


ISSN: 0973-7510
E-ISSN: 2581-690X
Antibiotic resistant bacteria are present in various environments and are concerning for public health. Antibiotic resistance genes in soil may proliferate throughout different ecosystems and be acquired by pathogens that pose health risks to humans and animals. This study investigated the antibiotic resistance of cultivable bacteria isolated from livestock-exposed and unexposed areas in a former perlite mining area in the Fa-La-Mee Mountain range, Lopburi, Thailand. Soil samples were collected in five different locations. Soil bacteria were isolated and tested for susceptibility against seven antibiotics, followed by 16S rRNA gene-based identification. A total of 56 bacterial isolates were isolated from the perlite-rich soil samples, including 34 isolates from the livestock-exposed areas and 22 isolates from the unexposed areas. Most of the isolates were resistant to cefepime (26.5% in exposed areas and 22.7% in unexposed areas) and ampicillin (23.5% in exposed areas and 22.7% in unexposed areas). 16S rRNA gene sequences revealed that most of the resistant bacteria isolated from perlite-rich soil were Bacillus spp. A multidrug-resistant isolate of Bacillus cereus resistant to ampicillin, trimethoprim/sulfamethoxazole, and cefepime was found in a livestock-unexposed area. The majority of the culturable antibiotic-resistant bacteria isolated from perlite-rich soil were cefepime-resistant Bacillus. Interestingly, the pattern of antibiotic resistance in exposed and unexposed areas was not different, which indicated that antibiotic resistance in perlite-rich soil is likely caused by other factors, such as physicochemical characteristics and/or the microbial population of the soil rather than by antibiotics used in livestock husbandry.
Antimicrobial Resistance, Perlite, Livestock Farming, Bacillus
Antimicrobial resistance (AMR) is a global issue that affects human, animal, and environmental health. According to the one health concept, AMR can spread from the environment to humans and have negative effects on human health.1,2 Previous studies have demonstrated that populations of antibiotic-treated livestock freely reared in local grassland carry residual antibiotics or antibiotic resistant bacteria/genes that spread AMR in the environment.1,3
Perlite is a type of volcanic aluminosilicate glass that is formed by rapid cooling of lava or magma. Perlite is used in many industries, including for concrete fillers, thermal insulators, catalysts, filter, and agriculture because of its low density, high porosity, chemical inertness, and lack of toxicity.4 Perlite mining occurs in many countries around the world, including China, Turkey, Greece, USA, Japan, Italy, Hungary, and Macedonia.4
A perlite mine in Thailand was operated in the Fa-La-Mee mountain range, Lopburi province between 1992 to 2017.5 The Fa-La-Mee mountain range was the only economic source of perlite in Thailand.6 Following the concession ending, the mining area was left without any restoration. After being abandoned for several years, the flat parts of the mine underwent natural rehabilitation and gradually turned to grasslands which were then used for livestock grazing. Antibiotics such as amoxicillin, kanamycin, gentamicin, streptomycin, ceftiofur, and sulfamethoxazole/trimethoprim, readily available in local veterinary pharmacies, are commonly used by farmers in the area to treat livestock infections without veterinary oversight. Importantly, many of these antibiotics are identical to, share the same mode of action with, or belong to the same antibiotic family as those used in human medicine.7,8 Improper use of antibiotic in livestock farming can increase the spread of AMR in the environment and raise the risk of cross-resistance in human pathogens.1,9
Previous reports demonstrated that mining activities disrupt soil microbial communities and can co-select for antibiotic resistance genes in bacteria. Mining waste, particularly heavy metals, can induce stress in bacteria, leading to hypermutation and increased antibiotic resistance.10,11 In this study, the flat parts of the mine were used for livestock grazing. The presence of grazing animals also affects the soil physicochemical properties, microbial communities as well as the spread of antibiotic resistance genes.12 Several studies reported that waste from antibiotic-treated livestock is associated with a rise in antibiotic-resistant bacteria in the soil, especially Enterobacteriaceae.3,13-15 Certain remaining areas of the mine remains steep slope or covered in dry dipterocarp forest which are inaccessible to livestock and thus remain free from contamination by commercial antibiotics.
As perlite naturally deposits near the Earth’s surface, the topsoil in both livestock-exposed and -unexposed areas contains a high amount of perlite, resulting in a distinct perlite-rich soil.16 Previous studies have demonstrated that plants cultivated in perlite tend to harbor higher levels of Enterobacteriaceae compared to those grown in other substrates. These findings suggest that perlite may provide a suitable environment for the proliferation of Enterobacteriaceae.17,18 Although perlite mining has occurred in several countries, data on the spread of AMR in perlite-rich soil are limited. Due to the unique characteristics of perlite, the grazing animal may influence the soil bacterial communities as well as increase the level of the soil AMR. Therefore, in this study, we investigated AMR in perlite-rich soil by comparing areas with livestock exposure to those without.
In this study, the levels of antibiotic-resistant bacteria from livestock-exposed areas and unexposed areas were evaluated to determine the relationship between AMR and livestock exposure in perlite-rich soil.
Study sites and soil sample collection
The soil samples were collected from the Fa-La-Mee mountain range in the Sa Bot district of Lopburi province, Thailand. Soil samples were collected in December 2022 (dry season) from livestock-exposed and unexposed areas in five different locations around the Fa-La-Mee mountain range (Figure 1). These locations were grazed by different herds of livestock. At each location, two areas were designated: one with livestock-exposed and one without. This resulted in a total of 10 sampling sites (5 exposed and 5 unexposed). To minimize bacterial cross-contamination, sampling sites in livestock-exposed and unexposed zones were located at least 10 meters apart and separated by natural barriers, such as forests or steep slopes. Five replicated plots, (1 × 1 m) located more than 2 m apart were selected at each of the sampling sites. Before sample collection, topsoil debris were removed. Five samples were collected at a depth of 10 cm in each plot. The samples were combined into one composite sample and sieved through a 2 mm mesh to remove large debris and organic litter. The sampling method was adapted from a previous study.19 Approximately 100 g of the combined sample was collected in a sterile tube, stored at 4°C and transported to the laboratory.
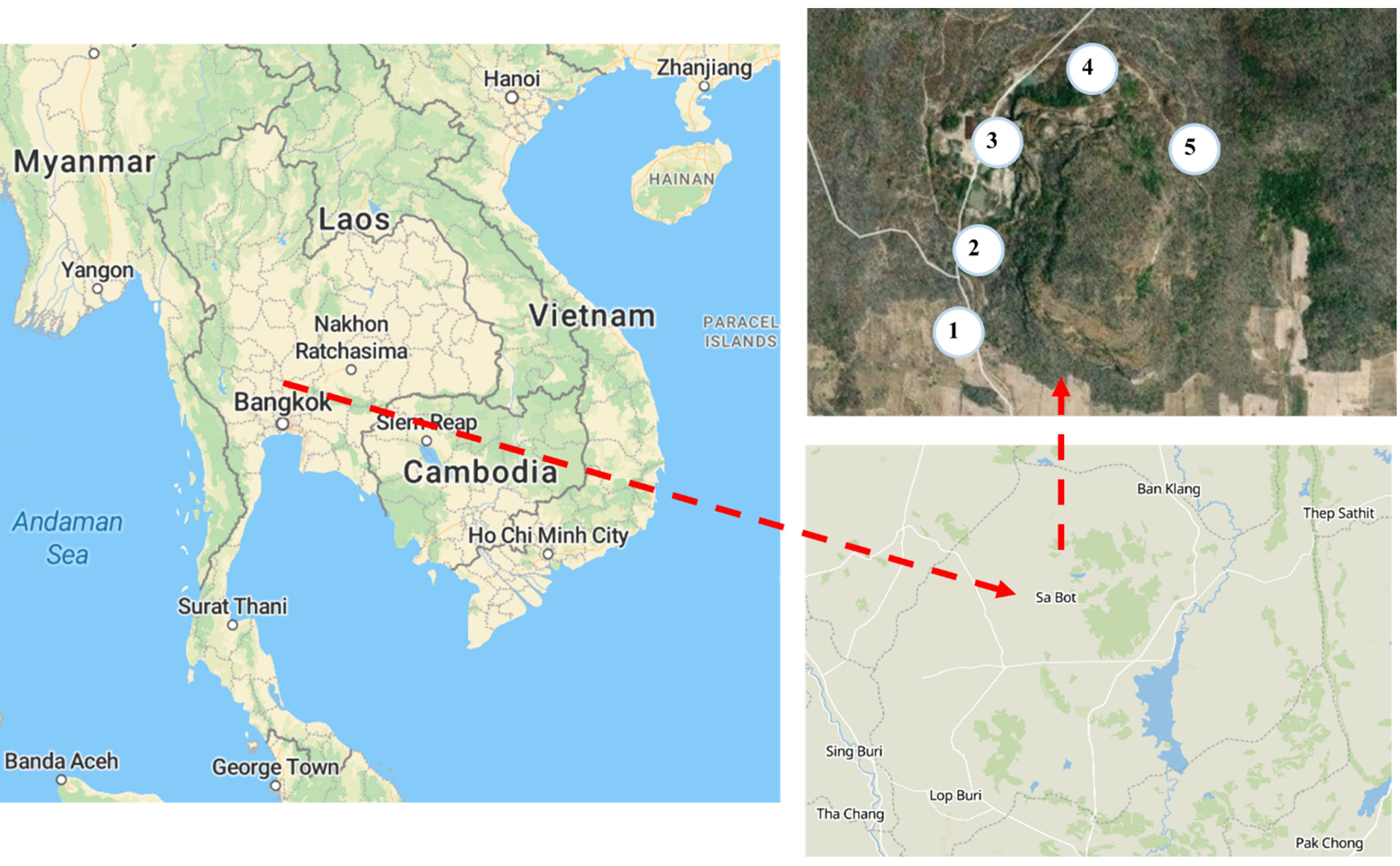
Figure 1. Study sites in the Fa-La-Mee mountain range, Sa Bot district, Lopburi province, Thailand. Livestock-exposed and -unexposed soil samples were collected at five sampling sites (1-5). The maps were obtained from Open Street Map (ODC Open Database License)
Isolation and maintenance of soil bacteria
To isolate soil bacteria, 1 g of each soil sample was dissolved in diluent (sterile normal saline solution), vortex homogenously and the volume was adjusted to 10 mL. Sample was mix thoroughly before performing serially dilution. The suspension was then serially diluted and 100 µL of the suspension at dilutions of 10-2 and 10-3 were spread on nutrient agar (Himedia) plates. The plates were incubated for up to 3 days at 37°C, according to previous study with some modifications.20-21 To acquire the pure isolate, the single colonies were picked up and sub-cultured on new nutrient agar plates. The pure isolate of bacteria was stored in 25% glycerol at -80°C for further studies.
Antimicrobial susceptibility testing
Pure isolates were tested for antibiotic susceptibility according to the Clinical and Laboratory Standards Institute (CLSI) standard by the disc diffusion method.20 Before preparing the bacterial suspension, the isolates were cultured in Mueller–Hinton broth (Himedia) overnight at 37°C with shaking at 150 rpm. The turbidity of the suspension was adjusted to McFarland 0.5 using Mueller–Hinton broth. After gently mixing the suspension, a sterile cotton swab was dipped into the suspension and any excess was removed by gently rotating the swab against the surface of the tube. The swab was then used to distribute the bacteria over surface of Mueller–Hinton agar (Himedia). The plates were left to stand at room temperature for 5 min and then antibiotic discs were placed on the designated area of the Mueller–Hinton agar. The antibiotics used in this study were ampicillin (10 µg), gentamicin (10 µg), cefepime (30 µg), ciprofloxacin (5 µg), trimethoprim/sulfamethoxazole (25 µg), tetracycline (30 µg), chloramphenicol (30 µg). All antibiotic discs were purchased from Oxoid Ltd. (Thermo Fisher Scientific). After incubation overnight at 37°C, the inhibition zones around the discs were measured and the isolates were classified as sensitive or resistant according to the standardized table supplied by the CLSI.
Molecular identification of bacteria by 16S rRNA gene sequencing
The universal primers 27F (5- AGAGTTTGATC(A/C)TGGCTCAG-3) and 1492R (5- TACGG(C/T)TACCTTGTTACGACTT-3) were used for amplification of the 16S rRNA gene fragment.21 DreamTaq PCR Master Mix (Thermo Fisher Scientific) was used for 16S rRNA gene amplification. The final concentration of each forward and reverse primer was 0.4 µmol. For DNA template, a single colony was picked up and dissolved in 50 µL of dH2O, heated at 95°C for 2 min, and centrifuged at 12,000 g for 1 min. An aliquot (2 µL) of the supernatant containing the DNA was used as the template. The PCR cycling conditions were as follows: an initial denaturation for 5 min at 95°C, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, extension at 72°C for 1 min, and then a final extension for 5 min at 72°C. The amplified PCR products were electrophoresed on 1% agarose gel. The amplified 16S rRNA gene fragment was purified and sequenced using DNA sequencing services (ATGC) using the same primer as for PCR amplification. The 16S rRNA gene sequences were exported into the Basic Local Alignment Search Tool (BLASTN) available at the National Center for Biotechnology Information (NCBI) website to identify matches with existing characterized reference sequences. A maximum likelihood phylogenetic tree was constructed using MEGA1122 with the 16S rRNA gene full-length sequences.
Statistic analysis
Fisher’s exact test was used to determine a statistically significant association between livestock exposure and AMR percentage in perlite-rich soil. The test was performed by RStudio.
Characteristics of the study site
The details for the study sites are given in Table 1. The livestock-exposed areas were plain areas with open grassland. The unexposed areas were steep slope or slope areas covered by dry dipterocarp forest. Livestock were not contained within set areas. The livestock were mixed-breed cattle or Siamese buffalo. The antibiotics use was reported by famers in the area which were amoxicillin, kanamycin, gentamicin, streptomycin, ceftiofur, and sulfamethoxazole/trimethoprim. Antibiotics are typically administered to livestock via injection for treatment of infections or wounds. The soil samples were sandy with white, light gray, or gray in color, which indicated that perlite was present in the soil. Examples of the study site geology and soil texture are shown in Figure 2.
Table (1):
Description of the study sites in the Fa-La-Mee mountain range
| Area | Coordinates | Area description | |
|---|---|---|---|
| Livestock-exposed area | Unexposed area | ||
| 1 | 15.289291, 100.876245 | Plain, open grassland Livestock: mixed-breed cattle |
Slope covered by dry dipterocarp forest |
| 2 | 15.290831, 100.876875 | Plain, open grassland Livestock: Siamese buffalos |
Slope covered by dry dipterocarp forest |
| 3 | 15.292203, 100.877317 | Plain, grassland Livestock: mixed-breed cattle |
Steep slope |
| 4 | 15.293443, 100.879061 | Plain, grassland Livestock: mixed-breed cattle |
Steep slope |
| 5 | 15.292420, 100.881079 | Plain, open grassland Livestock: mixed-breed cattle |
Slope covered by dry dipterocarp forest |

Figure 2. Examples of the geology of the study site and soil texture. (a) The geology of the study site 4 showing a livestock-exposed area (grassland) and unexposed area (steep slope). Textures of the soil samples collected from livestock exposed (b) and unexposed (c) areas at study site 4
Isolation of bacteria from the perlite-rich soil
A total of 56 cultivable bacteria were isolated from the perlite-rich soil samples, including 34 from livestock-exposed areas and 22 from unexposed areas. The colonies of isolated bacteria exhibited distinct characteristics for color, size, pigmentation, and texture. The isolates were then subjected to Gram staining and observed under a microscope to investigate their morphologies. Gram-positive bacteria (76.5%) were more abundant than gram-negative bacteria (23.5%) in the livestock-exposed area. Similarly, most of bacteria in the unexposed area were Gram-positive (86.4% compared with 13.64% for Gram-negative). Almost all the isolated bacteria were rod shaped, but some were filamentous (14% in the livestock-exposed areas and 4.3% in the unexposed areas).
Antibiotic sensitivity of cultivable bacteria from perlite-rich soil
The bacteria isolated from the perlite-rich soil in both livestock-exposed and unexposed areas were subjected to antibiotic susceptibility tests. The following seven antibiotics were used to test antibiotic sensitivity in accordance with the CLSI guidelines; ampicillin, gentamicin, cefepime, ciprofloxacin, trimethoprim/sulfamethoxazole, tetracycline, and chloramphenicol (Figure 3). The antibiotics used in this study were identical or relevant to the antibiotics used in livestock husbandry in the study area. The CLSI guidelines provide criteria for interpreting the clear zone diameter for discrimination between sensitive and resistant traits. The criteria depend on the bacterial species and the types of antibiotics. Because the isolated bacteria in this study were quite diverse and some genera were not listed in the guidelines, we decided to use the minimum zone diameter of each antibiotic to distinguish between sensitive and resistant traits. Isolates that had a clear zone diameter smaller than the minimum zone diameter of the tested antibiotic were categorized as resistant. Among the 34 bacteria isolated from the livestock-exposed area, nine isolates (26.5 %) showed resistance to cefepime, and eight isolates (23.5%) showed resistance to ampicillin. Three bacteria were resistant to one each of antibiotic which included chloramphenicol, gentamicin, and trimethoprim/sulfamethoxazole. Interestingly, bacteria from unexposed areas exhibited a similar resistance pattern. The percentage of isolates that showed resistance to cefepime and ampicillin resistance was 22.7% and three bacteria were resistant to one each of antibiotic which included chloramphenicol, gentamicin, and trimethoprim/sulfamethoxazole. All isolates showed high susceptibility to tetracycline and ciprofloxacin (Table 2). Fisher’s exact test revealed no significant association between livestock exposure and the percentage of AMR in perlite-rich soil (p = 1).
Table (2):
Number of isolate and percentage of antibiotic resistant bacteria isolated from perlite-rich soil in livestock-exposed and unexposed areas
| Area | Number of resistant isolate (percent) | ||||||
|---|---|---|---|---|---|---|---|
| AMP | C | TE | CN | CIP | SXT | FEP | |
| Exposed area | 8 (23.5) | 1 (2.9) | 0 (0) | 1 (2.9) | 0 (0) | 1 (2.9) | 9 (26.5) |
| Unexposed area | 5 (22.7) | 1 (4.5) | 0 (0) | 1 (4.5) | 0 (0) | 1 (4.5) | 5 (22.7) |
Abbreviations: AMP, ampicillin; C, chloramphenicol; TE, tetracycline; CN, gentamicin; CIP, ciprofloxacin; SXT, trimethoprim/sulfamethoxazole; and FEP, cefepime
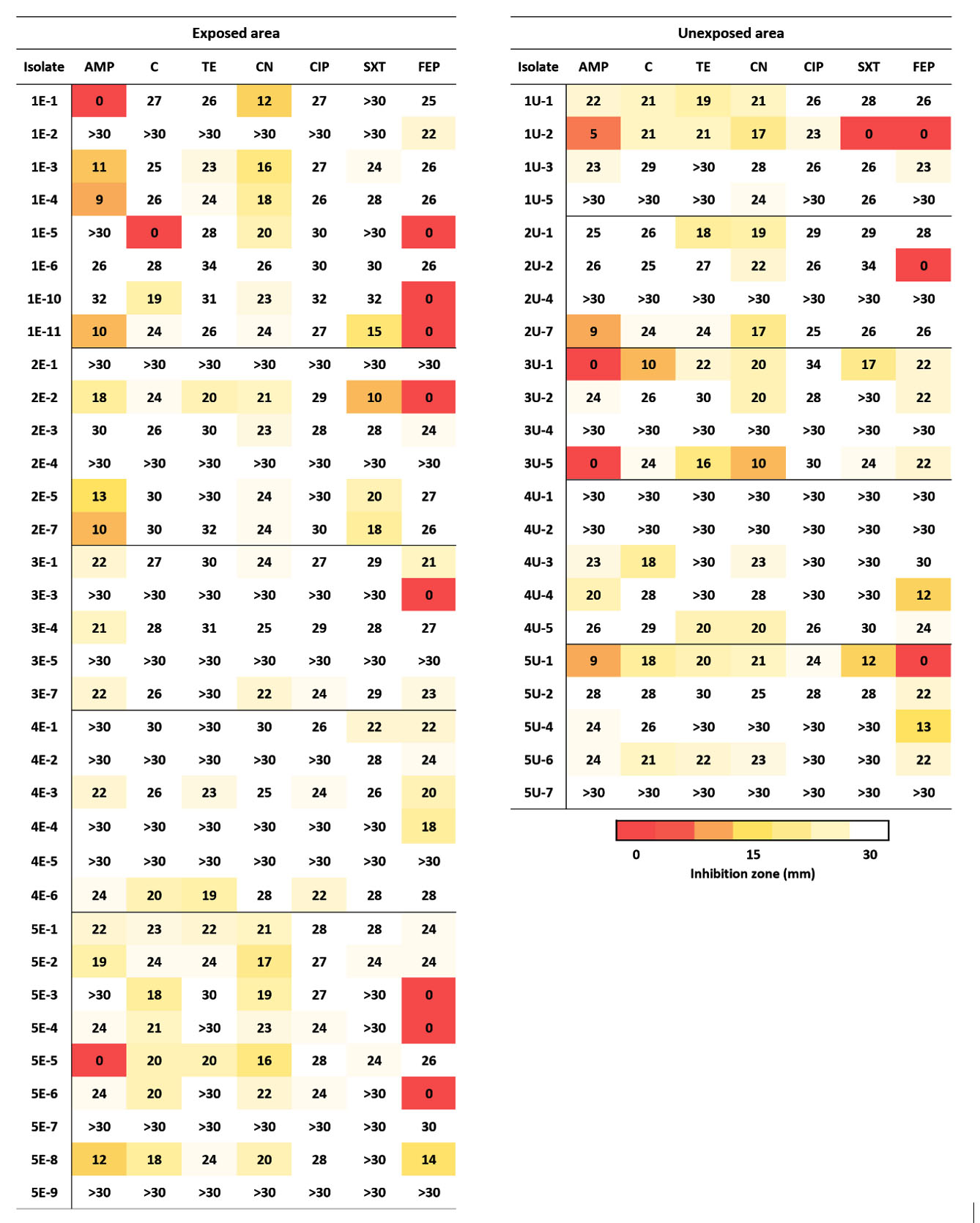
Figure 3. Diameter of the antibiotic inhibition zone. The diameter of the inhibition zone is shown in millimeters. Abbreviations: AMP, ampicillin; C, chloramphenicol; TE, tetracycline; CN, gentamicin; CIF, ciprofloxacin; SXT, trimethoprim/sulfamethoxazole; and FEP, cefepime
16S rRNA identification of resistant isolates
Resistant isolates were identified by 16S rRNA gene sequencing. The resistance pattern, identification result, and percent similarity of each isolate are presented in Table 3. A phylogenetic tree was drawn using the full-length 16S rRNA gene sequence (Figure 4). Most of the culturable bacteria isolated from the perlite-rich soil were Bacillus spp. Notably, all Bacillaceae (Bacillus, Peribacillus, and Fictibacillus) in both areas were resistant to cefepime. For the exposed areas, three Bacillus isolates were resistant to both ampicillin and cefepime, and an isolate of Chryseobacterium arthrosphaerae was resistant to both ampicillin and gentamicin. Ampicillin-resistant Enterobacter, Diaphorobacter, Stenotrophomonas, and Chryseobacterium were also found. In the unexposed areas, three isolates were resistant to two antibiotics. These were Pseudomonas nitroreducens, which showed resistant to ampicillin and chloramphenicol, Burkholderia sp. (non-pseudomallei), which were resistant to ampicillin and gentamicin, and Bacillus cereus, which was resistant to ampicillin and cefepime. Remarkably, an isolate of B. cereus was found to be resistant against three agents (ampicillin, trimethoprim/sulfamethoxazole, and cefepime) and was considered multi-drug resistant (MDR) (Table 3).
Table (3):
16S rRNA gene-based identification of antibiotic resistant bacteria isolated from perlite-rich soil
| Area | Isolate | Resistant pattern | Genus/species | % identity |
|---|---|---|---|---|
| Exposed | 1C-1 | AMP/CN | Chryseobacterium arthrosphaerae | 99.77 |
| 1C-3 | AMP | Enterobacter sp. | 99.46 | |
| 1C-4 | AMP | Enterobacter sp. | 98.82 | |
| 1C-5 | C/FEP | Bacillus licheniformis | 100 | |
| 1C-10 | FEP | Bacillus pumilus | 100 | |
| 1C-11 | AMP/FEP | Bacillus paramycoides | 100 | |
| 2C-2 | AMP/FEP | Bacillus cereus | 99.93 | |
| 2C-5 | AMP | Diaphorobacter sp. | 99.85 | |
| 2C-7 | AMP | Stenotrophomonas acidaminiphila | 99.8 | |
| 3C-3 | FEP | Bacillus sp. | 99.43 | |
| 5C-3 | FEP | Bacillus pumilus | 100 | |
| 5C-4 | FEP | Bacillus sp. | 100 | |
| 5C-5 | AMP | Chryseobacterium sp. | 99.85 | |
| 5C-6 | FEP | Bacillus pumilus | 100 | |
| 5C-8 | AMP/FEP | Bacillus licheniformis | 99.93 | |
| Unexposed | 1U-2 | AMP/SXT/FEP | Bacillus cereus | 99.93 |
| 2U-2 | FEP | Bacillus pumilus | 100 | |
| 2U-7 | AMP | Klebsiella variicola | 100 | |
| 3U-1 | AMP/C | Pseudomonas nitroreducens | 100 | |
| 3U-5 | AMP/CN | Burkholderia sp. | 99.93 | |
| 4U-4 | FEP | Peribacillus simplex | 100 | |
| 5U-1 | AMP/FEP | Bacillus cereus | 100 | |
| 5U-4 | FEP | Fictibacillus solisalsi | 100 |
Abbreviations: AMP, ampicillin; C, chloramphenicol; TE, tetracycline; CN, gentamicin; CIP, ciprofloxacin; SXT, trimethoprim/sulfamethoxazole; and FEP, cefepime
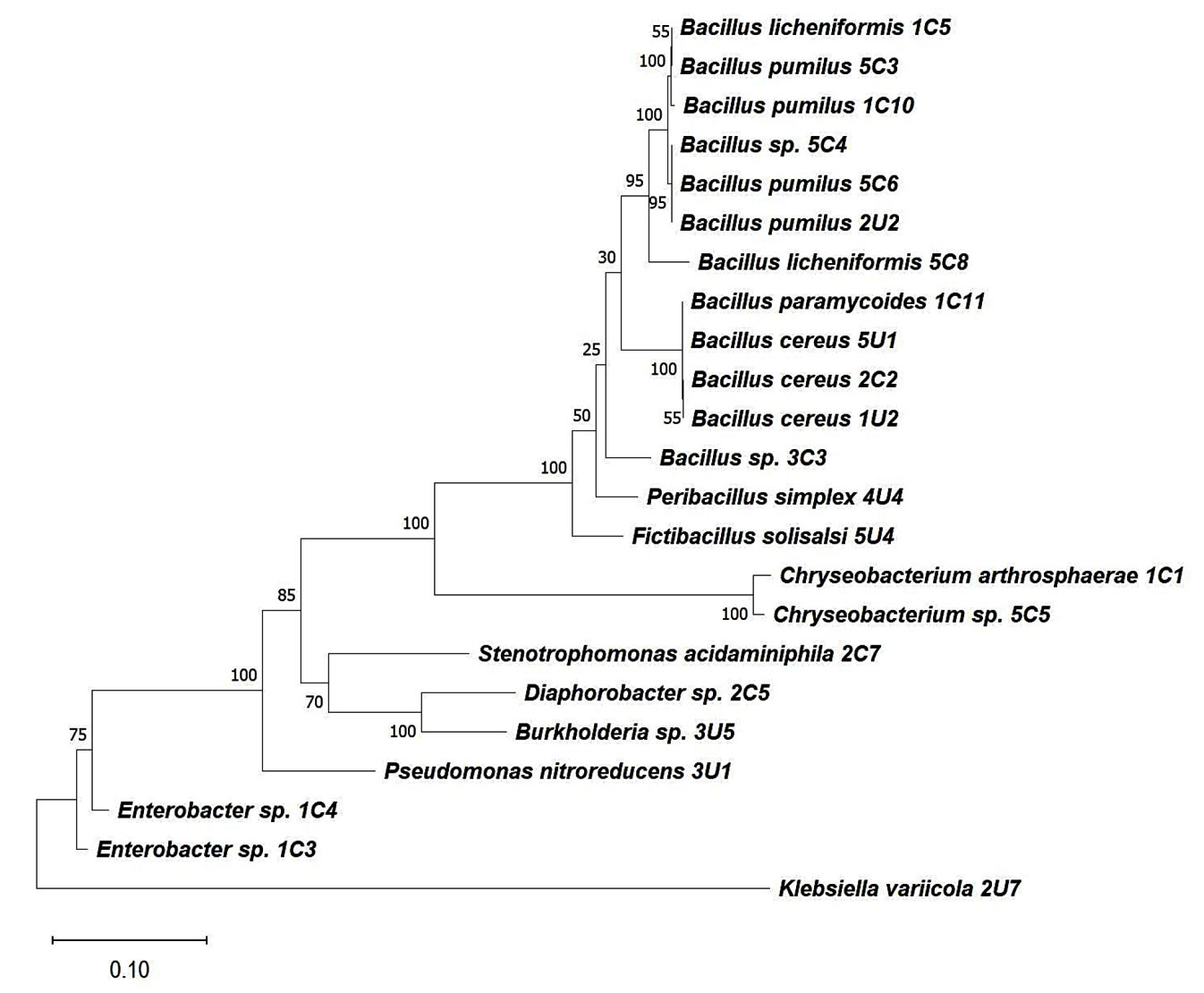
Figure 4. 16S rRNA phylogeny of resistant isolates. The phylogenetic tree was built with full-length 16S rRNA gene sequences using the Maximum Likelihood method in MEGA11. Bootstrap values (1000 replicates) are displayed for important internal nodes
Perlite is a valuable mineral that is used in many industries because of its properties.4 In Thailand, a perlite mine was historically operated in the Fa-La-Mee mountain range. After the end of the mining concession, parts of the mining area were gradually covered by grassland. In the past few years, people from nearby areas have used the grasslands for livestock grazing. The soil in these areas has a high perlite content, and this creates a unique environment that is not present in other parts of Thailand. In the current study, cultivable bacteria from perlite-rich soil in livestock-accessible and livestock-inaccessible areas were isolated and tested for antibiotic resistance. Most of the cultivable bacteria in the perlite-rich soil samples from both the livestock-exposed and unexposed areas were Gram-positive bacilli. The 16S rRNA gene sequences analysis revealed that most of the bacteria were Bacillus spp. Interestingly, an antibiotic susceptibility test revealed that AMR bacteria in perlite-rich soil appeared to be livestock independent because the resistant patterns were not different between livestock-exposed and unexposed areas. Many studies have suggested that soil bacteria spontaneously carry different types of antibiotic resistant genes that allow for survival against natural antibiotics present in the environment.23,24 For example, bacteria in remote untouched Antarctic soils spontaneously carry antibiotic resistant genes without antibiotic exposure.25 Almost all the ampicillin-resistant bacteria found in this study were previously reported to possess intrinsic ampicillin resistance.26-31 Several studies have showed that antibiotic-resistant bacteria are common in soil contaminated with livestock waste, particularly antibiotic-resistant Enterobacteriaceae. However, these studies lacked a non-contaminated control group.13-15 In this study, only few resistant Enterobacteriaceae were found (two isolates in the exposed areas and one isolate in the unexposed areas). A previous study demonstrated that the survival of Escherichia coli, a member of the Enterobacteriaceae family, varied with the soil type and that survivability of E. coli was minimal in sandy soil.32 Thus, the survival of Enterobacteriaceae in perlite-rich soil may be limited because of the physicochemical properties of the soil. Additionally, a study on the microbiome composition of soils from sandy-gravel mining complexes revealed that actinobacteria was the most abundant phyla across all samples.33 Members of actinobacteria such as Streptomyces are well recognized for their ability to synthesize many antibiotics that can alter soil bacterial communities.34,35 Previous studies that indicated perlite as a favorable environment for Enterobacteriaceae growth utilized heat-treated perlite as a planting medium, which seemingly eliminated the original microbial community. In contrast, our study employed unprocessed perlite directly from its natural environment, which contained complex microbial composition. This highlights the native microbial community’s potential role in controlling AMR Enterobacteriaceae within perlite-rich soils.16,17 Overall, our results indicated that AMR in perlite-rich soil is likely driven by intrinsic resistance, physicochemical properties, and/or the microbial community of the soil rather than antibiotic use in livestock farming. Investigation of the physicochemical properties and microbial communities of perlite-rich soil could offer deeper insights into spread of AMR within this unique environment.
Our results highlighted the resistant nature of Bacillus spp. against cefepime. Cefepime is a fourth-generation cephalosporin with broad activity against several Gram-positive and Gram-negative bacteria. Cefepime is normally used for treating nosocomial pneumonia, urinary tract infections, skin and skin structure infections, and intra-abdominal infections.36 A study on AMR profile of thermophilic Bacillus licheniformis isolated from soil in Iraq showed 100% resistant to cefepime.37 Bacillus cereus isolated from dairy products in China also exhibited a high level of resistance to cefepime (96.30%).38 Additionally, approximately 20 % of Bacillus species found in powdered infant milk in Saudi Arabia were cefepime resistant.39 Furthermore, many Bacillus species displayed a high level of resistance to other cephalosporins, including ceftazidime, cephalothin, and cefotaxime.40,41 In the present study, other members of the Bacillaceae family (Peribacillus simplex and Fictibacillus solisalsi) were also resistant to cefepime. Interestingly, a MDR B. cereus that was resistant to cefepime, ampicillin, and trimethoprim/sulfamethoxazole was isolated from the unexposed area. Resistance of B. cereus against different classes of antibiotics, including b-lactam antibiotics, cephalosporins, sulfonamides, and macrolides, has been reported worldwide.42-44 Bacillus spp. harbored several antibiotic resistant genes which involved several mechanisms included inactivation of antimicrobial agents, efflux pump, drug target modification such as enzymatic modifications by aminoglycoside 5-phosphotransferase (aph-5) and methylation in 23S rRNA.45-47 In addition, the antibiotic-resistant gene in manured soil is interconnected with the bacterial community in plants which is the potential risks of plant resistome migration to the food chain.48 Although it is well-known that B. cereus causes food poisoning by producing enterotoxins, there are increasing reports of it causing serious non-gastrointestinal tract infections, such as cutaneous and blood stream infections.49-51 Although B. cereus infection is rare, the spread of MDR B. cereus spread in the environment should be monitored because it may pose a health risk.
Most of culturable antibiotic resistant bacteria isolated from perlite-rich soil in this study were cefepime-resistant Bacillaceae. Antibiotic resistant patterns in livestock-exposed and unexposed areas are not different. These results indicated that antibiotic resistance in perlite-rich soil was likely not driven by antibiotic use in livestock farming but potentially by the physicochemical properties and/or microbial community of perlite-rich soil. Future study should include physicochemical analyses, microbial community profiling together with full antibiotic resistance profiling of both culturable and non-culturable bacteria to provide complete information about AMR, which will be useful for developing AMR prevention and control strategies in the future.
ACKNOWLEDGMENTS
The authors would like to thank Monchanok Makpum and Nichamon Parinyapon for laboratory assistance, and Gabrielle David, PhD, from Edanz for editing a draft of this manuscript.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
NL and JP conceptualized, designed the study and performed supervision. WH collected resources. NL, JP, SS, LY, JS, RM, AP and NJ performed investigation. NL, JP, SS, LY, JS, RM, AP, and NJ applied methodology and performed experiments. NL and JP performed formal analysis. NL and JP wrote the manuscript. WH reviewed, edited and approved the manuscript for publication.
FUNDING
This study was supported by Chulabhorn Royal Academy with funding grant number PSCM.011/66.
DATA AVAILABILITY
All datasets generated or analysed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies with human participants or animals performed by any of the authors.
- Tyrrell C, Burgess CM, Brennan FP, et al. Antibiotic resistance in grass and soil. Biochem Soc Trans. 2019;47(1):477-486.
Crossref - Aslam B, Khurshid M, Arshad MI, et al. Antibiotic resistance: One health one world outlook. Front Cell Infect Microbiol. 2021;11:771510.
Crossref - Wang H, Qi JF, Qin R, Ding K, Graham DW, Zhu YG. Intensified livestock farming increases antibiotic resistance genotypes and phenotypes in animal feces. Commun Earth Environ. 2023;4:123.
Crossref - Reka A, Pavlovski B, Lisichkov K, et al. Chemical, mineralogical and structural features of native and expanded perlite from Macedonia. Geologia Croatica. 2023;72(3):215-221.
Crossref - Department of Primary Industries and Mines. Thai Mining Concessions Database,2023. https://www1.dpim.go.th/mne/mn.php?pltname=&prov= 16&ore1=&ore 2=&status=%E0%BB% D4%B4%A1%D2%C3& a1dx=00&a1mx= 00&a1yx=0000&a2dx=00&a 2mx=00&a2yx=0000 &b1dx=00&b1mx=00&b1yx= 0000&b2dx=00&b2mx=00&b2yx= 0000&nox=& remexp=&pageid=2. Accessed 17 December 2023
- Saisuthichai, D. Property and uses of perlite. [in Thai]. STR2/2006. Bangkok: Department of Mineral Resources. 2006. http://library.dmr.go.th/elib/cgi-bin/opacexe.exe?op=mmvw&db=Main&sid=&skin=u&usid =&mmid=4350&bid= 20466, Accessed 17 December 2023
- Moulin G, Cavalie P, Pellanne I, et al. Antimicrobial resistance ad hoc group of the French food safety agency. A comparison of antimicrobial usage in human and veterinary medicine in France from 1999 to 2005. J Antimicrob Chemother. 2008;62(3):617-625.
Crossref - Prescott JF. History and current use of antimicrobial drugs in veterinary medicine. Microbiol Spectr. 2017;5(6).
Crossref - He Y, Yuan Q, Mathieu J, et al. Antibiotic resistance genes from livestock waste: occurrence, dissemination, and treatment. npj Clean Water. 2020;3(4):4.
Crossref - Gabay T, Petrova E, Gillor O, Ziv Y, Angel R. Only a minority of bacteria grow after wetting in both natural and post-mining biocrusts in a hyperarid phosphate mine. soil. 2023;9(1):231-242.
Crossref - Yi X, Liang JL, Su JQ, et al. Globally distributed mining-impacted environments are underexplored hotspots of multidrug resistance genes. ISME J. 2022;16(9):2099-2113.
Crossref - Mhuireach GA, Dietz L, Gillett T. One or many? Multi-species livestock grazing influences soil microbiome community structure and antibiotic resistance potential. Front Sustain Food Syst. 2022;6:926824.
Crossref - Amador P, Fernandes R, Prudencio C, Duare I. Prevalence of antibiotic resistance genes in multidrug-resistant Enterobacteriaceae on Portuguese livestock manure. Antibiotics. 2019;8(1):23.
Crossref - Mukuna W, Aniume T, Pokharel B, Khwatenge C, Basnet A, Kilonzo-Nthenge A. Antimicrobial susceptibility profile of pathogenic and commensal bacteria recovered from cattle and goat farms. Antibiotics. 2023;12(2):420.
Crossref - Jesumirhewe C, Springer B, Allerberger F, Ruppitsch W. Genetic characterization of antibiotic resistant Enterobacteriaceae isolates from bovine animals and the environment in Nigeria. Front Microbiol. 2022;13:793541.
Crossref - Lexa J, Varga P, Uhlik P, Kodira P, Biroo A. Rajnoha M. Perlite deposits of the Central Slovakia Volcanic Field (Western Carpathians): Geology and properties. Geologica Carpathica. 2021;72(3):253-281.
Crossref - Isik H, Topalcengiz Z, Guner S, Aksoy A. Generic and Shiga toxin-producing Escherichia coli (O157:H7) contamination of lettuce and radish microgreens grown in peat moss and perlite. Food Control. 2020;111:107079.
Crossref - Friesem G, Reznik N, Cohen MS, Carmi N, Kerem Z, Yedidia I. Root-Associated Microbiomes, Growth and Health of Ornamental Geophytes Treated with Commercial Plant Growth-Promoting Products. Microorganisms. 2021; 9(8).
Crossref - Sansupa C, Purahong W, Wubet T, et al. Soil bacterial communities and their associated functions for forest restoration on a limestone mine in northern Thailand. PLoS One. 2021;16(4):e0248806.
Crossref - Clinical and Laboratory Standards Institute (CLSI), Performance standards for antimicrobial susceptibility testing, 33rd edition. USA: Clinical and Laboratory Standards Institute. 2023.
- Heuer H, Krsek M, Baker P, Smalla K, Wellington EM. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl Environ Microbiol. 1997;63(8):3233-41.
Crossref - Tamura K, Stecher G, Kumar S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol. 2021;38(7):3022-3027.
Crossref - Canton R. Antibiotic resistance genes from the environment: a perspective through newly identified antibiotic resistance mechanisms in the clinical setting. Clin Microbiol Infect. 2009;15(1):20-25.
Crossref - Willms IM, Kamran A, Abmann NF, et al. Discovery of novel antibiotic resistance determinants in forest and grassland soil metagenomes. Front Microbiol. 2019;10:460.
Crossref - Van Goethem MW, Pierneef R, Bezuidt OKI, De peer YB, Cowan DA, Makhalanyane TP. A reservoir of ‘historical’ antibiotic resistance genes in remote pristine Antarctic soils. Microbiome. 2018;6:40.
Crossref - Rhodes KA, Schweizer HP. Antibiotic resistance in Burkholderia species. Drug Resist Updat. 2016;28:82-90.
Crossref - de Campos TA, de Almeida FM, de Almeida APC, et al. Multidrug-resistant (MDR) Klebsiella variicola strains isolated in a Brazilian hospital belong to new clones. Front Microbiol. 2021;16;12:604031.
Crossref - Damas MSF, Ferreira RL, Campanini EB, et al. Whole genome sequencing of the multidrug-resistant Chryseobacterium indologenes isolated from a patient in Brazil. Front Med. 2022;9:931379.
Crossref - Mojica MF, Humphries R, Lipuma JJ, et al. Clinical challenges treating Stenotrophomonas maltophilia infections: an update. JAC Antimicrob Resist. 2022;5;4(3):dla040.
Crossref - Intra J, Carcione D, Sala RM, Siracusa C, Brambilla P, Leoni V. Antimicrobial resistance patterns of Enterobacter cloacae and Klebsiella aerogenes strains isolated from clinical specimens: A twenty-year surveillance study. Antibiotics. 2023;12(4):775.
Crossref - Zhai Z, Cui C, Li X, et al. Prevalence, antimicrobial susceptibility, and antibiotic resistance gene transfer of Bacillus strains isolated from pasteurized milk. J Dairy Sci. 2023;106(1):75-83.
Crossref - Alegbeleye O, Sant’Ana AS. Survival behavior of six enterotoxigenic Escherichia coli strains in soil and biochar-amended soils. Environ Res. 2023;223:115443.
Crossref - Kimeklis AK, Gladkov GV, Tembotov RH, et al. Microbiome composition of disturbed soils from sandy-gravel mining complexes with different reclamation approaches. One Ecosystem. 2022;7:e83756.
Crossref - Abbasi S, Spor A, Sadeghi A, Safaie N. Streptomyces strains modulate dynamics of soil bacterial communities and their efficacy in disease suppression caused by Phytophthora capsici. Sci Rep. 2021;11(1):9317.
Crossref - Alam K, Mazumder A, Sikdar S, et al. Streptomyces: the biofactory of secondary metabolites. Front Microbiol. 2022;13:968053.
Crossref - Pais GM, Chang J, Barreto EF, et al. Clinical pharmacokinetics and pharmacodynamics of Cefepime. Clin Pharmacokinet. 2022;61(7):929-953.
Crossref - Banoon S, Ali Z, Salih T. Antibiotic resistance profile of local thermophilic Bacillus licheniformis isolated from Maysan province soil. Comunicata Scientiae. 2020;11:e3291.
Crossref - Zhao S, Chen J, Fei P, et al. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J Dairy Sci. 2020;103(5):3994-4001.
Crossref - Alanber MN, Alharbi NS, Khaled JM. Evaluation of multidrug-resistant Bacillus strains causing public health risks in powdered infant milk formulas. J Infect Public Health. 2020;13(10):1462-1468.
Crossref - Tarale P, Gawande S, Jambhulkar V. Antibiotic susceptibility profile of Bacilli isolated from the skin of healthy humans. Braz J Microbiol. 2015;46(4):1111-1118.
Crossref - Adamski P, Byczkowska-Rostkowska Z, Gajewska J, Zakrzewski AJ, Klebukowska L. Prevalence and antibiotic resistance of Bacillus sp. isolated from raw milk. Microorganisms. 2023;11(4):1065.
Crossref - Navaneethan Y, Effarizah ME. Prevalence, toxigenic profiles, multidrug resistance, and biofilm formation of Bacillus cereus isolated from ready-to eat cooked rice in Penang, Malaysia. Food Control. 2021;121; 107553.
Crossref - Algammal AM, Alfifi KJ, Mabrok M, et al. Newly Emerging MDR B. cereus in Mugil seheli as the first report commonly harbor nhe, hbl, cytK, and pc-plc Virulence Genes and bla1, bla2, tetA, and ermA Resistance Genes. Infect Drug Resist. 2022;15:2167-2185.
Crossref - Xiao D, Tong C, Yang T, et al. First insights into antimicrobial resistance, toxigenic profiles, and genetic diversity in Bacillus cereus isolated from Chinese sausages. LWT. 2023;181;114717.
Crossref - Bottone EJ. Bacillus cereus, a volatile human pathogen. Clin Microbiol Rev. 2010;23(2):382-398.
Crossref - Wash P, Batool A, Mulk S, et al. Prevalence of antimicrobial resistance and respective genes among Bacillus spp., a versatile Bio-fungicide. Int J Environ Res Public Health. 2022;19(22):14997.
Crossref - Blanco P, Hernando-Amado S, Reales-Calderon JA, et al. Bacterial multidrug efflux pumps: much more than antibiotic resistance determinants. Microorganisms. 2016;4(1):14.
Crossref - Zhang YJ, Hu HW, Chen QL, et al. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ Int. 2019;130:104912.
Crossref - Ko JH, Kang CI, Lee WJ, et al. Clinical features and risk factors for development of Bacillus bacteremia among adult patients with cancer: a case-control study. Support Care Cancer. 2015;23(2):377-384.
Crossref - Glasset B, Herbin S, Granier SA, et al. Bacillus cereus, a serious cause of nosocomial infections: Epidemiologic and genetic survey. PLoS One. 2018;13(5):e0194346.
Crossref - Esmkhani M, Shams S. Cutaneous infection due to Bacillus cereus: a case report. BMC Infect Dis. 2022;22(1):393.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.