


ISSN: 0973-7510
E-ISSN: 2581-690X
Existing antifungal drugs available today are low in efficacy, high in toxicity and cause multidrug resistance. Anti-Candida potential of geraniol, an acyclic monoterpene alcohol was evaluated and secretion of hydrolytic enzymes, adherence and biofilm formation were studied as virulence attributes. XTT reduction assay, inverted light microscopy and scanning electron microscopy (SEM) were used to study the effect of geraniol on biofilm formation. In vitro secretion of proteinases and phospholipases was observed on plates containing BSA and egg yolk, respectively. Geraniol showed minimum inhibitory concentration (MIC) of 130 µg/ml for C. albicans and C. glabrata and 80 µg/ml for C. tropicalis. Inhibition of biofilms was observed at sub-MIC values of geraniol. Proteinase secretion decreased in C. albicans, C. tropicalis and C. glabrata by 32.5-38.7%, 46.51%, 31.83%, respectively at MIC of geraniol. The clinical sensitive C. albicans D-27 and resistant S-1 showed 36.95% and 57.53% proteinase inhibition, respectively. Only minor inhibitory effects of geraniol were observed on phospholipase secretory activity. At MIC, the standard and resistant C. albicans strains showed 10-20% reduction in phospholipase inhibition, while the clinically sensitive strain showed an inhibition of 33%. Results show that geraniol interferes with the initial steps of fungal virulence and biofilm formation and can be used in the management and treatment of both topical and systemic Candida infections.
Candida, Geraniol, Biofilms, Proteinases, Phospholipases
Bloodstream Candida infections (Candidemia) become detrimental in immune-compromised patients and those in intensive care unit (ICU). The overall increase in incidence ranges from 75-400% depending on the size of the hospitals1, 2. Having high mortality rates and treatment costs, early investigation and management is essential to control these persistent and increasingly drug resistant fungal pathogens. Candida albicans is the most frequently isolated species but non-albicans species like C. tropicalis and C. glabrata are also to blame for nosocomial threats in mildly immunocompromised patients2, 3. The virulence factors that contribute to Candida pathogenesis include biofilm formation, phenotypic switching, adhesion to host tissues, hyphal formation, and secretion of hydrolytic enzymes4. The yeast form is associated with dissemination in blood, whereas the hyphal form is responsible for host tissue infiltration during early stages of infection and hence crucial for pathogenesis. Virulence in C. albicans is related to extracellular secretion of hydrolytic enzymes, mainly phospholipases and proteinases. Phospholipases are responsible for various aggressive and defensive actions on the host leading to tissue invasion and cell membrane remodelling. Proteinases contribute to pathogenesis by degrading or modifying target epithelial and mucosal barrier proteins and ligands on the yeast surface, thus allowing proper adherence of the fungus to host tissues5.
Candida cells adhere, colonize and form biofilms on medical implants such as blood and urinary catheters, dentures, artificial joints and voice prosthetics, causing high risk of infection in patients. Biofilm associated cells are much more resistant to existing antifungal drugs than planktonic cells due to complex biofilm structures, extracellular matrix and up-regulation of efflux pumps6, 7. Drug resistance associated with the biofilm mode of growth is multifactorial and involves production of persister cells, quorum sensing, production of extracellular matrix and drug binding and stress responses8, 9.
Plants and plant products have exhibited immense potential as antimicrobials10, 11. Terpenoids are a major class of natural compounds that have shown antifungal properties11-15. Geraniol, an acyclic monoterpene alcohol (Fig 1) is the major component of numerous essential oils like geranium oil, palmrosa oil, ninde oil, rose oil and citronella oil16. It possesses insecticidal, antihelminthic, antibacterial, antioxidant, anticancer and anti-inflammatory activities16-18 and is a good repellent for malarial, filarial and yellow fever vectors19. Recently, we showed that geraniol alters fungal cell morphology, significantly reduces ergosterol levels and inhibits glucose induced H+ efflux suggesting that it disrupts cell membrane integrity by interfering with ergosterol biosynthesis and H+ ATPase activity18. It has also been shown that geraniol causes mitochondrial dysfunction, impairs iron homeostasis and genotoxicity18, 20. This natural compound demonstrated negligible toxicity to human erythrocytes and hence is a good candidate to be studied as a potential antifungal drug. In the present study we have studied the effect of geraniol on hydrolytic enzymes secretion and biofilm formation as virulent factors on both sensitive and fluconazole (FLC)-resistant C. albicans strains along with standard strains of C. albicans, C. glabrata and C. tropicalis. To our knowledge the present study is the first one to show the effect of geraniol on the secretion of proteinases and phospholipases. These hydrolytic enzymes play a crucial role in facilitating adhesion, tissue damage and evasion of host immune responses21.
Strains and media
C. albicans (ATCC 10261 and ATCC 90028), C. glabrata ATCC 90030 and C. tropicalis ATCC 750 were the standard strains used in the present study. In addition, C. albicans D-27 (FLC sensitive) and C. albicans S-1 (FLC resistant) were isolated from vaginal swabs of diabetic patients obtained from Safdarjung Hospital, New Delhi, India. Strains showing MIC ³ 64 µg/ml for FLC were considered resistant. All the strains were maintained on YEPD (yeast extract 1%: peptone2%: dextrose 2%: agar 2.5%) plates at 4ºC. All the chemicals were of analytical grade and were procured from E. Merck (India). The media components were procured from Himedia (India) while geraniol was purchased from Aldrich (Germany).
Broth dilution method
Minimum inhibitory concentration (MIC) values of geraniol for the Candida strains were determined by the broth dilution method22. MIC was defined as the lowest concentration that causes 90% decrease in absorbance in comparison to that of the control (without test compound). After recording MIC values, 20 µl samples from all optically clear tubes (no growth) along with the last tube showing growth were sub-cultured on YEPD agar plates. The plates were incubated at 35ºC for a minimum of 3 days, until growth was clearly visible in the control samples, and MFC values were calculated as the lowest concentration of the test compound for which there was no visible growth23. Results were calculated as a mean of the two separate experiments done in triplicate.
Spot Assay
For studying susceptibility of geraniol to Candida cells24, cells were grown overnight in YEPD media at 37ºC. Cell culture was suspended into normal saline (optical density of 0.1 at 600 nm). For the assay, 5µl of fivefold serial dilution of each Candida strain was spotted on YEPD agar plates in the absence (control) and presence of ¼ MIC, ½ MIC, MIC, 2 MIC of geraniol. Difference in growth was observed after 48 hours of incubation at 30ºC. The solvent (1% DMSO) showed no effect on cell growth (data not shown).
In vitro Candida sensitivity to geraniol
Candida cells (105 cells/mL) were inoculated into YEPD agar at 40ºC and poured into 90-mm petri plates16. Sterile filter discs (4 mm) laden with the test compounds at different concentrations (3 MIC, 4 MIC and 5 MIC) were placed on agar plates. The diameter of inhibition zones (ZOI) was measured after 2 days. FLC (10 µg/mL) was also included as a positive control. The sensitivity index (SI) was calculated for all the three Candida species as ZOI (mm)/concentration of the antifungal compound (mg/mL). The SI values are shown as mean± SD25.
WST1 Cytotoxicity assay
Cell suspensions (1×105cells/ml) were cultured in 96-well plate with a final volume of 100µl/well along with appropriate concentrations of geraniol (≤ MIC) for 24 h followed by 2 h incubation at 37ºC with 10µl of WST- 1/CEC dye. Plates were gently shaken to mix the well constituents. The absorbance was recorded at 450 nm using a microplate Reader (BIORAD iMark, US) and reference was set at 655 nm. The absorbance values for culture medium were subtracted from the assay results. Each reading was taken in triplicate. Percentage cytotoxicity was calculated using the following equation:
% Cytotoxicity = [100 × (cell control – test value) ÷ (cell control)]
Proteinase assay
Candida cells were inoculated and incubated for 18 hrs at 37ºC in 5 ml YEPD media. Cells were centrifuged for 5 min at 3000 rpm and the pellets were re-suspended in saline, after washing twice for removing the residual culture medium. The cell suspensions, after standardization to a MacFarland 0.5 index, were exposed to desired concentrations of geraniol. After overlaying 2 µl on agar surface (agar 2%; BSA fraction V 0.2 g; yeast nitrogen base w/o amino acids; ammonium sulphate 1.45 g; glucose 20 g; sterile water to 1 L) at equidistant points, the plates were incubated for 2-3 days at 37°C. Proteinase secretion was estimated as Pz values, calculated by measuring the diameters of degradation zones and then by dividing the diameter of the colony by the diameter of the colony together with zone of clearance23, 26.
Phospholipase assay
Cell suspensions (MacFarland 0.5 index) were exposed to geraniol at MIC and sub-MIC and 2 µl pipette on agar peptone media (agar 2%, peptone 10g, glucose 30g, NaCl 57.3g, CaCl2 0.55g, distilled water added to 0.9 L) enriched with 10% (v/v) egg yolk emulsion (HiMedia). The plates were incubated for 2-4 days at 37°C and phospholipase secretion was estimated by measuring precipitation zones. Pz values were calculated as mentioned above 26, 27.
Adhesion assay and Biofilm formation
Cell suspension (100µl of 107cells/ml) in RPMI 1640 media was inoculated into each well of a sterile, 96-well microtiter plate28. Geraniol was added to each well at desired concentrations except in control. The plates were incubated at 37ºC for 90 min in a shaker at 75 rpm to allow adherence of cells. Each well was gently washed twice with PBS (150 µl) to remove non-adherent cells. The adherent cells were observed using Motic AE31 (Germany) Inverted microscope29. For biofilm formation, cell suspensions (1×107cells/ml in RPMI 1640) were inoculated into wells and incubated at 37°C for 1h. After the initial 1h adhesion period, fresh RPMI 1640 media, containing desired concentrations of geraniol were added to the plate wells containing adherent cells and incubated for 24 h at 37 °C. The adherent cells and biofilms were estimated using a semi-quantitative XTT reduction assay.
XTT- reduction assay
2,3-bis(2-methoxy-4-nitro-5-sulfo-phenyl)-2H-tetrazolium-5-carboxanilide (XTT) reduction assay is based on the reduction of XTT tetrazolium formazan by active mitochondria in Candida cells28. Briefly, pre-formed Candida biofilms were washed with PBS. XTT (300 mg/L) was added to 200 µl PBS. An aliquot of XTT (100µl) was then added to each pre-washed biofilms and control wells (to measure background XTT levels). The plates were incubated in dark for 5 h at 37°C without shaking. After 2min agitation the colorimetric change was observed at 450 nm (a direct correlation to metabolic activity of biofilms) using a microtiter plate reader (BIO-RAD, iMark, US). The wells containing PBS only or PBS plus XTT were used as blank. Biofilms were simultaneously examined using scanning electron microscopy (SEM).
Scanning Electron Microscopy
Post geraniol treatment, the biofilms formed as described above, were washed with PBS and fixed overnight in 2% glutaraldehyde (v/v) in PBS. The samples were washed twice in PBS and then placed in 1% osmium tetroxide for 30 min. Subsequently, the samples were dehydrated in a series of ethanol washes (70% for 10 min, 95% for 10 min, 100% for 20 min), and finally dried in a critical point dryer. The samples were then coated with gold/palladium (40%/60%). Biofilms were observed under Scanning electron microscope (ZEISS), following the given analytical conditions EHT = 3.00 kv, WD = 9.0mm, signal A = SE1, Mag = 3.07 K X.
Statistical analysis
All experiments in the present study were performed in triplicate and results were expressed as mean ± standard deviation (SD). Results were analysed employing Students t-test and P < 0.05 was considered statistically significant.
C. albicans and C. glabrata comprise nearly 70-80% of Candida species isolated from patients suffering from candidemia or invasive candidiasis. Among the non-albicans species, the highest mortality is linked with C. tropicalis and C. glabrata [30]. C. tropicalis, which comprises 6-12% of total Candida species, is apparently the most prevalent in India30 and is considered the causative agent of candidemia in cancer patients31. Hence these three Candida species were used to study the fungal virulence factors in the presence of geraniol (Fig. 1), a monoterpenoid alcohol, and one of the main components of several essential oils. In a previous study, we evaluated its anti-Candida potential and showed that it disrupts cell membrane integrity by interfering with ergosterol biosynthesis and H+-ATPase activity16. Here we further study the effect of this natural compound on secretion of hydrolytic enzymes (proteinases and phospholipases), adhesion and biofilm formation in both sensitive and resistant clinical strains of C. albicans and two other non-albicans Candida species.
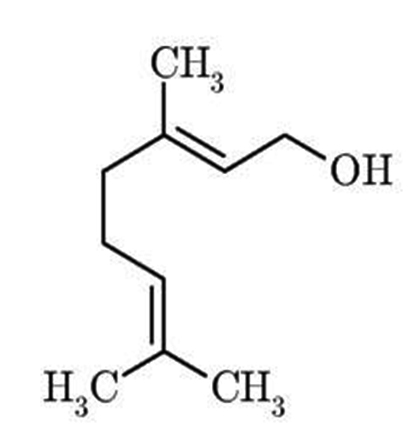
Fig. 1. Structure of geraniol, a monoterpenoid alcohol
Table 1 shows the MIC and MFC values of geraniol for the Candida strains studied here. The MIC of geraniol for C. albicans (ATCC 10261 and ATCC 90028) and C. glabrata (ATCC 90030) was 130 µg/ml while for C. tropicalis (ATCC 750) it was only 80 µg/ml. The corresponding MFC values for these standard strains were slightly higher – 160 µg/ml for C. albicans and C. glabrata, and 140 µg/ml for C. tropicalis. Interestingly, geraniol showed more sensitivity towards the clinical sensitive strains than the standard strains. FLC-sensitive C. albicans D-27 showed an MIC of 80 µg/ml while FLC-resistant C. albicans S-1 gave a higher value of 175 µg/ml. Although resistant Candida strains showed less sensitivity in comparison to standard and sensitive strains, still the antifungal potential was high enough to perform further studies. The MFC values were again only slightly higher with 140 µg/ml in FLC-sensitive and 250 µg/ml for FLC-resistant clinical strains of C. albicans. FLC, the conventional antifungal drug showed an MIC of 10 µg/ml for standard Candida strains. The value was 6 µg/ml for FLC-sensitive and 125 µg/ml for FLC-resistant C. albicans as shown in table 1.
Table (1):
Minimum inhibitory and fungicidal concentrations for geraniol against Candida strains.
| Candida strains | Type | MIC(µg/ml) | MFC (µg/ml) |
|---|---|---|---|
| C. albicans ATCC 10261 | Standard (American Type Cell Culture) | 130 | 160 |
| C. albicans ATCC 90028 | 130 | 160 | |
| C. glabrata ATCC 90030 | 130 | 160 | |
| C. tropicalis ATCC 750 | 80 | 140 | |
| C. albicans D-27 | Clinical FLC-sensitive | 80 | 140 |
| C. albicans S-1 | Clinical FLC-resistant | 175 | 250 |
Fluconazole (FLC) showed an MIC of 10 µg/mL for all standard strains, 6 µg/ml for FLC-sensitive and 125 µg/ml for FLC-resistant Candida strains
Antifungal susceptibility was also investigated on solid media by spot assay (Fig. 2) as one of the confirmatory methods. Candida cells were grown on YEPD agar for 48 h at 30ºC. Five spots in horizontal direction represent the serial dilution of cell culture. There is a significant reduction in the growth of Candida cells, which is concentration dependent. At MIC, the strains showed little or no proliferation at fourth dilution. C. albicans did not show any proliferation even at third dilution. No growth was observed in all the tested strains at 2 MIC, strongly recommending geraniol an antifungal agent. The results summarized in table 2 gives an estimate of the sensitivity of the strains for the antifungal compound. All three types of standard Candida strains showed sensitivity evident from zone of clearance (results not shown here). Sensitivity index (SI) was calculated as the ratio of the diameter of ZOI (mm) to the concentration of test compound (mg/ml). Sensitivity of geraniol increased with increasing concentration and C. tropicalis was more sensitive than C. albicans followed by C. glabrata. In all three cases sensitivity was less than that of FLC but the results are still encouraging as the test compound has low toxicity and is fungicidal in nature while FLC is fungistatic and leads to resistance.
Table (2):
In vitro Candida sensitivity to geraniol as determined by disc diffusion assay. Each strain was tested in triplicate. Sensitivity Index (SI) is expressed as mean ± SD and was calculated as the ratio of diameter of ZOI (mm) to concentration of test compound (mg/ml).
| Candida species | Test compound | Sensitivity Index (SI) |
|---|---|---|
|
C. albicans ATCC 10261 |
3 MIC | 2.2703± 0.017 |
| 4 MIC | 2.37± 0.06 | |
| 5 MIC | 2.52± 0.052 | |
| FLC (10 µg/ml) | 5.68± 0.06 | |
|
C. glabrata ATCC 90030 |
3 MIC | 1.935± 0.091 |
| 4 MIC | 2.12± 0.061 | |
| 5 MIC | 2.48± 0.080 | |
| FLC (10 µg/ml) | 4.866± 0.55 | |
|
C. tropicalis ATCC 750 |
3 MIC | 2.311± 0.062 |
| 4 MIC | 2.466±0.0353 | |
| 5 MIC | 2.65± 0.027 | |
| FLC (10 µg/ml) | 5.435±0.065 |
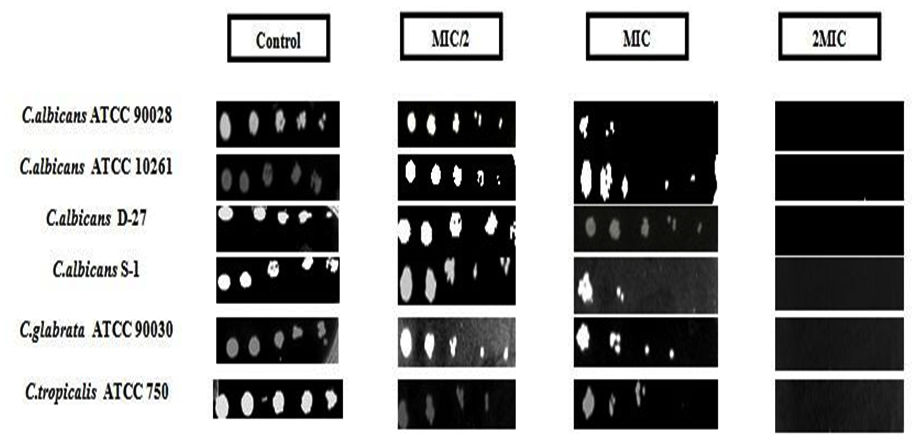
Fig. 2. Spot assay showing susceptibility of C. albicans (ATCC 90028, ATCC 10261, sensitive strain (D-27), resistant strain (S-1)), C. glabrata and C. tropicalis to vary-ing concentrations of geraniol. Control has no test compound
WST1 cytotoxic assay was performed to check yeast cell viability in the presence of geraniol, by a colorimetric method based on the principle of the metabolizing activity of mitochondrial succinic oxidase, cytochrome P-450 systems and flavoprotein oxidase of living fungal cells32, 33. This leads to the conversion of XTT to formazan, which is a water soluble product and hence can be easily measured in supernatants. The generation of the dark yellow coloured formazan is measured at 420-480 (optimal at 440 nm) and is directly correlated to cell number. This way biofilms can be studied as intact structures, and can be observed for drug susceptibility without disruption of their structures. Cells were cultured in a microplate and then incubated with the reagent WST-1 and the assay was monitored spectrophotometrically. It showed ≥90% cytotoxicity at MIC values for all tested strains (Fig. 3). This correlates well with the growth data presented in Fig 2. At concentrations of ½ MIC and ¼ MIC, average range of cytotoxicity was 68-88% and 30-45%, respectively in all three species. The above mentioned study revealed that geraniol shows high cytotoxic activity and affects the metabolic activities of yeast cells leading to decreased growth and viability, which can be explored with further studies.
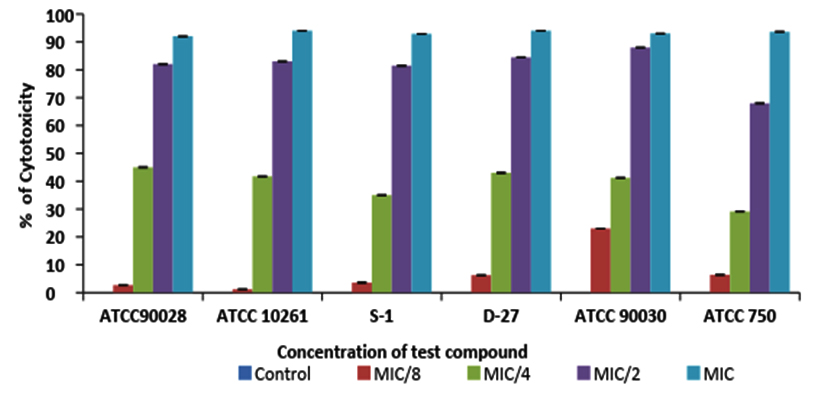
Fig. 3. The cytotoxic activity of geraniol against C. albicans (ATCC 90028 &10261), C. albicans D-27, C. albicans S-1, C. glabrata (ATCC 90030), C. tropicalis (ATCC 750). Error bar lines represents ± SD to the mean value from 3 different record-ings
Extracellular proteinases and phospholipases are important for the invasion of host tissues by Candida cells. Strains were tested for proteinase activity in the presence and absence of geraniol at MIC and sub-MIC concentrations (1/2 MIC, 1/4 MIC and 1/8MIC). Fig. 4 (a) shows degradation zones produced on solid media as a result of proteinase secretion by Candida species. At MIC, geraniol showed 32.5-38.7%, 46.51%, 31.83% proteinase inhibition in C. albicans (ATCC 10261 and ATCC 90028), C. tropicalis ATCC 750 and C. glabrata ATCC 90030 respectively. The clinical strains, C. albicansD-27 and S-1 showed 36.95% and 57.53% proteinase inhibition, respectively. At 1/2 MIC, this compound showed enzyme inhibition in the range of 30-40% for all tested strains (Fig 4b). Results showed that at ¼ MIC and 1/8 MIC, geraniol does not have any significant effect on secretion of proteinases. A decrease in secretion of proteinases under the effect of geraniol points towards the less virulent nature of C. albicans and non-albicans species in the presence of this natural compound. FLC-resistant isolates showed even lower secretory activity of proteinases in comparison to standard and sensitive strains, indicating high antifungal potential of geraniol in the resistant strain.
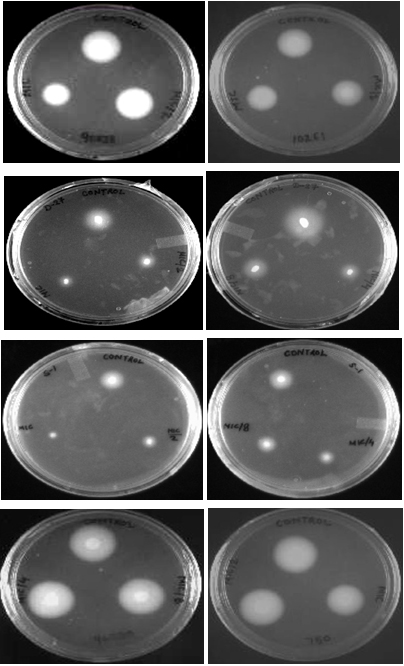
Fig. 4. (a) BSA agar plates showing the proteinase secretion in C. albicans ATCC 90028 and 10261 (A, B), C. albicans D-27 (C, D), C. albicans S-1 (E, F), C. glabrata ATCC 90030 (G), C. tropicalis ATCC 750 (H), in the presence of geraniol at 1/8 MIC, 1/4MIC, ½ MIC and MIC
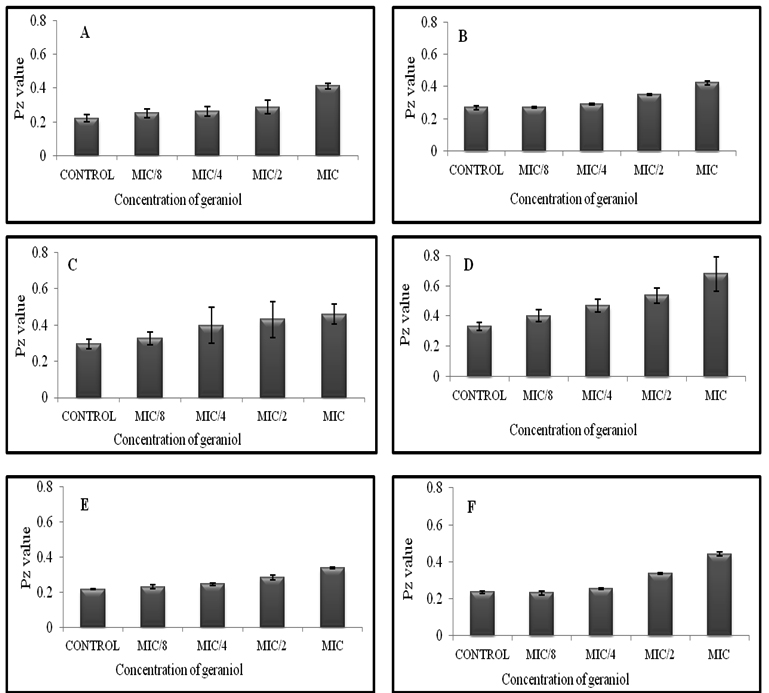
Fig. 4 (b). Effect of geraniol on proteinase secretion by C. albicans ATCC 90028 and 10261 (A and B), C. albicans D-27 (C), C. albicans S-1 (D), C. glabrata ATCC 90030 (E), C. tropicalis ATCC 750 (F). Errors bars represent ± SD to the mean value from 3 different readings
The secretory activity of phospholipases was not displayed by all the strains. Secretion was observed only in C. albicans ATCC 10261 and C. tropicalis ATCC 750 while C. glabrata ATCC 90030 failed to secrete this hydrolytic enzyme. Our results correlate with previously reported data, where screening of 41 Candida isolates showed phospholipase secretion in only 79% C. albicans, while none of the strains of C. glabrata, C. tropicalis, C. parapsilosis secreted the enzyme34. Hence, only secretory activity of C. albicans ATCC 90028, D-27 and S-1 have been shown here. Our data shows that the inhibitory effect of geraniol on phospholipase secretory activity was not as significant as on proteinase secretion. At MIC, geraniol showed 10%, 20% and 33% reduction in phospholipase secretion in case of C. albicans ATCC 90028, C. albicans S-1 and C. albicans D-27 respectively (Fig 5). The percentage inhibition for C. albicans ATCC 10261 was 15-21% (Figure not shown). Secretion of phospholipases is not significantly inhibited in comparison to proteinases, which may be due to the reason that geraniol has inhibitory effect on SAP genes rather than PLB genes1.
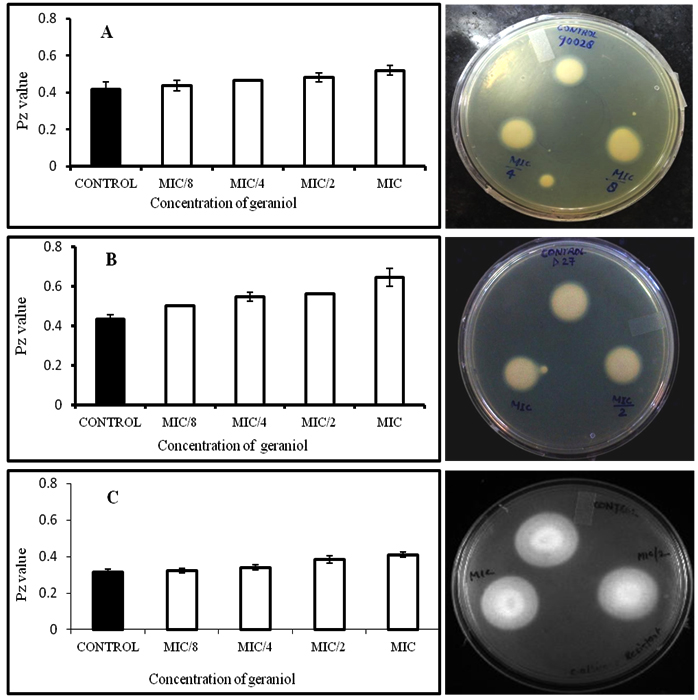
Fig. 5. Effect of geraniol on phospholipase secretion in C. albicans ATCC 90028 (A), C. albicans D-27 (B), C. albicans S-1 (C) in the presence of different concentrations of geraniol (1/8 MIC, ¼ MIC, ½ MIC and MIC)
Fungal cell adherence to host tissues is the initial step crucial to the process of biofilm formation. The amount and the characteristics of biofilms were microscopically studied with the semi-quantitative colorimetric technique based on XTT-reduction assay. Microscopic analysis and colorimetric readings revealed that the formation of hyphae and biofilms was inhibited by geraniol in a concentration dependant manner. Fig 6 shows microscopic pictures of cell adherence on polystyrene surface. The control (6A) shows a large number of adherent cells in comparison to the treated case (6B). More than 60% microbial infections in humans are biofilm related, where C. albicans biofilm is a major virulence factor responsible for high risk of mortality in some patients. Chronic biofilms facilitate carcinogenesis by causing inflammation35 and chronic mucocutaneous candidiasis is a risk for oral cancer36. The present study shows that adherence of C. albicans cells to polystyrene surface was significantly inhibited by geraniol. Adherence was inhibited by 45%, 38%, 52% and 44.6% in C. albicans ATCC 90028, C. albicans D-27 and C. albicans S-1 at their respective MIC values. At 1/2 MIC, inhibition was 20.54%, 33.72 and 27.4 % in the same order. In case of C. albicans ATCC 10261, cell adherence was inhibited by 50%, 30 % and 33 % at MIC, 1/2 MIC and 1/4 MIC, respectively (Fig. 7).
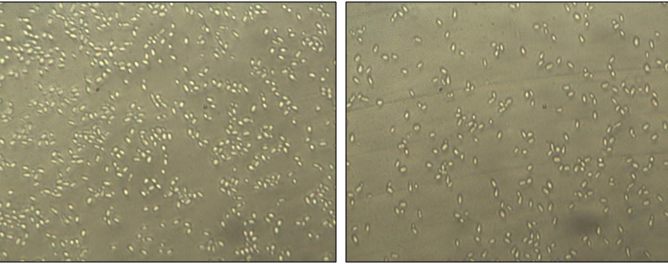
Fig. 6. Inverted microscopic pictures of adhesion, showing the decrease in number of adherent C. albicans (ATCC 10261) cells treated with MIC of geraniol (B), while cells in control (A) show a large number of adherent cells in the absence of geraniol
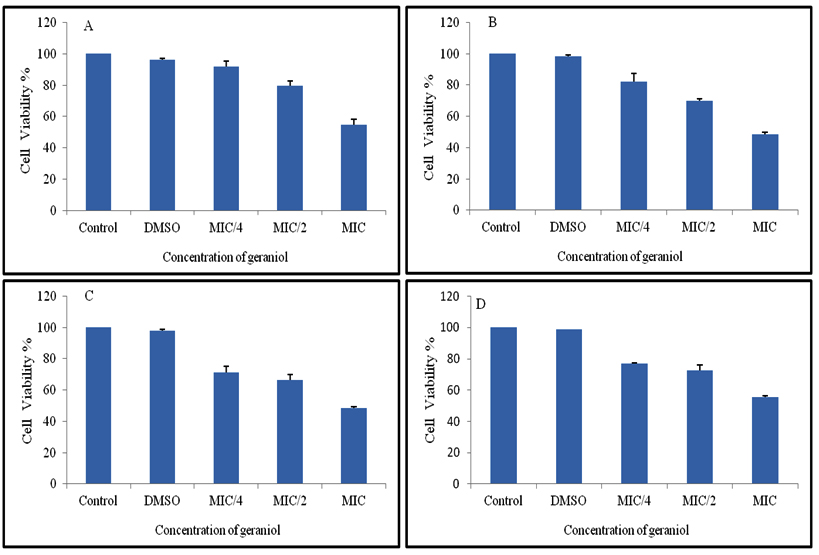
Fig. 7. Effect of varying concentrations of geraniol (MIC, ½ MIC, ¼ MIC) on adhesion in C. albicans ATCC 90028 (A), C. albicans ATCC 10261 (B), C. albicans FLC sensitive D-27 (C) and FLC resistant C. albicans S-1(D ). Cell viability was estimated using the XTT reduction assay after 1h incubation. 1% DMSO was also used as a control
Biofilms formed by C. albicans are highly structured and composed of round, budding yeast cells along with pseudohyphae and hyphae encased within an extracellular matrix. Their formation compromises of four different stages that include adherence to host surfaces. This is the initial stage followed by proliferation and growth of pseudohyphae and hyphae. Finally the yeast cells disperse to seed biofilms to new sites in the host17. C. albicans associated biofilms are difficult to treat due to acquired resistance for existing antibiotics. Hence, a search for novel compounds against fungal biofilms is a big necessity now and natural compounds are good candidates as they are non-toxic, easily available and do not lead to multidrug resistance. Majority of biofilms are commonly formed by C. albicans rather than non-albicans Candida species37. XTT reduction by the metabolic activity of cells is used to study the inhibitory activity of geraniol on biofilms. Exposure to geraniol at MIC, caused inhibition in biofilm formation of 71.8%, 75-79% and 63% respectively in C. albicans ATCC 90028, C. albicans D-27 and C. albicans S-1. Biofilm inhibition in the same strains at lower concentration of 1/2 MIC was ≤55%, ≤58% and 43%.
C. albicans ATCC 10261 showed 85% inhibition at MIC of geraniol followed by ½ MIC (79) and ¼ MIC (60%) (Fig.8). C. albicans D-27 (FLC-sensitive) and C. albicans ATCC 10261 showed higher sensitivity towards adherence as well as biofilm formation when treated with varying concentration of geraniol. Inhibitory effect of geraniol on cell adherence and biofilm formation was dose dependent. Significant reduction in biofilm formation may be due to the inhibition of yeast to hyphal transition. The morphological transition provides dense structural integrity to biofilm complexes. Geraniol may be responsible for reducing cell adherence which is crucial for biofilm formation. Previous studies have also linked inhibition of yeast to hyphal transition to anti-biofilm activity in C. albicans18. The contribution of yeast to hyphal transition is significant in the formation of biofilms and hence is responsible for the virulence and pathogenicity of this opportunistic fungus.
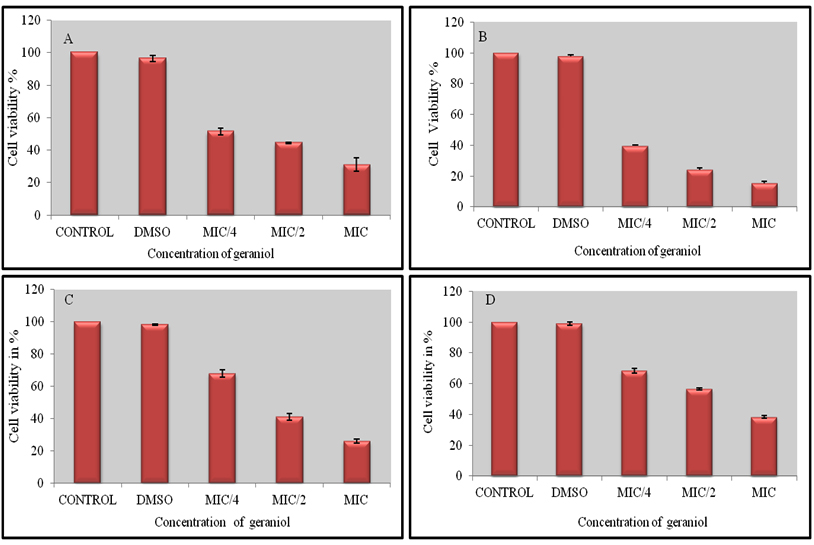
Fig. 8. The effect of varying concentrations of geraniol (MIC, ½ MIC and ¼ MIC) on biofilm formation in C. albicans ATCC 90028 (A), C. albicans ATCC 10261 (B), C. albi-cans FLC sensitive D-27 (C) and FLC resistant C. albicans S-1(D ). Cell viability was estimat-ed using the XTT reduction assay after 24 h incubation. 1% DMSO (control) had no effect on biofilms. Errors bars represent ± SD to the mean value from 3 different values

Fig. 9. Scanning electron micrographs of C. albicans ATCC 10261 biofilms grown for 24h on silicon discs in the absence (A) and presence (B) of geraniol at MIC. In the absence of geraniol a dense population of yeast cells and hyphae could be seen while in the presence, a loss in the biofilm structural elements could be observed
Figure 9 shows SEM visualization of biofilm formation by C. albicans on plastic cover slips. Cells treated with geraniol at MIC produced less dense biofilms, which were composed mainly of yeast cells and pseudohyphae. Poor biofilm architecture was observed in treated cells due to absence of true hyphae (8B). The control biofilms (8A) formed in the absence of geraniol were denser and constituted mainly true hyphae and pseudohyphae forming a fairly good architecture. The data was consistent with biofilm results estimated by XTT reduction assays.
In conclusion, our work demonstrates that geraniol exhibits potent antifungal activity against Candida species, inhibits biofilm formation, reduces cell adherence and decreases the secretion of hydrolytic enzymes especially proteinases. Results clearly suggest that geraniol is clinically significant and may be used in treating biofilm-associated Candida infections. Further research is required both in vivo and in vitro to validate the diverse mode of antifungal action of geraniol as a promising antifungal agent.
ACKNOWLEDGMENTS
Yamini Sharma is thankful to ICMR, New Delhi for ICMR-SRF fellowship reference no- 45/55/2014-PHA/BMS.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
- Ghannoum M A. Potential Role of Phospholipases in Virulence and Fungal Pathogenesis. Clin Microbiol Rev., 2000; 13 (1): 122-143.
- Sardi JCO, Scorzoni L, Bernardi T, Fusco-Almeida AM, Mendes Giannini MJS. Candida species: current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J Med Microbiol., 2013; 62 (1): 10-24.
- Chakrabarti A, Chatterjee SS, Rao KL, Zameer MM, Shivaprakash MR, Singhi S, Singh R, Varma SC. Recent experience with fungaemia: change in species distribution and azole resistance. Scand J Infect Dis., 2009; 41: 275–284.
- Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence 2013; 4 (2): 119–128.
- Monod M, Borg-Von Zepelin M. Secreted proteinases and other virulence mechanisms of Candida albicans. Chem Immunol., 2002; 81: 114-28.
- Gulati M, Nobile CJ. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes and Infection., 2016; 18: 310-321.
- Ramage G, Vandewalle K, Wickes BL, Lopez-Ribot JL. Characteristics of biofilm formation by Candida albicans. Rev Iberoam Micol., 2001; 18: 163 –170.
- La Fleur MD, Kumamoto CA, Lewis K. Candida albicans biofilms produce antifungal-tolerant persister cells. Antimicrob Agents Chemother., 2006; 50: 3846-3839.
- Taff HT, Mitchell KF, Edward JA, Andes DR. Mechanisms of Candida biofilm drug resistance. Future Microbiol., 2013; 8: 1325–1337.
- Cowan MM. Plant products as antimicrobial agents. Clin Micro-biol Rev., 1999; 12: 564-82.
- Zore GB, Thakre AD, Jadhav S, Karuppayil SM. Terpenoids inhibit Candida albicans growth by affecting membrane integrity and arrest of cell cycle. Phytomedicine., 2011; 18: 1181-90.
- Vengurlekar S, Sharma R, Trivedi P. Efficacy of some natural compounds as antifungal agents. Pharmacogn Rev., 2012; 6 (12): 91–99.
- Ahmad A, Khan A, Akhtar F, Yousuf S, Xess I, Khan LA, Manzoor N. Fungicidal activity of thymol and carvacrol by disrupting ergosterol biosynthesis and membrane integrity against Candida. Eur J Clin Microbiol Infect Dis., 2011; 30: 41 – 50.
- Ahmad A, Khan A, Manzoor N. Reversal of efflux mediated antifungal resistance underlies synergistic activity of two monoterpenes with fluconazole. Eur J Pharm Sci., 2013; 48: 80-86.
- Khan A, Ahmad A, Manzoor N, Khan LA. Antifungal activities of Ocimum sanctum Essential Oil and its Lead Molecules. Nat Prod Commun., 2010; 5 (2) 345-349.
- Sharma Y, Khan LA, Manzoor N. Anti-Candida activity of geraniol involves disruption of cell membrane integrity and function. J Mycol Med., 2016; 26: 244-254.
- Nobile CJ, Johnson AD. Candida albicansbiofilms and human disease. Annu Rev Microbiol., 2015; 69: 71–92.
- Singh S, Fatima Z, Hameed S. Insights into the mode of action of anticandidal herbal monoterpenoid geraniol reveal disruption of multiple MDR mechanisms and virulence attributes in Candida albicans. Arch Microbiol., 2016; 198 (5): 459–472.
- Maia MF, Moore SJ. Plant-based insect repellents: a review of their efficacy, development and testing. Malar J., 2011; 10: S11.
- Leite MC, de BritoBezerra AP, de Sousa JP, de Oliverira Lima E. Investigating the antifungal activity and mechanisms of geraniol against Candida albicans strains. Med Mycol., 2015; 53: 275-84.
- Yang YL. Virulence factors of Candida species. J Microbiol Immunol Infect., 2003; 36: 223-228.
- CLSI-Clinical and Laboratory Standards Institute, 2008. Reference method for broth dilution antifungal susceptibility testing of yeast, Approved Standard M27-A3, Clinical and Laboratory Standards Institute Standards, Wayne, PA, USA, 40 pp.
- Samber N, Khan Amber, Varma A, Manzoor N. Synergistic anti-Candidal activity and mode of action of Mentha piperita essential oil and its major components. Pharm Biol., 2015; 53 (10): 1496-504.
- Ansari MA, Fatima Z, Hameed S. Sesamol: a natural phenolic compound with promising anticandidal potential. J Pathog., 2014; 895193: 1–12.
- Ahmad A, Khan A, Khan LA, Manzoor N. In vitro synergy of eugenol and methyl eugenol with fluconazole against clinical Candida isolates. J Med Microbiol., 2010; 59: 1178-1184.
- Price MF, Wilkinson ID, Gentry LO. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia., 1982; 20: 7-14.
- Samber N, Varma A, Manzoor N. Evaluation of Mentha piperita essential oil and its major constituents for antifungal activity in Candida spp. Int. J. Innov. Res. Sci. Eng., 2014; 3 (2): 9404-9411.
- Ramage G, Saville SP, Wickes BL, Lopez-Ribot JL. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl Environ Microbiol., 2002; 68: 5459-5463.
- Khan A, Ahmad A, Xess I, Khan LA, Manzoor N. Ocimum sanctum essential oil inhibits virulence attributes in Candida albicans. Phytomedicine., 2014; 21: 448-452.
- Kothavade RJ, Kura MM, Valand AG, Panthaki MH. Candida tropicalis: its prevalence, pathogenicity and increasing resistance to fluconazole. J Med Microbiol., 2010; 59: 873-80.
- Vazquez JA, Sobel JD. 2003. Candidiasis, pp 143–87. In Dismukes WE, Pappas PG, Sobel JD. (eds.) Clinical Mycology, Oxford University Press, New York.
- Kuhn DM, Balkis M, Chandra J, Mukherjee PK, Ghannoum MA. Uses and limitations of the XTT assay in studies of Candida growth and metabolism. J Clin Microbiol., 2003; 41 (1): 506-508.
- Altman FP. Tetrazolium salts and formazans. Prog. Histochem. Cytochem., 1976; 9: 1-56.
- Samaranayake LP, Raeside JM, MacFarlane TW. Factors affecting the phospholipase activity of Candida species in vitro. Sabouraudia., 1984; 22: 201-207.
- Coussens LM, Werb Z. Inflammation and cancer. Nature., 2002; 420: 860–867.
- Marttila E, Bowyer P, Sanglard D, Uittamo J, Kaihovaara P, Salaspuro M, Richardson M, Rautemaa R. Fermentative 2-carbon metabolism produces carcinogenic levels of acetaldehyde in Candida albicans. Mol Oral Microbiol.,2013; 28 (4): 281-91.
- Kuhn DM, Chandra J, Mukherjee PK, Ghannoum MA. Comparison of Biofilms Formed by Candida albicans and Candida parapsilosis on Bioprosthetic Surfaces. Infect Immun., 2002; 70 (2):878-888.
© The Author(s) 2018. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.