures were streaked onto Sabouraud Dextrose Agar (SDA) plates without antibiotics to obtain pure cultures. Yeast-like colonies were microscopically examined and repeatedly sub-cultured on SDA to ensure purity.
The obtained isolates were preliminarily identified according to the methods of Abdel-Hafez et al.19 and Kurtzman et al.20 For the separation of the culture liquid (supernatant), the yeast cultures were centrifuged at 6000 rpm for 15 minutes. The supernatant was collected for further analyses.
Identification of yeast isolates
Yeast isolates were preliminarily identified based on morphological and cultural characteristics. For this purpose, the shape, color, size, and texture of 3-5 day-old colonies grown on Sabouraud Dextrose Agar were examined, along with the microscopic morphology of cells.
For more accurate identification, isolates were subcultured twice under the original isolation conditions. MALDI-TOF MS analysis was performed on the third generation of pure cultures using a 4800 Plus MALDI TOF/TOF™ Analyzer (AB SCIEX, Framingham, MA, USA), essentially as described previously.21
In brief, a small amount of fresh biomass from 48-72 hour single colonies grown on Sabouraud medium was suspended in 70% ethanol, vortexed, and centrifuged. The pellet was subjected to formic acid/acetonitrile protein extraction according to the manufacturer’s recommendations. One microliter of the extract was spotted onto a polished steel target plate, air-dried, and overlaid with 1 µL of α-cyano-4-hydroxycinnamic acid (CHCA) matrix solution (10 mg/mL in 50% acetonitrile containing 0.1% trifluoroacetic acid).
Mass spectra were acquired in positive linear mode. Data Explorer 4.0 software (AB SCIEX) was used to convert the mass spectra into .txt files, which were subsequently imported into a BioNumerics 5.1 database (Applied Maths, Sint-Martens-Latem, Belgium). Spectral profiles were compared using the Pearson product moment correlation coefficient, and a dendrogram was constructed using the unweighted pair group method with arithmetic mean (UPGMA) clustering algorithm. Homogeneous clusters consisting of isolates with visually identical or virtually identical mass spectra were delineated and assigned to genus and, where possible, species level.
Molecular identification by ITS/18S rRNA gene sequencing
Molecular identification of yeast isolates was performed by sequencing the 18S rRNA gene and the ITS1-ITS2 regions. Yeast cultures were first grown in liquid medium for 48-72 hrs, after which cells were harvested for genomic DNA extraction using a standard extraction protocol.
Amplification of the 18S rRNA and ITS regions was carried out by polymerase chain reaction (PCR) using universal fungal primers targeting conserved flanking regions. The presence and size of PCR products were confirmed by agarose gel electrophoresis.
Purified amplicons were subjected to Sanger sequencing to obtain high-quality nucleotide sequences. Although next-generation sequencing platforms such as Illumina or PacBio offer higher throughput, Sanger sequencing remains the gold standard for routine fungal identification due to its accuracy and cost-effectiveness in single-isolate analysis.
The obtained sequences were analyzed using BLAST searches against the NCBI GenBank database for taxonomic identification. Phylogenetic relationships were inferred using multiple sequence alignment and dendrogram construction based on reference sequences retrieved from public databases.22
Extraction of crude secondary metabolites
0.5 g of lyophilized whole-culture was ground using a Potter homogenizer. The resulting homogenate was transferred into a 100 mL flask, and 20 mL of ethyl acetate was added. The mixture was then placed on a shaker at room temperature with a rotation speed of 160 rpm. After 24 hours, the mixture was filtered through Whatman filter paper. The obtained ethyl acetate extract was concentrated to dryness using a rotary evaporator, and the dried extracts were accurately weighed and dissolved in DMSO to prepare stock solutions at 1 mg/mL.
Determination of antibacterial activity of endophytic yeast extracts
The antibacterial activity of the total extracts was determined by the agar diffusion method. Gram-positive bacteria: Staphylococcus aureus and Bacillus subtilis; Gram-negative bacteria: Pseudomonas aeruginosa and Escherichia coli, and yeast Candida albicans were used. Gram-positive bacteria: Staphylococcus aureus and Bacillus subtilis; Gram-negative bacteria: Pseudomonas aeruginosa and Escherichia coli were inoculated into meat-peptone medium and grown in a thermostat at 37 °C for 18-20 hours. All bacterial suspensions were prepared according to the McFarland Standard at 0.5 (~1.5 x 108 CFU/mL). Bacterial suspensions were inoculated onto the surface of meat-peptone agar medium in a plate in an amount of 0.1 mL and incubated for 30 minutes. 10 mm diameter wells were formed, and 100 µL of 1 mg/mL solutions of the extracts in DMSO were poured. As a positive control, the antibiotic Cefazolin 1 mg/mL was used. DMSO was used as a negative control. After incubation of the Petri dishes in a thermostat at 37 °C for 24 hours, the zone of inhibition of the growth of test microorganisms was measured. Each experiment was repeated three times.
Antimicrobial activity against Candida albicans was evaluated using Sabouraud Dextrose Agar plates. The yeast inoculum was adjusted to 0.5 McFarland (~1 × 106 CFU/mL) and plates were incubated at 28 °C for 48 hrs. Clotrimazole 1 mg/mL were used as a positive control.
Identification and characterization of samples using reverse phase Nano-LC-MS/MS
Reverse-phase nano-liquid chromatography tandem mass spectrometry (Nano-LC-MS/MS) analysis was performed using an Agilent 1200 nano-flow liquid chromatography system coupled to an Agilent 6520B Q-TOF mass spectrometer. Chromatographic separation was achieved using a Zorbax SB-C18 CHIP column
(5 µm, 75 µm × 43 mm).
Samples were dissolved in methanol and filtered through a 0.22 µm syringe filter prior to analysis. The mobile phase consisted of solvent A (0.1% formic acid in water with 5% acetonitrile) and solvent B (acetonitrile containing 0.1% formic acid and 10% deionized water). A gradient elution program was applied as follows: 0-3 min, 0% B; 3-12 min, linear increase to 60% B; 12-18 min, 60% B; 18-20 min, return to 0% B. The flow rate was set at 0.6 µL/min during elution. A sample volume of 2.0 µL was injected using an Agilent Micro WPS autosampler.
Mass spectrometric detection was carried out using electrospray ionization (ESI) in positive ion mode. The operating conditions were as follows: drying gas (N₂) flow rate 4.0 L/min, temperature 350 °C, capillary voltage 2000 V, fragmentor voltage 175 V. Data were acquired over an m/z range of 50-3000 for MS and 50-2500 for MS/MS. MS/MS fragmentation was performed using collision-induced dissociation with collision energy ranging from 10-40 eV.
The mass spectrometer was calibrated prior to analysis using an Agilent ESI calibration mixture (m/z 118-2722). Caffeine (100 fmol/µL) was used as an internal standard to monitor system performance and stability. Data acquisition was performed in automatic MS/MS mode, selecting the most intense precursor ions.
Mass spectrometry data were processed using Agilent MassHunter Qualitative Analysis software (version B.07.00). Metabolite identification was carried out by comparing accurate mass (m/z values) and MS/MS fragmentation patterns with reference spectra available in the MassBank database. All compounds were reported as putatively identified based on spectral similarity.
Statistical analysis
All antimicrobial activity assays were performed in triplicate (n = 3), and the results were expressed as mean values ± standard deviation (SD). Prior to statistical analysis, data normality was assessed using the Shapiro Wilk test, and homogeneity of variances was evaluated using Levene’s test. Differences in inhibition zone diameters among the tested yeast strains were analyzed using one-way analysis of variance (ANOVA). When statistically significant differences were observed (P < 0.05), Tukey’s honestly significant difference (HSD) post hoc test was applied to determine pairwise differences between groups. All statistical analyses were performed using appropriate statistical software packages. A P-value of less than 0.05 was considered statistically significant.
Isolation and identification of endophytic Yeasts
A total of more than 50 endophytic yeast isolates were obtained from the generative parts of 18 plant species, including Malus domestica (apple), Prunus avium (sweet cherry), Prunus persica (peach), Fragaria vesca (strawberry), Morus alba (white mulberry), Morus nigra (black mulberry), Prunus armeniaca (apricot), Cerasus vulgaris (cherry), Ficus carica (fig), Punica granatum (pomegranate), Diospyros kaki (persimmon), Crataegus pontica (hawthorn), Prunus domestica (plum), Rubus idaeus (raspberry), Pyrus armeniacifolia (pear), Actinidia deliciosa (kiwi), Vitis vinifera (grape), and Ribes aureum (currant). Among the isolates, 41 were successfully identified to the species level using MALDI-TOF MS.
Table (1):
Identification result of endophytic yeasts by MALDI-TOF mass spectrometry
Yeast strains |
No. of isolates |
Host plant |
Isolate IDs |
Identification method |
|---|---|---|---|---|
Pichia kluyveri |
11 |
Fragaria vesca, Prunus persica, Malus domestica, Pyrus armeniacifolia, Cerasus vulgaris, Prunus domestica, Rubus idaeus |
59, 72, 73, 90, 76, 77, 98, 81, 91, 93, 99 |
MALDI-TOF mass spectrometry |
Pichia kudriavzevii |
5 |
Morus alba, Fragaria vesca, Morus nigra, Prunus avium |
56, 58, 65, 66, 62 |
MALDI-TOF mass spectrometry |
Hanseniaspora opuntiae |
3 |
Ficus carica, Fragaria vesca, Crataegus pontica. |
57, 92, 69 |
MALDI-TOF mass spectrometry |
Kluyveromyces marxianus |
1 |
Prunus avium |
61 |
MALDI-TOF mass spectrometry |
Nakazawae holstii |
1 |
Prunus avium |
63 |
MALDI-TOF mass spectrometry |
Hanseniaspora uvarum |
4 |
Diospyros kaki Thunb., Prunus armeniaca |
96, 17, 97, 14 |
MALDI-TOF mass spectrometry |
Metschnikowia pulcherrima |
8 |
Prunus armeniaca, Diospyros kaki Thunb., Prunus persica, Punica granatum |
1, 2, 3, 5, 6, 02, 021, 67 |
MALDI-TOF mass spectrometry |
Meyerozyma guilliermondii |
4 |
Prunus persica |
01, 03, 18, 101 |
MALDI-TOF mass spectrometry |
Meyerozyma carpophila |
2 |
Actinidia deliciosa |
1,03,108 |
MALDI-TOF mass spectrometry |
Candida dubliniensis |
2 |
Vitis vinifera |
107, 109 |
MALDI-TOF mass spectrometry |
Quambalaria cyanescens |
1 |
Malus domestica |
51 |
ITS rDNA sequencing |
Starmerella bacillaris |
1 |
Rubus idaeus |
80 |
ITS rDNA sequencing |
Naganishia adeliensis |
1 |
Diospyros kaki Thunb. |
95 |
ITS rDNA sequencing |
Total |
44 |
As shown in Table 1, MALDI-TOF MS can mainly identify certain groups of endophytic yeasts; it was not possible to identify strains 022, 023, 025, 028, 103 and 110. Furthermore, 3 isolates: 51, 80 and 95 were identified using molecular genetic methods as they exhibited distinct morphological features and were not successfully identified by MALDI-TOF MS. In this, they were identified using the sequences of the 18S rRNA gene, ITS1 and ITS2. The identification methods of the 18S rRNA gene, ITS1 and ITS2 play a very important role in microbial genetic analysis and taxonomic classification. Identification of the 18S rRNA gene: The 18S rRNA gene (ribosomal RNA) is an essential molecule located on the ribosome in eukaryotic cells and is involved in protein synthesis. The 18S rRNA gene is highly conserved and has a similar structure in many eukaryotic organisms. Identification of ITS1 and ITS2: Internal Transcribed Spacer (ITS) regions are non-transcribed regions located between ribosomal genes (18S, 5.8S, and 28S). ITS regions are highly variable in many eukaryotic organisms, particularly yeast and plants, but they are highly similar and are well suited for use in taxonomic classification.23 Also, samples for analysis must be very pure. Using the 18S rRNA gene, ITS1, and ITS2.
Based on molecular analyses, isolates 51, 80, and 95 were confirmed as Quambalaria cyanescens (PV153718), Starmerella bacillaris (PV153713) and Naganishia adeliensis (PV153716), respectively.
Antimicrobial activity of endophytic Yeast extracts
While the antibacterial activity initially did not show a specific inhibition zone when tested in the culture fluid of the isolates, the freeze-dried suspension showed inhibition zones.
In this study, antibacterial activity was tested in 29 strains of endophytic yeasts. Equal amounts of dry mass were obtained to extract for all strains, and all samples were standardized to a final concentration of 1 mg/mL. The potential effect of DMSO was evaluated using solvent controls, which showed no inhibitory activity.
Table (2):
Antimicrobial activity of endophytic yeast extracts against test microorganisms (mean ± SD, n = 3). Different letters within each column indicate statistically significant differences at P < 0.05 (one-way ANOVA followed by Tukey’s HSD test)
| No. | Yeast strains | Antibacterial activity (growth inhibition zones, mm) | ||||
|---|---|---|---|---|---|---|
| S. aureus | B. subtilis | P. aeruginosa | E. coli | C. albicans | ||
| Control | 30 ± 0.01 | 30 ± 0.01 | 23 ± 0.02 | 32 ± 0.01 | 22 ± 0.03 | |
| 1 | Meyerozyma guilliermondii 01 | — | 18 ± 0.8 | — | 16 ± 1.1 | — |
| 2 | Metschnikowia pulcherrima 02 | 35 ± 0.8 | 17 ± 0.8 | 32 ± 1.8 | 18 ± 1.1 | 19 ± 1.1 |
| 3 | Meyerozyma guilliermondii 03 | 36 ± 1.8 | 17 ± 1.2 | 32 ± 1.4 | 25 ± 0.6 | 18 ± 1.1 |
| 4 | Metschnikowia pulcherrima 3 | — | — | — | — | 12 ± 1.4 |
| 5 | Metschnikowia pulcherrima 5 | 18 ± 1.4 | 12 ± 0.6 | 15 ± 1.2 | 15 ± 0.8 | 15 ± 1.8 |
| 6 | Hanseniaspora uvarum 14 | 16 ± 1.4 | 17 ± 1.2 | 15 ± 1.8 | 16 ± 1.2 | 16 ± 1.1 |
| 7 | Meyerozyma guilliermondii 18 | 16 ± 1.2 | 18 ± 0.6 | 16 ± 1.4 | 16 ± 1.2 | — |
| 8 | Metschnikowia pulcherrima 021 | 22 ± 1.1 | 23 ± 0.8 | 25 ± 0.8 | 16 ± 1.2 | 20 ± 1.2 |
| 9 | 022 | 17 ± 1.2 | 25 ± 1.4 | 22 ± 1.2 | 25 ± 1.1 | 21 ± 0.6 |
| 10 | 023 | 25 ± 1.4 | 25 ± 0.6 | 25 ± 1.2 | 23 ± 1.8 | 23 ± 0.8 |
| 11 | 25 | 30 ± 1.8 | 22 ± 1.4 | 17 ± 0.8 | 13 ± 1.2 | 18 ± 0.8 |
| 12 | 28 | 33 ± 1.2 | 25 ± 0.8 | 22 ± 0.6 | 13 ± 1.4 | 18 ± 1.8 |
| 13 | Quambalaria cyanescens 51 | 17 ± 1.8 | 13 ± 1.4 | 18 ± 0.8 | 15 ± 1.1 | 17 ± 1.2 |
| 14 | Pichia kudriavzevii 56 | 17 ± 1.2 | 18 ± 1.4 | 15 ± 0.8 | — | — |
| 15 | Pichia kudriavzevii 58 | — | 16 ± 1.8 | 16 ± 0.6 | — | 15 ± 0.8 |
| 16 | Pichia kudriavzevii 62 | — | — | 18 ± 1.8 | — | — |
| 17 | Pichia kudriavzevii 66 | 17 ± 0.6 | 20 ± 0.8 | 18 ± 1.4 | 13 ± 1.2 | 17 ± 1.1 |
| 18 | Metschnikowia pulcherrima 67 | — | 20 ± 1.8 | 15 ± 1.1 | 15 ± 0.6 | — |
| 19 | Hanseniaspora opuntiae 69 | 14 ± 1.1 | 15 ± 0.8 | 14 ± 0.6 | 15 ± 1.1 | 16 ± 0.8 |
| 20 | Starmerella bacillaris 80 | 18 ± 1.2 | 15 ± 0.6 | 15 ± 0.8 | 15 ± 1.2 | 15 ± 1.1 |
| 21 | Naganishia adeliensis 95 | 15 ± 0.6 | 14 ± 0.8 | 17 ± 1.2 | 20 ± 1.1 | 17 ± 1.1 |
| 22 | Pichia kluyveri 98 | — | 18 ± 1.4 | — | 15 ± 1.8 | 14 ± 0.5 |
| 23 | Meyerozyma guilliermondii 101 | — | 15 ± 1.4 | — | — | 16 ± 0.5 |
| 24 | 103 | 18 ± 1.1 | 14 ± 0.8 | 16 ± 1.4 | 13 ± 1.1 | — |
| 25 | Candida dubliniensis 107 | 17 ± 0.6 | 17 ± 1.2 | 18 ± 1.1 | 15 ± 1.4 | 21 ± 1.1 |
| 26 | Meyerozyma carpophila 108 | 16 ± 1.1 | 17 ± 0.8 | 20 ± 0.6 | 15 ± 0.8 | 22 ± 1.4 |
| 27 | Candida dubliniensis 109 | — | 18 ± 0.8 | 18 ± 0.6 | 16 ± 0.8 | 23 ± 0.8 |
| 28 | 110 | 14 ± 1.2 | 16 ± 1.8 | 19 ± 1.4 | 17 ± 1.1 | 23 ± 1.1 |
As shown in Table 2, (“–” indicates absence of inhibition or inhibition zone diameter below the measurable threshold (10 mm)), several endophytic yeast isolates exhibited pronounced and broad-spectrum antimicrobial activity, characterized by large inhibition zone diameters against multiple test microorganisms. These findings are consistent with previous reports demonstrating that plant-associated and endophytic microorganisms possess strong antagonistic potential against pathogenic bacteria and yeasts.24 Similar inhibition patterns have been reported for microbial antagonists isolated from plant tissues, where inhibition levels reached up to 50%-70% against diverse pathogens.2,25 The relatively lower susceptibility of Gram-negative bacteria can be attributed to their outer membrane barrier; however, the detectable inhibition observed indicates that yeast-derived metabolites may partially overcome this intrinsic resistance.26 The inhibition zone data confirm that endophytic yeasts represent competitive antimicrobial agents comparable to, and in some cases exceeding, other reported microbial biocontrol systems.
Among the tested strains, Metschnikowia pulcherrima 02, Meyerozyma guilliermondii 03, and isolates 023 and 028 demonstrated the highest antibacterial activity overall. In particular, Metschnikowia pulcherrima 02 showed marked inhibition against Staphylococcus aureus (35 mm) and Pseudomonas aeruginosa (32 mm), indicating considerable activity against both Gram-positive and Gram-negative bacteria.
Inhibition zone diameters (mm) of selected yeast strains against Staphylococcus aureus, Bacillus subtilis, Pseudomonas aeruginosa, Escherichia coli, and Candida albicans. Data are presented as mean ± SD (n = 3).
Similarly, Meyerozyma guilliermondii 03 exhibited high inhibitory effects against S. aureus (36 mm), P. aeruginosa (32 mm), and Escherichia coli (25 mm), suggesting the presence of potent antimicrobial metabolites. Isolate 023 showed consistently high activity across all tested microorganisms, with inhibition zones reaching 25 mm against S. aureus, B. subtilis, and P. aeruginosa, and 23 mm against E. coli and Candida albicans, reflecting a broad-spectrum antimicrobial profile (Figure). Notably, isolates 028 and 025 also demonstrated considerable activity, particularly against S. aureus (33-30 mm) and B. subtilis
(25-22 mm), further supporting their antibacterial potential. High antifungal activity against Candida albicans was observed in isolates 023, 110, and Candida dubliniensis 109, with inhibition zones up to 23 mm, indicating effective anti-yeast properties.
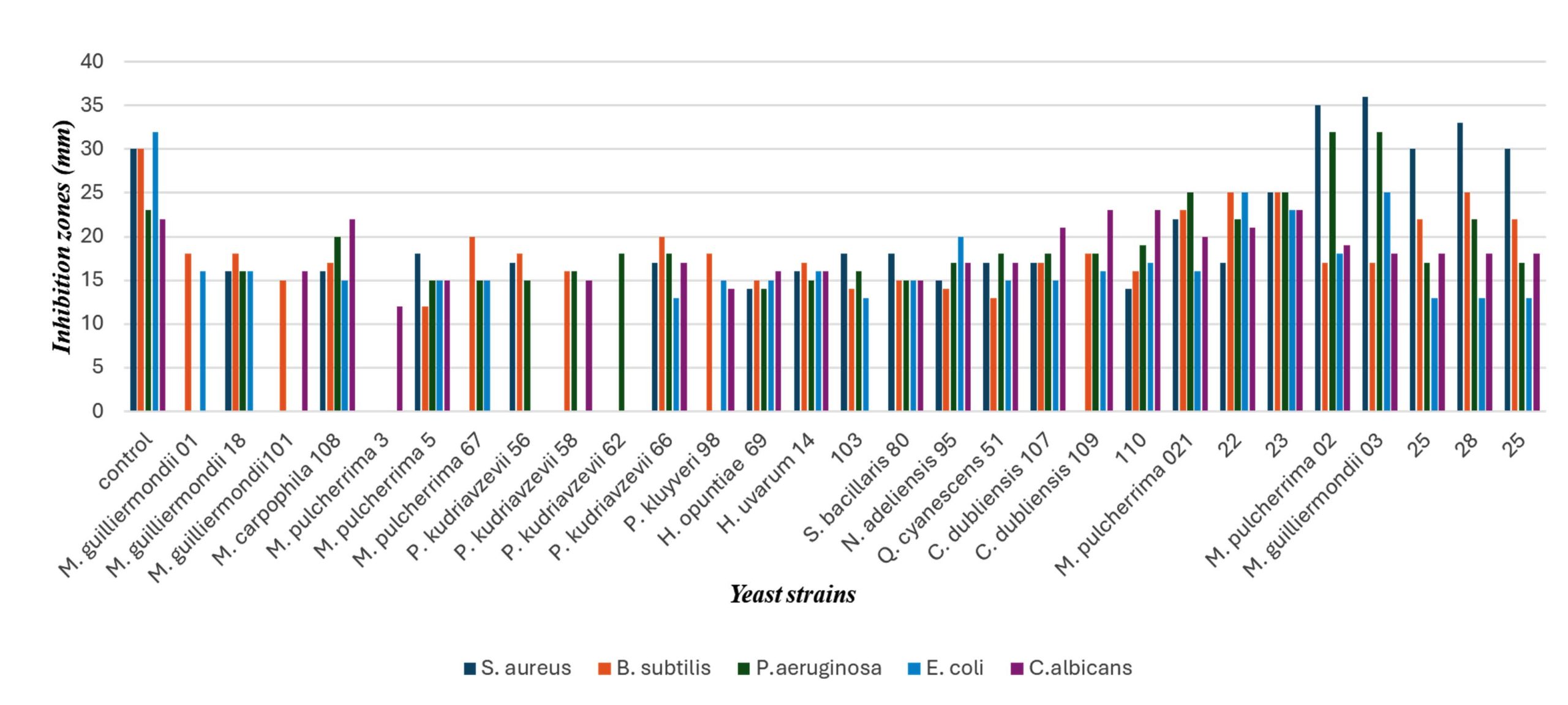
Figure. Antimicrobial activity of endophytic yeast extracts against test microorganisms
Overall, strains such as Metschnikowia pulcherrima 02, Meyerozyma guilliermondii 03, Meyerozyma carpophila 108, Pichia kudriavzevii 66, and isolate 023 can be considered the most promising candidates due to their notable and broad-spectrum antimicrobial activity.
Identification and characterization of samples using reverse phase Nano-LC-MS/MS
To elucidate the biochemical basis of the observed antimicrobial activity, extracts of the most active endophytic yeast strains were subjected to reverse-phase Nano-LC-MS/MS analysis. The analysis revealed the presence of several known antimicrobial and bioactive secondary metabolites, suggesting that the inhibitory effects observed in the agar diffusion assays are closely associated with metabolite production, as previously reported for endophytic yeasts and fungi.1,7,11,27,28
As shown in Table 3, the extract of Meyerozyma guilliermondii 03, Nano-LC-MS/MS analysis putatively identified andrastin D (m/z 428.25), lanostenol (m/z 428.4) and apigenin (m/z 270.17). Apigenin is a well-known flavonoid with documented broad-spectrum antibacterial,29 and antifungal activity, particularly against Staphylococcus aureus, Escherichia coli, and Pseudomonas aeruginosa, and is also known to inhibit biofilm formation.5,7,30 The presence of this compound may explain the marked inhibition zones observed for this strain against both Gram-positive and Gram-negative bacteria.
Table (3):
Putatively identification of antimicrobial metabolites produced by selected endophytic yeast strains using reverse-phase Nano-LC-MS/MS analysis
| No. | Compound name | Time (min) | Mass (g/mol) | Molecular Formula | Chemical structures | Fragments | Link | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Meyerozyma guilliermondii 03 | ||||||||||
| 1 | Andrastin D | 2.137 | 428.25629 | C₂₆H₃₆O₅ | 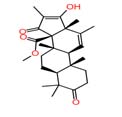 |
132.0912 195.1181 243.1389 343.1926 |
https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-AAFC-AC000821 Megan J. Kelman, Justin B.Renaud, Mark W. Sumarah, Agriculture and Agri-Food Canada | |||
| 2 | Lanostenol | 2.618 | 428.40182 | C₃₀H₅₂O | 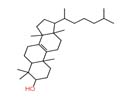 |
137.1328 413.3913 |
https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-mFam-MC02_000300 C. Broeckling, Colorado State University Proteomics and Metabolomics Facility |
|||
| 3 | Apigenin | 4.394 | 270.0528 | C₁₅H₁₀O₅ | 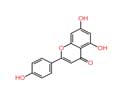 |
274.295 318.8723 391.7261 |
https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-mFam-MC21_000255 Stefanie Doell IPB |
|||
| Hanseniaspora uvarum 14 | ||||||||||
| 4 | Altersolanol A | 1.726 | 336.0845 | C₁₆H₁₆O₈ | 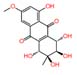 |
151.0385 | https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-AAFC-AC000070 Justin B. Renaud, Mark W. Sumarah, Agriculture and Agri-Food Canada |
|||
| 5 | Fumiquinazoline F | 1.809 | 358.14297 | C₂₁H₁₈N₄O₂ | 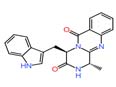 |
363.1670 381.1322 |
https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-AAFC-AC000912 Justin B. Renaud, Mark W. Sumarah, Agriculture and Agri-Food Canada |
|||
| 6 | Amygdalin | 2.991 | 457.432 | C₂₀H₂₇NO₁₁ | 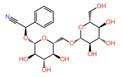 |
325.1138 | https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-RIKEN-PR300660 Tetsuya Mori, Center for Sustainable Resource Science, RIKEN |
|||
| 7 | 1,4-naphthalene-dione | 1.779 | 158.0368 | C₁₀H₆O₂ | 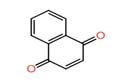 |
159.0443 | https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-UFZ-UA001803 C. Gallampois (Umea), E. Schymanski (Eawag), W. Brack (UFZ) | |||
| S. bacillaris 80 | ||||||||||
| 8 | 9(11)-Dehydroglycyrrhetic acid | 18.361 | 454.6993 | C₃₀H₄₆O₃ | 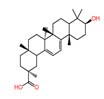 |
186.8000 275.1000 456.2000 |
https://massbank.eu/MassBank/RecordDisplay?id=MSBNK-RIKEN_NPDepo-CB000464 Nogawa T, Okano A, CSRS, RIKEN | |||
In isolate of Hanseniaspora uvarum 14, altersolanol A (m/z 336.08) and Fumiquinazoline F (m/z 358.14) were putatively identified. Altersolanol A is a naturally occurring biologically active compound derived from fungi that exhibits antimicrobial properties, particularly showing activity against Pseudomonas aeruginosa.31,32 Fumiquinazoline F is a member of the fumiquinazoline subgroup of quinoline alkaloids. This compound has been reported to exhibit both antibacterial and antifungal activities.33
In the extract of 028, Nano-LC-MS/MS analysis revealed a diverse metabolite profile, including amygdalin and 1,4-naphthoquinone. The coexistence of multiple antimicrobial metabolites in this strain correlates well with its broad-spectrum inhibitory activity against both bacterial and fungal test organisms. Although amygdalin is not considered a primary antibiotic compound, previous studies suggest that it may enhance antimicrobial activity indirectly or act synergistically with other bioactive metabolites.7,8
Similarly, Starmerella bacillaris 80 was found to produce 9(11)-Dehydroglycyrrhetic acid (m/z 454.69) 9(11)-Dehydroglycyrrhetic acid is a naturally derived triterpenoid compound obtained as an oxidized derivative of 18α-glycyrrhetinic acid. It has been investigated for antimicrobial activities.34
It should be noted that certain compounds, such as apigenin and amygdalin, may originate from plant material or medium components rather than being exclusively produced by yeast, and therefore their biosynthetic origin requires further validation.
Overall, the detection of 1,4- naphthoquinone, altersolanol A, fumiquinazoline F, and 9(11)-Dehydroglycyrrhetic acid. These compounds are known for antibacterial, antifungal, and antibiofilm activities, indicating that the observed inhibition effects may be associated with metabolite production.
The antimicrobial activity observed only after freeze-drying can be attributed to the comprehensive recovery of metabolites from both the culture supernatant and the biomass. Lyophilization was applied to the entire suspension, ensuring that extracellular as well as intracellular compounds were retained and subsequently extracted.
The present study demonstrates that endophytic yeasts isolated from diverse plant species in Uzbekistan represent a promising source of bioactive metabolites with antimicrobial potential. The LC–MS/MS analysis revealed the presence of several secondary metabolites, including 1,4-naphthoquinone, altersolanol A, fumiquinazoline F, and andrastin D, which are known to exhibit diverse biological activities. The antimicrobial activity observed in selected yeast isolates is therefore likely metabolite-mediated, consistent with previous reports highlighting the importance of fungal secondary metabolites in biocontrol mechanisms.
Among the detected compounds, 1,4-naphthoquinone derivatives represent the most significant contributors to antifungal activity. Recent studies have demonstrated that naphthoquinones exhibit strong and broad-spectrum antifungal effects, particularly against Candida albicans, including drug-resistant strains. These compounds act through multiple mechanisms, such as induction of oxidative stress, disruption of mitochondrial function, inhibition of glycolysis, and suppression of biofilm formation.35 Certain naphthoquinone derivatives have demonstrated higher antifungal efficacy than conventional drugs such as fluconazole and exert their activity through multiple mechanisms, including disruption of fungal cell membranes, induction of reactive oxygen species (ROS), and inhibition of biofilm formation in pathogenic yeasts.35,36
Polyketide-derived metabolites such as altersolanol A may also contribute to the observed antimicrobial activity. In this study, altersolanol A was putatively identified in Meyerozyma guilliermondii 03 based on MassBank spectral matching, consistent with previous reports describing its antimicrobial activity.37 However, its potential cytotoxicity requires careful evaluation.
Another important class of compounds identified in this study includes alkaloid metabolites such as fumiquinazoline F, which are commonly produced by Aspergillus species. Alkaloids of this class are known to possess a wide range of biological activities, including antimicrobial, anticancer, and enzyme-inhibitory effects.38 Their structural diversity and bioactivity make them valuable candidates for drug discovery, although their antimicrobial mechanisms are often complex and may involve interference with DNA synthesis, enzyme activity, or cellular signaling pathways.
Meroterpenoids such as andrastin D may also contribute to the observed antimicrobial activity. In this study, andrastin D was putatively identified in active yeast extracts, consistent with previous reports describing the antimicrobial potential of related andrastin derivatives.39 Its presence supports a possible role in the bioactivity observed, although further validation is required. In addition, regulatory studies have demonstrated that the production of such metabolites in fungi is controlled by specific biosynthetic gene clusters, highlighting the importance of genetic regulation in secondary metabolite production.40
The presence of multiple bioactive compounds within the same yeast extract suggests that the observed antimicrobial activity is likely the result of synergistic or additive interactions between metabolites, rather than the effect of a single compound. This is consistent with current understanding of microbial biocontrol systems, where multiple mechanisms—including production of diffusible antimicrobial compounds, competition for nutrients, and interference with pathogen virulence—act simultaneously to suppress pathogenic microorganisms.41 Such multi-target activity is particularly advantageous in preventing the development of resistance and enhancing overall antimicrobial efficacy. In particular, metabolites associated with Metschnikowia species, such as iron-chelating pigments (e.g., pulcherrimin), have been shown to suppress microbial growth by inducing nutrient deprivation and oxidative stress.42, 43
It is also important to distinguish between bioactive metabolites and non-contributing compounds detected in the LC–MS analysis. While compounds such as lanosterol were identified, they are primarily structural sterols involved in membrane biosynthesis and do not contribute directly to antimicrobial activity. Similarly, some detected compounds may originate from plant material rather than yeast metabolism, emphasizing the need for careful interpretation of metabolomic data.
Overall, the results highlight the significant potential of endophytic yeasts as sources of antimicrobial metabolites. Compounds such as quinone derivatives (e.g., 1,4-naphthoquinone), along with polyketides, alkaloids, and meroterpenoids, are known for their antimicrobial properties and may contribute to the observed bioactivity. However, this study does not provide direct evidence of causality, as no bioassay-guided fractionation, purification of individual compounds, or testing of isolated metabolites was performed. Therefore, the relationship between detected metabolites and antimicrobial activity should be considered as a hypothesis based on co-occurrence rather than a confirmed mechanism. The observed effects are likely associated with a combination of metabolites acting synergistically. Further studies are required to validate the role of individual compounds and establish definitive structure–activity relationships.
Importantly, this study represents one of the first reports from Uzbekistan describing the isolation and diversity assessment of endophytic yeasts from plant hosts, along with their screening for antimicrobial activity. In addition, the metabolite profiles of selected strains were analyzed using Nano-LC-MS/MS and putatively identified through MassBank database matching, highlighting the significance of this work in linking microbial diversity with bioactive potential. These findings provide a basis for future studies focused on the isolation of individual antimicrobial compounds, comparison with standard substances, and their potential application in pharmaceutical development.
This study demonstrates that endophytic yeasts isolated from diverse plant hosts in Uzbekistan represent a previously underexplored and promising source of antimicrobial activity. Several strains, particularly Metschnikowia pulcherrima 02, Meyerozyma guilliermondii 03, Pichia kudriavzevii 66, and isolate 023, exhibited pronounced and broad-spectrum inhibitory effects against both bacterial and fungal pathogens.
Nano-LC–MS/MS analysis, combined with MassBank-based spectral matching, revealed the presence of multiple bioactive secondary metabolites, including naphthoquinones, polyketides, alkaloids, and meroterpenoids, suggesting that the observed antimicrobial activity may be associated with metabolite production. However, as individual compounds were not isolated or experimentally validated, this relationship should be interpreted as a putative association rather than a confirmed mechanism.
The findings highlight the importance of Uzbekistan’s endophytic yeast biodiversity as a potential reservoir of bioactive compounds and support its relevance for the development of metabolite-based biocontrol agents and natural antimicrobial products. Importantly, this study provides one of the first integrated insights into the diversity, antimicrobial potential, and metabolite profiles of endophytic yeasts from this region.
Future research should focus on bioassay-guided fractionation, purification of active metabolites, quantitative structure–activity analysis, and comparison with standard antimicrobial agents to validate their practical applicability in pharmaceutical and agricultural systems.
ACKNOWLEDGMENTS
The authors thank the Institute of Bioorganic Chemistry of the Academy of Sciences of Uzbekistan, the U.A. Arifov Institute of Ion-Plasma and Laser Technologies, and the Sanitary and Epidemiological Surveillance Service under the Administration of the President of Uzbekistan for their assistance in metabolite analysis and MALDI-TOF-based microbial identification.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
DS designed the study and performed experiments. LA performed data analysis. DS wrote the manuscript. LA reviewed and revised the manuscript. TG supervised the study and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
ETHICS STATEMENT
Not applicable.
- Ling L, Tu Y, Ma W, et al. A potentially important resource: endophytic yeasts. World J Microbiol Biotechnol. 2020;36(8):1-7.
Crossref - Hagaggi NSA, Khalaf MEA, El Rady EA. Bioactivity of endophytic bacteria inhabiting the Egyptian medicinal plant Hyoscyamus muticus. Novel Res Microbiol J. 2025;9(2):51-62.
Crossref - Gulyamova T, Abdulmyanova L, Ruzieva D, et al. Diversity of endophytic fungi associated with medicinal plants of Uzbekistan and their biological properties. Acta Microbiol Bulg. 2022;38(2):91-97.
- Tayung K. Studies on endophytic microorganisms associated with Taxus spp. [PhD thesis]. Gauhati University. 2008.
- Siddiqui MS, Thodey K, Trenchard I, Smolke CD. Advancing secondary metabolite biosynthesis in yeast with synthetic biology tools. FEMS Yeast Res. 2012;12(2):144-170.
Crossref - Jahangeer M, Mahmood Z, Mashook A, et al. Yeast cells secondary metabolites and their industrial applications: a review. Pak J Med Biol Sci. 2019;2(1):62-67.
- Silva FV, Souza GB, Nogueira AR. Use of yeast crude extract for sequential injection determination of carbohydrates. Anal Lett. 2001;34(8):1377-1388.
Crossref - Utama GL, Rahmah SA, Kayaputri IL, Balia RL. Antibacterial activities of indigenous yeasts isolated from pomegranate peels (Punica granatum L.). J Adv Pharm Technol Res. 2022;13(1):56-60.
Crossref - Hicks RH, Moreno-Beltrán M, Gore-Lloyd D, Chuck CJ, Henk DA. The oleaginous yeast Metschnikowia pulcherrima displays killer activity against avian-derived pathogenic bacteria. Biology. 2021;10(12):1227.
Crossref - Csutak O, Corbu V. The yeasts – versatile antimicrobial agents. Rev Biol Biomed Sci. 2020;3(1):1-9.
Crossref - Zhao J, Mou Y, Shan T, et al. Antimicrobial metabolites from the endophytic fungus Pichia guilliermondii isolated from Paris polyphylla var. yunnanensis. Molecules. 2010;15(11):7961-7970.
Crossref - Sipiczki M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl Environ Microbiol. 2006;72(10):6716-6724.
Crossref - He Y, Degraeve P, Oulahal N. Bioprotective yeasts: Potential to limit postharvest spoilage and to extend shelf life or improve microbial safety of processed foods. Heliyon. 2024;10(3).
Crossref - Cecarini V, Cuccioloni M, Bonfili L, et al. Identification of a killer toxin from Wickerhamomyces anomalus with β-glucanase activity. Toxins. 2019;11(10):568.
Crossref - Leyva Salas M, Mounier J, Valence F, Coton M, Thierry A, Coton E. Antifungal microbial agents for food biopreservation: a review. Microorganisms. 2017;5(3):37.
Crossref - Rahman M, Islam R, Hasan S, et al. A comprehensive review on bio-preservation of bread: an approach to adopt wholesome strategies. Foods. 2022;11(3):319.
Crossref - Atakhanov A, Ashurov N, Abdurazakov M, et al. Preparation and properties of nanocomposite films based on carboxymethyl-cellulose and melanin from Cladosporium endophytic fungus. Polym Bull. 2025;82(17):12031-12049.
Crossref - Bhuyan B, Hansepi PK, Akhtar S, et al. Isolation of yeast endophytes from healthy seeds of Capsicum annuum L. and assessment of their antimicrobial activity. J Appl Biol Biotechnol. 2023;11(6):111-116.
Crossref - Abdel-Hafez SI, Abo-Elyousr KA, Abdel-Rahim IR. Leaf surface and endophytic fungi associated with onion leaves and their antagonistic activity against Alternaria porri. Czech Mycol. 2015;67(1):1-22.
Crossref - Kurtzman CP, Fell JW, Boekhout T, eds. The Yeasts: A Taxonomic Study. 4th ed. Elsevier. 2000.
- Bizzini A, Durussel C, Bille J, Greub G, Prodhom G. Performance of matrix-assisted laser desorption ionization–time of flight mass spectrometry for identification of bacterial strains routinely isolated in a clinical microbiology laboratory. J Clin Microbiol. 2010;48(5):1549-1554.
Crossref - Wagner K, Springer B, Pires VP, Keller PM. Molecular detection of fungal pathogens in clinical specimens by 18S rDNA high-throughput screening in comparison to ITS PCR and culture. Sci Rep. 2018;8(1):6964.
Crossref - Polomska X, Juszczyk P, Cadez N, et al. Comparison of physiological and PCR-RFLP rDNA identification of yeast species commonly found in cheese. Pol J Food Nutr Sci. 2007;57(2):221-226
- Prima SR, Elfahmi EE, Julianti E, Fidrianny I. Evaluation of antibacterial and antioxidant activity of endophytic fungi isolated from Capsicum annuum L. and Allium cepa L. Pharmacogn J. 2022;14(2):329-334.
Crossref - Abdulmyanova LI, Gulyamova TG, Ruzieva DM, Rasulova GA, Yusupov UK. The impact of plant extracts on the metabolic profile of endophytic fungi isolated from vinca plants. Plant Cell Biotechnol Mol Biol. 2020;21(57-58):109-117.
- Younis G, Awad A, Dawod RE, Yousef NE. Antimicrobial activity of yeasts against some pathogenic bacteria. Vet World. 2017;10(8):979.
Crossref - Hussein MA, Khalaf KJ, Salman JAS. Antibiofilm activity of dextran purified from Saccharomyces boulardii against MDR-Pseudomonas aeruginosa isolated from wounds and burns. Novel Res Microbiol J. 2025;9(6):438-453.
Crossref - Pohsnem JM, Nath BJ, Sarma S, Parasar DP. Isolation, identification and characterization of an indigenous pigmented yeast isolated from Assam, North East, India. J Pure Appl Microbiol. 2025;19(3):1908-1918.
Crossref - Benalach L, Boukada F, Cherifi K, Latreche A, Messellem I, Bellatreche M. Investigating the bioactive potential of Ononis spinosa L.: phytochemical profiling and antibacterial effectiveness evaluation. Novel Res Microbiol J. 2025;9(2):82-91.
Crossref - Karpiński TM, Adamczak A, Ożarowski M. Antibacterial activity of apigenin, luteolin, and their C-glucosides. In: Proceedings of the 5th International Electronic Conference on Medicinal Chemistry. Basel, Switzerland: MDPI; 2019.
Crossref - Haraguchi H, Abo T, Hashimoto K, Yagi A. Action mode of antimicrobial altersolanol A in Pseudomonas aeruginosa. Biosci Biotechnol Biochem. 1992;56(8):1221-1224.
Crossref - Sharma N, Kushwaha M, Qayum A, Singh SK, Goutam U, Jaglan S. Exploring bioactive metabolites from endophytic fungi of Albizia lebbeck (L.) Benth. J Pure Appl Microbiol. 2026;20(1):612-621.
Crossref - Rystsov GK, Antipova TV, Renfeld ZV, et al. Fumiquinazolines F and G from the fungus Penicillium thymicola demonstrate anticancer efficacy against triple-negative breast cancer MDA-MB-231 cells by inhibiting epithelial-mesenchymal transition. Int J Mol Sci. 2025;26(15):7582.
Crossref - Liu Y, Sheng R, Fan J, Guo R. A review on structure-activity relationships of glycyrrhetinic acid derivatives with diverse bioactivities. Mini Rev Med Chem. 2022;22(15):2024-2066.
Crossref - Chen Q, Yang M, Lai X, et al. Antifungal potential of naphthoquinone derivatives: screening of shikonin-based compounds and mechanistic insights into 5,8-dihydroxy-1,4-naphthoquinone against Candida albicans in vitro and in vivo. Microbiol Spectr. 2026;14(4):e02438-25.
Crossref - Borba-Santos LP, Nicoletti CD, Vila T, et al. A novel naphthoquinone derivative shows selective antifungal activity against Sporothrix yeasts and biofilms. Braz J Microbiol. 2022;53(2):749-758.
Crossref - Aly AH, Debbab A, Edrada-Ebel RA, et al. Protein kinase inhibitors and other cytotoxic metabolites from the fungal endophyte Stemphylium botryosum isolated from Chenopodium album. Mycosphere. 2010;1(2):153-162.
- Wang H, Sun L, Ma X, et al. Polyketides and alkaloids from the fungus Aspergillus fumigatus YB4-17 and ent-fumiquinazoline J induce apoptosis and paraptosis in human hepatoma HepG2 cells. Front Pharmacol. 2024;15:1487977.
Crossref - Qin YY, Huang XS, Liu XB, et al. Three new andrastin derivatives from the endophytic fungus Penicillium vulpinum. Nat Prod Res. 2022;36(13):3262-3270.
Crossref - Marcano Y, Montanares M, Gil-Duran C, et al. Pr laeA affects the production of roquefortine C, mycophenolic acid, and andrastin A in Penicillium roqueforti but has little impact on asexual development. J Fungi. 2023;9(10):954.
Crossref - Roca-Couso R, Flores-Felix JD, Rivas R. Mechanisms of action of microbial biocontrol agents against Botrytis cinerea. J Fungi. 2021;7(12):1045.
Crossref - Moreno Beltran M. Using Diverse Methods to Expand the Sustainable Industrial Promise of an All-Rounder Yeast, Metschnikowia pulcherrima [PhD thesis]. University of Bath; 2021. https://researchportal.bath.ac.uk/en/studentTheses/using-diverse-methods-to-expand-the-sustainable-industrial-promis/
- Devi RP, Yamunasri P, Balachandar D, Murugananthi D. Potentials of soil yeasts for plant growth and soil health in agriculture: a review. J Pure Appl Microbiol. 2025;19(1):1-18.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.