


ISSN: 0973-7510
E-ISSN: 2581-690X
Biological control of plant pathogens is assuming increasing importance in organic and low input sustainable agriculture. Forty one Actinobacterial strains isolated from the arid, semi arid and humid regions of Rajasthan, Karnataka, Andhra Pradesh in India were tested for their antagonism to four plant pathogenic fungi viz., Macrophomina phaseolina, Sclerotium rolfsii, Rhizoctonia solani and Fusarium oxysporum. The Streptomyces strains from humid regions were more effective in inhibiting mycelial growth of Macrophomina phaseolina (62.8%). Arid and semi arid region strains (40.8 and 40.6%) were almost similar in inhibiting mycelial growth of Sclerotium rolfsii compared to humid region strains. Semi arid region strains (24.2%) were more effective in inhibiting the mycelial growth of Rhizoctonia solani followed by arid and humid region strains. However only the arid region strains (44.5%) showed inhibition towards Fusarium oxysporum. The strains with best antogonistic ability (10 no.s) also exhibited other plant growth promoting traits like production of Indole acetic acid (1.9 – 5.7 mg ml-1) and Gibberellic acid (24.1-41.4 mg ml-1). All the strains except Streptomyces A30 could solubilize P from inorganic tri-calcium phosphate. The amount of P solubilization ranged from 3.2 – 6.8%. All the strains except Streptomyces A40 produced alkaline phosphatase which ranged from 60.0-160.9 µg p-nitrophenol/ml /h. Some strains produced ammonia whereas none of the strains produced HCN, siderophore or exhibited chitinase activity. In a glass house evaluation in soil microcosms, among the ten strains Streptomyces A6 was found best for promoting maize and Streptomyces A17 for chickpea growth.
Actinobacteria, Streptomyces, Biocontrol, PGPR, dryland crops.
Pathogenic micro-organisms affecting plant health are a major and chronic threat to food production worldwide. Use of pesticides to control the pathogens is an inevitable part of food production strategy but their unscientific usage in developing countries has often led to adverse effects on soil and human health. Several negative effects like development of pathogen resistance and adverse influence on non-target populations have been reported. Furthermore, the growing cost of pesticides and consumer demand for pesticide-free food has led to a search for substitutes for these products. There are also a number of fastidious diseases for which chemical solutions are few, ineffective or nonexistent. Biological control is thus considered as an alternative or a supplemental way of reducing the use of chemicals in agriculture. Some of the Actinobacteria naturally occurring in soils and on rhizosphere of various crops act as potential biocontrol agents. Actinobacteria possess several properties which help in plant growth promotion and as biocontrol agent in combating several plant diseases. They colonize plant root surfaces, exhibit antibiosis against plant root pathogens, synthesis of particular extracellular enzymes, hydrogen cyanide production and siderophore production (Tokala et al. 2002). They are able to tolerate unfavourable growth conditions by forming spores (Alexander 1977). Actinobacteria, which have the ability to thrive under dry conditions, could be a valuable biocontrol resource to the poor farmers of dry land areas. Since Actinobacteria prefer dry to semi-dry soils, the crops grown in arid and semi-arid regions are likely to benefit by their inoculation. In the present study, Actinobacteria were isolated from arid, semi arid and also humid regions (for comparison) of western and peninsular India and screened for antagonism against some important fungal plant pathogens; their plant growth promoting attributes were also tested in vitro and in vivo on maize and chickpea in soil microcosms.
Isolation of Actinobacteria
Soil samples were collected from the rhizosphere of Sorghum (Sorghum vulgare), Pearl millet (Pennisetum typhoideum), Pigeon pea (Cajanus cajan), Finger millet (Eleusine coracana) and Groundnut (Arachis hypogea) grown in arid region of Jaisalmer in Rajasthan; arid and semi arid regions of Belgaum, Hubli, Bijapur and Tumkur districts of Karnataka and Anantapur in Andhra Pradesh. Two soil samples from pristine forest in humid region of Karnataka (Kegdal, Vibuthi) were also taken. Four different media were used for isolation of Actinobacteria viz., actinomycetes isolation agar, starch casein agar, arginine glycerol salts medium and humic acid vitamin agar. Isolates were characterized by referring to the 9th edition of Bergey’s manual of determinative bacteriology (Holt et al. 1994) and identified by standard methods (International Streptomyces project)
In vitro antagonism to fungal pathogens
The Actinobacterial strains were screened for the antagonism (biocontrol potential) to soil borne plant pathogens viz., Macrophomina phaseolina, Sclerotium rolfsii, Rhizoctonia solani and Fusarium oxysporum. The fungal cultures were obtained from the Directorate of Oilseeds Research, Hyderabad and Central Research Institute of Dryland Agriculture (CRIDA), Hyderabad, India. The mycelia plug of the pathogen (grown on PDA for two days) was cut and placed at the centre of fresh PDA plates using a cork borer. In the same PDA plate at 3 cm distance from the pathogen a plug of Actinobacteria (grown on oat meal agar for five days) was placed. This test was performed separately for all the Actinobacterial strains. A control was maintained without inoculation of Actinobacterial strains but grown only with pathogen. In vitro screening for antagonism was made in three replica plates. The inoculated plates were incubated at 30 ± 2 ºC for 7 days. Colony growth inhibition (%) was calculated by using the formula: C – T/C *100, where C is the colony growth (in mm) of pathogen in control, and T is the colony growth of pathogen in dual culture. All strains were tested in triplicate.
Growth Promoting Characteristics
The strains were tested for plant growth promoting attributes like Indole acetic acid (IAA) production (Gordon and Weber 1951); Gibberellic acid (GA) production (Paleg 1965); P solubilisation on Pikovskaya agar (Pikovskaya, 1948) followed by spectrophotometric quantiûcation of P solubilized in broth after 10 days growth at 28±2°C in shake cultures at 125rpm; K solubilization on Aleksandrov agar (Hu et al. 2006) followed by flame photometric quantification of K solubilized in broth after 10 days growth at 28±2°C in shake cultures at 125rpm; Alkaline phosphatase production (Tabatabai and Bremner, 1969); Ammonia production (Cappuccino and Sherman, 2005); Siderophore production (growth in chrome azurol S-CAS medium (Schwyn and Neilands, 1987); and chitinase production (Hamid et al. 2013) were determined.
Plant Bioassay
Ten efficient biocontrol strains Streptomyces A1, A2, A6, A10, A11, A17, A18, A30, A40 and A45 were screened for their ability to promote plant growth in soil microcosms under glass house conditions at Indian Institute of Soil Science, Bhopal. The experiment had five replications and laid out in completely randomized design. 330 ml paper cups were taken and filled with 300g soil (Vertic Eustochrept). The Actinobacterial strains were grown separately on starch casein broth for 7 days with shaking at 125 rpm. Farm yard manure (FYM) used as a carrier material was air dried for 3-4 days and passed through 0.2 mm sieve and sterilized three times by steam sterilization (121°C for 20 min) on successive days and then two times dry heat sterilization (160°C for 3 h each time). Forty milliliter of the culture broth was added to 100 gm FYM in a plastic pouch, mixed by hands and sealed. One gram of the FYM based inoculants was added to 10 ml of 1% carboxy methyl cellulose (CMC). Maize Seeds (var. JM-216) were surface sterilized by dipping them in 95% ethanol solution for 5 min and in 3% sodium hypochlorite (NaClO) solution for 5 minutes and subsequently washed in sterile distilled water for five times. Then 24 seeds of maize were transferred to the CMC culture suspension and kept overnight. The seeds were removed aseptically and air dried in a laminar air flow work station. The inoculants coated seeds were sown @ 4 seeds per cup. After germination, the plants were thinned to maintain three plants in each cup. After 15 days urea was applied as solution to the cups at 40 µg N g-1 soil. The cups were watered regularly with tap water (boiled for 30 min and cooled) to maintain optimum moisture. The plant height, number of leaves and plant dry mass was recorded at 4 weeks and 6 weeks growth stage (30 and 45 days after sowing). The same methodology was used for chickpea (JG-16) seed treatment. Observations like plant height recorded both at 30 and 60 days after sowing (DAS), nodule number and dry weight, dry weight of plants was recorded at 60 DAS.
The soil samples obtained from different crop rhizosphere at different places of arid, semi arid and humid regions had a wide variation in pH from 6.0 to 9.0. The organic carbon content ranged from 0.12 to 3.95 per cent (Table 1). The Actinobacterial count of the soil samples on four different nutrient media employed for enumeration is given in Table 1. Higher populations were recorded on Humic acid vitamin agar (14.8X104cfu/g soil) while the lowest Actinobacterial population was recorded in Argnine glycerol salts medium (3.5 X 104cfu/g soil). Humic acid vitamin agar was the best medium for enumeration of Actinobacteria from all soils. These results agree with Hayakawa and Nonomura (1987) who showed that Humic acid vitamin (HV) agar was superior to the other media including colloidal chitin agar, glycerol-arginine agar and starch-casein-nitrate agar for the isolation and enumeration of soil Actinobacteria. Similarly, Coombs and Franco (2003) observed that the nutrient poor media such as HV agar, TWYE (Tap water yeast extract) and YECH (Yeast extract- casein hydrolysate) agar are most effective for isolation of endophytic Actinobacteria from wheat root. When the average counts of Actinobacteria obtained from different type of soils grown on four different media were compared, the highest Actinobacterial population was observed in rhizosphere of crops sampled in black soils (Vertisols) which may be due to soil type and higher pH. Varalakshmi et al. (2010) who observed that black soils harboured more population of actinomycetes in the rhizosphere of fox tail millet as compared to red soil.
Table (1):
Enumeration of Actinobacteria (x 104cfu/g soil ±SEm) in different growth media*
| Climatic region | Soil order | Place and state | Origin | pH | OC (%) | AIA | SCA | HVA | AGS |
|---|---|---|---|---|---|---|---|---|---|
| Arid | Alfisol | Tumkur (Karnataka) Ananthpur (Andhra Pradesh) | Finger millet Groundnut | 6.3 | 0.36 | 5.2±0.12 | 4.5±0.18 | 11.3±0.25 | 2.6±0.05 |
| Aridisol | Jaisalmer (Rajasthan) Sam (Rajasthan) | Pearl millet Desert | 9.0 | 0.12 | 6.7±0.16 | 6.3±0.23 | 5.8±0.23 | 5±0.01 | |
| Semi-arid | Vertisol | Hubli, Bijapur (Karnataka) | Sorghum, Pigeon pea, Pearl millet | 8.6 | 0.66 | 8.5±0.17 | 7.2±0.26 | 31.7±0.30 | 4.2±0.17 |
| Alfisol | Gadag (Karnataka) | Sorghum | 8.4 | 0.65 | 5.2±0.17 | 4.6±0.08 | 10.0±0.32 | 2.8±0.17 | |
| Humid | Lateritic | Uttarakannada (Karnataka) | Forest | 6.0 | 3.95 | 7.5±0.14 | 7.2±0.23 | 9.0±0.26 | 3.0±0.31 |
| Vertisol | Uttarakannada (Karnataka) Belgaum (Karnataka) | Forest Sorghum | 6.2 | 0.81 | 8.2±0.15 | 7.7±0.26 | 21.2±0.48 | 3.5±0.22 | |
| Average | 7.4 | 1.1 | 6.88±0.15 | 6.25±0.20 | 14.83±0.30 | 3.51±0.15 |
*AIA: Actinomycetes Isolation agar, SCA: Starch casein agar, HVA: Humic acid vitamin agar, AGS: Arginine glycerol salts medium.
OC: organic carbon
Forty one Actinobacterial isolates were obtained in total from all the soil. Based on morphological, biochemical and microscopic observations, these isolates were assigned to four genera (data not shown) – Streptomyces (26 no.), Nocardia (12), Micromonospora (1) and Saccharopolyspora (2). Highest number of isolates belonged to the genus Streptomyces (63%), followed by Nocardia (29%) while rest of the isolates belonged to Micromonospora (5%) and Saccharopolyspora (2%). Dominance of Streptomyces among the actinomycetes was also earlier reported by many research workers (Alexander, 1961; Gupta1991; Gesheva and Gesheva 2000).
In this study, 41 Actinobacterial strains were tested for their in vitro antagonistic activity against four plant pathogenic fungi viz., Macrophomina phaseolina, Sclerotium rolfsii, Rhizoctonia solani and Fusarium oxysporum. Some of the Streptomyces strains inhibited all the four pathogens whereas Nocardia, Saccharopolyspora and Micromonospora strains inhibited all the pathogens except Fusarium oxysporum (Table 2). The strains from humid regions were more effective in inhibiting mycelial growth of Macrophomina phaseolina (inhibited mycelial growth by 62.8%) (Fig.1) followed by arid and semi arid region strains. Arid and semi arid region strains (40.8 and 40.6%) are almost similar in inhibiting mycelia growth of Sclerotium rolfsii compared to humid region strains. Semi arid region strains (24.2%) are more effective in inhibiting the mycelia growth of Rhizoctonia solani followed by arid and humid region strains, whereas only arid region strains (44.5%) showed inhibition towards Fusarium oxysporum. This biocontrol potentiality can be attributed to the production of antimicrobial compounds including ammonia production. Khamna et al. (2009) observed biocontrol activity of 23 Streptomyces isolates against five phytopathogenic fungi viz., Alternaria brassicicola, Colletotrichum gloeosporioides, Fusarium oxysporum, Penicillium digitatum and Sclerotium rolfsii. Subhashini (2010) isolated 40 actinomycetes from tobacco soils of India and reported that 10 Streptomyces isolates showed high antagonistic activity against Pythium aphanidermatum under in vitro conditions.
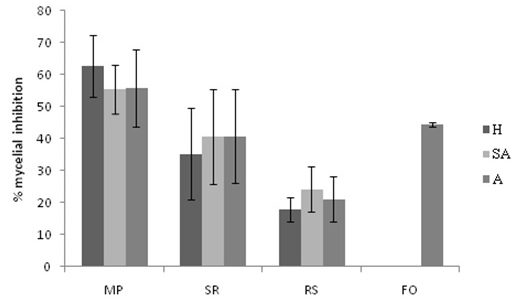
Fig 1. Biocontrol ability of Actinobacteria against fungal plant pathogens (dual culture). MP: Macrophomina phaseolina; SR: Sclerotium rolfsii; RS: Rhizoctonia solani; FO: Fusarium oxysporum; H: Humid; SA: Semi arid; A: Arid
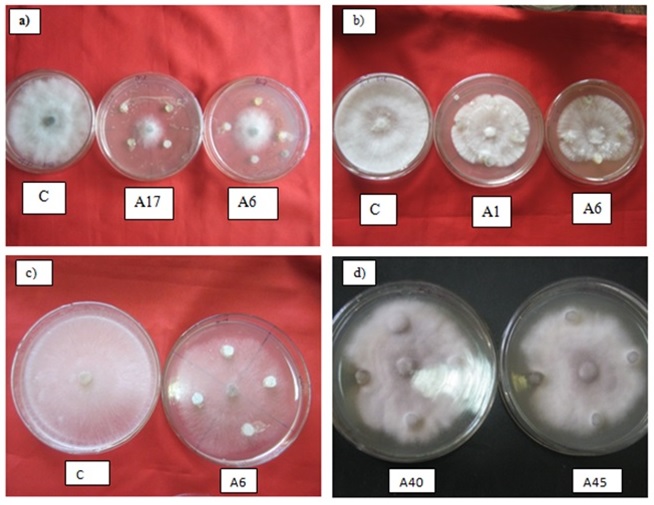
Fig. 2. Antagonism of Actinobacteria against fungal plant pathogens; a) Macrophomina phaseolina b) Sclerotium rolfsii c) Rhizoctonia solani d) Fusarium oxysporum
Table (2):
Biocontrol potential of different genera of Actinobacteria against fungal plant Pathogens
Genera |
Number of strains inhibited MP |
Number of strains inhibited SR |
Number of strains inhibited RS |
Number of strains inhibited FO |
|---|---|---|---|---|
Streptomyces (26 no’s) |
20 |
20 |
18 |
2 |
Nocardia (12 no’s) |
7 |
7 |
6 |
0 |
Saccharoployspora (2 no’s) |
2 |
2 |
1 |
0 |
Micromonospora (1 no) |
1 |
1 |
1 |
0 |
* MP: Macrophomina phaseolina, SR: Sclerotium rolfsii, RS: Rhizoctonia solani, FO: Fusarium oxysporum
Ten Actinobacterial strains showing best biocontrol ability were screened for plant growth promoting attributes and also tested for their influence on plant growth of maize (Table 4) and chickpea (Table 5) under glass house conditions. The Indole acetic acid (IAA) production of the strains under in vitro conditions ranged from 1.9- 5.7 mg ml-1, Gibberellic acid (GA) production ranged from 24.1-41.4 mg ml-1. Solans et al. (2009) found that three rhizoactinomycetes Streptomyces MM40, Actinoplanes ME3 and Micromonospora MM18 produced and released IAA, GA3, and zeatine in a defined media. All the strains except Streptomyces A30 could solubilize P from inorganic tri-calcium phosphate. The amount of P solubilzation ranged from 3.2-6.8%. Among the 10 strains 7 strains could solubilize K from muscovite mica which ranged from 3.7-6.3 mg ml-1. Some strains produced ammonia whereas none of the strains produced HCN, siderophore or exhibited chitinase activity. All the strains except Streptomyces A40 produced alkaline phosphatase which ranged from 60.0-160.9 µg p-nitrophenol -1ml-h.
Table (3):
Plant growth promoting attributes of Actinobacteria (Streptomyces spp.)
Isolates |
IAA (mg ml-1) |
GA (mg ml-1) |
% Pi released at 10 days |
Alkaline Phosphatase (µg p-nitrophenol/ml /h) |
K solubilization (mg ml-1) at 10 days |
Ammonia production |
|---|---|---|---|---|---|---|
A1 |
5.1 |
34.8 |
6.0 |
83.6 |
6.1 |
+ |
A2 |
1.9 |
24.3 |
4.8 |
60.0 |
3.7 |
+ |
A6 |
5.7 |
35.3 |
6.5 |
141.6 |
6.3 |
+ |
A10 |
5.6 |
33.0 |
6.02 |
106.9 |
5.9 |
+ |
A11 |
3.9 |
24.1 |
4.9 |
78.6 |
4.0 |
– |
A17 |
5.42 |
41.4 |
6.8 |
160.9 |
6.3 |
+ |
A18 |
– |
– |
3.2 |
79.2 |
3.9 |
– |
A30 |
3.3 |
29.5 |
– |
74.4 |
– |
– |
A40 |
2.5 |
– |
5.6 |
– |
– |
– |
A45 |
3.4 |
28.6 |
4.3 |
62.3 |
– |
+ |
*All strains negative for HCN production, siderophore production and chitinase activity.
Table (4):
Effect of Actinobacteria (Streptomyces spp.)on Maize (Var. JM-216) at different growth stages
Isolates |
Plant height (cm plant-1) 30 DAS |
Plant height (cm plant-1) 45 DAS |
No. of leaves (plant-1) 30 DAS |
No. of leaves (plant-1) 45 DAS |
Total DM (mg plant-1) 45 DAS |
|---|---|---|---|---|---|
A1 |
14.4 |
19.2 |
6.3 |
8.4 |
560 |
A2 |
12.6 |
16.8 |
6.0 |
8.0 |
554 |
A6 |
15.0 |
20.0 |
6.7 |
8.9 |
1044 |
A10 |
13.1 |
17.5 |
6.3 |
8.4 |
485 |
A11 |
14.1 |
18.9 |
6.0 |
8.0 |
467 |
A17 |
12.8 |
17.0 |
6.0 |
8.0 |
493 |
A18 |
12.9 |
17.2 |
5.0 |
6.7 |
364 |
A30 |
13.5 |
18.0 |
5.0 |
6.7 |
551 |
A40 |
11.7 |
15.6 |
5.7 |
7.6 |
456 |
A45 |
15.7 |
21.0 |
5.0 |
6.7 |
417 |
Control |
10.5 |
14.0 |
4.0 |
5.3 |
380 |
S.Em ± |
0.15 |
0.20 |
NS |
NS |
2.5 |
CD @ 1% |
0.48 |
0.63 |
8.0 |
Franco-Correa et al. (2010) isolated 30 actinomycetes from the rhizosphere of Trifolium repens L. and found that 21 isolates exhibited phosphate solubilizing ability on Pikovskaya’s medium, all the isolates produced acid phosphatase and only thirteen isolates produced alkaline phosphatase.
Results of glass house screening showed that Actinobacterial strains significantly differed in influencing plant growth attributes like plant height, number of leaves and total plant dry weight. Among all the strains the highest plant height (20.0 cm), number of leaves (8.9) and plant dry weight (1044 mg plant -1) was obtained in plants inoculated with Actinobacterial strain A6 (Streptomyces isolated from arid soils) at 45 DAS. Gopalakrishnan et al., (2013) evaluated five strains of Streptomyces on Sorghum under green house conditions and found that all the five strains had significantly enhanced plant growth parameters over un-inoculated control. In Chickpea, strain A17 (Streptomyces) showed highest plant height (30.5 cm) number of nodules (29.0) nodule dry weight (26.7 mg plant-1) and plant dry weight (936 mg plant -1). The increase in nodule numbers could be due to improvement of Rhizobium symbiosis caused by the plant growth promoting action of the Actinobacteria. Solans et al. (2009) co-inoculated actinomycetes with Siniorhizobium meliloti and found that nodulation and growth of Alfalfa (Medicago sativa) plants were significantly increased when compared to plants inoculated only with Siniorhizobium meliloti. The findings of this study is of much significance to crop production in utilizing efficient Actinobacterial strains like Streptomyces A6 for maize and Streptomyces A17 in chickpea cultivation to obtain higher crop yields. The identified strains have high potential for use particularly in dry land agriculture as bioinoculants for suppressing plant pathogens, solubilzation of P and K with the added advantage to crops in terms of improved crop growth and yields due to plant growth promoting hormones elaborated by the bacteria in crop rhizosphere.
Table (5):
Effect of Actinobacteria (Streptomyces spp.) on Chickpea var (JG-16) at different growth stages
Isolates |
Plant height (cm plant-1)30 DAS |
Plant height (cm plant-1)60 DAS |
Nodule number (No.plant-1)60 DAS |
Nodule dry weight (mg plant-1)60 DAS |
Total Dw (g plant-1) 60 DAS |
|---|---|---|---|---|---|
A1 |
21.8 |
28.9 |
23 |
13.3 |
797 |
A2 |
20.2 |
28.2 |
29 |
23.3 |
861 |
A6 |
21.7 |
29.0 |
22 |
16.7 |
808 |
A10 |
17.2 |
30.8 |
29 |
23.7 |
896 |
A11 |
24.2 |
27.5 |
28 |
11.7 |
863 |
A17 |
20.3 |
30.5 |
29 |
26.7 |
936 |
A18 |
21.5 |
29.6 |
23 |
16.7 |
848 |
A30 |
19.2 |
28.7 |
21 |
16.7 |
793 |
A40 |
21.2 |
29.8 |
21 |
20.0 |
860 |
A45 |
21.8 |
30.2 |
21 |
11.7 |
736 |
Control |
11.3 |
27.0 |
19 |
10.3 |
661 |
S.Em ± |
0.10 |
0.36 |
1.0 |
0.2 |
3.2 |
CD @ 1% |
0.31 |
1.08 |
3.0 |
0.6 |
9.6 |
- Alexander, M (ed): Introduction to Soil Microbiology, Wiley International edn. Tropan Company Ltd., Japan, 1961; pp 472.
- Alexander, M (ed): Introduction to soil microbiology, 2nd edn. Wiley, New York, 1977; pp 467.
- Cappuccino, J.G., Sherman, N. In: Microbiology: a laboratory manual, 6thedn. Pearson education, Singapore, 2005.
- Coombs, J., T., Franco, C.,M. Isolation and identification of actinobacteria from surface-sterilized wheat roots. Appl. Environ. Microbiol., 2003; 69(9): 5603-5608.
- Gesheva, V., Geshava, R. Physiological and antagonistic potential of actinomycetes from Ioquat rhizosphere. Microbiol. Res., 2000; 155:133-135.
- Gopalakrishnan, S., Srinivas, V., Vidya, M.,S., Rathore, A. Plant growth-promoting activities of Streptomyces spp. in sorghum and rice. Springer Plus, 2013; 2(1): 574.
- Gordon, S.,A., Weber, R.,P. Colorimetric estimation of indoleacetic acid. Plant Physiol., 1951; 26: 192-195.
- Gupta, A. Characterization of soil actinomycetes of Agra region. Actabotanica Indica., 1991.
- Hamid, R., Mahboob, A., Ahmad, M.,M., Abdin, M.,Z., Javed, S. Purification and characterization of thermostable chitinase from a novel Stenotrophomonas maltophilia strain. Malays. J. Microbiol., 2013; 9(1): 7-12.
- Hayakawa, M., Nonomura, H. Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes. J. Ferment. Technol., 1987; 65: 501-509.
- Hu, X.,F., Chen, J., Guo, J.,F. Two phosphate and potassium solubilizing bacteria isolated from Tiannu mountain, Zhejiang, China. World J. Micro. Biotech., 2006; 22: 983-990.
- Khamna, S., Yokota, A., Peberdy, J. F., Lumyong, S. Antifungal activity of Streptomyces spp. isolated from rhizosphere of Thai medicinal plants. Intl. J. Integrative Biol., 2009; 6(3):143-147.
- Paleg, L., G. Physiological effects of gibberellins. Ann. Rev. Plant Physiol., 1965; 16: 291-322.
- Pikovskaya, R., I. Mobilization of phosphates in soil in connection with the vital activities of some microbial species. Mikrobiol., 1948; 17: 362-370.
- Schwyn, B., Neilands, J., B. Universal assay for the detection and determination of siderophores. Annal. Biochem., 1987; 160:47-56.
- Solans, M., Vobis, G., Wall, L.G. Saprophytic actinomycetes promote nodulation in Medicagosativa- Sinorhizobium meliloti symbiosis in the presence of high N. J. Plant Growth Reguln., 2009; 28(2): 106-114.
- Subhashini, D., V. Biological control of damping off (Pythium aphanidermatum) using Streptomyces lavendulae isolate 21 on tobacco seedbeds. J. Biological control., 2010; 24(4): 338-342.
- Tabatabai, M., A., Bremner, J., M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Boil. Biochem., 1969; 1(4): 301-307.
- Tokala, R.,K., Strap, J.,L., Jung, C.M., Crawford, D.L., Salove, M.,H., Deobald, L.,A., Bailey, J.,F., Morra, M.,J. Novel plant-microbe rhizosphere interaction involving Streptomyces lydicusWYEC108 and the pea plant (Pisum sativum). Appl. Environ. Microbiol., 2002; 68: 2161-2171.
- Varalakshmi, T., Sekhar, K., M., Rafi, M., M., Charyulu, P.,B.,B.,N. Population studies of actinomycetes isolated from the rhizosphere of Foxtail millet. Curr. Biotica., 2010; 3(1):305-317.
© The Author(s) 2017. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.