


ISSN: 0973-7510
E-ISSN: 2581-690X
Tomato cultivation faces significant challenges from foliar fungal diseases such as early blight caused by Alternaria spp., resulting in substantial reduction in yield. In this study, we examined the suppressive effects of the antagonistic fungus Clonostachys rosea on early blight of tomato under controlled experimental conditions. The results of dual culture experiments revealed the inhibitory effects of five C. rosea isolates on Alternaria spp., with the TNAU CR04 isolate exhibiting the highest inhibition (77.22%). Scanning electron microscopy provided good insight into antagonistic effects of C. rosea against A. alternata, revealing hyphal interactions and structural alterations. Further investigations focused on the suppression of Alternaria alternata mycelial growth (86.78%) by culture filtrates of C. rosea. The results revealed that TNAU CR04 at a 50% concentration strongly inhibited mycelial growth. Through GC-MS analysis, we identified key compounds involved in the interaction between C. rosea TNAU CR04 and A. alternata, shedding light on metabolic pathways and defense mechanisms. Overall, this study showed that C. rosea and its metabolites strongly act against Alternaria spp., revealing its mode of action and mechanisms underlying disease suppression in tomato plants.
Plant Pathogenic Fungi, Antifungal Activity, Metabolites, Pathways, Interaction, Defense
Tomato (Solanum lycopersicum L.) is a commonly cultivated vegetable from the Solanaceae family. It is a major crop grown extensively in China, India, the United States, Turkey and Italy. China ranks as the top producer, whereas United States and Mexico are key players in export markets. Its adaptability to both temperate and tropical regions makes it a crop of global importance. It is considered to be a crucial “protective food” because of its rich nutritional profile.1 Various biotic obstacles including viruses, oomycetes, bacteria, fungi and root-knot nematodes limit the yield of tomatoes. Early blight induced by Alternaria solani and Alternaria alternata is one of the most harmful and prevalent diseases among the numerous biotic factors that degrade tomato quality and yield globally.2 Due to its detrimental effects, this pathogen has received significant attention over the years, primarily because of the substantial yield loss of about 80%.3 To manage early blight disease, farmers predominantly depend on fungicides. However, the excessive use and reliance on these chemicals have sparked concerns regarding food safety, resistance to pathogens, degradation of soil and environmental sustainability. Hence, innovative disease management strategies that promote sustainable agriculture without compromising the environment need to be developed.4 Various mycoparasitic fungi can counteract Alternaria species in plants5,6 and their utilization as biological control agents against foliar diseases induced by Alternaria is underexplored and needs further investigation.
Clonostachys rosea (Gliocladium roseum)7 was initially identified as the anamorph of Bionectria ochroleuca, a teleomorph based on its morphology and supported by DNA sequence data. In temperate and tropical regions, the Clonostachys genus harbors predatory fungi that exhibit versatile lifestyles and thrive as soilborne, saprophytic and endophytic organisms, with potential implications for ecological dynamics and agricultural practices. Antagonistic isolates of C. rosea are used as biological control measures against various significant foliar plant diseases, including gray mold (Botrytis cinerea), gray leaf spot in maize (Cercospora zeae-maydis),8 leaf blotch in barley (Bipolaris sorokiniana),9 leaf spots (Alternaria spp.) and various other pathogens.5 While engaging in antagonistic mechanisms, isolates of C. rosea generate and release antimicrobial compounds and cell wall-degrading enzymes.10,11 These fungi can also strongly tolerate pathogenic toxins and chemical fungicides, probably due to the presence and activity of numerous membrane-associated transporters.12
The effectiveness of C. rosea strains in terms of biocontrol activity has been widely established, as indicated by their ability to generate a broad spectrum of metabolic compounds. These include enzymatically active proteins, small molecules linked with fungal or plant cell walls, and various secondary metabolites. Moreover, these metabolites can stimulate the defensive systems of plants, supporting their resilience against infections.13 Therefore, this study was performed to evaluate the effects of C. rosea on the development of leaf spots on tomatoes caused by Alternaria spp. under laboratory conditions. This study also involved metabolomic analyses to identify key metabolites and pathways of C. rosea involved in pathogen inhibition.
Isolation, identification and characterization of Alternaria spp.
Tomato leaves showing characteristic early blight infections were collected from several major tomato cultivation areas in Tamil Nadu, India. After confirming the presence of fungal spores, isolation was performed and Alternaria spp. were isolated from infected leaves using the tissue segment method. The pathogen was identified by morphological methods and sequencing of the ITS region. The virulence of these strains was subsequently tested and confirmed through Koch’s postulates. The most virulent isolate in pathogenicity studies was selected for assessing antagonistic activity. Following purification on PDA media, fungal cultures were stored at 4°C for subsequent studies.
Collection of biocontrol organisms
Five strains of C. rosea, were designated as TNAU CR01 (Accession No.: OK147890), TNAU: CR02 (Accession No.: ON926972), TNAU: CR03 (Accession No.: ON926975), TNAU: CR04 (Accession No: ON926968) and TNAU CR05 (Accession No.: ON926986), which were characterized by Gowrisri et al. were acquired from Culture Collection Centre, Department of Plant Pathology, TNAU, Coimbatore, Tamil Nadu, India. These isolates were then grown on PDA media at 27°C for seven days, and the actively growing colonies were stored at 4°C for future experiments.
Effect of Clonostachys rosea on the growth of Alternaria spp. in vitro
Dual culture technique
The ability of C. rosea to suppress pathogen growth was recorded in vitro using a dual culture technique. A 6 mm mycelial disc was excised from the rapidly growing edges of 12-day-old cultures of C. rosea using a sterilized cork borer. The mycelial disc was inoculated onto a sterilized PDA medium placed 1 cm away from the edge of the Petri dish. Concurrently, the fungal mycelial disc of the pathogen was placed at the far end of the Petri dish, opposite the C. rosea disc. Petri plates solely grown with the pathogen were used as controls. The treatments were replicated four times and the plates were kept at 28 ± 2°C. The inhibition caused by C. rosea was assessed by calculating the percentage (%) of mycelial growth inhibition using the formula described by Rapilly (1968).14
% I = [(C-T)/C] X 100)
Here,
I= percentage inhibition of pathogen by antagonist
C= radial growth of the pathogen in the control (mm)
T= radial growth of the pathogen in the treatment (mm)
Scanning electron microscopy
We performed SEM to observe hyphal interactions between C. rosea and Alternaria spp. First, fungal disks (5 mm) of both microorganisms were grown on PDA for 8-10 days. These cultures were observed under a light microscope to look at the early contact stage, after which 1 cm agar blocks containing the labeled mycelial interactions were excised for SEM sample preparation. Control blocks were collected from the two peripheries of the blocks comprising the antagonist and the pathogen and these blocks were fixed with osmium tetroxide. Images were captured using a FAI QUANTA 250 Model SEM at 15 kV in the Department of Nanotechnology, TNAU, Coimbatore, Tamil Nadu, India. Through SEM imaging, the mycelial growth of C. rosea over Alternaria spp. was documented.
Preparation of crude culture filtrates of Clonostachys rosea
To prepare crude culture filtrates, a 5 mm disc from a 12-day culture of C. rosea grown on PDA medium was added to Erlenmeyer flasks containing 100 mL of potato dextrose broth (PDB). The conical flasks containing culture broth were then kept at 28 ± 2°C for 15 days. The fungal extract was acquired through filtration using Whatman filter paper No. 4. After filtration, the fungal filtrate was stored at 4°C for further use.
Poisoned food technique
To evaluate the in vitro inhibition of Alternaria spp. mycelium, the filtrates of C. rosea were mixed with PDA medium. PDA plates were augmented with culture filtrate (CF) from C. rosea, which consisted of 20 ml of PDA per Petri dish at various percentages (20%, 30%, 40% and 50%). Next, the plates were inoculated with a 5 mm mycelial disc from the periphery of a seven day-old Alternaria spp. culture. Each treatment was repeated four times. For comparison, control plates devoid of culture filtrate suspension were also included. The plates were incubated at 28°C for 7-10 days until the pathogen reached the periphery of the control plate. The colony diameter of each plate was subsequently recorded and compared to that of the control.
Metabolomic analysis of antagonist, pathogen and their interactions
Crude culture metabolites from the potent strain of C. rosea. TNAU CR04 which presented the greatest reduction in mycelial development of the pathogen was selected from dual culture to determine the non-volatile organic compounds or metabolites responsible for suppressing Alternaria spp. PDB broth was inoculated with the respective antagonist and pathogen to determine their interaction. The control broth was also maintained with only the antagonist or pathogen. Cultures were incubated at 28°C for 12-15 days, followed by filtration through two layers of filter paper. An equal volume of ethyl acetate was added in to the extracted culture filtrate, and the mixture was incubated overnight in a shaker at 150 rpm. The ethyl acetate fraction was then separated using a separating funnel. The collected upper phase was evaporated under a rotary evaporator and dried. The dried residue was dissolved in 1 mL of HPLC-grade methanol and subjected to GC-MS analysis using a Perkin Elmer Clarus SQ 8C gas chromatograph-mass spectrometry equipment. The primary compounds were detected using of a computer-based method, and their mass spectrum was checked using the NIST library. Principal component analysis (PCA) was performed and a heat map was constructed for metabolite distribution patterns using R software. Metabolite set enrichment analysis and network-based analysis were conducted out using Metaboanalyst 6.0 and Cytoscape 3.3, respectively.
Statistical analysis
The data were statistically analyzed using R software version 2.14.1, where an ANOVA was conducted and the mean values were compared by Duncan Multiple Range Test.
Isolation, morphological and molecular identification of the pathogen
Alternaria spp. were isolated from diseased tomato leaves collected from various regions of Tamil Nadu, and their pure culture was maintained on PDA media. The characterization of these isolates revealed predominantly gray or brown colonies with consistent growth patterns (Figure 1a). On the underside of the same medium, the colonies displayed central black coloration surrounded by a hazy yellowish-brown rim and white margins (Figure 1b). Microscopic examination revealed the presence of solitary conidia, varying from straight to slightly flexuous, muriform or ellipsoidal with a tapered beak (Figure 1c). These conidia were generally pale and occasionally branched. The pathogenic fungal isolates were confirmed by ITS sequence analysis. A 531 bp band was amplified with ITS primers, and sequence alignment was performed using the Basic Local Alignment Search Tool (BLAST) to assess the phylogenetic relationships of the strains. The fungal strains showed 100% identity with both A. alternata and A. solani. The ITS sequences of A. alternata and A. solani were submitted to GenBank under the accession numbers PP763705, PP763571, PP760124, PP767753, PP767412 and PP784700. In pathogenicity studies, the strain A. alternata (CBE Alt1- PP767412) was found to be more virulent, leading to its selection for further antagonistic assays.
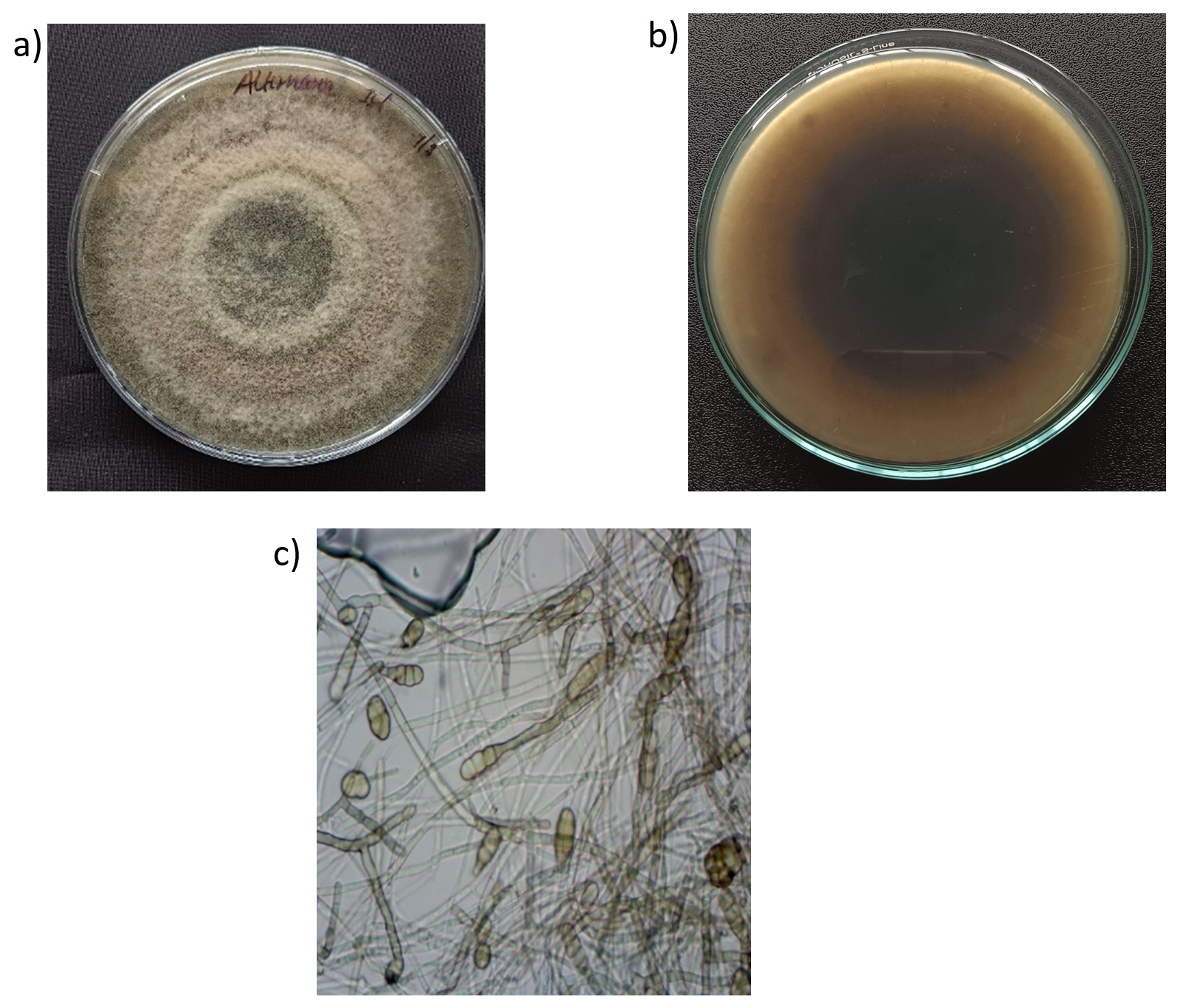
Figure 1. (a) Pure culture of A. alternata (CBE Alt 1) grown on PDA medium, displaying characteristic concentric rings; (b) Production of dark pigmentation by the pure culture of A. alternata on the back side of the PDA medium; (c) Microscopic view of A. alternata showing septate hyphae and muriform conidia
Efficacy of Clonostachys rosea isolates on Alternaria spp. in vitro (dual culture)
A dual culture assay was performed to determine the ability of C. rosea to suppress A. alternata CBE Alt1 growth in vitro. Five antagonistic isolates were tested against the early blight pathogen A. alternata. All C. rosea isolates inhibited the growth on A. alternata, with the C. rosea isolate TNAU CR04 showing the highest inhibition rate of 77.22%, followed by TNAU CR05, TNAU CR03 and TNAU CR01 with inhibition rates of 69.00%, 57.78% and 53.18% respectively. The lowest inhibitory effect was recorded for the C. rosea isolate TNAU CR02 (50.33%) (Table 1 and Figure 2). Antifungal testing confirmed that C. rosea can hinder the growth of A. alternata, with TNAU CR04 being the most effective. Consequently, TNAU CR04 was selected for further in vitro and in vivo studies.
Table (1):
Antagonistic effect of C. rosea on the mycelial growth of A. alternata
Isolate |
Radial mycelial growth (mm) |
Mycelial inhibition % |
|---|---|---|
TNAU CR01 |
42.300d |
53.18d (46.82) |
TNAU CR02 |
44.667e |
50.33e (45.18) |
TNAU CR03 |
37.967c |
57.78c (49.47) |
TNAU CR04 |
23.267a |
77.22a (59.48) |
TNAU CR05 |
27.900b |
69.00b (56.17) |
Control |
90.00f |
0.00f (0.28) |
CD (p = 0.05) |
1.775 |
0.879 |
SED |
0.814 |
0.403 |
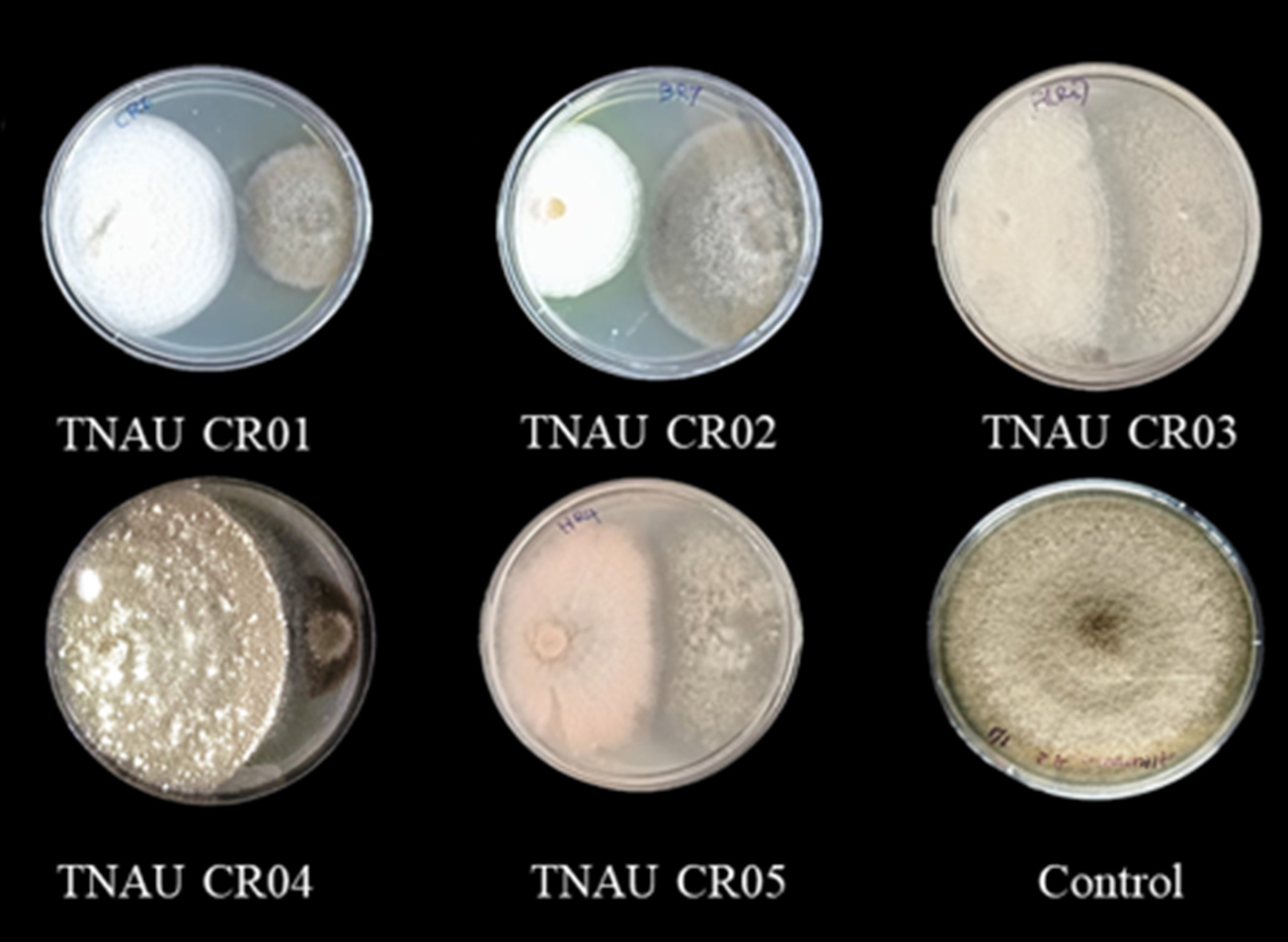
Figure 2. In vitro antagonistic potential of C. rosea isolates against A. alternata in dual-culture assay
Scanning electron microscopy
The mycoparasitic behavior of C. rosea TNAU CR04 toward A. alternata in dual culture was investigated via SEM. For observation, mycelial samples were scraped from the interaction zone of the dual plate. The hyphae of the antagonistic fungus C. rosea formed dense coils and tightly encircled the hyphae of A. alternata. This encircling led to a wrinkled appearance or collapse of the A. alternata hyphae. The hyphae of pathogen and the mycoparasitic hyphae were attached longitudinally. We also recorded several other structures, including hyphal depressions, hooks, small contact branches and coils in the interaction region (Figure 3).

Figure 3. SEM analysis of mycoparasitic C. rosea TNAU CR04 interacting with hyphae of A. alternata (a) Mycelia of C. rosea growing over A. alternata (b) Mycelial shrinkage and disintegration of mycelia of A. alternata (c) Conidiogenous cells with conidia of C. rosea emerging from hyphae of A. alternata (d) Control- hyphae of A. alternata
Mycelial growth inhibition by culture filtrates of Clonostachys rosea TNAU CR04
Poisoned food technique
The effective C. rosea isolate TNAU CR04 was also studied to determine the potential of its non-volatile compounds. The percentage inhibition of the pathogenic isolate (CBE Alt1- PP767412) was calculated and compared to their full growth on control plates. Evaluation of these non-volatile components at different concentrations revealed different inhibitory activities against growth of pathogenic mycelia. TNAU CR04 at a 50% concentration of the culture filtrate exhibited the most potent inhibition against Alternaria spp., with an inhibition rate of 86.78% followed by the 40% concentration (85.11%). The culture filtrates at a concentration of 20% presented the lowest inhibition of about 71.89% (Table 2 and Figure 4).
Table (2):
Inhibition rate of culture filtrates of Clonostachys rosea TNAU CR04 against Alternaria alternata by poisoned food technique
Culture filtrate concentrations (%) |
Radial mycelial growth(mm) |
Mycelial inhibition% |
|---|---|---|
20% |
27.12d |
71.89c (57.982) |
30% |
19.73c |
78.11b (62.104) |
40% |
13.46b |
85.11a (67.36) |
50% |
11.90a |
86.78a (68.72) |
Control |
90.00f |
0.00d (0.286) |
CD (p = 0.05) |
1.345 |
2.372 |
SED |
0.604 |
1.31 |

Figure 4. In vitro efficacy of C. rosea TNAU CR04 culture filtrates on mycelial growth inhibition of A. alternata
Profiling of secondary metabolites of Clonostachys rosea TNAU CR04 and its interaction with Alternaria alternata
GC-MS analysis of non-volatile secondary metabolic compounds in C. rosea TNAU CR04 revealed the presence of formic acid, 2(3H)-furanone, Di-n-octyl phthalate and 5-hexadecenoic acid. These compounds were also detected during the interaction between A. alternata and C. rosea. However, non-volatile metabolites including dl-glyceraldehyde dimer, 5-hydroxymethylfurfural, acetidine and cyclopropanecarboxamide were exclusively detected during the association between C. rosea and A. alternata, and not when C. rosea was present alone. The heat map (Figure 5) delineated the presence or absence of compounds, as well as their upregulation and downregulation, highlighting interactions among them. The gradation from blue (low) to red (high) in the heat map visually represented the relative intensity of the measured compounds. Metabolite enrichment analysis revealed that specific pathways were upregulated during the interaction between antagonist and pathogen. When grown alone (Figure 6a), C. rosea prioritizes phenylalanine metabolism, a critical pathway for producing phenylpropanoids, which can enhance its biocontrol potential through the synthesis of antifungal compounds. During the interaction with A. alternata (Figure 6b), C. rosea exhibited a broader metabolic reconfiguration. The significant enrichment in butanoate and glycerolipid metabolism indicated a shift toward lipid and membrane-related processes, which may be critical for maintaining cellular integrity under the stress of fungal antagonism. Pathway maps with network diagrams were used to assess the relevance of the key pathways, providing insights into the significance of the results. The compounds present in the TCA cycle and the synthesis of terpenes are shown in Figure 7. Key enzymes such as citrate synthase, aconitase and malate dehydrogenase indicate active energy metabolism through the TCA cycle, whereas enzymes in the terpene biosynthesis pathway suggest the production of terpenoids, possibly for antifungal defense. Additionally, the presence of ATP-producing enzymes and redox-related enzymes such as NADH dehydrogenase, indicates the role of energy production and redox reactions in supporting this metabolic interaction. Overall, the analysis highlighted the metabolic coordination of C. rosea in utilizing both primary and secondary metabolic pathways during its interaction with A. alternata, likely contributing to its biocontrol efficacy.
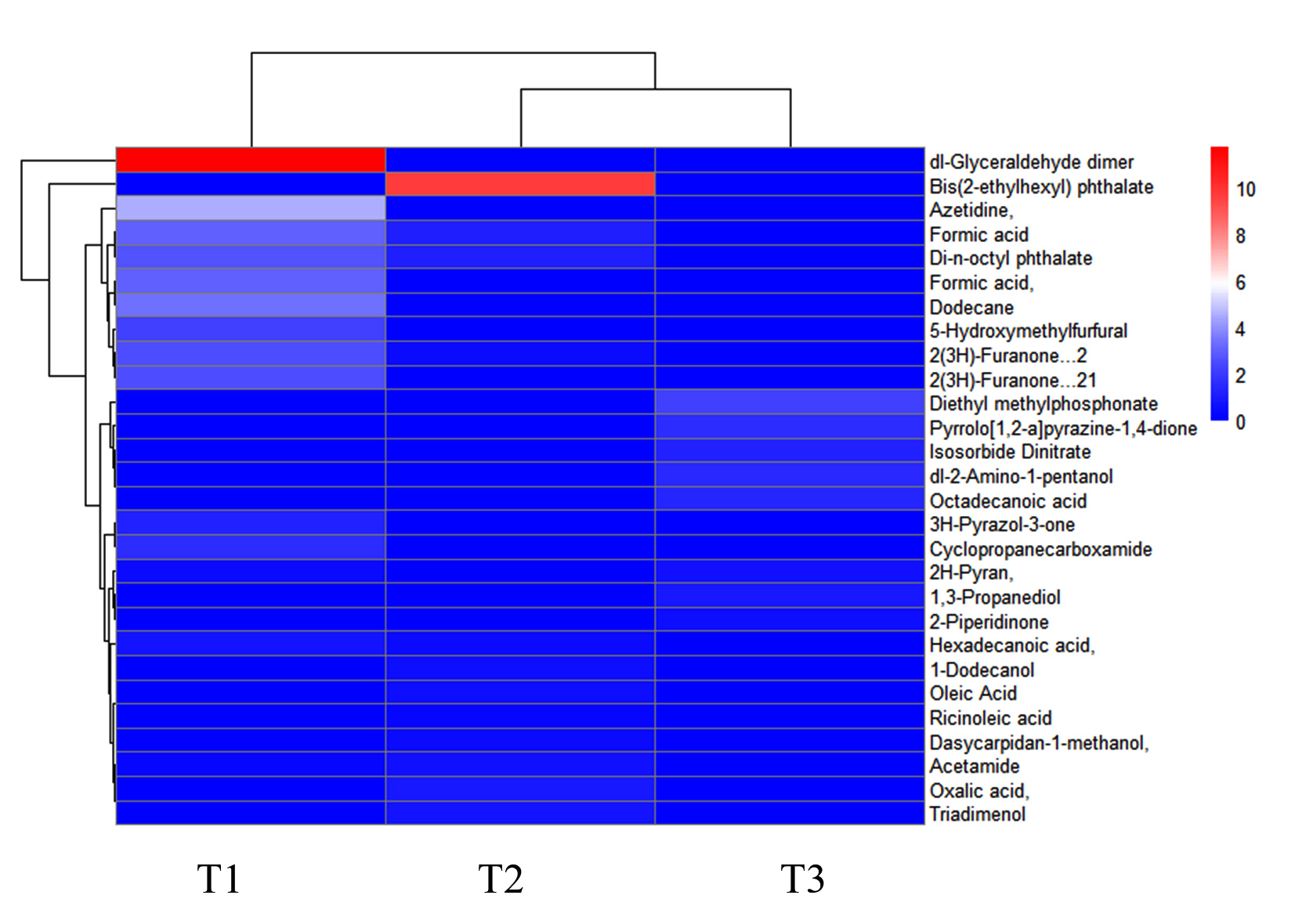
Figure 5. The heat map illustrates the secondary metabolites released by C. rosea TNAU CR04 and their interaction with A. alternata. The scale on the right ranges from 0 to 10. Blue represents low values (near 0), indicating low or no abundance of the compound in the treatment. Red represents high values (up to 10), indicating high abundance of the compound. The gradation from blue (low) to red (high) visually encodes the relative intensity of the measured compounds. T1- C. rosea + A. alternata, T2- A. alternata and T3- C. rosea
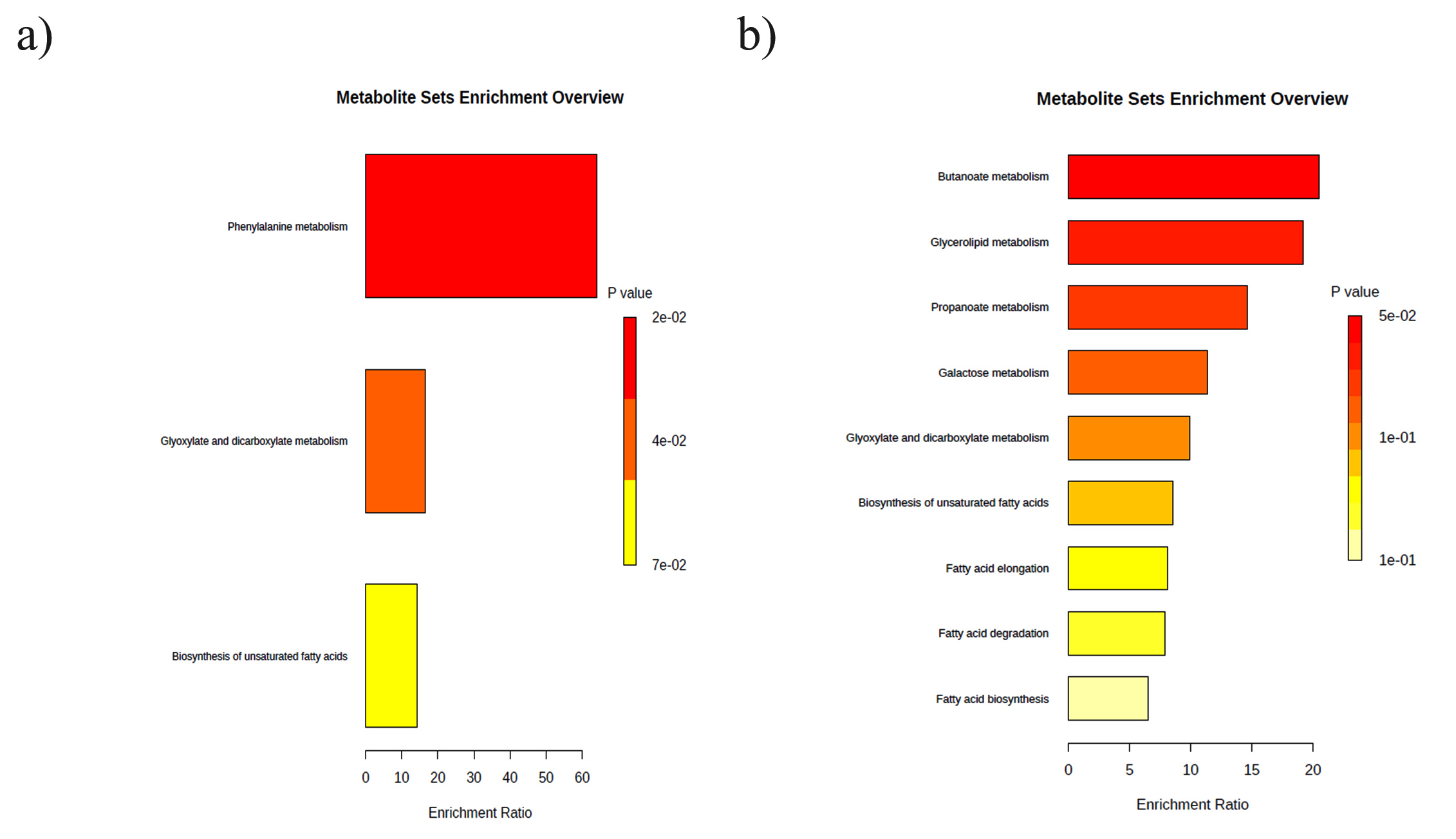
Figure 6. KEGG enrichment classification of non-volatile secondary metabolites for various treatments (a) KEGG enrichment classification of metabolites of C. rosea TNAU CR04 alone (b) KEGG enrichment classification of metabolites produced during the interaction between C. rosea TNAU CR04 and A. alternata
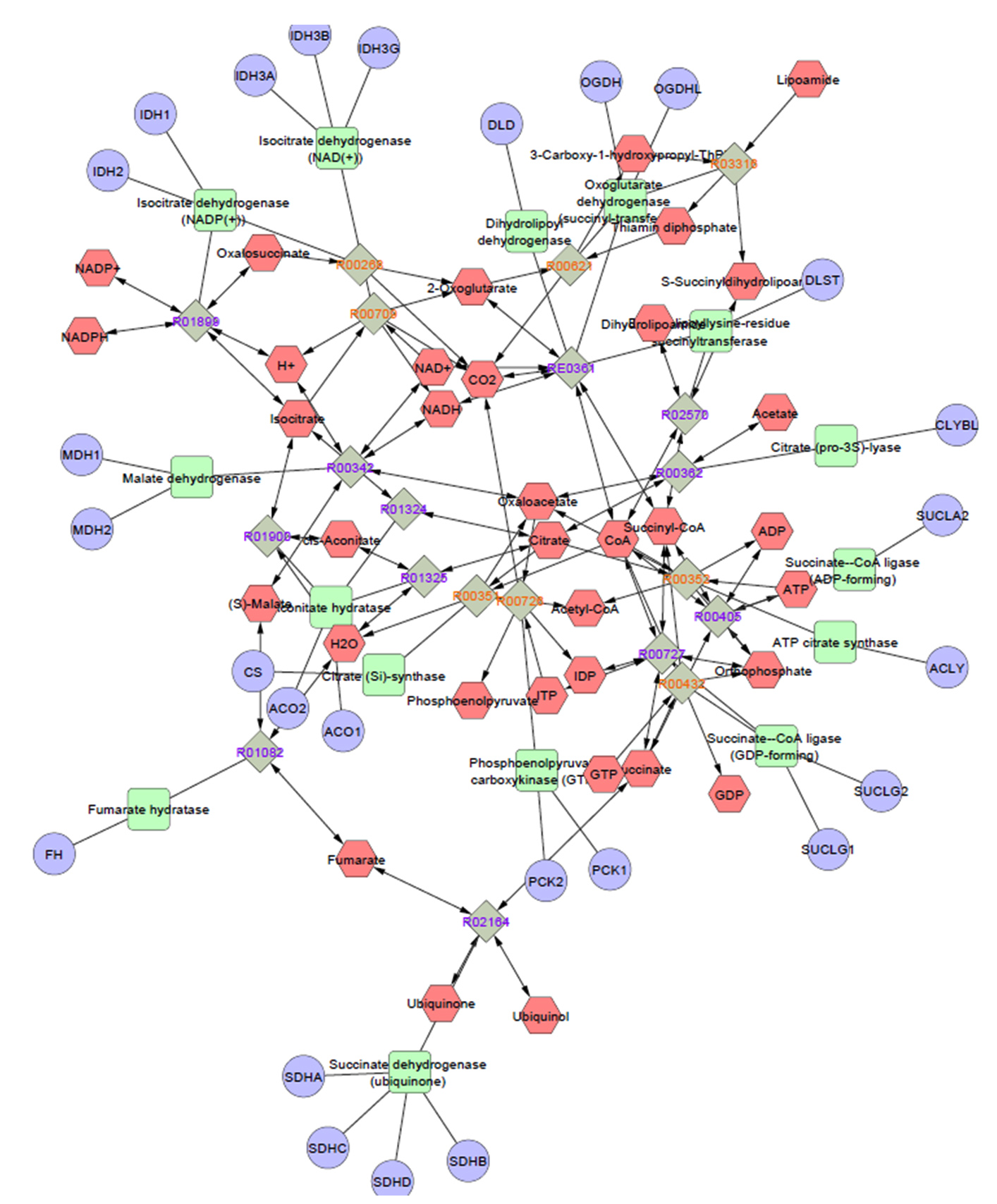
Figure 7. Pathway enrichment analysis. Various pathways involved in the interaction between the secondary metabolites of C. rosea TNAU CR04 and A. alternata was identified using Metscape in Cytoscape 3.3. Enzymes that participate in the biosynthesis of the TCA cycle and terpenes are indicated in green
Tomato plants are frequently afflicted by early blight caused by Alternaria spp., which is a widespread leaf disease, that greatly threatens the health of plants. This disease reduces the size and number of fruits and causes economic losses in yield of up to 79%.15,16 Fungicides serve as common chemical interventions used to effectively combat early blight disease. However, the utilization of chemicals to combat fungal infections in plants may not always be favorable due to associated risks. As an alternative approach, biocontrol agents comprising various microorganisms are preferred. These microorganisms are sustainable and have positive environmental effects, making them suitable for disease control in plants. Clonostachys rosea, known as a mycoparasite, that can antagonize various plant pathogens, such as Alternaria spp., Bipolaris sorokiniana, Botrytis cinerea, Fusarium culmorum, Rhizoctonia solani, Sclerotinia sclerotiorum, Pythium spp., and Verticillium dahliae as reported in studies by Yu and Sutton.,17 Jensen et al.,5 Huang,18 Knudsen et al.,19 McQuilken et al.,20 Rodriguez et al.11 Chatterton and Punja.10 and Keinath et al.21 Although many Clonostachys strains have been identified, only a small subset of fungi has been used in metabolomic profiling studies. Additionally, in this study we systematically investigated the inhibitory effect of C. rosea and its metabolites on the growth of Alternaria spp., via a combination of in vitro assays conducted in a laboratory environment.
The isolation and characterization of Alternaria spp. from diseased tomato leaves highlighted the prevalence and importance of this pathogen in agricultural settings. The observed colony morphology and growth patterns are consistent with previous descriptions of Alternaria species. The inhibitory effects observed under in vitro dual culture conditions across different C. rosea isolates, with isolate TNAU CR04 exhibiting the highest efficacy, revealed the promising biocontrol capabilities of this fungus. These results matched those of previous studies demonstrating the antagonistic activity of C. rosea toward plant pathogens, including seed-borne Alternaria spp. and Sclerotinia sclerotiorum.5,11 SEM images demonstrated the mycoparasitic behavior of C. rosea toward A. alternata, with C. rosea forming dense coils around A. alternata hyphae, leading to their collapse. These findings highlighted the ability of C. rosea to effectively control organism towards plant pathogens.22 This observation was also consistent with SEM observations of the association between C. rosea and B. cinerea, which revealed the penetration of conidia and germ tubes directly into B. cinerea. This penetration occurred without the formation of an appressorium, ultimately leading to cytoplasmic disintegration.8 We also assessed the inhibitory effects of secondary metabolites released by C. rosea isolates against pathogens. The isolate TNAU CR04 demonstrated the greatest inhibition rate of 77.22% at a 50% concentration of culture filtrate. This finding indicated that C. rosea acts as an antagonistic fungus by secreting inhibitory compounds.23,24 Han et al.25 and Saraiva et al.26 reported C. rosea generates diverse secondary metabolic compounds with strong antagonistic activity against plant pathogens. Clonostachys fungi produce copious amounts of various metabolites, including nitrogenous compounds, polyketide derivatives and terpenes. These metabolites exhibit a wide array of biological mechanisms including germicidal, insecticidal, nematicidal and cytocidal properties.27-29 The identification and characterization of non-volatile organic metabolites in C. rosea and their interactions with A. alternata offer valuable insights into the biochemical dynamics of fungal interactions. GC-MS analysis revealed a spectrum of compounds, including formic acid, 2(3H)-furanone, Di-n-octyl phthalate and 5-hexadecenoic acid in C. rosea. During its interaction with A. alternata, additional compounds such as
dl-glyceraldehyde dimer, 5-hydroxymethylfurfural, acetidine and cyclopropanecarboxamide were detected, indicating a shift in chemical composition.
A heat map showed significant metabolic shifts in C. rosea when interacting with A. alternata. KEGG analysis revealed that metabolites were enriched during the interaction between
C. rosea and A. alternata that participate in fatty acid-mediated and lipid-mediated signaling contribute to defense responses. These findings align to those of previous studies highlighting the metabolic shifts that occur in biocontrol fungi during pathogen antagonism.30-32
Pathway analysis depicted that metabolites from C. rosea play a crucial roles in enhancing plant immunity by regulating hormone signaling, influencing the tricarboxylic acid (TCA) cycle and stimulating the biosynthesis of defense-related metabolites. Similar results were found for other biocontrol agents such as Trichoderma spp., where energy metabolism is upregulated during antagonistic interactions to sustain the production of antifungal compounds and maintain cellular homeostasis.33 Manganiello et al.32 reported that Trichoderma harzianum stimulates the TCA cycle and the hexose monophosphate (HMP) pathway to increase tomato growth by increasing the activities of succinate dehydrogenase and glucose-6-phosphate dehydrogenase. In C. rosea, terpenoid production during interaction with A. alternata may increase its ability to suppress the growth of this pathogen. Similar findings have been reported in other studies, where terpenoid production was linked to the biocontrol efficacy of fungal antagonists.34 These findings can help understand fungal interactions and may aid in development of innovative biocontrol strategies.25,35,36 We further determined the molecular mechanisms underlying the antagonistic efficacy of C. rosea and its culture filtrates, including the identification of key bioactive compounds and biosynthetic genes and elucidated of their modes of action and the interaction dynamics between C. rosea and Alternaria spp.
To summarize, this study highlighted the promising potential of C. rosea as an effective antagonistic agent against plant pathogens using Alternaria spp. as a model system. By adopting a comprehensive approach involving in vitro assays, metabolomic profiling and detailed characterization of culture filtrates, we obtained significant insights into the inhibitory mechanisms of C. rosea against Alternaria spp. Our findings highlighted the multifaceted mode of action exhibited by C. rosea, including the production of inhibitory metabolites. In the future, further identification and characterization of the bioactive compounds, optimization of production methods and field trials to validate their efficacy in real-world agricultural settings needs to be performed. Overall, this study provided deeper insights into the antagonistic potential of C. rosea and laid the groundwork for its application in integrated disease management strategies, thus our findings may help to enhance crop productivity and sustainability.
ACKNOWLEDGMENTS
The authors acknowledge the Department of Plant Pathology, Agricultural College & Research Institute, Coimbatore, Tamil Nadu Agricultural University, Tamil Nadu, India, for providing necessary facilities in carrying out the above research work.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
VS and SH conceptualized the study and performed validation. RK performed project administration, supervision and applied methodology. AA performed investigation and experiments. US and NS collected resources and performed data interpretation. US, NS and AA performed analysis. VS and SH reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
ETHICS STATEMENT
Not applicable.
- Tiwari JK, Behera TK, Rai N, Yerasu SR, Singh MK, Singh PM. Tomato breeding for processing in India: Current status and prospects. Veg Sci. 2022;49(02):123-132.
Crossref - Panno S, Davino S, Caruso AG, et al. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the mediterranean basin. Agronomy. 2021;11(11):2188.
Crossref - Bauske MJ, Mallik I, Yellareddygari SKR, Gudmestad NC. Spatial and temporal distribution of mutations conferring QoI and SDHI resistance in Alternaria solani across the United States. Plant Dis. 2018;102(2):349-358.
Crossref - Awan ZA, Shoaib A, Iftikhar MS, Jan BL, Ahmad P. Combining biocontrol agent with plant nutrients for integrated control of tomato early blight through the modulation of physio-chemical attributes and key antioxidants. Front Microbiol. 2022;13: 807699.
Crossref - Jensen B, Knudsen IMB, Madsen M, Jensen DF. Biopriming of infected carrot seed with an antagonist, Clonostachys rosea, selected for control of seedborne Alternaria spp. Phytopathology. 2004;94(6):551-560.
Crossref - Abo-Elyousr KAM, Abdel-Hafez SII, Abdel-Rahim IR. Isolation of Trichoderma and evaluation of their antagonistic potential against Alternaria porri. J Phytopathol. 2014;162(9):9:567-574.
Crossref - Schroers HJ, Samuels GJ, Seifert KA, Gams W. Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia. 1999;91(2): 365-385.
Crossref - Hasan R, Lv B, Uddin MJ, et al. Monitoring mycoparasitism of Clonostachys rosea against Botrytis cinerea using GFP. J Fungi. 2022 8(6):567.
Crossref - Jensen B, Lubeck PS, Jorgensen HJ. Clonostachys rosea reduces spot blotch in barley by inhibiting prepenetration growth and sporulation of Bipolaris sorokiniana without inducing resistance. Pest Manag Sci. 2016;72(12):2231-2239.
Crossref - Chatterton S, Punja ZK. Chitinase and b-1,3-glucanase enzyme production by the mycoparasite Clonostachys rosea f. catenulata against fungal plant pathogens. Can J Microbiol. 2009;55(4):356-367.
Crossref - Rodriguez MA, Cabrera G, Gozzo FC, Eberlin MN, Godeas A. Clonostachys rosea BAFC3874 as a Sclerotinia sclerotiorum antagonist: mechanisms involved and potential as a biocontrol agent. J Appl Microbiol. 2011;110(5):1177-1186.
Crossref - Karlsson M, Durling MB, Choi J, et al. Insights on the evolution of mycoparasitism from the genome of Clonostachys rosea. Genome Biol Evol. 2015;7(2):465-480.
Crossref - Gowrisri N, Rengasamy K, Thiruvengadam R, Karthikeyan M, Shanmugam V, Narayanan S. Exploration of antimicrobial and antifungal compounds in Clonostachys rosea using gas chromatography-mass spectrometry. Indian J Chem. Technol. 2023;30(6):6541.
Crossref - Rapilly F. Mycology Techniques in Plant Pathology. Annals of Epiphyta. Vol 19. Edition INRA, Paris, 1988: 25-39.
- Sherf F, MacNab AA. Vegetable diseases and their control: John Wiley & Sons, 2nd ed. 1986.
- Gwary DM, Nahunnaro H. Epiphytotics of early blight of tomatoes in Northeastern Nigeria. Crop Prot. 1998;17(8):619-624.
Crossref - Yu H, Sutton JC. Morphological development and interactions of Gliocladium roseum and Botrytis cinerea in raspberry. Can J Plant Pathol. 1997;19(3):237-246.
Crossref - Huang HC. Gliocladium catenulatum: hyperparasite of Sclerotinia sclerotiorum and Fusarium species. Can J. Bot. 1978;56(18):2243-2246.
Crossref - Knudsen IMB, Hockenhull J, Jensen DF. Biocontrol of seedling diseases of barley and wheat caused by Fusarium culmorum and Bipolaris sorokiniana: effects of selected fungal antagonists on growth and yield components. Plant Pathol. 1995;44(3):467-477.
Crossref - Mcquilken, Gemmell, Lahdenpera. Gliocladium catenulatum as a potential biological control agent of damping off in bedding plants. J Phytopathol. 2001;149(3-4)171-178.
Crossref - Keinath AP, Fravel DR, Papavizas GC. Potential of Gliocladium roseum for biocontrol of Verticillium dahliae. Phytopathology. 1991;81(6):644-648.
Crossref - Ruppavalli VM, Karthikeyan M, Johnson I, et al. First Report of Clonostachys rosea as a Mycoparasite on Sclerotinia sclerotiorum Causing Head Rot of Cabbage in India. Plants. 2023;12(1)199.
Crossref - Kohl J, Kolnaar R, Ravensberg WJ. Mode of action of microbial biological control agents against plant diseases: relevance beyond efficacy. Front Plant Sci. 2019;10:454982.
Crossref - Alizadeh M, Vasebi Y, Safaie N. Microbial antagonists against plant pathogens in Iran: A review. Open Agric. 2020;5(1):404-440.
Crossref - Han Z, Ghanizadeh H, Zhang H, et al. Clonostachys rosea promotes root growth in tomato by secreting auxin produced through the tryptamine pathway. J Fungi. 2022;8(11):1166.
Crossref - Saraiva RM, Borges AV, Borel FC, Maffia LA. Compounds produced by Clonostachys rosea deleterious to Botrytis cinerea. Braz J Agric. 2020;95(1):34-47.
Crossref - Han P, Zhang X, Xu D, Zhang B, Lai D, Zhou L. Metabolites from Clonostachys fungi and their biological activities. J Fungi. 2020;6(4):229.
Crossref - Gowrisri N, Kannan R, Raguchander T, Swarnakumari N, Saravanakumar D. Nematicidal action of Clonostachys rosea against Meloidogyne incognita: in-vitro and in silico analyses. J Taibah Univ Sci. 2024;18(1):2288723.
Crossref - Zhang L, Yang J, Niu Q, et al. Investigation on the infection mechanism of the fungus Clonostachys rosea against nematodes using the green fluorescent protein. Appl Microbiol Biotechnol. 2008;78(6):983-990.
Crossref - Dedyukhina EG, Kamzolova SV, Vainshtein MB. Arachidonic acid as an elicitor of the plant defense response to phytopathogens. Chem Biol Technol Agric. 2024;1(8):1-6.
Crossref - Liu T, Chen Y, Tian S, Li B. Crucial roles of effectors in interactions between horticultural crops and pathogens. Horticulturae. 2023;9(2):250.
Crossref - Manganiello G, Sacco A, Ercolano MR, et al. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front Microbiol. 2018;9:1966.
Crossref - Dubey MK, Broberg A, Sooriyaarachchi S, Ubhayasekera W, Jensen DF, Karlsson M. The glyoxylate cycle is involved in pleotropic phenotypes, antagonism and induction of plant defence responses in the fungal biocontrol agent Trichoderma atroviride. Fungal Genet Biol. 2013;58:33-41.
Crossref - Wawrzyn GT, Bloch SE, Schmidt-Dannert C. Discovery and characterization of terpenoid biosynthetic pathways of fungi. Methods Enzymol. 2012;515:83-105.
Crossref - Lysoe E, Dees MW, Brurberg MB. A three-way transcriptomic interaction study of a biocontrol agent (Clonostachys rosea), a fungal pathogen (Helminthosporium solani), and a potato host (Solanum tuberosum). Mol Plant Microbe Interact. 2017;30(8):646-655.
Crossref - Wang Q, Chen X, Chai X, et al. The involvement of jasmonic acid, ethylene, and salicylic acid in the signaling pathway of Clonostachys rosea-induced resistance to gray mold disease in tomato. Phytopathology. 2019;109(7):1102-1114.
Crossref
© The Author(s) 2024. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.